Cerebral Sulcal Asymmetry in Macaque Monkeys


Abstract

{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Asymmetrical Development of Cerebral Sulci
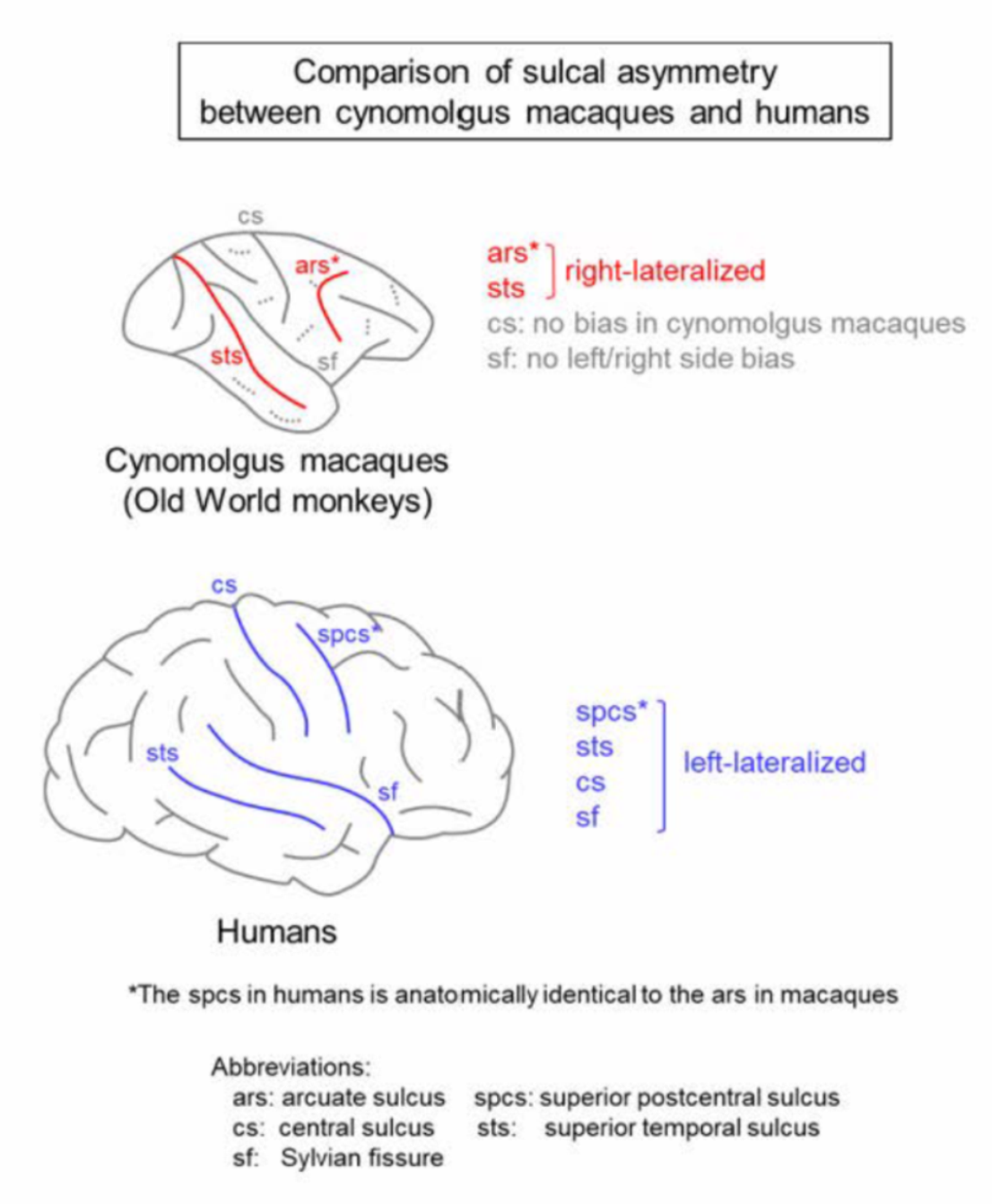
3. Sulcal Asymmetry in Macaques
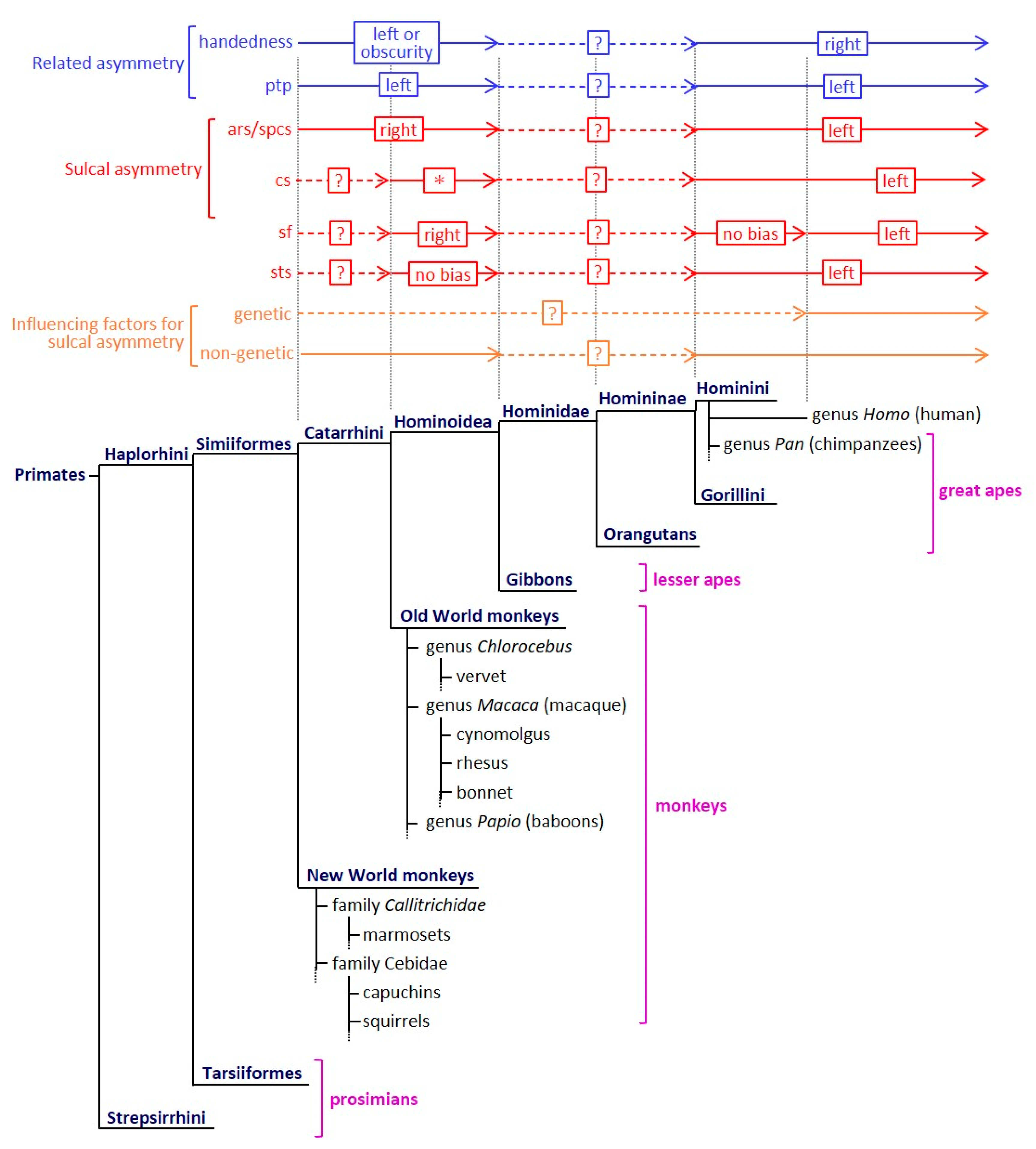
4. Comparison to Other Nonhuman Primates
5. Possible Mediators for Evaluating Sulcal Asymmetry
6. Conclusions
Funding
Conflicts of Interest
References
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains-The Biology and Behaviour of Brain Asymmetries; Cambridge Univ. Press: New York, NY, USA, 2013. [Google Scholar]
- Chi, J.G.; Dooling, E.C.; Gilles, F.H. Gyral development of the human brain. Ann. Neurol. 1977, 1, 86–93. [Google Scholar] [CrossRef]
- Pillay, P.; Manger, P.R. Order-specific quantitative patterns of cortical gyrification. Eur. J. Neurosci. 2007, 25, 2705–2712. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Fukunishi, K.; Kashima, M.; Imai, N.; Saito, S.; Sakata-Haga, H.; Aoki, I.; Fukui, Y. Neuroanatomic and MRI references for normal development of cerebral sulci of laboratory primate, cynomolgus monkeys (Macaca fascicularis). Congenit. Anom. (Kyoto) 2012, 52, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Hikishima, K.; Murayama, A.Y.; Okano, H.J.; Sasaki, E.; Okano, H. Fetal sulcation and gyrification in common marmosets (Callithrix jacchus) obtained by ex vivo magnetic resonance imaging. Neuroscience 2014, 257, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, I.; Hernández-Martí, M.; Bernet, E.; Galofré, E. Formation and growth of the cerebral convolutions I. Postnatal development of the median-suprasylvian gyrus and adjoining sulci in the cat. J. Anat. 1988, 160, 89–100. [Google Scholar]
- Wosinski, M.; Schleicher, A.; Zilles, K. Quantitative analysis of gyrification of cerebral cortex in dogs. Neurobiology 1996, 4, 441–468. [Google Scholar]
- Sawada, K.; Watanabe, M. Development of cerebral sulci and gyri in ferrets (Mustela putorius). Congenit. Anom. (Kyoto) 2012, 52, 168–175. [Google Scholar] [CrossRef]
- Sawada, K.; Aoki, I. Biphasic aspect of sexually dimorphic ontogenetic trajectory of gyrification in the ferret cerebral cortex. Neuroscience 2017, 364, 71–81. [Google Scholar] [CrossRef]
- Louw, G.J. The development of the sulci and gyri of the bovine cerebral hemispheres. Anat. Histol. Embryol. 1989, 18, 246–264. [Google Scholar] [CrossRef]
- Marino, L.; Connor, R.C.; Fordyce, R.E.; Herman, L.M.; Hof, P.R.; Lefebvre, L.; Lusseau, D.; McCowan, B.; Nimchinsky, E.A.; Pack, A.A.; et al. Cetaceans have complex brains for complex cognition. PLoS Biol. 2007, 5, e139. [Google Scholar] [CrossRef]
- Mortensen, H.S.; Pakkenberg, B.; Dam, M.; Dietz, R.; Sonne, C.; Mikkelsen, B.; Eriksen, N. Quantitative relationships in delphinid neocortex. Front. Neuroanat. 2014, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Kochounov, P.; Castro, C.; Davis, D.; Dudley, D.; Brewer, J.; Zhang, Y.; Kroenke, C.D.; Purdy, D.; Fox, P.T.; Simerly, C.; et al. Mapping primary gyrogenesis during fetal development in primate brains: High-resolution in utero structural MRI of fetal brain development in pregnant baboons. Front. Neurosci. 2010, 4, 20. [Google Scholar]
- Kochounov, P.; Glahn, D.C.; Fox, P.; Lancaster, J.L.; Saleem, K.; Shelledy, W.; Zilles, K.; Thompson, P.M.; Coulon, O.; Mangin, J.F.; et al. Genetics of primary cerebral gyrification: Heritability of length, depth and area of primary sulci in an extended pedigree of baboons. Neuroimage 2010, 53, 1126–1134. [Google Scholar] [CrossRef]
- Clark, G.M.; Mackay, C.E.; Davidson, M.E.; Iversen, S.D.; Collinson, S.L.; James, A.C.; Roberts, N.; Crow, T.J. Paracingulate sulcus asymmetry; sex difference, correlation with semantic fluency and change over time in adolescent onset psychosis. Psychiatry Res. 2010, 184, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Fukunishi, K.; Kashima, M.; Imai, N.; Saito, S.; Aoki, I.; Fukui, Y. Regional difference in sulcal infolding progression correlated with cerebral cortical expansion in cynomolgus monkey fetuses. Congenit. Anom. (Kyoto) 2017, 57, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Kochunov, P.; Zilles, K.; Shelledy, W.; Lancaster, J.; Thompson, P.; Duggirala, R.; Blangero, J.; Fox, P.T.; Glahn, D.C. On the genetic architecture of cortical folding and brain volume in primates. Neuroimage 2010, 53, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Sun, X.Z.; Fukunishi, K.; Kashima, M.; Satio, S.; Sakata-Haga, H.; Tokado, H.; Aoki, I.; Fukui, Y. Ontogenetic pattern of gyrification in fetuses of cynomolgus monkeys. Neuroscience 2010, 167, 735–740. [Google Scholar] [CrossRef]
- Liu, T.; Wen, W.; Zhu, W.; Trollor, J.; Reppermund, S.; Crawford, J.; Jin, J.S.; Luo, S.; Brodaty, H.; Sachdev, P. The effects of age and sex on cortical sulci in the elderly. Neuroimage 2010, 51, 19–27. [Google Scholar] [CrossRef]
- Imai, N.; Sawada, K.; Fukunishi, K.; Sakata-Haga, H.; Fukui, Y. Sexual dimorphism of sulcal length asymmetry in cerebrum of adult cynomolgus monkeys (Macaca fascicularis). Congenit. Anom. (Kyoto) 2011, 51, 161–166. [Google Scholar] [CrossRef]
- Foundas, A.L.; Hong, K.; Leonard, C.M.; Heilman, K.M. Hand preference and magnetic resonance imaging asymmetries of the central sulcus. Neuropsychiatry Neuropsychol. Behav. Neurol. 1998, 11, 65–71. [Google Scholar]
- Klöppel, S.; Mangin, J.F.; Vongerichten, A.; Frackowiak, R.S.; Siebner, H.R. Nurture versus nature: Long-term impact of forced right-handedness on structure of pericentral cortex and basal ganglia. J. Neurosci. 2010, 30, 3271–3275. [Google Scholar] [CrossRef] [PubMed]
- Fornito, A.; Wood, S.J.; Whittle, S.; Fuller, J.; Adamson, C.; Saling, M.M.; Velakoulis, D.; Pantelis, C.; Yücel, M. Variability of the paracingulate sulcus and morphometry of the medial frontal cortex: Associations with cortical thickness, surface area, volume, and sulcal depth. Hum. Brain Mapp. 2008, 29, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Amiez, C.; Wilson, C.R.E.; Procyk, E. Variations of cingulate sulcal organization and link with cognitive performance. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bonte, M.; Frost, M.A.; Rutten, S.; Ley, A.; Formisano, E.; Goebel, R. Development from childhood to adulthood increases morphological and functional inter-individual variability in the right superior temporal cortex. Neuroimage 2013, 83, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Bodin, C.; Takerkart, S.; Belin, P.; Coulon, O. Anatomo-functional correspondence in the superior temporal sulcus. Brain Struct. Funct. 2018, 223, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Corballis, M.C. The Lopsided Brain: Evolution of the Generative Mind; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Cantalupo, C.; Pilcher, D.L.; Hopkins, W.D. Are planum temporale and sylvian fissure asymmetries directly related? A MRI study in great apes. Neuropsychologia 2003, 41, 1975–1981. [Google Scholar] [CrossRef]
- Bogart, S.L.; Mangin, J.F.; Schapiro, S.J.; Reamer, L.; Bennett, A.J.; Pierre, P.J.; Hopkins, W.D. Cortical sulci asymmetries in chimpanzees and macaques: A new look at an old idea. Neuroimage 2012, 61, 533–541. [Google Scholar] [CrossRef]
- Bonan, I.; Argenti, A.M.; Duyme, M.; Hasboun, D.; Dorion, A.; Marsault, C.; Zouaoui, A. Magnetic resonance imaging of cerebral central sulci: A study of monozygotic twins. Acta Genet. Med. Gemellol. (Roma) 1998, 47, 89–100. [Google Scholar] [CrossRef]
- Kasprian, K.; Langs, G.; Brugger, P.C.; Bittner, M.; Weber, M.; Arantes, M.; Prayer, D. The prenatal origin of hemispheric asymmetry: An in utero neuroimaging study. Cereb. Cortex 2011, 21, 1076–1083. [Google Scholar] [CrossRef]
- Sawada, K.; Fukunishi, K.; Kashima, M.; Saito, S.; Sakata-Haga, H.; Aoki, I.; Fukui, Y. Fetal gyrification in cynomolgus monkeys: A concept of developmental stages of gyrification. Anat. Rec. (Hoboken) 2012, 295, 1065–1074. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sawada, K.; Fukunishi, K.; Imai, N.; Sakata-Haga, H.; Fukui, Y. Postnatal change in sulcal length asymmetry in cerebrum of cynomolgus monkeys (Macaca fascicularis). Anat. Rec. (Hoboken) 2014, 297, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K.; Palomero-Gallagher, N.; Amunts, K. Development of cortical folding during evolution and ontogeny. Trends Neurosci. 2013, 36, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Lonsdorf, E.V.; Hopkins, W.D. Wild chimpanzees show population level handedness for tool use. Proc. Natl. Acad. Sci. USA 2005, 102, 12634–12638. [Google Scholar] [CrossRef] [PubMed]
- Corp, N.; Byrne, R.W. Sex difference in chimpanzee handedness. Am. J. Phy. Anthropol. 2004, 123, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Chatagny, P.; Badoud, S.; Kaeser, M.; Gindrat, A.D.; Savidan, J.; Fregosi, M.; Moret, V.; Roulin, C.; Schmidlin, E.; Rouiller, E.M. Distinction between hand dominance and hand preference in primates: A behavioral investigation of manual dexterity in nonhuman primates (macaques) and human subjects. Brain Behav. 2013, 3, 575–595. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.; Hopkins, W.D. Exploring the relationship between cerebellar asymmetry and handedness in chimpanzees (Pan troglodytes) and capuchins (Cebus apella). Neuropsychologia 2007, 45, 2333–2339. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.A.; Thompson, C.R. Hand preference for tool-use in capuchin monkeys (Cebus apella) is associated with asymmetry of the primary motor cortex. Am. J. Primatol. 2013, 75, 435–440. [Google Scholar] [CrossRef]
- Margiotoudi, K.; Marie, D.; Claidière, N.; Coulon, O.; Roth, M.; Nazarian, B.; Lacoste, R.; Hopkins, W.D.; Molesti, S.; Fresnais, P.; et al. Handedness in monkeys reflects hemispheric specialization within the central sulcus. An in vivo MRI study in right- and left-handed olive baboons. Cortex 2019, 118, 203–211. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Nir, T. Planum temporale surface area and grey matter asymmetries in chimpanzees (Pan troglodytes): The effect of handedness and comparison within findings in humans. Behav. Brain Res. 2010, 208, 436–443. [Google Scholar] [CrossRef]
- Gannon, P.J.; Holloway, R.L.; Broadfield, D.C.; Braum, A.R. Asymmetry of chimpanzee planum temporale: Humanlike pattern of Wernicke’s brain language area homolog. Science 1998, 279, 220–222. [Google Scholar] [CrossRef]
- Marie, D.; Roth, M.; Lacoste, R.; Nazarian, B.; Bertello, A.; Anton, J.L.; Hopkins, W.D.; Margiotoudi, K.; Love, S.A.; Meguerditchian, A. Left brain asymmetry of the planum temporale in a nonhominid primate: Redefining the origin of brain specialization for language. Cereb. Cortex 2018, 28, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- Lyn, H.L.; Pierre, P.; Bennett, A.J.; Fears, S.C.; Woods, R.P.; Hopkins, W.D. Planum temporale grey matter asymmetries in chimpanzees (Pan troglodytes), vervet (Chlorocebus aethiops sabaeus), rhesus (Macaca mulatta) and bonnet (Macaca radiata) monkeys. Neuropsychologia 2011, 49, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, W.D. Neuroanatomical asymmetries and handedness in chimpanzees (Pan troglodytes): A case for continuity in the evolution of hemispheric specialization. Ann. N. Y. Acad. Sci. 2013, 1288, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K.; Armstrong, E.; Schleicher, A.; Kretschmann, H.J. The human pattern of gyrification in the cerebral cortex. Anat. Embryol. 1988, 179, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Rosch, R.E.; Ronan, L.; Cherkas, L.; Gurd, J.M. Cerebellar asymmetry in a pair of monozygotic handedness-discordant twins. J. Anat. 2010, 217, 38–47. [Google Scholar] [CrossRef]
- Kelava, I.; Lewitus, E.; Huttner, W.B. The aecondary loss of gyrencephaly as an example of evolutionary phenotypical reversal. Front. Neuroanat. 2013, 7, 16. [Google Scholar] [CrossRef]
- Yeni-Komshian, G.H.; Benson, D.A. Anatomical study of cerebral asymmetry in the temporal lobe of humans, chimpanzees, and rhesus monkeys. Science 1976, 192, 387–389. [Google Scholar] [CrossRef]
- Falk, D.; Cheverud, J.; Vannier, M.W.; Conroy, G.C. Advanced computer graphics technology reveals cortical asymmetry in endocasts of rhesus monkeys. Folia Primatol. 1986, 4, 98–103. [Google Scholar] [CrossRef]
- Falk, D.; Hildebolt, C.; Cheverud, J.; Vannier, M.; Helmkamp, R.C.; Konigsberg, L. Cortical asymmetries in frontal lobes of rhesus monkeys (Macaca mulatta). Brain Res. 1990, 512, 40–45. [Google Scholar] [CrossRef]
- Heilbroner, P.L.; Holloway, R.L. Anatomical brain asymmetries in New World and Old World monkeys: Stages of temporal lobe development in primate evolution. Am. J. Phys. Anthropol. 1988, 76, 39–48. [Google Scholar] [CrossRef]
- Heilbroner, P.L.; Holloway, R.L. Anatomical brain asymmetry in monkeys: Frontal, temporoparietal, and limbic cortex in Macaca. Am. J. Phys. Anthropol. 1989, 80, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Goulas, A.; Stiers, P.; Hutchison, R.M.; Everling, S.; Petrides, M.; Margulies, D.S. Intrinsic functional architecture of the macaque dorsal and ventral lateral frontal cortex. J. Neurophysiol. 2017, 117, 1084–1099. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, E.; Schleicher, A.; Omran, H.; Curtis, M.; Zilles, K. The ontogeny of human gyrification. Cereb. Cortex 1995, 5, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Sholl, S.A.; Kim, K.L. Androgen receptors are differentially distributed between right and left cerebral hemispheres of the fetal male rhesus monkey. Brain Res. 1990, 516, 122–126. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Pilcher, D.L.; MacGregor, L. Sylvian fissure asymmetries in nonhuman primates revisited: A comparative MRI study. Brain Behav. Evol. 2000, 56, 293–299. [Google Scholar] [CrossRef]
- Li, L.; Preuss, T.M.; Rilling, J.K.; Hopkins, W.D.; Glasser, M.F.; Kumar, B.; Nana, R.; Zhang, X.; Hu, X. Chimpanzee (Pan troglodytes) precentral corticospinal system asymmetry and handedness: A diffusion magnetic resonance imaging study. PLoS ONE 2010, 21, e12886. [Google Scholar] [CrossRef]
- Humle, T.; Matsuzawa, T. Laterality in hand use across four tool use behaviors among the wild chimpanzees of Bossou, Guinea, West Africa. Am. J. Primatol. 2009, 71, 40–48. [Google Scholar] [CrossRef]
- Sawada, K.; Horiuchi-Hirose, M.; Saito, S.; Aoki, I. Male prevalent enhancement of leftward asymmetric development of the cerebellar cortex in ferrets (Mustela putorius). Laterality 2015, 20, 723–737. [Google Scholar] [CrossRef]
- Sawada, K.; Aoki, I. Age-dependent sexually-dimorphic asymmetric development of the ferret cerebellar cortex. Symmetry 2017, 9, 40. [Google Scholar] [CrossRef]
- Sawada, K.; Kamiya, S.; Aoki, I. Asymmetry of cerebellar lobular development in ferrets. Symmetry 2020, 12, 735. [Google Scholar] [CrossRef]
- Koyun, N.; Aydinlioğlu, A.; Aslan, K. A morphometric study on dog cerebellum. Neurol. Res. 2011, 33, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Buckner, R.L.; Liu, H. Cerebellar asymmetry and its relation to cerebral asymmetry estimated by intrinsic functional connectivity. J. Neurophysiol. 2013, 109, 46–57. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawada, K. Cerebral Sulcal Asymmetry in Macaque Monkeys. Symmetry 2020, 12, 1509. https://doi.org/10.3390/sym12091509
Sawada K. Cerebral Sulcal Asymmetry in Macaque Monkeys. Symmetry. 2020; 12(9):1509. https://doi.org/10.3390/sym12091509
Chicago/Turabian StyleSawada, Kazuhiko. 2020. "Cerebral Sulcal Asymmetry in Macaque Monkeys" Symmetry 12, no. 9: 1509. https://doi.org/10.3390/sym12091509
APA StyleSawada, K. (2020). Cerebral Sulcal Asymmetry in Macaque Monkeys. Symmetry, 12(9), 1509. https://doi.org/10.3390/sym12091509