
Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
3. Results
Systematic Paleontology
- Order Coleoptera Linnaeus, 1758
- Superfamily Cucujoidea Latreille, 1802
- Family Erotylidae Latreille, 1802
- Subfamily Xenoscelinae Ganglbauer, 1899
- Genus Xenophagus Lyubarsky & Perkovsky, 2017
- Type species: Xenophagus popovi Lyubarsky & Perkovsky, 2017
- Species composition. Type species, Xenophagus simutniki sp.n.
- Xenophagus simutniki Lyubarsky & Perkovsky, sp.n.
- http://zoobank.org/urn:lsid:zoobank.org:act:97E43669-56D8-4103-BC4E-D273399E242A (accessed on 20 February 2023)
- Etymology. Named in honor of our friend, hymenopterist Dr. Sergej A. Simutnik.
- MATERIAL. Holotype: SIZK UA-28134, Rovno amber, late Eocene.
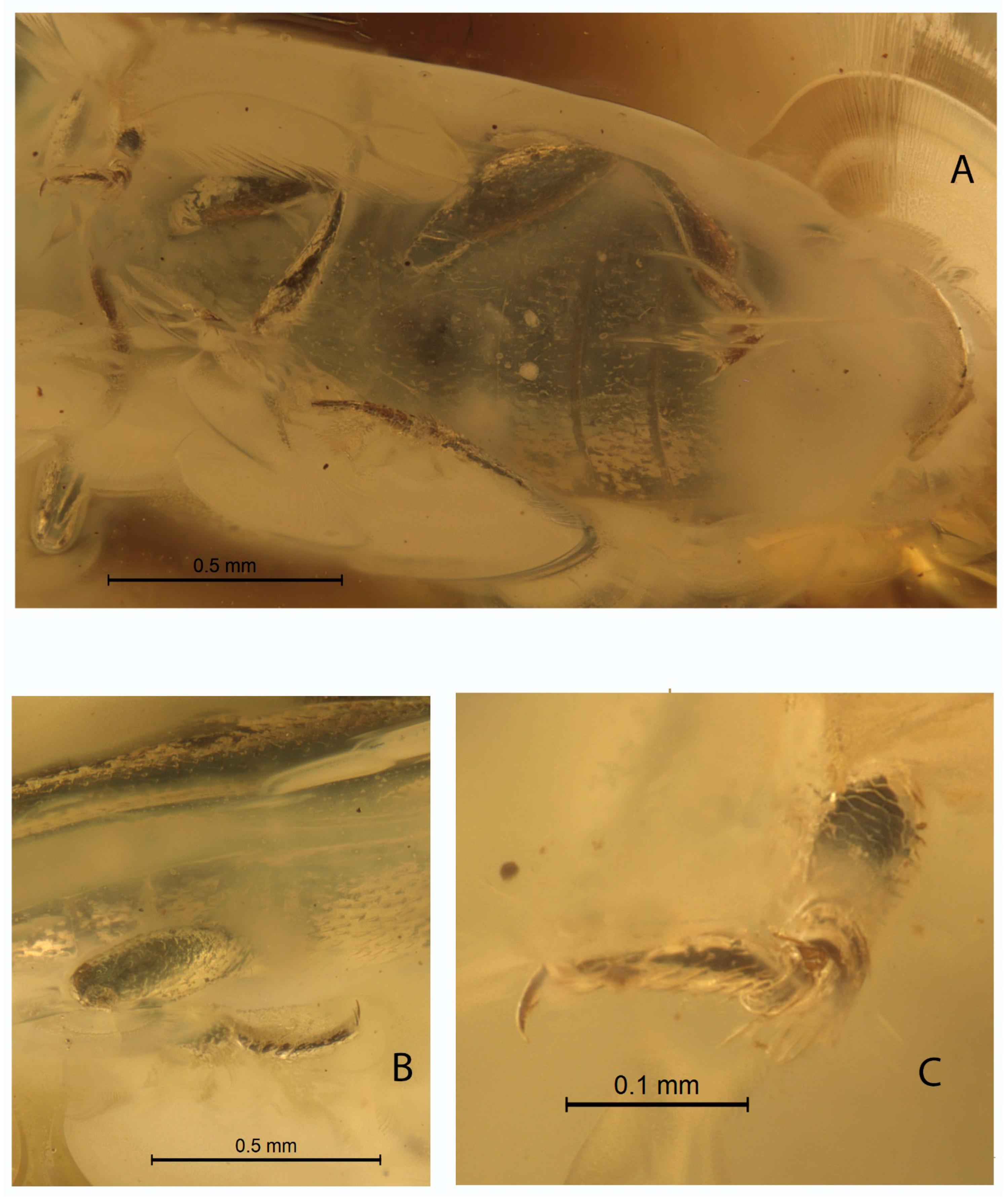
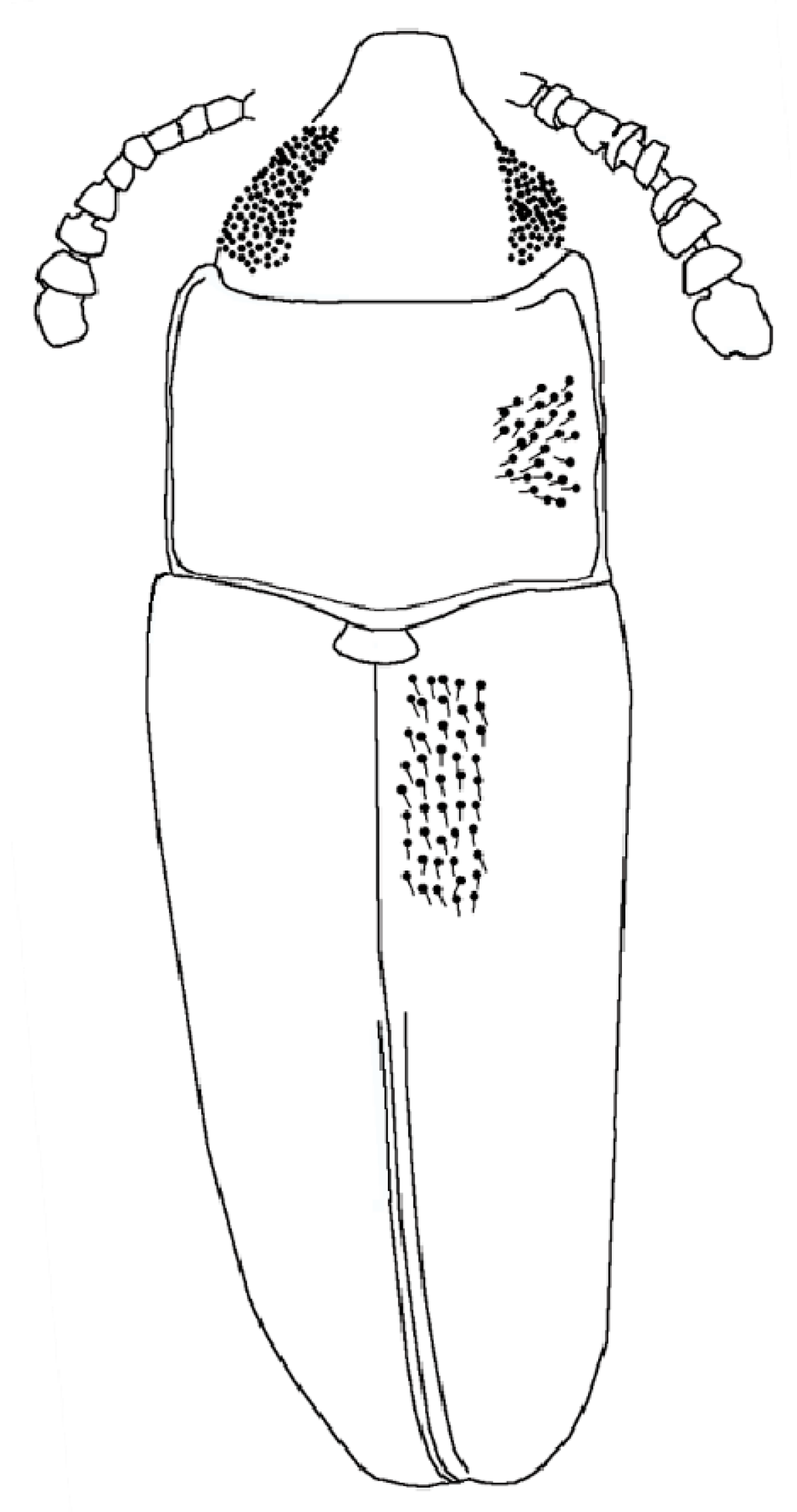
- DESCRIPTION. Body parallel-sided, 2.5 times longer than wide, dorsal vestiture short and decumbent (Figure 1A), dorsum weakly convex. Body length 2.7 mm, width 0.9 mm.
- 1.
- —
- Submetacoxal lines not present……………………………………………………………….……4
- 2.
- Elytral punctures irregular. .…………………………………………………………………………3
- —
- Elytral punctures arranged in rows. Baltic amber ………… Warnis tvanksticus Lyubarsky, Perkovsky & Alekseev, 2016
- 3.
- Anterior margin of pronotum notched (Figure 1A). Rovno amber..……Xenophagus simutniki Lyubarsky & Perkovsky, sp. n.
- —
- Anterior margin of pronotum without notch. Baltic amber …………Xenophagus popovi Lyubarsky & Perkovsky, 2017
- 4.
- Pronotum widest across apical one third. ..……………………………………………………5
- —
- Pronotum almost completely parallel-sided. Rovno amber..………… Xenohimatium rovnense Lyubarsky & Perkovsky, 2012
- 5.
- Antenna long, extending beyond posterior edge of pronotum when directed posteriad. Rovno amber ..………Zavaljus lyubarskyi Alekseev & Bukeis, 2022
- —
- Antenna short, extending towards midlength of pronotum when directed posteriad. Bitterfeld amber ………Microzavaljus saxonicus Lyubarsky & Perkovsky, 2018
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Węgrzynowicz, P. Morphology, phylogeny and classification of the family Erotylidae based on adult characters (Coleoptera: Cucujoidea). Genus 2002, 13, 435–504. [Google Scholar]
- Leschen, R.A.B. Erotylidae (Insecta: Coleoptera: Cucujoidea): Phylogeny and review. Fauna New Zealand 2003, 47, 1–108. [Google Scholar]
- Leschen, R.A.B.; Węgrzynowicz, P. Generic catalogue and taxonomic status of Languriidae (Coleoptera: Cucujoidea). Ann. Zool. 1998, 48, 221–243. [Google Scholar]
- Robertson, J.A.; McHugh, J.V.; Whiting, M.F. A molecular phylogenetic analysis of the pleasing fungus beetles (Coleoptera: Erotylidae): evolution of colour patterns, gregariousness, and mycophagy. Syst. Entomol. 2004, 29, 173–187. [Google Scholar] [CrossRef]
- Leschen, R.A.B.; Buckley, T.R. Multistate characters and diet shifts: Evolution of Erotylidae (Coleoptera). Syst. Biol. 2007, 56, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Skelley, P.E. Termitophilous pleasing fungus beetles: Chasmatodera Arrow and Rhamphidera Skelley, gen. nov. (Coleoptera: Erotylidae: Tritominae). Ann. Zool. 1999, 49, 255–263. [Google Scholar]
- Leschen, R.A.B.; Ashe, J.S. New species, phylogenetic placement, and mammal associations of Loberopsyllus (Languriidae: Xenoscelinae). In Entomological Contributions in Memory of Byron Alexander; Byers, G.W., Hagen, R.H., Brooks, R.W., Eds.; University of Kansas Natural History Museum Special Publication: Lawrence, KS, USA, 1999; Volume 24, pp. 171–177. [Google Scholar]
- Leschen, R.A.B. The Empocryptus-group (Languriidae: Toraminae) and a new genus associated with a lepidopteran cocoon. Coleopt. Bull. 1997, 51, 303–318. [Google Scholar]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Microzavaljus, a second extinct genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Bitterfeld Amber. Russ. Entomol. J. 2018, 27, 271–276. [Google Scholar] [CrossRef]
- Alekseev, V.I. A new species of Triplax Herbst, 1793 (Coleoptera: Erotylidae) from Bitterfeld amber. Balt. J. Coleopterol. 2014, 14, 171–177. [Google Scholar]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Serramorphus, a new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia) from Late Eocene Bitterfeldian amber. Russ. Entomol. J. 2017, 26, 37–40. [Google Scholar] [CrossRef]
- Kupryjanowicz, J.; Lyubarsky, G.Yu.; Perkovsky, E.E. A new genus of the subfamily Languriinae (Coleoptera: Erotylidae) from the Late Eocene Baltic amber. Far East. Entomol. 2022, 447, 1–9. [Google Scholar] [CrossRef]
- Alekseev, V.; Bukejs, A. First fossil representatives of Pharaxonothinae Crowson (Coleoptera: Erotylidae): indirect evidence for cycads existence in Baltic amber forest. Zootaxa 2017, 4337, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Lyubarsky, G.Yu.; Perkovsky, E.E.; Alekseev, V.I. The first record of the subfamily Xenoscelinae (Coleoptera, Erotylidae) from the Baltic amber. Paleontol. J. 2016, 50, 963–969. [Google Scholar] [CrossRef]
- Lyubarsky, G.Yu.; Perkovsky, E.E. Xenophagus, a new genus of pleasing fungus beetles (Coleoptera: Erotylidae) from Baltic amber. Russ. Entomol. J. 2017, 26, 147–150. [Google Scholar] [CrossRef]
- Lyubarsky, G.Yu.; Perkovsky, E.E. A new genus of Erotylidae from Eocene amber (Coleoptera: Clavicornia). Russ. Entomol. J. 2012, 21, 35–38. [Google Scholar] [CrossRef]
- Alekseev, V.I.; Bukejs, A. A new extinct species of Zavaljus Reitter (Coleoptera: Erotylidae) from Rovno amber: boreal distribution range since the Eocene. Hist. Biol. 2022, 35, 315–319. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Zosimovich, V.Y.; Vlaskin, A.P. Chapter 7: Rovno amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Legalov, A.A.; Nazarenko, V.Yu.; Perkovsky, E.E. A new species of the genus Dorytomus Germar, 1817 (Coleoptera, Curculionidae) from Rovno amber. Zootaxa 2021, 5006, 95–100. [Google Scholar] [CrossRef]
- Martynov, A.V.; Vasilenko, D.V.; Perkovsky, E.E. First Odonata from Upper Eocene Rovno amber (Ukraine). Hist. Biol. 2022, 34, 2182–2187. [Google Scholar] [CrossRef]
- Perkovsky, E.E. Rovno amber caddisflies (Insecta, Trichoptera) from different localities, with information about three new sites. Vestn. Zool. 2017, 51, 15–22. [Google Scholar] [CrossRef]
- Jałoszyński, P.; Perkovsky, E.E. Diversity of Scydmaeninae (Coleoptera: Staphylinidae) in Upper Eocene Rovno amber. Zootaxa 2016, 4157, 1–85. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Ignatov, M.S.; Remizowa, M.V.; Nuraliev, M.S.; Blagoderov, V.; Garbout, A.; Perkovsky, E.E. Staminate flower of Prunus s. l. (Rosaceae) from Eocene Rovno amber (Ukraine). J. Plant. Res. 2018, 131, 925–943. [Google Scholar] [CrossRef]
- Perkovsky, E.E. Only a half of species of Hymenoptera in Rovno amber is common with Baltic amber. Vestn. Zool. 2018, 52, 353–360. [Google Scholar] [CrossRef]
- Mitov, P.G.; Perkovsky, E.E.; Dunlop, J.A. Harvestmen (Arachnida: Opiliones) in Eocene Rovno amber (Ukraine). Zootaxa 2021, 4984, 43–72. [Google Scholar] [CrossRef]
- Bukejs, A.; Háva, J.; Alekseev, V.I. A new fossil species of Attagenus Latreille (Coleoptera: Dermestidae) in Rovno and Baltic ambers, with a brief review of known fossil beetles from the Rovno amber Lagerstätte. Foss. Rec. 2020, 23, 95–104. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Kairišs, K. The first record of Cupedidae (Coleoptera: Archostemata) from Eocene Rovno amber: Cupes groehni Kirejtshuk, 2005 examined using X-ray microtomography. Balt. J. Coleopterol. 2021, 21, 111–116. [Google Scholar]
- Sokolov, A.V.; Perkovsky, E.E. The first Eocene species of Bacanius (Coleoptera: Histeridae: Dendrophilinae) from Rovno amber. Russ. Entomol. J. 2020, 29, 157–160. [Google Scholar] [CrossRef]
- Kazantsev, S.V.; Perkovsky, E.E. The first Cacomorphocerus species (Coleoptera, Cantharidae) from Rovno amber: A second species of the genus with 11-segmented antennae. Zootaxa 2020, 4751, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, S.V.; Perkovsky, E.E. Imprint of a Helcophorus Fairmaire, 1881: The first net-winged beetle (Coleoptera: Lycidae) from Rovno amber. Zootaxa 2022, 5128, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Kupryjanowicz, J.; Lyubarsky, G.Y.; Perkovsky, E.E. Heterhelus buzina sp.n. (Coleoptera: Kateretidae) from Rovno amber: The first proxy for Sambucus in the Eocene of Eastern Europe. Invertebr. Zool. 2021, 18, 16–24. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Perkovsky, E.E. Protomauroania mikhailovi—A new species of malachite beetles (Coleoptera, Dasytidae) in Rovno amber. Zootaxa 2021, 5006, 189–194. [Google Scholar] [CrossRef]
- Jałoszyński, P.; Perkovsky, E.E. A new bizarre species of Euconnus (Cladoconnus) in Upper Eocene Rovno amber (Coleoptera: Staphylinidae: Scydmaeninae). Zootaxa 2021, 5004, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Alekseev, V.I. Cathartosilvanus perkovskyi sp. nov.—first record of Silvanidae (Coleoptera: Cucujoidea) from Eocene Rovno amber. Balt. J. Coleopterol. 2021, 21, 19–24. [Google Scholar]
- Alekseev, V.I.; Bukejs, A. A new fossil genus of minute-clubbed beetles (Coleoptera: Cucujoidea: Monotomidae) from Paleogene amber of Europe. Hist. Biol. 2022, 35, 430–435. [Google Scholar] [CrossRef]
- Alekseev, V.I.; Bukejs, A. Extant genus of flat bark beetle (Coleoptera: Silvanidae) with a present-day Australian-southern South American disjunction discovered in Eocene Rovno amber. Zootaxa 2022, 5129, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Telnov, D.; Perkovsky, E.E.; Vasilenko, D.V.; Yamamoto, S. The first fossil Coleoptera record from the Volyn Region, Ukraine, with description of a new Glesoconomorphus (Coleoptera, Mycteridae) in syninclusion with Winterschmidtiidae (Acari) and a key to species. ZooKeys 2021, 1068, 189–201. [Google Scholar] [CrossRef]
- Telnov, D.; Perkovsky, E.E.; Kundrata, R.; Kairišs, K.; Vasilenko, D.V.; Bukejs, A. Revealing Palaeogene distribution of the Ptilodactylidae (Insecta: Coleoptera): The first Ptilodactyla Illiger, 1807 records from Rovno amber of Ukraine. Hist Biol. 2022, in press. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new species of the genus Glaesotropis Gratshev and Zherikhin, 1995 (Coleoptera, Anthribidae) from Rovno amber. Foss. Rec. 2021, 24, 1–7. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. Ceutorhynchus Germar (Coleoptera, Curculionidae) as proxy for Eocene Brassicaceae: First record of the genus from Rovno amber. J. Paleontol. 2022, 96, 379–386. [Google Scholar] [CrossRef]
- Legalov, A.A.; Vasilenko, D.V.; Perkovsky, E.E. The American tribes Anypotactini and Eudiagogini (Coleoptera, Curculionidae) in Eocene of Europe as indicators of Eocene climate with description a new species. Diversity 2022, 14, 767. [Google Scholar] [CrossRef]
- Legalov, A.A.; Vasilenko, D.V.; Perkovsky, E.E. New proxy for Moraceae in Priabonian of Europe: First record of the genus Demimaea Pascoe, 1870 (Coleoptera: Curculionidae) from Eocene Rovno amber. Hist. Biol. 2022, 1–7. [Google Scholar] [CrossRef]
- Legalov, A.A. Fossil history of bark-beetles (Coleoptera: Scolytidae) with descriptions of two new species. Hist. Biol. 2022, 1–11. [Google Scholar] [CrossRef]
- Lyubarsky, G.Y.; Bukejs, A. Two new fossil species of the genus Atomaria Stephens (Coleoptera: Cryptophagidae) from Eocene European amber with a key to species described from fossil resins. Zootaxa 2022, 5188, 283–289. [Google Scholar] [CrossRef]
- Matalin, A.V.; Perkovsky, E.E.; Vasilenko, D.V. First record of tiger beetles (Coleoptera, Cicindelidae) from Rovno amber with the description of a new genus and species. Zootaxa 2021, 5016, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, S.V.; Bocak, L. New genus of erotine net-winged beetles, Damzenium gen. nov. (Coleoptera: Lycidae), from Eocene Rovno amber. Zootaxa 2022, 5154, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Kirichenko-Babko, M.; Perkovsky, E.E.; Vasilenko, D.V. A new genus and species of Lebiini (Coleoptera: Carabidae) from late Eocene Rovno amber. Hist. Biol. 2022, 34, 436–442. [Google Scholar] [CrossRef]
- Kirichenko-Babko, M.; Perkovsky, E.E.; Vasilenko, D.V. Antephilorhizus zerovae sp. nov. (Carabidae: Lebiini), the second ground beetle species from Rovno amber. Hist. Biol. 2022, 34, 2424–2429. [Google Scholar] [CrossRef]
- Schmidt, J.; Scholz, S.; Will, K. Character analysis and descriptions of Eocene sphodrine fossils (Coleoptera, Carabidae) using light microscopy, micro-CT scanning, and 3D imaging. Dtsch. Entomol. Z. 2022, 69, 19–44. [Google Scholar] [CrossRef]
- Vitali, F.; Perkovsky, E.E. Poliaenus europaeus n. sp., the first cerambycid from Rovno amber (Coleoptera Cerambycidae). Hist. Biol. 2022, in press. [Google Scholar] [CrossRef]
- Haug, C.; Baranov, V.A.; Hörnig, M.K.; Gauweiler, J.; Hammel, J.U.; Perkovsky, E.E.; Haug, J.T. 35 million-year-old solid-wood-borer beetle larvae support the idea of stressed Eocene amber forests. Palaeobiodivers. Palaeoenviron. 2022, 1–10. [Google Scholar] [CrossRef]
- Yamamoto, S.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. First fossil species of ship-timber beetles (Coleoptera: Lymexylidae) from Eocene Rovno amber (Ukraine). Foss. Rec. 2022, 25, 65–74. [Google Scholar] [CrossRef]
- Nabozhenko, M.V.; Perkovsky, E.E. Oracula campbelli sp. n.—The first fossil darkling beetle (Coleoptera: Tenebrionidae: Alleculinae) from the late Eocene Rovno amber (Ukraine). Zootaxa 2023, 5230, 245–250. [Google Scholar] [CrossRef]
- Lyubarsky, G.Y.; Perkovsky, E.E. New findings of Cryptophagidae (Coleoptera: Clavicornia) from Baltic amber in the unbiased collection of the Paleontological Institute of RAS. Russ. Entomol. J. 2021, 30, 282–287. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Odnosum, V.K.; Nazarenko, V.Yu.; Vasilenko, D.V. First record of the genus Cyrtanaspis Emery (Insecta: Coleoptera: Scraptiidae) from Baltic amber. Paleontol. J. 2022, 56, 208–212. [Google Scholar] [CrossRef]
- Simutnik, S.A.; Perkovsky, E.E.; Vasilenko, D.V. Protaphycus shuvalikovi Simutnik gen. et sp. n. (Chalcidoidea, Encyrtidae, Encyrtinae) from Rovno amber. J. Hymenopt. Res. 2022, 91, 1–9. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Nel, A. A new Rovno amber termite genus (Isoptera, Rhinotermitidae) from Styr River basin. Palaeont. Electron. 2021, 24, a05. [Google Scholar] [CrossRef] [PubMed]
- Giłka, W.; Harbach, R.E.; Perkovsky, E.E. Mosquitoes (Diptera: Culicidae) in Eocene amber from the Rovno region, Ukraine. Zootaxa 2021, 5016, 257–270. [Google Scholar] [CrossRef]
- Olmi, M.; Guglielmino, A.; Vasilenko, D.V.; Perkovsky, E.E. Discovery of the first apterous pincer wasp from amber, with description of a new tribe, genus and species of Apodryininae (Hymenoptera, Dryinidae). Zootaxa 2022, 5162, 54–66. [Google Scholar] [CrossRef]
- Gimmel, M.L.; Ferro, M.L. General overview of saproxylic Coleoptera. In Saproxylic Insects: Diversity, Ecology and Conservation; Ulyshen, M.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 51–128. [Google Scholar]
- Horion, A. Faunistik der Mitteleuropäischer Käfer. Band VII.; Clavicornia, I. Teil (Sphaeritidae bis Phalacridae); Feyel: Überlingen, Germany, 1960; p. 346. [Google Scholar]
- Ivanov, V.D.; Melnitsky, S.I.; Perkovsky, E.E. Caddisflies from Cenozoic resins of Europe. Paleontol. J. 2016, 50, 485–493. [Google Scholar] [CrossRef]
- Radchenko, A.G.; Perkovsky, E.E. Wheeler’s dilemma revisited: First Oecophylla–Lasius syninclusion and other ant syninclusions in the Bitterfeld amber (late Eocene). Invertebr. Zool. 2021, 18, 47–65. [Google Scholar] [CrossRef]
- Radchenko, A.G.; Perkovsky, E.E.; Vasilenko, D.V. Formica species (Hymenoptera, Formicidae, Formicinae) in late Eocene Rovno amber. J. Hymen. Res. 2021, 82, 237–251. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Wegierek, P. Aphid-Buchnera-Ant symbiosis; or why are aphids rare in the tropics and very rare further south? Earth. Environ. Sci. Trans. R. Soc. Edinb. 2018, 107, 297–310. [Google Scholar] [CrossRef]
- Hilszczański, J.; Jaworski, T.; Plewa, R.; Ługowoj, J. Zavaljus brunneus (Gyllenhal, 1808)—a beetle species new to the Polish fauna (Coleoptera: Erotylidae). Genus 2014, 25, 421–424. [Google Scholar]
- Mänd, K.; Muehlenbachs, K.; McKellar, R.C.; Wolfe, A.P.; Konhauser, K.O. Distinct origins for Rovno and Baltic ambers: Evidence from carbon and hydrogen stable isotopes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 265–273. [Google Scholar] [CrossRef]
- Perkovsky, E.E. A new species of Micromalthidae (Coleoptera) from the Rovno amber: 1. Adult morphology. Paleontol. J. 2016, 50, 293–296. [Google Scholar] [CrossRef]
- Legalov, A.A.; Nazarenko, V.Y.; Perkovsky, E.E. A new genus of fungus weevils (Coleoptera: Anthribidae) in Rovno amber. Foss. Rec. 2018, 21, 207–212. [Google Scholar] [CrossRef]
- Olmi, M.; Eggs, B.; Capradossi, L.; van de Kamp, T.; Perkovsky, E.E.; Guglielmino, A.; Vasilenko, D.V. A new species of Bocchus from upper Eocene Rovno amber (Hymenoptera: Dryinidae). J. Hymenopt. Res. 2022, 92, 257–272. [Google Scholar] [CrossRef]
- Gumovsky, A.; Perkovsky, E.; Rasnitsyn, A. Laurasian ancestors and “Gondwanan” descendants of Rotoitidae (Hymenoptera: Chalcidoidea): What a review of Late Cretaceous Baeomorpha revealed. Cretac. Res. 2018, 84, 286–322. [Google Scholar] [CrossRef]
- Kolesnikov, V.B.; Turbanov, I.S.; Eskov, K.Yu.; Propistsova, E.A.; Bashkuev, A.S. First non-amber Mesozoic pseudoscorpion from Upper Triassic deposits of eastern Europe, with a description of two new fossil subfamilies (Arachnida, Pseudoscorpiones, Feaellidae). Pap. Palaeontol. 2022, 8, e1466. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyubarsky, G.Y.; Perkovsky, E.E.; Vasilenko, D.V. Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber. Life 2023, 13, 636. https://doi.org/10.3390/life13030636
Lyubarsky GY, Perkovsky EE, Vasilenko DV. Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber. Life. 2023; 13(3):636. https://doi.org/10.3390/life13030636
Chicago/Turabian StyleLyubarsky, Georgy Yu., Evgeny E. Perkovsky, and Dmitry V. Vasilenko. 2023. "Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber" Life 13, no. 3: 636. https://doi.org/10.3390/life13030636
APA StyleLyubarsky, G. Y., Perkovsky, E. E., & Vasilenko, D. V. (2023). Unexpected Diversity of Xenoscelinae in Priabonian European Amber: The Third Xenosceline Species from Rovno Amber. Life, 13(3), 636. https://doi.org/10.3390/life13030636