
Combined Transcriptome and Lipidomic Analyses of Lipid Biosynthesis in Macadamia ternifolia Nuts

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. RNA Sequencing, De Novo Assembly, and Annotations
2.2. Transcriptome Changes in Different Fruit Kernel Stages
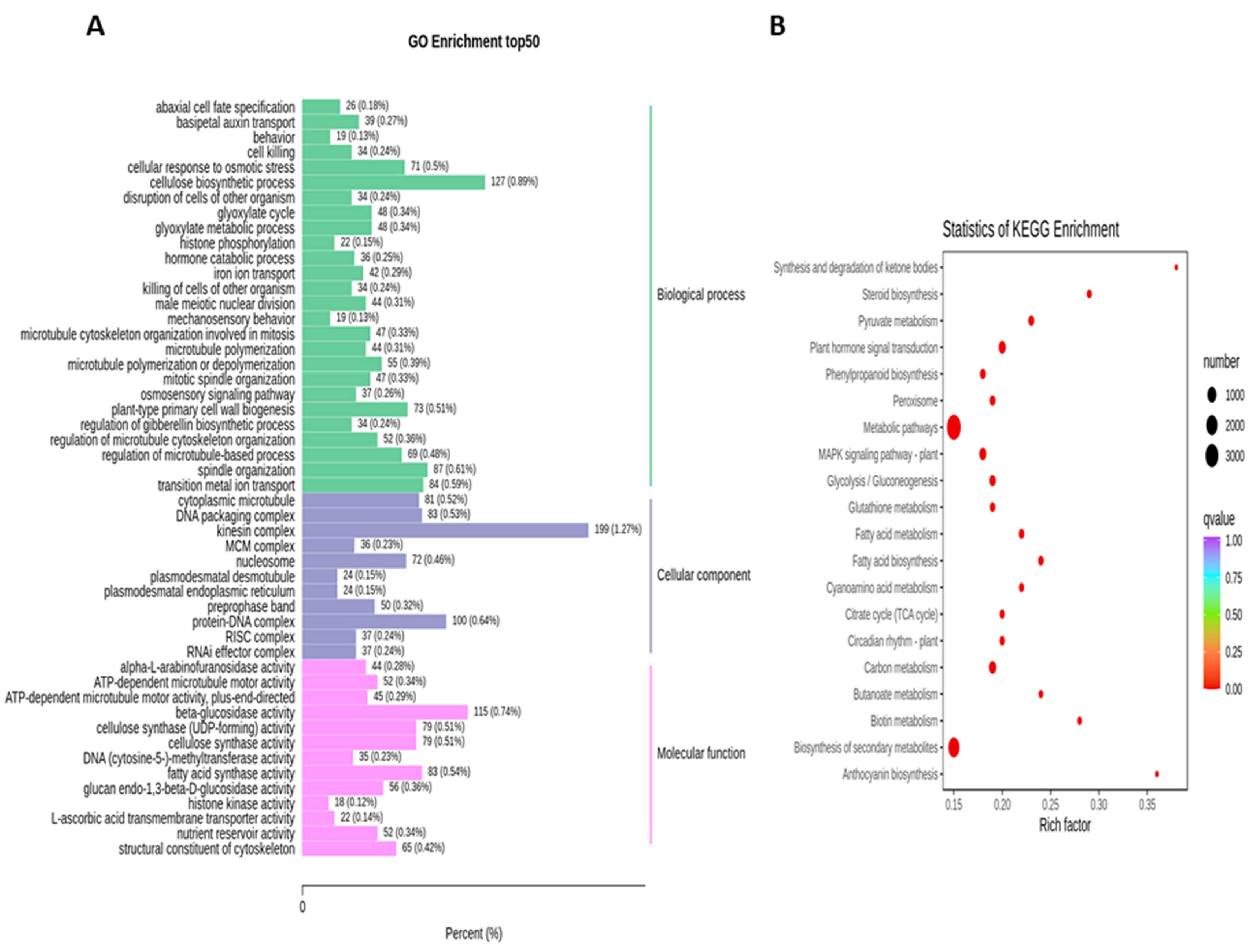
2.3. Functional Enrichment Analysis of DEGs
2.4. Overview of the Lipid Composition at Different Fruit Kernel Developmental Stages
2.5. Lipid-Related Gene Expression Changes during Fruit Kernel Development
2.6. Validation of Key Genes Involved in Lipid Regulation Using qRT-PCR Analysis
3. Discussion
3.1. Divergence of Lipid Composition in the Kernels of M. ternifolia
3.2. Lipid Biosynthesis in Fruit Kernels of M. ternifolia
4. Materials and Methods
4.1. Plant Materials, RNA Extraction, and Sequencing
4.2. Transcript Assembly, Annotation, and Analysis of DEGs
4.3. Real-Time qRT-PCR Analysis
4.4. Lipid Extraction and Analysis Using UPLC-MS/MS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Niu, Y.-F.; Li, G.-H.; Ni, S.-B.; He, X.-Y.; Zheng, C.; Liu, Z.-Y.; Gong, L.-D.; Liu, J. Genome assembly and annotation of Macadamia tetraphylla. bioRxiv 2020. [Google Scholar] [CrossRef]
- Toft, B.D.; Alam, M.; Topp, B. Estimating genetic parameters of architectural and reproductive traits in young macadamia cultivars. Tree Genet. Genomes 2018, 14, 1–10. [Google Scholar] [CrossRef]
- Mast, A.R.; Willis, C.L.; Jones, E.H.; Downs, K.M.; Weston, P.H. A smaller Macadamia from a more vagile tribe: Inference of phylogenetic relationships, divergence times, and diaspore evolution in Macadamia and relatives (tribe Macadamieae; Proteaceae). Am. J. Bot. 2008, 95, 843–870. [Google Scholar] [CrossRef]
- Hu, W.; Fitzgerald, M.; Topp, B.; Alam, M.; O’Hare, T.J. A review of biological functions, health benefits, and possible de novo biosynthetic pathway of palmitoleic acid in macadamia nuts. J. Funct. Foods 2019, 62, 103520. [Google Scholar] [CrossRef]
- Navarro, S.L.; Rodrigues, C.E. Macadamia oil extraction methods and uses for the defatted meal byproduct. Trends Food Sci. Technol. 2016, 54, 148–154. [Google Scholar] [CrossRef]
- Insanu, M.; Hartati, R.; Bajri, F.; Fidrianny, I. Macadamia Genus: An Updated Review of Phytochemical Compounds and Pharmacological Activities. Biointerface Res. Appl. Chem. 2021, 11, 14898–14905. [Google Scholar]
- Hong, M.Y.; Groven, S.; Marx, A.; Rasmussen, C.; Beidler, J.J.M. Anti-inflammatory, antioxidant, and hypolipidemic effects of mixed nuts in atherogenic diet-fed rats. Molecules 2018, 23, 3126. [Google Scholar] [CrossRef] [PubMed]
- Hardner, C.M.; Wall, M.; Cho, A.J.H. Global macadamia science: Overview of the special section. HortScience 2019, 54, 592–595. [Google Scholar] [CrossRef]
- Hardner, C.; e Silva, J.C.; Williams, E.; Meyers, N.; McConchie, C.J.H. Breeding new cultivars for the Australian macadamia industry. HortScience 2019, 54, 621–628. [Google Scholar] [CrossRef]
- Penoni, E.d.S.; Pio, R.; Rodrigues, F.A.; Maro, L.A.C.; Costa, F.C.J.C.R. Analysis of fruits and nuts of macadamia walnut cultivars. Ciência Rural 2011, 41, 2080–2083. [Google Scholar] [CrossRef]
- Piza, P.L.B.T.; Moriya, L.M. Cultivo da macadâmia no Brasil. Rev. Bras. Frutic. 2014, 36, 39–45. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Sathe, S.K. Chemical composition of selected edible nut seeds. J. Agric. Food Chem. 2006, 54, 4705–4714. [Google Scholar] [CrossRef]
- Rengel, A.; Pérez, E.; Piombo, G.; Ricci, J.; Servent, A.; Tapia, M.S.; Gibert, O.; Montet, D. Lipid profile and antioxidant activity of macadamia nuts (Macadamia integrifolia) cultivated in Venezuela. Nat. Sci. 2015, 12, 535–547. [Google Scholar]
- Knothe, G. Biodiesel derived from a model oil enriched in palmitoleic acid, macadamia nut oil. Energy Fuels 2010, 24, 2098–2103. [Google Scholar] [CrossRef]
- Wang, D.D.; Hu, F.B. Dietary fat and risk of cardiovascular disease: Recent controversies and advances. Annu. Rev. Nutr. 2017, 37, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.L.; Blake, R.J.; Wills, R.B. Macadamia nut consumption lowers plasma total and LDL cholesterol levels in hypercholesterolemic men. J. Nutr. 2003, 133, 1060–1063. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Yu-Poth, S.; Sabaté, J.; Ratcliffe, H.E.; Zhao, G.; Etherton, T.D. Nuts and their bioactive constituents: Effects on serum lipids and other factors that affect disease risk. Am. J. Clin. Nutr. 1999, 70, 504s–511s. [Google Scholar] [CrossRef] [PubMed]
- Vadivel, V.; Kunyanga, C.N.; Biesalski, H.K. Health benefits of nut consumption with special reference to body weight control. Nutrition 2012, 28, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Kaijser, A.; Dutta, P.; Savage, G. Oxidative stability and lipid composition of macadamia nuts grown in New Zealand. Food Chem. 2000, 71, 67–70. [Google Scholar] [CrossRef]
- Kochhar, S.P.; Henry, C.J.K. Oxidative stability and shelf-life evaluation of selected culinary oils. Int. J. Food Sci. Nutr. 2009, 60, 289–296. [Google Scholar] [CrossRef]
- Yoon, W.-J.; Kim, M.-J.; Moon, J.-Y.; Kang, H.-J.; Kim, G.-O.; Lee, N.H.; Hyun, C.-G. Effect of palmitoleic acid on melanogenic protein expression in murine b16 melanoma. J. Oleo Sci. 2010, 59, 315–319. [Google Scholar] [CrossRef]
- Kuo, T.M.; Gardner, H. Perspectives on the Production of Industrial Oils in Genetically Engineered Oilseeds. In Lipid Biotechnology; CRC Press: Boca Raton, FL, USA, 2002; pp. 107–117. [Google Scholar]
- Drackley, J. Lipid Metabolism; CABI: Wallingford, UK, 2000; Volume 1, pp. 97–119. [Google Scholar]
- Mutz, K.-O.; Heilkenbrinker, A.; Lönne, M.; Walter, J.-G.; Stahl, F. Transcriptome analysis using next-generation sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K.; Zhang, X.; Guo, L.; Qi, T.; Tang, H.; Zhang, M.; Zhang, B.; Wang, H.; Qiao, X.; Feng, J. Comparative transcriptome analysis of inbred lines and contrasting hybrids reveals overdominance mediate early biomass vigor in hybrid cotton. BMC Genom. 2020, 21, 1–16. [Google Scholar]
- Beisson, F.; Koo, A.J.; Ruuska, S.; Schwender, J.; Pollard, M.; Thelen, J.J.; Paddock, T.; Salas, J.J.; Savage, L.; Milcamps, A. Arabidopsis genes involved in acyl lipid metabolism. A 2003 census of the candidates, a study of the distribution of expressed sequence tags in organs, and a web-based database. Plant Physiol. 2003, 132, 681–697. [Google Scholar] [PubMed]
- Severin, A.J.; Woody, J.L.; Bolon, Y.-T.; Joseph, B.; Diers, B.W.; Farmer, A.D.; Muehlbauer, G.J.; Nelson, R.T.; Grant, D.; Specht, J.E. RNA-Seq Atlas of Glycine max: A guide to the soybean transcriptome. BMC Plant Biol. 2010, 10, 1–16. [Google Scholar] [CrossRef]
- O’Hara, P.; Slabas, A.R.; Fawcett, T. Fatty acid and lipid biosynthetic genes are expressed at constant molar ratios but different absolute levels during embryogenesis. Plant Physiol. 2002, 129, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.; Yi, C.; Tian, D.; Sangha, J.S.; Hong, Y.; Yin, Z. Expression of fatty acid and lipid biosynthetic genes in developing endosperm of Jatropha curcas. Biotechnol. Biofuels 2012, 5, 1–15. [Google Scholar] [CrossRef]
- Gupta, K.; Kayam, G.; Faigenboim-Doron, A.; Clevenger, J.; Ozias-Akins, P.; Hovav, R. Gene expression profiling during seed-filling process in peanut with emphasis on oil biosynthesis networks. Plant Sci. 2016, 248, 116–127. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, K.; Wu, C.; Zhao, Y.; Yin, X.; Zhang, B.; Grierson, D.; Chen, K.; Xu, C. Effect of ethylene on cell wall and lipid metabolism during alleviation of postharvest chilling injury in peach. Cells 2019, 8, 1612. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.P.; Kroon, J.T.; Swarbreck, D.; Febrer, M.; Larson, T.R.; Graham, I.A.; Caccamo, M.; Slabas, A.R. Tissue-specific whole transcriptome sequencing in castor, directed at understanding triacylglycerol lipid biosynthetic pathways. PLoS ONE 2012, 7, e30100. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ni, J.; Shah, F.A.; Wang, Q.; Wang, Z.; Wu, L.; Fu, S. Transcriptome analysis of pecan seeds at different developing stages and identification of key genes involved in lipid metabolism. PLoS ONE 2018, 13, e0195913. [Google Scholar] [CrossRef]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Dussert, S.; Guerin, C.; Andersson, M.; Joët, T.; Tranbarger, T.J.; Pizot, M.; Sarah, G.; Omore, A.; Durand-Gasselin, T.; Morcillo, F. Comparative transcriptome analysis of three oil palm fruit and seed tissues that differ in oil content and fatty acid composition. Plant Physiol. 2013, 162, 1337–1358. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Simpson, J.P.; Li-Beisson, Y.; Beisson, F.; Pollard, M.; Ohlrogge, J.B. A land-plant-specific glycerol-3-phosphate acyltransferase family in Arabidopsis: Substrate specificity, sn-2 preference, and evolution. Plant Physiol. 2012, 160, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Welti, R.; Shah, J. The Arabidopsis thaliana dihydroxyacetone phosphate reductase gene Suppressor of fatty acid desaturase deficiency1 is required for glycerolipid metabolism and for the activation of systemic acquired resistance. Plant Cell 2004, 16, 465–477. [Google Scholar] [CrossRef]
- Li, R.-J.; Gao, X.; Li, L.-M.; Liu, X.-L.; Wang, Z.-Y.; Lü, S.-Y. De novo assembly and characterization of the fruit transcriptome of Idesia polycarpa reveals candidate genes for lipid biosynthesis. Front. Plant Sci. 2016, 7, 801. [Google Scholar] [CrossRef]
- Nock, C.J.; Baten, A.; Barkla, B.J.; Furtado, A.; Henry, R.J.; King, G.J. Genome and transcriptome sequencing characterises the gene space of Macadamia integrifolia (Proteaceae). BMC Genom. 2016, 17, 1–12. [Google Scholar] [CrossRef]
- Howlett, B.G.; Nelson, W.R.; Pattemore, D.E.; Gee, M. Pollination of macadamia: Review and opportunities for improving yields. Sci. Hortic. 2015, 197, 411–419. [Google Scholar] [CrossRef]
- Monaghan, E.K. Chemical Composition and Protein Antigenicity–Almond (Prunus dulcis) and Macadamia Nut (Macadamia integrifolia) Seeds; The Florida State University: Tallahassee, FL, USA, 2008. [Google Scholar]
- Tu, X.-H.; Wu, B.-f.; Xie, Y.; Xu, S.-L.; Wu, Z.-Y.; Lv, X.; Wei, F.; Du, L.-Q.; Chen, H. A comprehensive study of raw and roasted macadamia nuts: Lipid profile, physicochemical, nutritional, and sensory properties. Food Sci. Nutr. 2021, 9, 1688–1697. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Y.; Wu, G.; Tang, W.; Peng, Z.; Bu, X.; Chao, L.; Yin, X.; Xiong, J.; Zhang, H.; Zhao, X. Oil biosynthesis and transcriptome profiles in developing endosperm and oil characteristic analyses in Paeonia ostii var. lishizhenii. J. Plant Physiol. 2018, 228, 121–133. [Google Scholar] [CrossRef]
- Kaup, M.T.; Froese, C.D.; Thompson, J.E. A role for diacylglycerol acyltransferase during leaf senescence. Plant Physiol. 2002, 129, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Hsieh, K.; Ratnayake, C.; Huang, A.H. A novel group of Oleosins is present inside the pollen ofArabidopsis. J. Biol. Chem. 2002, 277, 22677–22684. [Google Scholar] [CrossRef]
- Yu, S.-Y.; Zhang, Y.; Lyu, Y.-P.; Yao, Z.-J.; Hu, Y.-H. Lipidomic profiling of the developing kernel clarifies the lipid metabolism of Paeonia ostii. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Butka, E.; Wang, X. Comprehensive quantification of triacylglycerols in soybean seeds by electrospray ionization mass spectrometry with multiple neutral loss scans. Sci. Rep. 2014, 4, 1–11. [Google Scholar] [CrossRef]
- Woodfield, H.K.; Cazenave-Gassiot, A.; Haslam, R.P.; Guschina, I.A.; Wenk, M.R.; Harwood, J.L. Using lipidomics to reveal details of lipid accumulation in developing seeds from oilseed rape (Brassica napus L.). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Poxleitner, M.; Rogers, S.W.; Lacey Samuels, A.; Browse, J.; Rogers, J.C. A role for caleosin in degradation of oil-body storage lipid during seed germination. Plant J. 2006, 47, 917–933. [Google Scholar] [CrossRef]
- Volk, G.M.; Crane, J.; Caspersen, A.M.; Hill, L.M.; Gardner, C.; Walters, C. Massive cellular disruption occurs during early imbibition of Cuphea seeds containing crystallized triacylglycerols. Planta 2006, 224, 1415–1426. [Google Scholar] [CrossRef]
- Lin, J.-T.; Liu, S.-C.; Hu, C.-C.; Shyu, Y.-S.; Hsu, C.-Y.; Yang, D.-J. Effects of roasting temperature and duration on fatty acid composition, phenolic composition, Maillard reaction degree and antioxidant attribute of almond (Prunus dulcis) kernel. Food Chem. 2016, 190, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.M. Functional lipid characteristics, oxidative stability, and antioxidant activity of macadamia nut (Macadamia integrifolia) cultivars. Food Chem. 2010, 121, 1103–1108. [Google Scholar] [CrossRef]
- Colzato, M.; Scramin, J.; Forato, L.; Colnago, L.; Assis, O. 1h Nmr investigation of oil oxidation in macadamia nuts coated with zein-based films. J. Food Process. Preserv. 2011, 35, 790–796. [Google Scholar] [CrossRef]
- Ha, K.S.; Thompson, G.A., Jr. Diacylglycerol metabolism in the green alga Dunaliella salina under osmotic stress: Possible role of diacylglycerols in phospholipase C-mediated signal transduction. Plant Physiol. 1991, 97, 921–927. [Google Scholar] [CrossRef][Green Version]
- Degenkolbe, T.; Giavalisco, P.; Zuther, E.; Seiwert, B.; Hincha, D.K.; Willmitzer, L. Differential remodeling of the lipidome during cold acclimation in natural accessions of Arabidopsis thaliana. Plant J. 2012, 72, 972–982. [Google Scholar] [CrossRef]
- Chen, M.; Thelen, J.J. Acyl-lipid desaturase2 is required for chilling and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 1430–1444. [Google Scholar] [CrossRef]
- Zhang, Q.; Lin, F.; Mao, T.; Nie, J.; Yan, M.; Yuan, M.; Zhang, W. Phosphatidic acid regulates microtubule organization by interacting with MAP65-1 in response to salt stress in Arabidopsis. Plant Cell 2012, 24, 4555–4576. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, Y.; Shao, Y.; Li, M.; Ma, F. Comprehensive genomic analysis and expression profiling of diacylglycerol kinase gene family in Malus prunifolia (Willd.) Borkh. Gene 2015, 561, 225–234. [Google Scholar] [CrossRef]
- Ruelland, E.; Cantrel, C.; Gawer, M.; Kader, J.-C.; Zachowski, A. Activation of phospholipases C and D is an early response to a cold exposure in Arabidopsis suspension cells. Plant Physiol. 2002, 130, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yao, N.; Song, J.T.; Luo, S.; Lu, H.; Greenberg, J.T. Ceramides modulate programmed cell death in plants. Genes Dev. Cell 2003, 17, 2636–2641. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P. Acyl-lipid metabolism. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2013; Volume 11. [Google Scholar]
- Andre, C.; Froehlich, J.E.; Moll, M.R.; Benning, C. A heteromeric plastidic pyruvate kinase complex involved in seed oil biosynthesis in Arabidopsis. Plant Cell 2007, 19, 2006–2022. [Google Scholar] [CrossRef]
- Cai, Y.; Li, S.; Jiao, G.; Sheng, Z.; Wu, Y.; Shao, G.; Xie, L.; Peng, C.; Xu, J.; Tang, S. Os PK 2 encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, compound granule formation and grain filling. Plant Biotechnol. J. 2018, 16, 1878–1891. [Google Scholar] [CrossRef]
- Moreno-Pérez, A.J.; Sánchez-García, A.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Acyl-ACP thioesterases from macadamia (Macadamia tetraphylla) nuts: Cloning, characterization and their impact on oil composition. Plant Physiol. Biochem. 2011, 49, 82–87. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Lal, M.A. Plant Physiology, Development and Metabolism; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Massengo-Tiassé, R.P.; Cronan, J.E. Diversity in enoyl-acyl carrier protein reductases. Cell. Mol. Life Sci. 2009, 66, 1507–1517. [Google Scholar] [CrossRef]
- Cagliari, A.; Margis-Pinheiro, M.; Loss, G.; Mastroberti, A.A.; de Araujo Mariath, J.E.; Margis, R. Identification and expression analysis of castor bean (Ricinus communis) genes encoding enzymes from the triacylglycerol biosynthesis pathway. Plant Sci. 2010, 179, 499–509. [Google Scholar] [CrossRef]
- Eichmann, T.O.; Lass, A. DAG tales: The multiple faces of diacylglycerol-stereochemistry, metabolism, and signaling. Cell. Mol. Life Sci. 2015, 72, 3931–3952. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Su, Y.; Wang, X. Phosphatidic acid-mediated signaling. Lipid Mediat. Protein Signal. 2013, 991, 159–176. [Google Scholar]
- Wang, X.; Chapman, K.D. Lipid signaling in plants. Front. Plant Sci. 2013, 4, 216. [Google Scholar] [CrossRef]
- Akoh, C.C.; Lee, G.-C.; Liaw, Y.-C.; Huang, T.-H.; Shaw, J.-F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, F.; Huang, X.; Wang, W.; Zhang, S.; Feng, F. Identification and integrated analysis of mRNAs, lncRNAs, and microRNAs of developing seeds in high oleic acid sunflower (Helianthus annuus L.). Acta Physiol. Plant. 2021, 43, 1–13. [Google Scholar] [CrossRef]
- Oenel, A.; Fekete, A.; Krischke, M.; Faul, S.C.; Gresser, G.; Havaux, M.; Mueller, M.J.; Berger, S. Enzymatic and non-enzymatic mechanisms contribute to lipid oxidation during seed aging. Plant Cell Physiol. 2017, 58, 925–933. [Google Scholar]
- Opassiri, R.; Pomthong, B.; Onkoksoong, T.; Akiyama, T.; Esen, A.; Cairns, J.R.K. Analysis of rice glycosyl hydrolase family 1 and expression of Os4bglu12 β-glucosidase. BMC Plant Biol. 2006, 6, 1–19. [Google Scholar] [CrossRef]
- Ketudat Cairns, J.R.; Esen, A. β-Glucosidases. Cell. Mol. Life Sci. 2010, 67, 3389–3405. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Dossa, K.; Zhou, R.; Li, D.; Liu, A.; Qin, L.; Mmadi, M.A.; Su, R.; Zhang, Y.; Wang, J.; Gao, Y. A novel motif in the 5′-UTR of an orphan gene ‘Big Root Biomass’ modulates root biomass in sesame. Plant Biotechnol. J. 2021, 19, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Liu, H.; Zhang, J.; Lyu, Y.; Li, Z.; He, Y.; Zhang, X.; Deng, X.; Brotman, Y.; Fernie, A.R. Lipidomic and transcriptomic analysis reveals reallocation of carbon flux from cuticular wax into plastid membrane lipids in a glossy “Newhall” navel orange mutant. Hortic. Res. 2020, 7, 1–18. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Base (Gb) | Mapped Reads (%) | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|---|
| S1-1 | 55,604,268 | 53,433,146 | 8.0 | 85 | 97.9 | 93.8 | 44.8 |
| S1-2 | 56,740,048 | 54,978,858 | 8.3 | 83 | 97.5 | 92.8 | 45.4 |
| S1-3 | 50,745,980 | 49,452,818 | 7.4 | 83 | 97.9 | 93.7 | 45.2 |
| S2-1 | 49,242,862 | 45,820,088 | 6.8 | 82 | 97.8 | 93.5 | 44.1 |
| S2-2 | 46,406,958 | 44,626,700 | 6.7 | 82 | 97.8 | 93.5 | 44.3 |
| S2-3 | 48,601,196 | 46,373,882 | 7.0 | 81 | 97.9 | 93.7 | 44.1 |
| S3-1 | 55,107,466 | 53,593,048 | 8.0 | 79 | 97.8 | 93.6 | 45.7 |
| S3-2 | 46,080,428 | 44,529,822 | 6.7 | 78 | 97.8 | 93.6 | 45.2 |
| S3-3 | 56,758,326 | 54,256,348 | 8.1 | 76 | 98.0 | 94.2 | 44.0 |
| ID | S1 | S2 | S3 | Annotations |
|---|---|---|---|---|
| Unigene 73227 | 61.9 | 1.8 | 0.0 | 3-ketoacyl-acyl carrier protein reductase |
| Unigene 211387 | 11.5 | 0.2 | 0.0 | 3-ketoacyl-acyl carrier protein reductase |
| Unigene 73228 | 10.6 | 0.0 | 0.0 | 3-ketoacyl-acyl carrier protein reductase |
| Unigene 163200 | 0.0 | 1.2 | 0.0 | 3-ketoacyl-acyl carrier protein reductase |
| Unigene 26385 | 41.6 | 0.0 | 0.0 | 3-ketoacyl-acyl carrier protein reductase |
| Unigene 170150 | 108.4 | 640.6 | 3.6 | Beta-glucosidase 11 |
| Unigene 41085 | 27.4 | 0.2 | 0.3 | Beta-glucosidase 11 |
| Unigene 81638 | 0.3 | 18.4 | 0.0 | Beta-glucosidase 12 |
| Unigene 81652 | 1.3 | 22.1 | 0.0 | Beta-glucosidase 12 |
| Unigene 91684 | 0.0 | 0.8 | 0.0 | Beta-glucosidase 44 |
| Unigene 35002 | 2.6 | 0.1 | 0.0 | Beta-glucosidase 41 |
| Unigene 101847 | 0.0 | 0.0 | 1.7 | Choline/ethanolaminephosphotransferase 1 |
| Unigene 81642 | 56.3 | 479.2 | 0.1 | Cyanogenic beta-glucosidase |
| Unigene 147364 | 0.0 | 5.8 | 2.7 | Diacylglycerol kinase 2 |
| Unigene 150581 | 0.0 | 0.7 | 0.0 | Enoyl-acyl-carrier-protein reductase (NADH) |
| Unigene 150580 | 0.0 | 1.4 | 0.0 | Enoyl-acyl-carrier-protein reductase (NADH) |
| Unigene 140441 | 0.0 | 3.0 | 0.0 | Enoyl-acyl-carrier-protein reductase (NADH) |
| Unigene 240247 | 4.6 | 0.0 | 0.1 | GDSL esterase lipase |
| Unigene 213504 | 3.7 | 10.1 | 0.0 | GDSL esterase lipase |
| Unigene 82774 | 7.5 | 5.7 | 0.0 | GDSL esterase lipase |
| Unigene 183098 | 6.6 | 0.9 | 0.0 | GDSL esterase lipase |
| Unigene 62756 | 0.0 | 3.2 | 0.1 | GDSL esterase lipase |
| Unigene 38911 | 1.4 | 0.0 | 0.0 | GDSL esterase lipase |
| Unigene 113245 | 1.4 | 0.0 | 0.0 | GDSL esterase lipase |
| Unigene 238268 | 12.8 | 0.5 | 0.0 | GDSL esterase lipase |
| Unigene 69671 | 18.5 | 40.2 | 0.5 | GDSL esterase lipase APG |
| Unigene 103298 | 5.3 | 11.0 | 0.0 | GDSL esterase lipase APG |
| Unigene 152617 | 1.5 | 0.0 | 0.0 | Glycerol-3-phosphate acyltransferase |
| Unigene 128368 | 1.1 | 0.0 | 0.0 | Glycerol-3-phosphate acyltransferase |
| Unigene 163240 | 1.8 | 1.0 | 0.0 | Hydroxymethylglutaryl-CoA synthase |
| Unigene 122230 | 71.4 | 2.0 | 60.9 | Inositol-3-phosphate synthase |
| Unigene 9629 | 0.7 | 0.0 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 66716 | 9.1 | 0.1 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 54520 | 1.3 | 0.0 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 59928 | 20.7 | 0.1 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 213589 | 5.5 | 0.0 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 37584 | 19.1 | 0.3 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 34267 | 15.0 | 0.1 | 0.0 | Linoleate 13S-lipoxygenase 2-1 |
| Unigene 13471 | 2.4 | 0.1 | 0.0 | Linoleate 9S-lipoxygenase A |
| Unigene 5404 | 0.9 | 0.0 | 0.0 | Linoleate 9S-lipoxygenase A |
| Unigene 139752 | 0.5 | 0.0 | 0.5 | Lipoxygenase 4 |
| Unigene 227869 | 4.6 | 11.8 | 0.1 | Phosphatidylinositol 4-kinase alpha 1 |
| Unigene 171746 | 72.5 | 95.5 | 0.6 | Phospholipase A2-alpha |
| Unigene 56285 | 0.0 | 0.0 | 0.8 | Phospholipid hydroperoxide |
| Unigene 63669 | 24.8 | 20.0 | 0.5 | Pyruvate kinase 2 |
| Unigene 63675 | 2.4 | 2.4 | 0.0 | Pyruvate kinase 2 |
| Unigene 86831 | 2.8 | 1.0 | 0.0 | Pyruvate kinase 2 |
| Unigene 158347 | 4.21 | 0.00 | 0.26 | Non-specific phospholipase C6; EC |
| Unigene 190439 | 5.87 | 6.86 | 0.08 | Non-specific phospholipase C2; EC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, R.; Bai, H.; Li, B.; Liu, C.; Ying, Z.; Xiong, Z.; Wang, W. Combined Transcriptome and Lipidomic Analyses of Lipid Biosynthesis in Macadamia ternifolia Nuts. Life 2021, 11, 1431. https://doi.org/10.3390/life11121431
Shi R, Bai H, Li B, Liu C, Ying Z, Xiong Z, Wang W. Combined Transcriptome and Lipidomic Analyses of Lipid Biosynthesis in Macadamia ternifolia Nuts. Life. 2021; 11(12):1431. https://doi.org/10.3390/life11121431
Chicago/Turabian StyleShi, Rui, Haidong Bai, Biao Li, Can Liu, Zhiping Ying, Zhi Xiong, and Wenlin Wang. 2021. "Combined Transcriptome and Lipidomic Analyses of Lipid Biosynthesis in Macadamia ternifolia Nuts" Life 11, no. 12: 1431. https://doi.org/10.3390/life11121431
APA StyleShi, R., Bai, H., Li, B., Liu, C., Ying, Z., Xiong, Z., & Wang, W. (2021). Combined Transcriptome and Lipidomic Analyses of Lipid Biosynthesis in Macadamia ternifolia Nuts. Life, 11(12), 1431. https://doi.org/10.3390/life11121431