Brain Activity in Response to Visual Symmetry


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. The Importance of Symmetry
1.2. Models of Symmetry Processing in the Brain
2. Six Questions
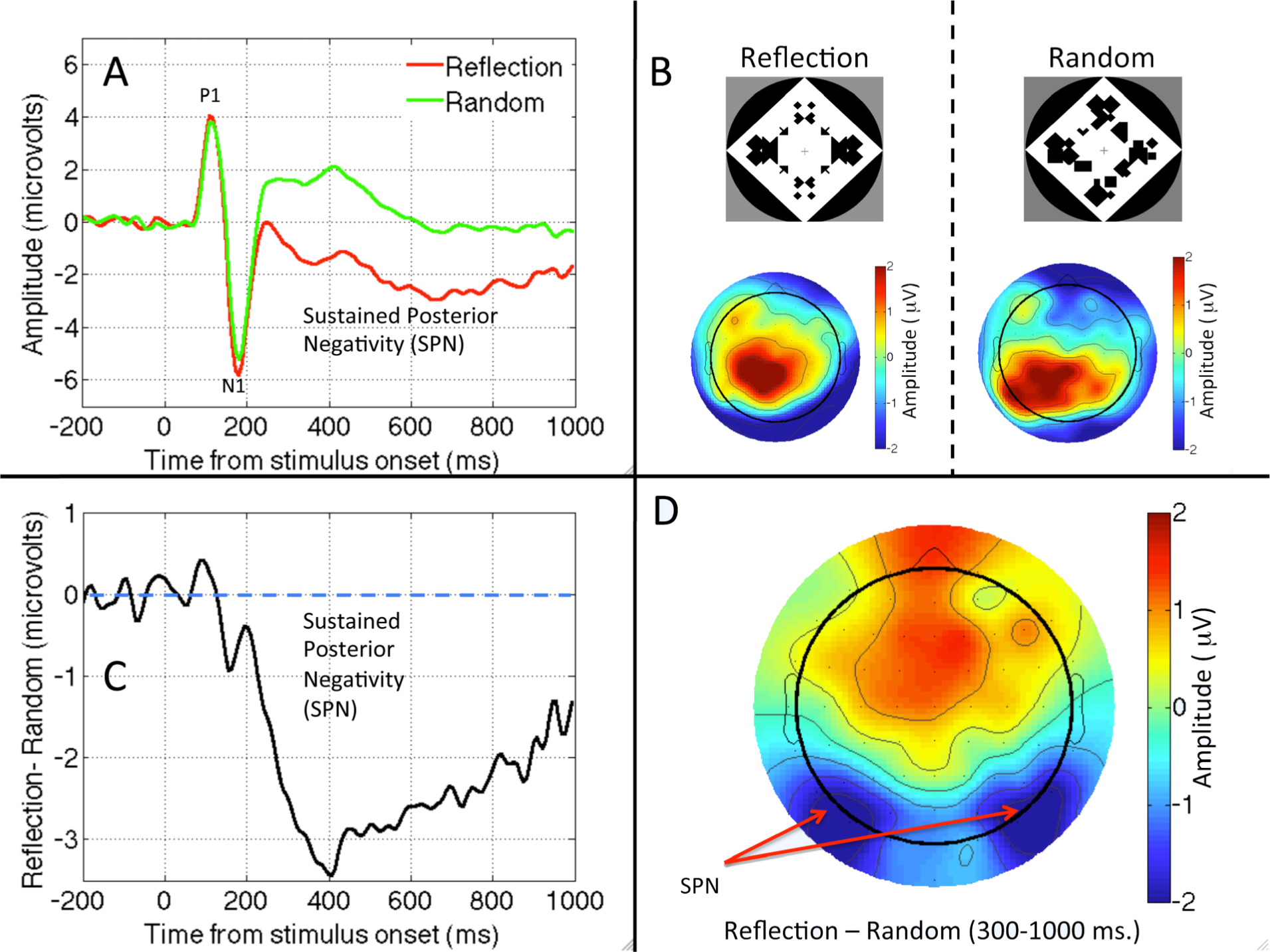
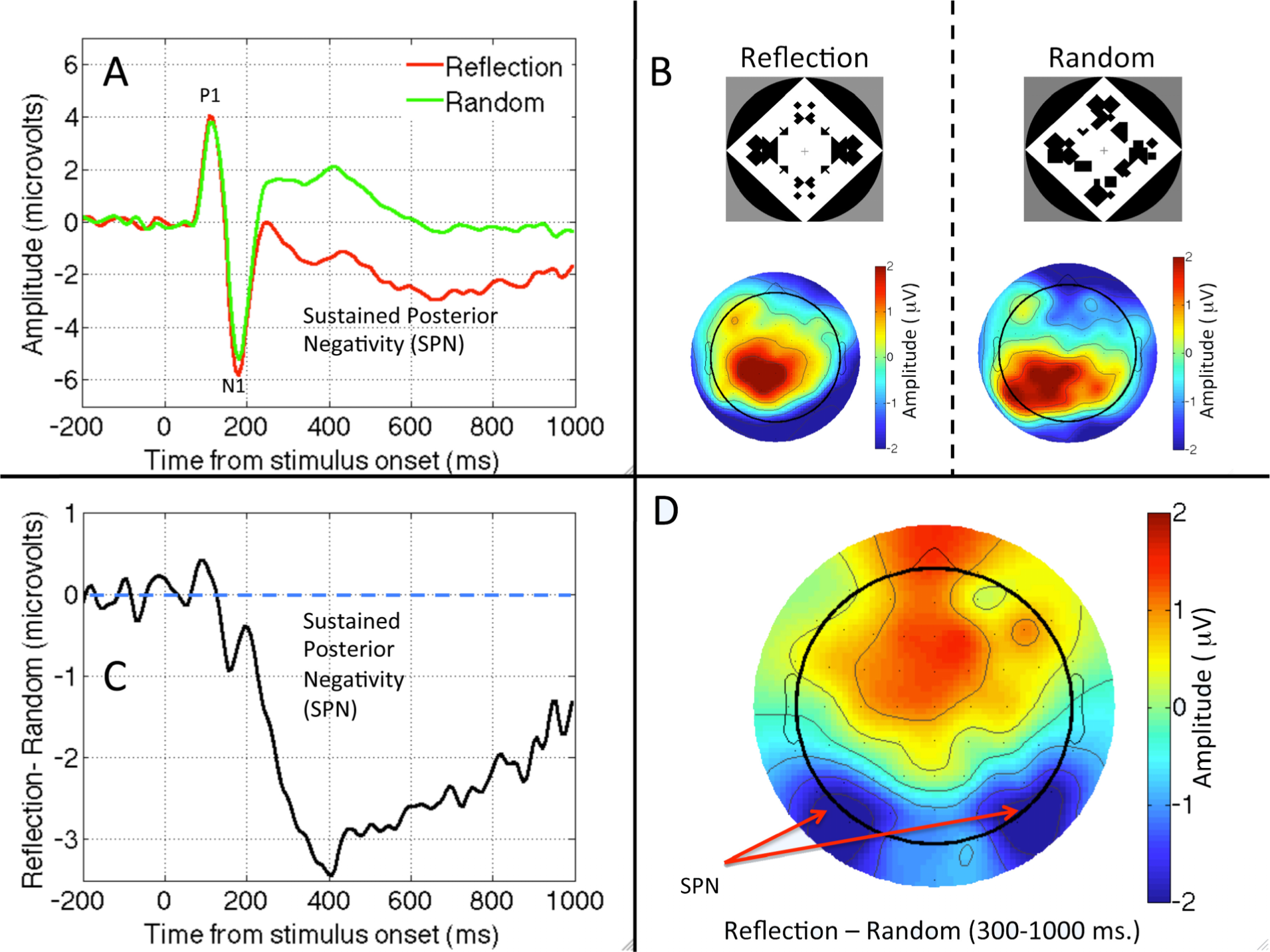
2.1. Question One: Is There an Automatic and Sustained Response to Symmetry in Visual Areas?
2.2. Question Two: Which Brain Areas Are Involved in Symmetry Perception?
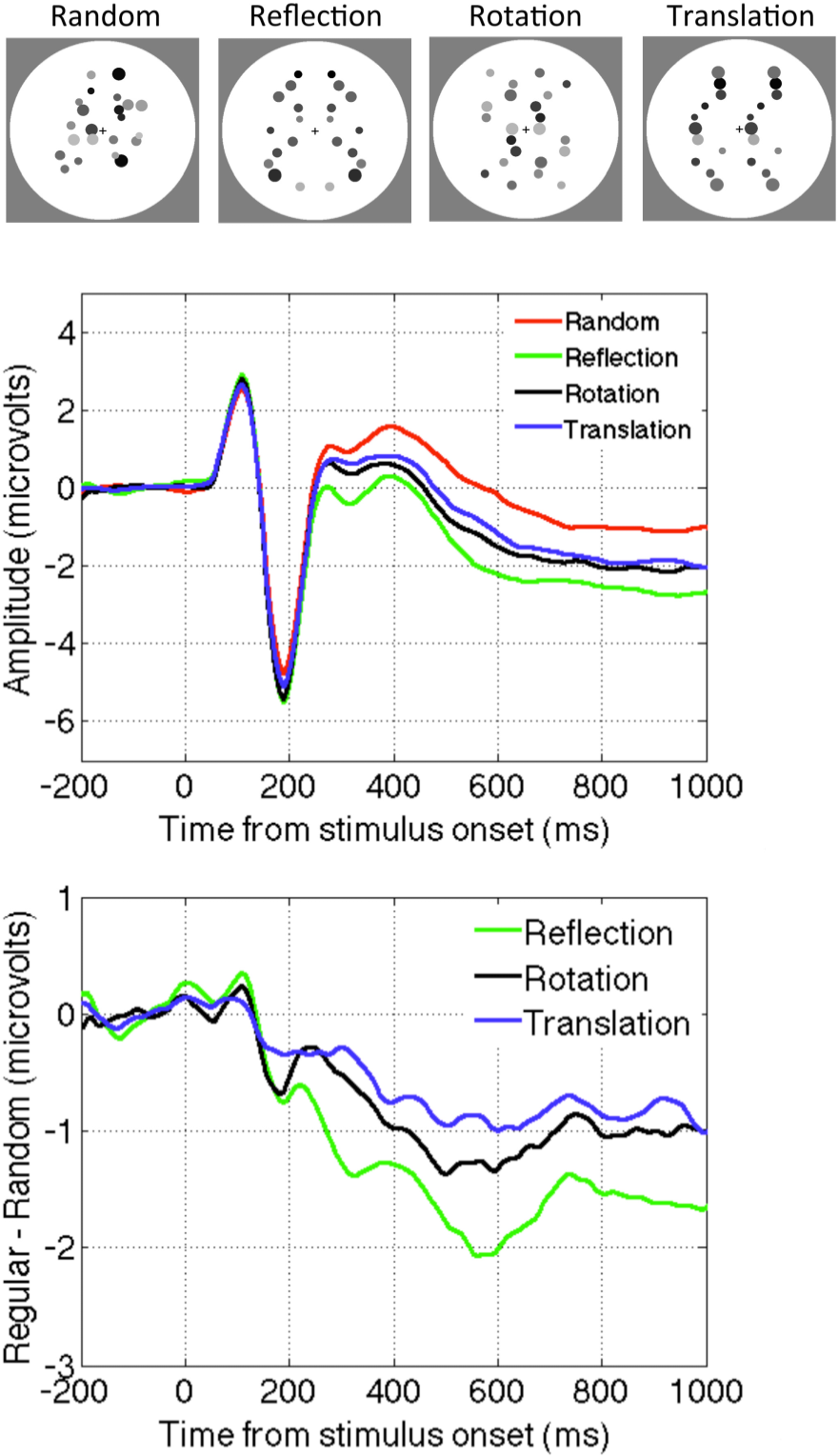
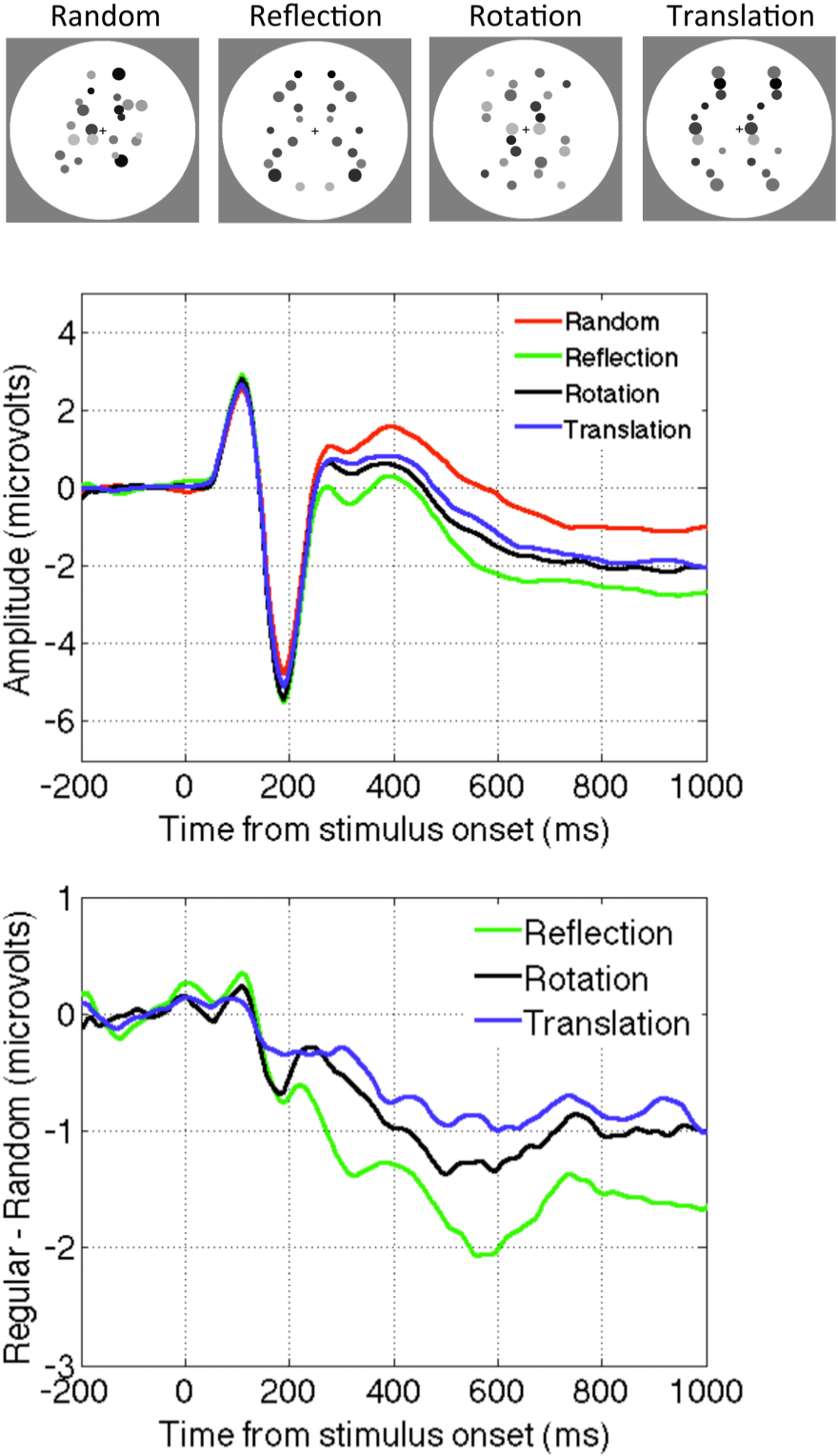
2.3. Question Three: Is Reflection Special?
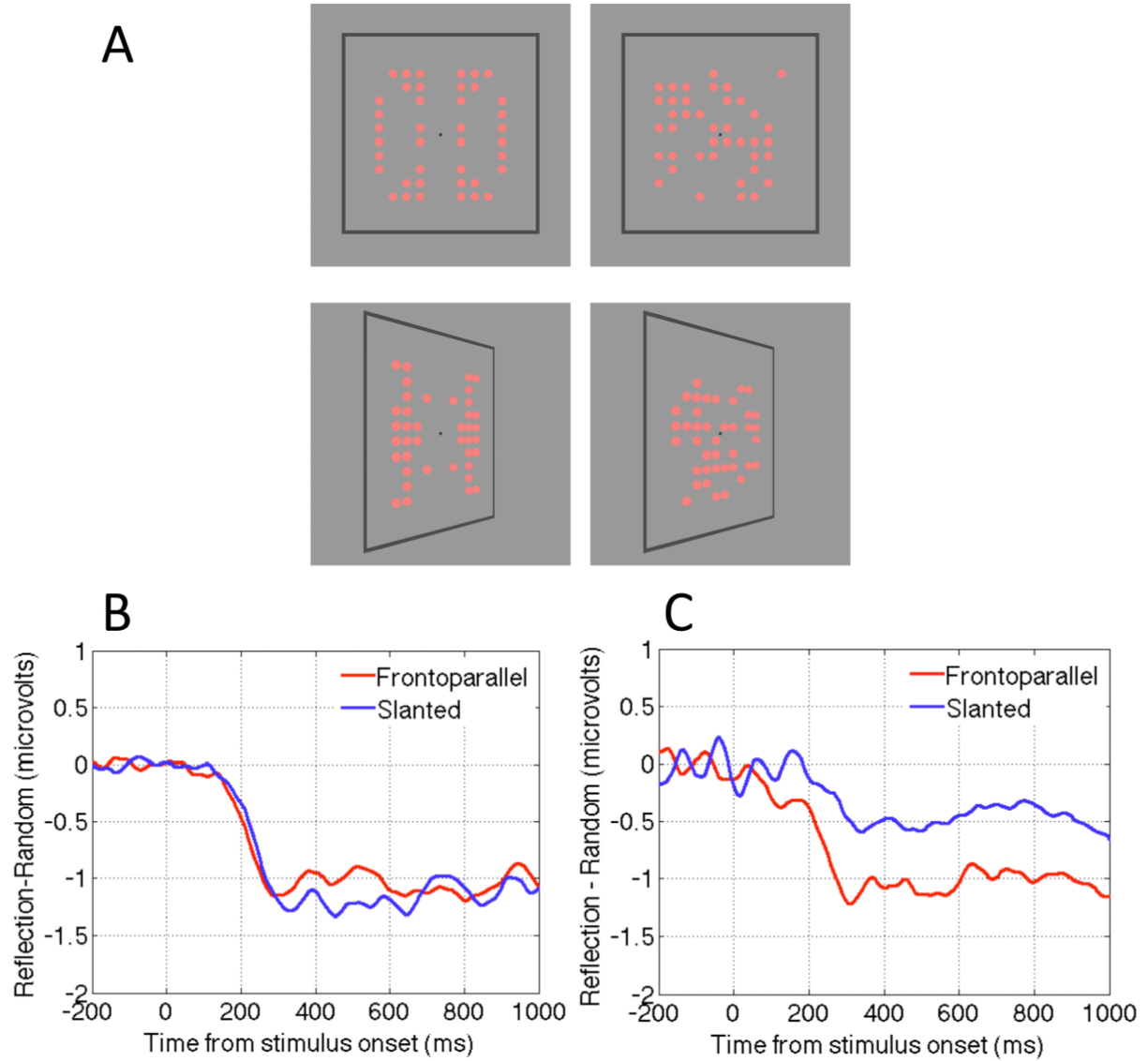
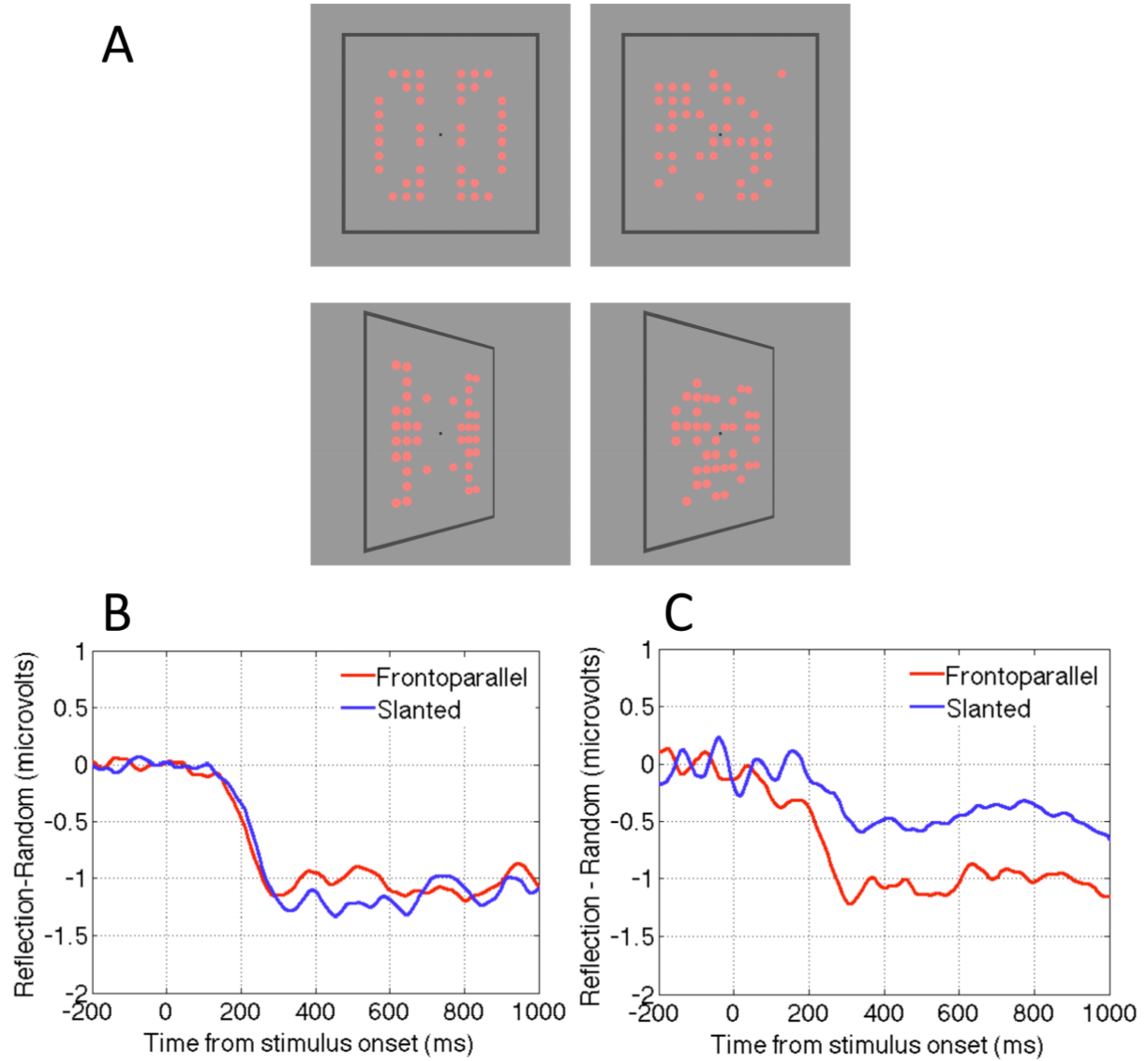
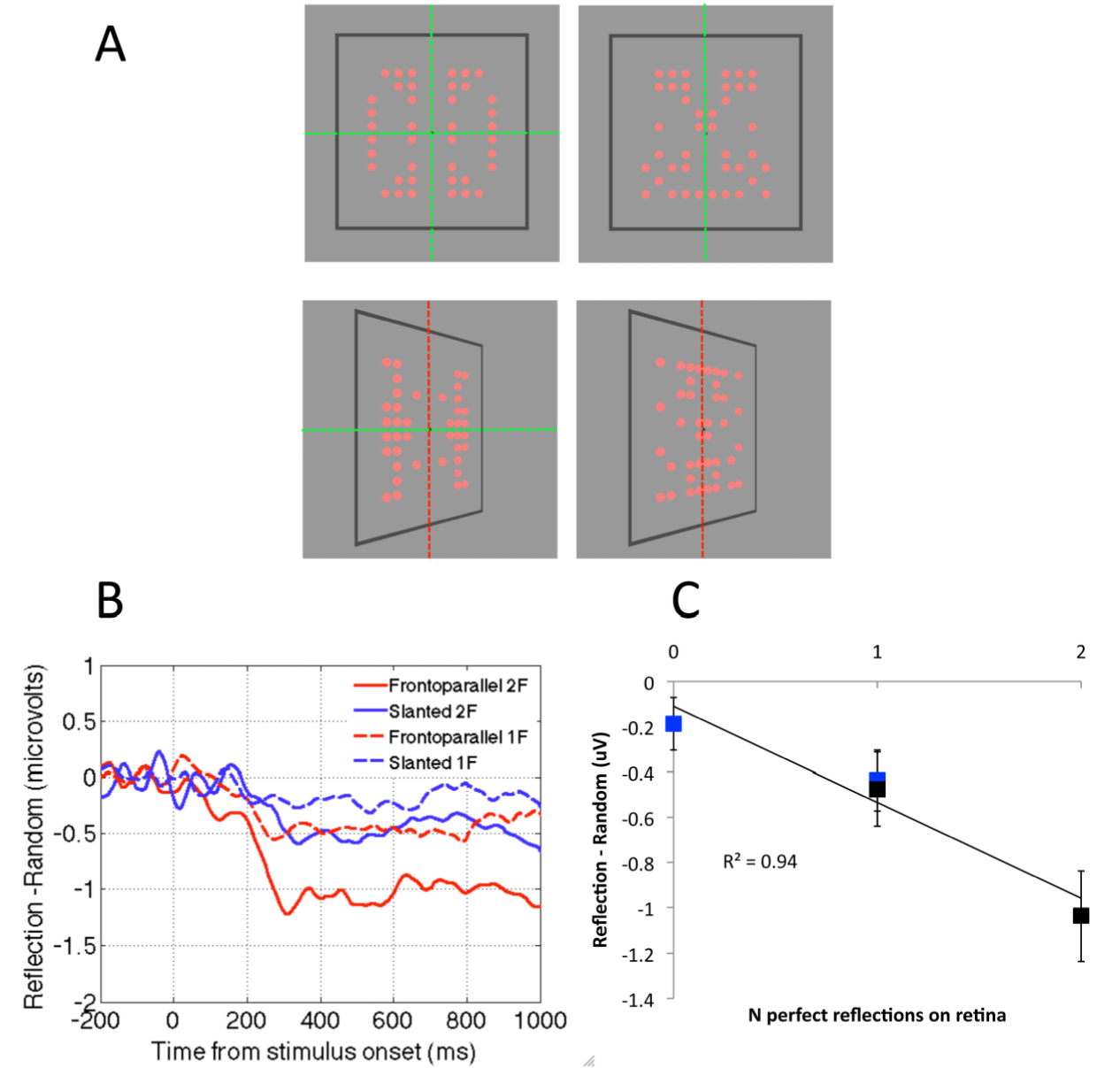
2.4. Question Four: Is the Neural Response to Symmetry View Invariant?
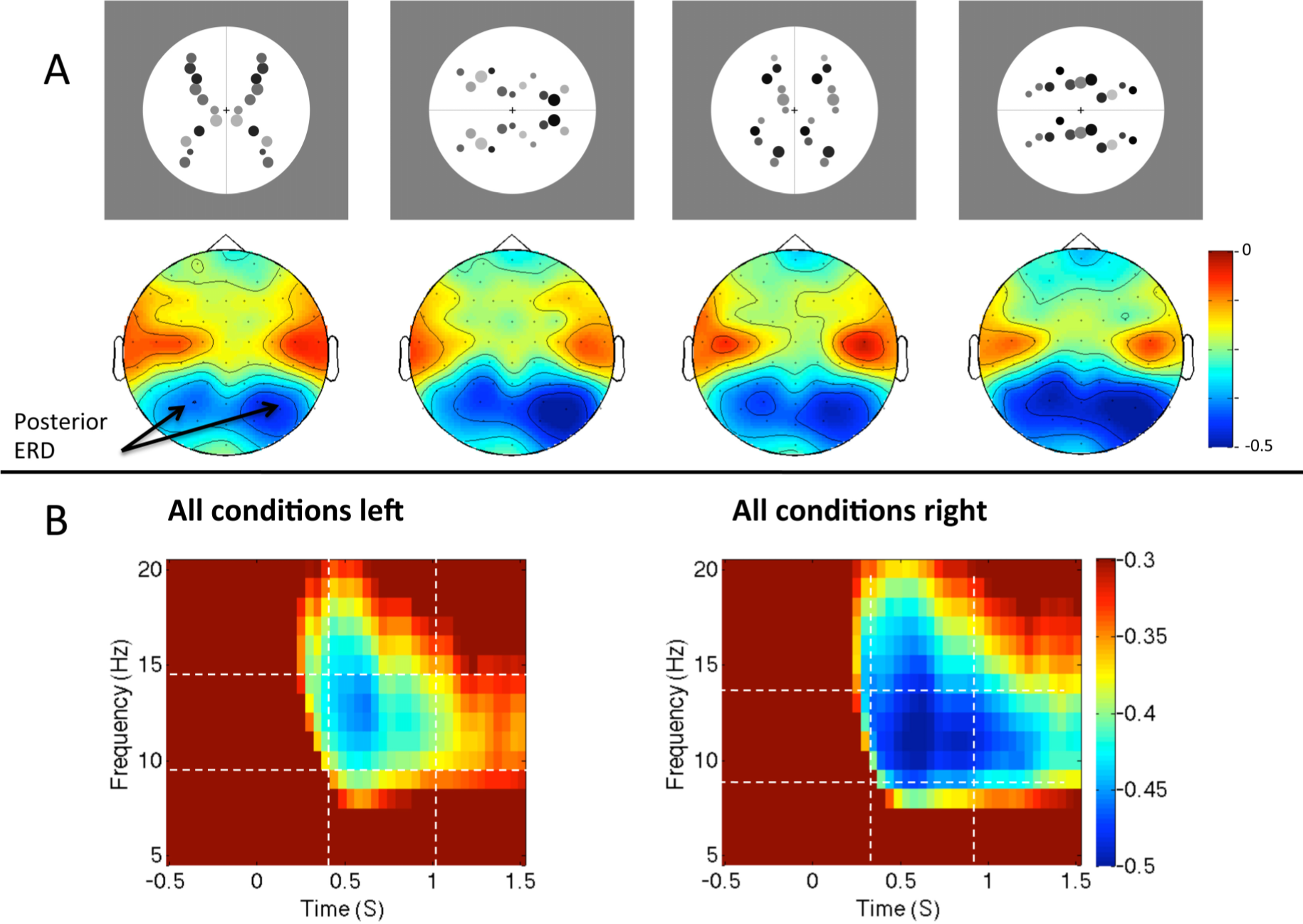
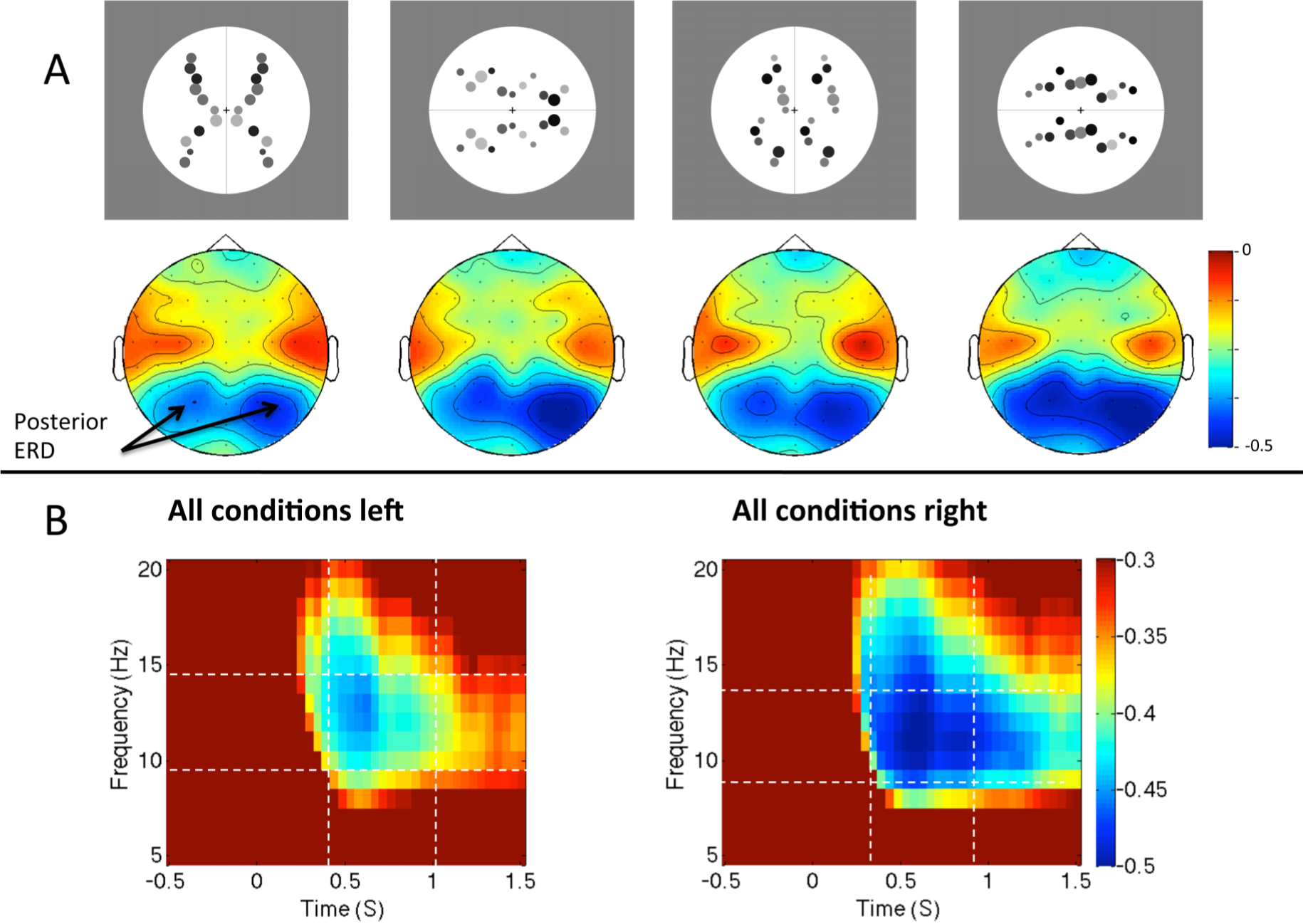
2.5. Question Five: How Are Brain Rhythms in the Two Hemispheres Altered During Symmetry Perception?
2.6. Question Six: Does Symmetry Processing Produce an Automatic Affective Response?
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Tyler, C.W. Empirical aspects of symmetry perception. Spat. Vis 1995, 9, 1–7. [Google Scholar]
- Wagemans, J. Detection of visual symmetries. Spat. Vis 1995, 9, 9–32. [Google Scholar]
- Wagemans, J. Characteristics and models of human symmetry detection. Trends Cogn. Sci 1997, 1, 346–352. [Google Scholar]
- Treder, M.S. Behind the looking glass: A review on human symmetry perception. Symmetry 2010, 2, 510–543. [Google Scholar]
- Van der Helm, P.A. Symmetry perception. In The Oxford Handbook of Perceptual Organization; Wagemans, J., Ed.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Mach, E. The analysis of sensations and the relation of the physical to the psychical; Dover Publications: New York, NY, USA, 1959. [Google Scholar]
- Bertamini, M. Sensitivity to reflection and translation is modulated by objectness. Perception 2010, 39, 27–40. [Google Scholar]
- Wenderoth, P. The salience of vertical symmetry. Perception 1994, 23, 221–236. [Google Scholar]
- Grammer, K.; Fink, B.; Møller, A.P.; Thornhill, R. Darwinian aesthetics: Sexual selection and the biology of beauty. Biol. Rev 2003, 78, 385–407. [Google Scholar]
- Rhodes, G.; Proffitt, F.; Grady, J.M.; Sumich, A. Facial symmetry and the perception of beauty. Psychon. Bull. Rev 1998, 5, 659–669. [Google Scholar]
- Bertamini, M.; Byrne, C.; Bennett, K.M. Attractiveness is influenced by the relationship between postures of the viewer and the viewed person. i-Perception 2013, 4, 170–179. [Google Scholar]
- Tovee, M.J.; Tasker, K.; Benson, P.J. Is symmetry a visual cue to attractiveness in the human female body? Evol. Hum. Behav 2000, 21, 191–200. [Google Scholar]
- Van Dongen, S. Associations between asymmetry and human attractiveness: Possible direct effects of asymmetry and signatures of publication bias. Ann. Hum. Biol 2011, 38, 317–323. [Google Scholar]
- Møller, A.; Thornhill, R. Bilateral symmetry and sexual selection: A meta-analysis. Am. Nat 1998, 151, 174–192. [Google Scholar]
- Wignall, A.E.; Heiling, A.M.; Cheng, K.; Herberstein, M.E. Flower symmetry preferences in honeybees and their crab spider predators. Ethology 2006, 112, 510–518. [Google Scholar]
- Swaddle, J.P.; Cuthill, I.C. Female zebra finches prefer males with symmetrical chest plumage. Proc. R. Soc. B Biol. Sci 1994, 258, 267–271. [Google Scholar]
- Rodriguez, I.; Gumbert, A.; de Ibarra, N.H.; Kunze, J.; Giurfa, M. Symmetry is in the eye of the “beholder”: Innate preference for bilateral symmetry in flower-naive bumblebees. Naturwissenschaften 2004, 91, 374–377. [Google Scholar]
- Delius, J.D.; Habers, G. Symmetry: Can pigeons conceptualize it? Behav. Biol 1978, 22, 336–342. [Google Scholar]
- Swaddle, J.P.; Ruff, D.A.; Page, L.C.; Frame, A.M.; Long, V.A. A test of receiver perceptual performance: European starlings’ ability to detect asymmetry in a naturalistic trait. Anim. Behav 2008, 76, 487–495. [Google Scholar]
- Tudor, M.S.; Morris, M.R. Experience plays a role in female preference for symmetry in the swordtail fish xiphophorus malinche. Ethology 2009, 115, 812–822. [Google Scholar]
- Clara, E.; Regolin, L.; Vallortigara, G. Preference for symmetry is experience dependent in newborn chicks (Gallus gallus). J. Exp. Psychol. Anim. Behav. Process 2007, 33, 12–20. [Google Scholar]
- Bornstein, M.H.; Ferdinandsen, K.; Gross, C.G. Perception of symmetry in infancy. Dev. Psychol 1981, 17, 82–86. [Google Scholar]
- Humphrey, G.K.; Humphrey, D.E. The role of structure in infant visual-pattern perception. Can. J. Psychol 1989, 43, 165–182. [Google Scholar]
- Machilsen, B.; Pauwels, M.; Wagemans, J. The role of vertical mirror symmetry in visual shape detection. J. Vis 2009, 9, 1–11. [Google Scholar]
- Li, Y.F.; Pizlo, Z.; Steinman, R.M. A computational model that recovers the 3D shape of an object from a single 2D retinal representation. Vis. Res 2009, 49, 979–991. [Google Scholar]
- Reber, R. Processing fluency, aesthetic pleasure, and culturally shared taste. In Aesthetic Science: Connecting Mind, Brain, and Experience; Shimamura, A.P., Palmer, S.E., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 223–249. [Google Scholar]
- Reber, R.; Winkielman, P.; Schwartz, N. Effects of perceptual fluency on affective judgments. Psychol. Sci 1998, 9, 45–48. [Google Scholar]
- Makin, A.D.J.; Pecchinenda, A.; Bertamini, M. Implicit affective evaluation of visual symmetry. Emotion 2012, 12, 1021–1030. [Google Scholar]
- Washburn, D.K.; Crowe, D.S. Symmetries of Culture; University of Washington Press: Seattle, WA, USA, 1988. [Google Scholar]
- Arnheim, R. Art and Visual Perception: A Psychology of the Creative Eye; University of California Press: Berkley, CA, USA, 1974. [Google Scholar]
- Ramachandran, V.S.; Hirstein, W. The science of art: A neurological theory of aesthetic experience. J. Conscious. Stud 1999, 6, 15–31. [Google Scholar]
- Chatterjee, A. Neuroaesthetics: A coming of age story. J. Cogn. Neurosci 2011, 23, 53–62. [Google Scholar]
- Leder, H.; Belke, B.; Oeberst, A.; Augustin, D. A model of aesthetic appreciation and aesthetic judgments. Br. J. Psychol 2004, 95, 489–508. [Google Scholar]
- Zeki, S. Art and the brain. J. Conscious. Stud 1999, 6, 76–96. [Google Scholar]
- Zeki, S. Neural concept formation and art: Dante, Michelangelo, Wagner. J. Conscious. Stud 2002, 9, 53–76. [Google Scholar]
- Herbert, A.M.; Humphrey, G.K. Bilateral symmetry detection: Testing a “callosal” hypothesis. Perception 1996, 25, 463–480. [Google Scholar]
- Dakin, S.C.; Hess, R.F. The spatial mechanisms mediating symmetry perception. Vis. Res 1997, 37, 2915–2930. [Google Scholar]
- Dakin, S.C.; Watt, R.J. Detection of bilateral symmetry using spatial filters. Spat. Vis 1994, 8, 393–413. [Google Scholar]
- Movshon, J.A.; Thompson, I.D.; Tolhurst, D.J. Spatial summation in the receptive fields of simple cells in the cat’s striate cortex. J. Physiol 1978, 283, 53–77. [Google Scholar]
- Wenderoth, P. The effects of the contrast polarity of dot-pair partners on the detection of bilateral symmetry. Perception 1996, 25, 757–771. [Google Scholar]
- Tyler, C.W.; Hardage, L. Mirror symmetry detection: Predominance of second-order pattern processing throughout the visual field. In Human Symmetry Perception and Its Computational Analysis; Tyler, C.W., Ed.; VSP; Utrecht, The Netherlands, 1996; pp. 157–172. [Google Scholar]
- Mancini, S.; Sally, S.L.; Gurnsey, R. Detection of symmetry and anti-symmetry. Vis. Res 2005, 45, 2145–2160. [Google Scholar]
- Poirier, F.J.A.M.; Wilson, H.R. A biologically plausible model of human shape symmetry perception. J. Vis 2010, 10, 1–16. [Google Scholar]
- Wagemans, J.; van Gool, L.; Swinnen, V.; van Horebeek, J. Higher-order structure in regularity detection. Vis. Res 1993, 33, 1067–1088. [Google Scholar]
- Makin, A.D.J.; Wilton, M.M.; Pecchinenda, A.; Bertamini, M. Symmetry perception and affective responses: A combined EEG/EMG study. Neuropsychologia 2012, 50, 3250–3261. [Google Scholar]
- Norcia, A.M.; Candy, T.R.; Pettet, M.W.; Vildavski, V.Y.; Tyler, C.W. Temporal dynamics of the human response to symmetry. J. Vis 2002, 2, 132–139. [Google Scholar]
- Jacobsen, T.; Höfel, L. Descriptive and evaluative judgment processes: Behavioral and electrophysiological indices of processing symmetry and aesthetics. Cogn. Affective Behav. Neurosci 2003, 3, 289–299. [Google Scholar]
- Höfel, L.; Jacobsen, T. Electrophysiological indices of processing aesthetics: Spontaneous or intentional processes? Int. J. Psychophysiol 2007, 65, 20–31. [Google Scholar]
- Höfel, L.; Jacobsen, T. Electrophysiological indices of processing symmetry and aesthetics: A result of judgment categorization or judgment report? J. Psychophysiol 2007, 21, 9–21. [Google Scholar]
- Makin, A.D.J.; Rampone, G.; Pecchinenda, A.; Bertamini, M. Electrophysiological responses to visuospatial regularity. Psychophysiology 2013, 50, 1045–1056. [Google Scholar]
- Rampone, G.; Makin, A.D.J.; Bertamini, M. Electrophysiological analysis of the affective congruence between pattern regularity and word valence. Neuropsychologia 2014, 58, 107–117. [Google Scholar]
- Royer, F.L. Detection of symmetry. J. Exp. Psychol. Hum. Percept. Perform 1981, 7, 1186–1210. [Google Scholar]
- Makin, A.D.J.; Rampone, G.; Wright, A.; Martinovic, J.; Bertamini, M. Visual symmetry in objects and gaps. J. Vis 2014, 14, 1–12. [Google Scholar]
- Bertamini, M.; Friedenberg, J.D.; Kubovy, M. Detection of symmetry and perceptual organization: The way a lock-and-key process works. Acta Psychol 1997, 95, 119–140. [Google Scholar]
- Beh, H.C.; Latimer, C.R. Symmetry detection and orientation perception: Electrocortical responses to stimuli with real and implicit axes of orientation. Aust. J. Psychol 1997, 49, 128–133. [Google Scholar]
- Tyler, C.W.; Hardage, L.; Miller, R.T. Multiple mechanisms for the detection of mirror symmetry. Spat. Vis 1995, 9, 79–100. [Google Scholar]
- Wright, D.; Makin, A.D.J.; Bertamini, M. Right-lateralized alpha desynchronization during symmetry discrimination: Hemispheric specialization or directed spatial attention? Psychophysiology 2015, in press. [Google Scholar]
- Makin, A.D.J.; Rampone, G.; Bertamini, M. Conditions for view invariance in the neural response to visual symmetry. Psychophysiology 2014. [Google Scholar] [CrossRef]
- Oka, S.; Victor, J.D.; Conte, M.M.; Yanagida, T. VEPs elicited by local correlations and global symmetry: Characteristics and interactions. Vis. Res 2007, 47, 2212–2222. [Google Scholar]
- Verleger, R.; Görgen, S.; Jaśkowski, P. An ERP indicator of processing relevant gestalts in masked priming. Psychophysiology 2005, 42, 677–690. [Google Scholar]
- De Wit, T.C.J.; Bauer, M.; Oostenveld, R.; Fries, P.; van Lier, R. Cortical responses to contextual influences in amodal completion. Neuroimage 2006, 32, 1815–1825. [Google Scholar]
- Gruber, T.; Müller, M.M. Oscillatory brain activity dissociates between associative stimulus content in a repetition priming task in the human EEG. Cereb. Cortex 2005, 15, 109–116. [Google Scholar]
- Martinovic, J.; Mordal, J.; Wuerger, S.M. Event-related potentials reveal an early advantage for luminance contours in the processing of objects. J. Vis 2011, 11, 1–15. [Google Scholar]
- Tyler, C.W.; Baseler, H.A.; Kontsevich, L.L.; Likova, L.T.; Wade, A.R.; Wandell, B.A. Predominantly extra-retinotopic cortical response to pattern symmetry. Neuroimage 2005, 24, 306–314. [Google Scholar]
- Sasaki, Y.; Vanduffel, W.; Knutsen, T.; Tyler, C.W.; Tootell, R. Symmetry activates extrastriate visual cortex in human and nonhuman primates. Proc. Natl. Acad. Sci. USA 2005, 102, 3159–3163. [Google Scholar]
- Chen, C.C.; Kao, K.L.C.; Tyler, C.W. Face configuration processing in the human brain: The role of symmetry. Cereb. Cortex 2007, 17, 1423–1432. [Google Scholar]
- Bona, S.; Herbert, A.; Toneatto, C.; Silvanto, J.; Cattaneo, Z. The causal role of the lateral occipital complex in visual mirror symmetry detection and grouping: An fMRI-guided TMS study. Cortex 2014, 51, 46–55. [Google Scholar]
- Cattaneo, Z.; Mattavelli, G.; Papagno, C.; Herbert, A.; Silvanto, J. The role of the human extrastriate visual cortex in mirror symmetry discrimination: A TMS-adaptation study. Brain Cogn 2011, 77, 120–127. [Google Scholar]
- Kourtzi, Z.; Kanwisher, N. Representation of perceived object shape by the human lateral occipital complex. Science 2001, 293, 1506–1509. [Google Scholar]
- Clark, A. Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behav. Brain Sci 2013, 36, 181–204. [Google Scholar]
- Rao, R.P.N.; Ballard, D.H. Predictive coding in the visual cortex: A functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci 1999, 2, 79–87. [Google Scholar]
- Joo, S.J.; Boynton, G.M.; Murray, S.O. Long-range, pattern-dependent contextual effects in early human visual cortex. Curr. Biol 2012, 22, 781–786. [Google Scholar]
- Hochstein, S.; Ahissar, M. View from the top: Hierarchies and reverse hierarchies in the visual system. Neuron 2002, 36, 791–804. [Google Scholar]
- Van der Zwan, R.; Leo, E.; Joung, W.; Latimer, C.; Wenderoth, P. Evidence that both area V1 and extrastriate visual cortex contribute to symmetry perception. Curr. Biol 1998, 8, 889–892. [Google Scholar]
- Palmer, S.E. On goodness, Gestalt, groups, and Garner: Local symmetry subgroups as a theory of figural goodness. In The Structure of Perception; Lockhead, G., Pomerantz, J., Eds.; American Psychological Association: Washington, DC, USA, 1991. [Google Scholar]
- Biederman, I.; Gerhardstein, P.C. Recognizing depth-rotated objects—Evidence and conditions for 3-dimensional viewpoint invariance. J. Exp. Psychol. Hum. Percept. Perform 1993, 19, 1162–1182. [Google Scholar]
- Foster, D.H.; Gilson, S.J. Recognizing novel three-dimensional objects by summing signals from parts and views. Proc. R. Soc. B Biol. Sci 2002, 269, 1939–1947. [Google Scholar]
- Szlyk, J.P.; Rock, I.; Fisher, C.B. Level of processing in the perception of symmetrical forms viewed from different angles. Spat. Vis 1995, 9, 139–150. [Google Scholar]
- Van der Vloed, G.; Csatho, A.; van der Helm, P.A. Symmetry and repetition in perspective. Acta Psychol 2005, 120, 74–92. [Google Scholar]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends Cogn. Sci 2005, 9, 474–480. [Google Scholar]
- Klimesch, W.; Sauseng, P.; Hanslmayr, S. EEG alpha oscillations: The inhibition-timing hypothesis. Brain Res. Rev 2007, 53, 63–88. [Google Scholar]
- Laufs, H.; Holt, J.L.; Elfont, R.; Krams, M.; Paul, J.S.; Krakow, K.; Kleinschmidt, A. Where the BOLD signal goes when alpha EEG leaves. Neuroimage 2006, 31, 1408–1418. [Google Scholar]
- Sauseng, P.; Klimesch, W.; Stadler, W.; Schabus, M.; Doppelmayr, M.; Hanslmayr, S.; Gruber, W.R.; Birbaumer, N. A shift of visual spatial attention is selectively associated with human EEG alpha activity. Eur. J. Neurosci 2005, 22, 2917–2926. [Google Scholar]
- Parsons, L.M. Superior parietal cortices and varieties of mental rotation. Trends Cogn. Sci 2003, 7, 515–517. [Google Scholar]
- Mesulam, M.M. Functional anatomy of attention and neglect: From neurons to networks. In The Cognitive and Neural Bases of Spatial Neglect; Karnath, H., Milner, D., Vallar, G., Eds.; Oxford University Press: New York, NY, USA, 2002; pp. 33–45. [Google Scholar]
- Nicholls, M.E.; Roberts, G.R. Can free-viewing perceptual asymmetries be explained by scanning, pre-motor or attentional biases? Cortex 2002, 38, 113–136. [Google Scholar]
- Jacobsen, T.; Schubotz, R.I.; Höfel, L.; Cramon, D.Y. Brain correlates of aesthetic judgment of beauty. NeuroImage 2006, 29, 276–285. [Google Scholar]
- Corballis, M.C.; Roldan, C.E. Perception of symmetrical and repeated patterns. Percept. Psychophys 1974, 16, 136–142. [Google Scholar]
- Verma, A.; van der Haegen, L.; Brysbaert, M. Symmetry detection in typically and atypically speech lateralized individuals: A visual half-field study. Neuropsychologia 2013, 51, 2611–2619. [Google Scholar]
- Brysbaert, M. Lateral preferences and visual-field asymmetries—Appearances may have been overstated. Cortex 1994, 30, 413–429. [Google Scholar]
- Wilkinson, D.T.; Halligan, P.W. The effects of stimulus symmetry on landmark judgments in left and right visual fields. Neuropsychologia 2002, 40, 1045–1058. [Google Scholar]
- Jensen, O.; Kaiser, J.; Lachaux, J.-P. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci 2007, 30, 317–324. [Google Scholar]
- Jacobsen, T.; Höfel, L. Aesthetic judgments of novel graphic patterns: Analyses of individual judgments. Percept. Mot. Skills 2002, 95, 755–766. [Google Scholar]
- Winkielman, P.; Cacioppo, J.T. Mind at ease puts a smile on the face: Psychophysiological evidence that processing facilitation elicits positive affect. J. Personal. Soc. Psychol 2001, 81, 989–1000. [Google Scholar]
- Rothermund, K.; Wentura, D. Underlying processes in the implicit association test: Dissociating salience from associations. J. Exp. Psychol. Gen 2004, 133, 139–165. [Google Scholar]
- Duckworth, K.L.; Bargh, J.A.; Garcia, M.; Chaiken, S. The automatic evaluation of novel stimuli. Psychol. Sci 2002, 13, 513–519. [Google Scholar]
- Nosek, B.A.; Greenwald, A.G.; Banaji, M.R. The implicit association test at age 7: A methodological and conceptual review. In Automatic Processes in Social Thinking and Behavior; Bargh, J.A., Ed.; Psychology Press: Philadelphia, PA, USA, 2007; pp. 265–292. [Google Scholar]
- Bertamini, M.; Makin, A.; Rampone, G. Implicit association of symmetry with positive valence, high arousal and simplicity. i-Perception 2013, 45, 317–327. [Google Scholar]
- Bertamini, M.; Makin, A.D.J.; Pecchinenda, A. Testing Whether and When Abstract Symmetric Patterns Produce Affective Responses. PLoS One 2013, 87, e68403. [Google Scholar] [CrossRef]
- Pecchinenda, A.; Bertamini, M.; Makin, A.D.J.; Ruta, N. The pleasantness of visual symmetry: Always, never or sometimes. PLoS One 2014, 9, e92685. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertamini, M.; Makin, A.D.J. Brain Activity in Response to Visual Symmetry. Symmetry 2014, 6, 975-996. https://doi.org/10.3390/sym6040975
Bertamini M, Makin ADJ. Brain Activity in Response to Visual Symmetry. Symmetry. 2014; 6(4):975-996. https://doi.org/10.3390/sym6040975
Chicago/Turabian StyleBertamini, Marco, and Alexis D.J. Makin. 2014. "Brain Activity in Response to Visual Symmetry" Symmetry 6, no. 4: 975-996. https://doi.org/10.3390/sym6040975
APA StyleBertamini, M., & Makin, A. D. J. (2014). Brain Activity in Response to Visual Symmetry. Symmetry, 6(4), 975-996. https://doi.org/10.3390/sym6040975