A Supersymmetry and Quantum Cryptosystem with Path Integral Approach in Biology




Abstract
1. Introduction
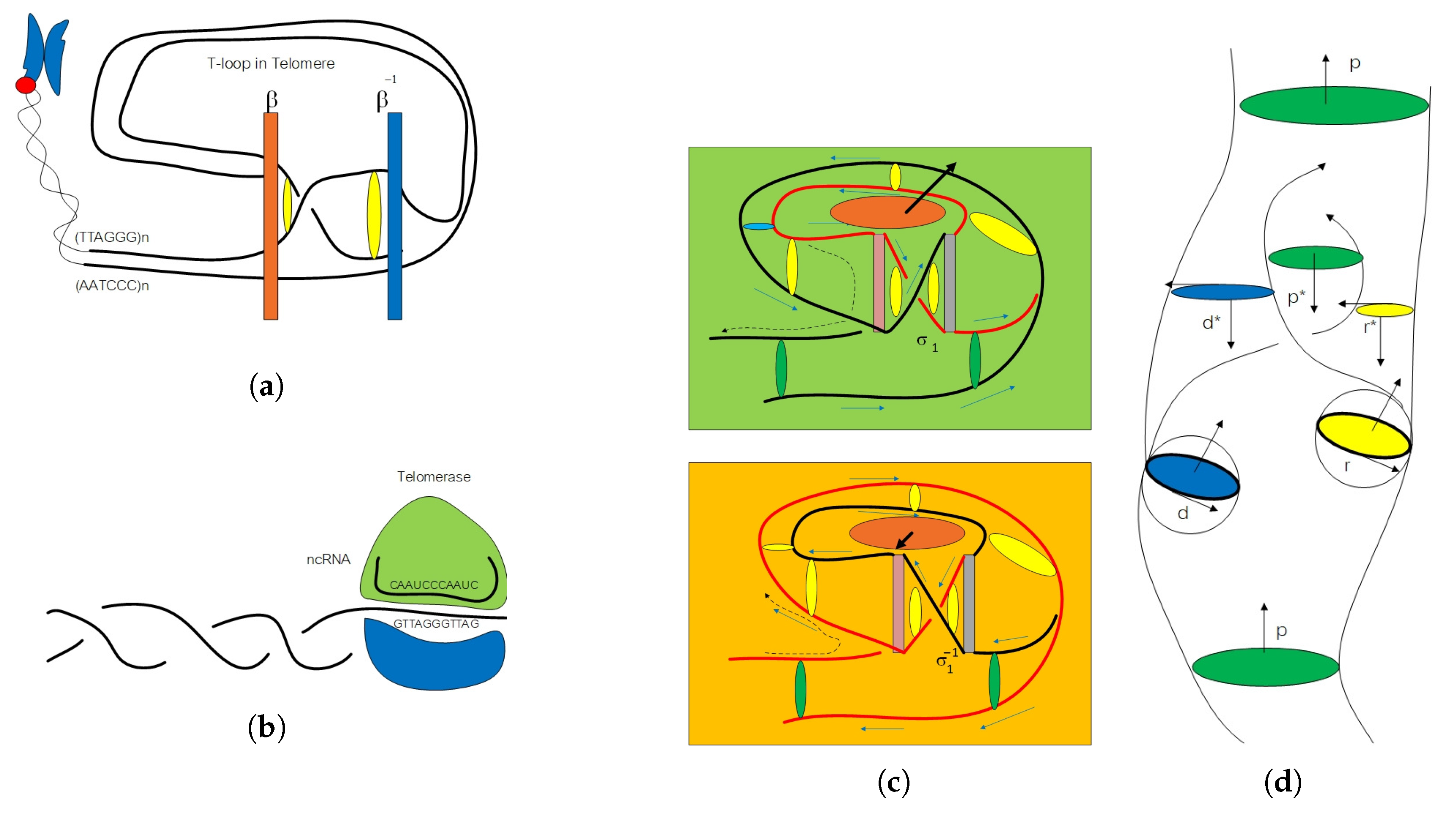
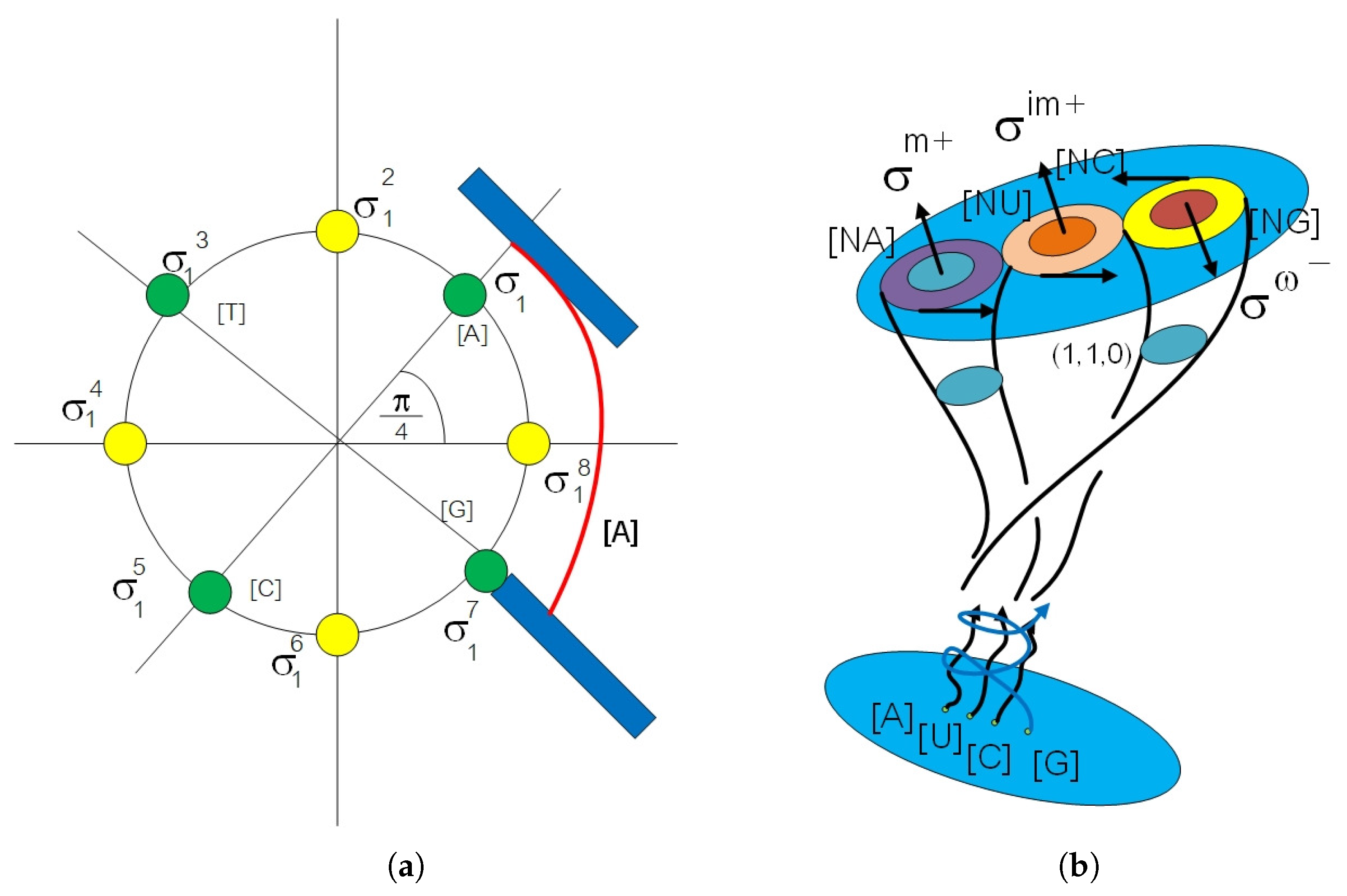
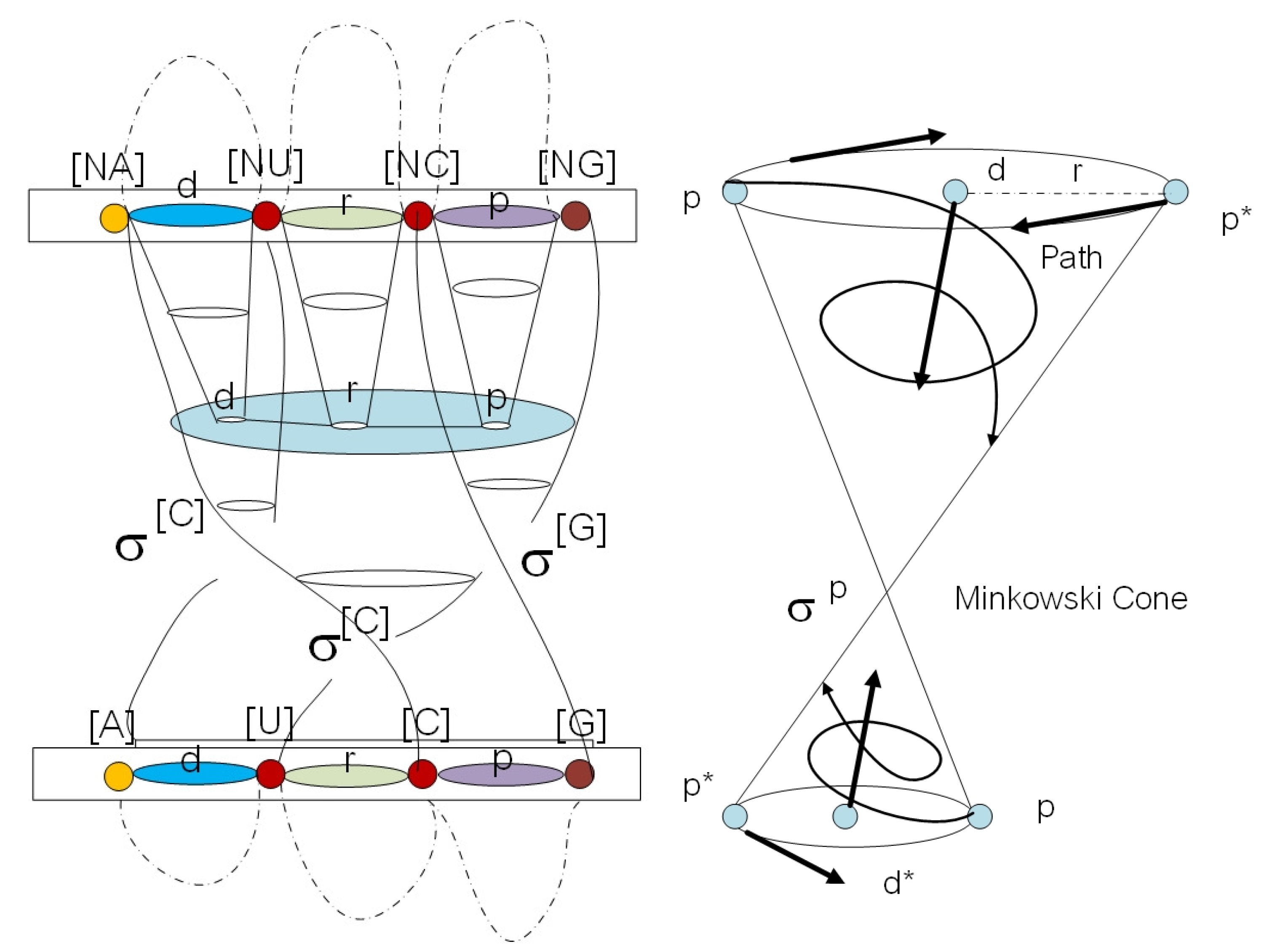
2. Algebraic Construction of G-Quadruplex in Telomere
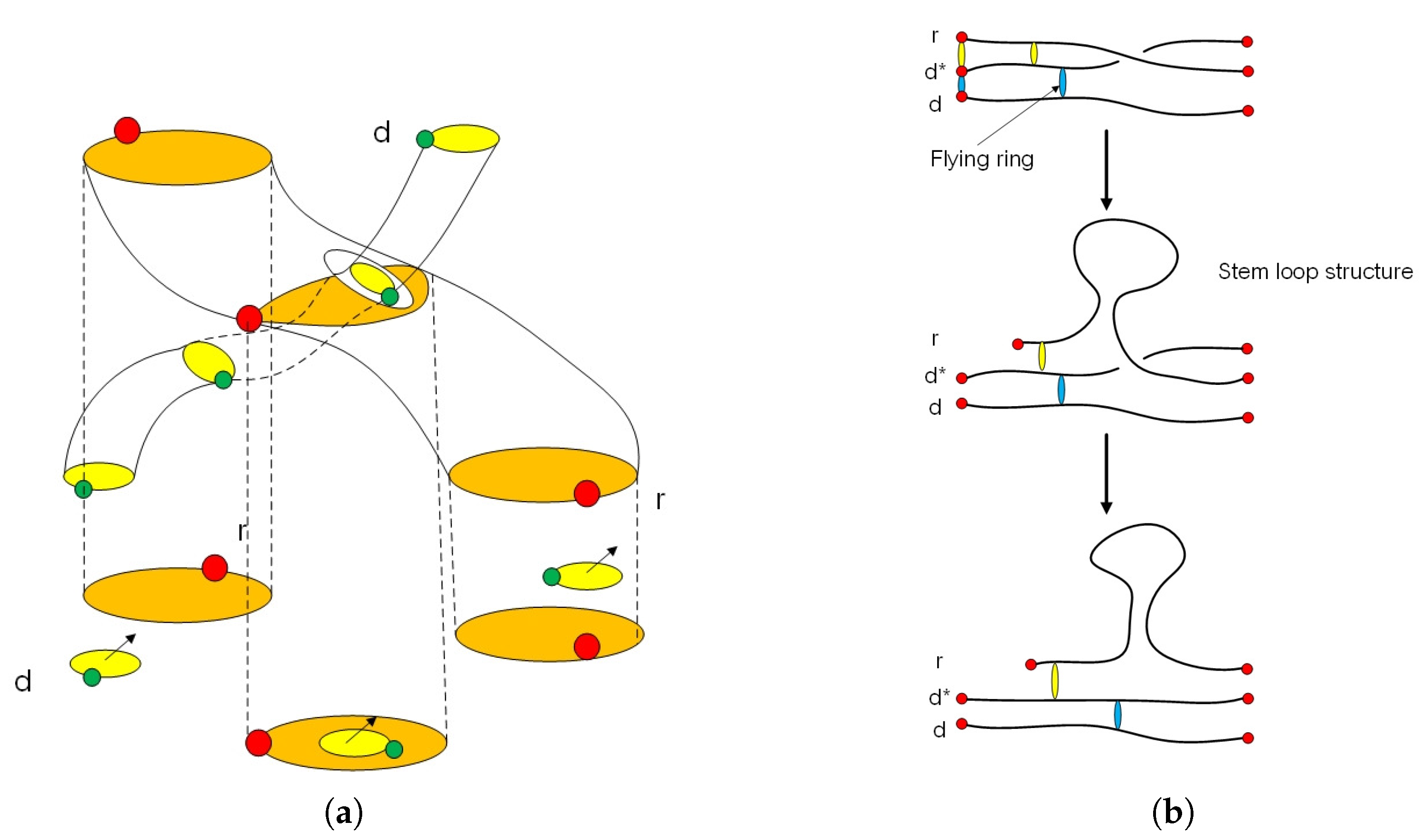
2.1. Anyon in Biology and Configuration Phase Space for Protein Folding
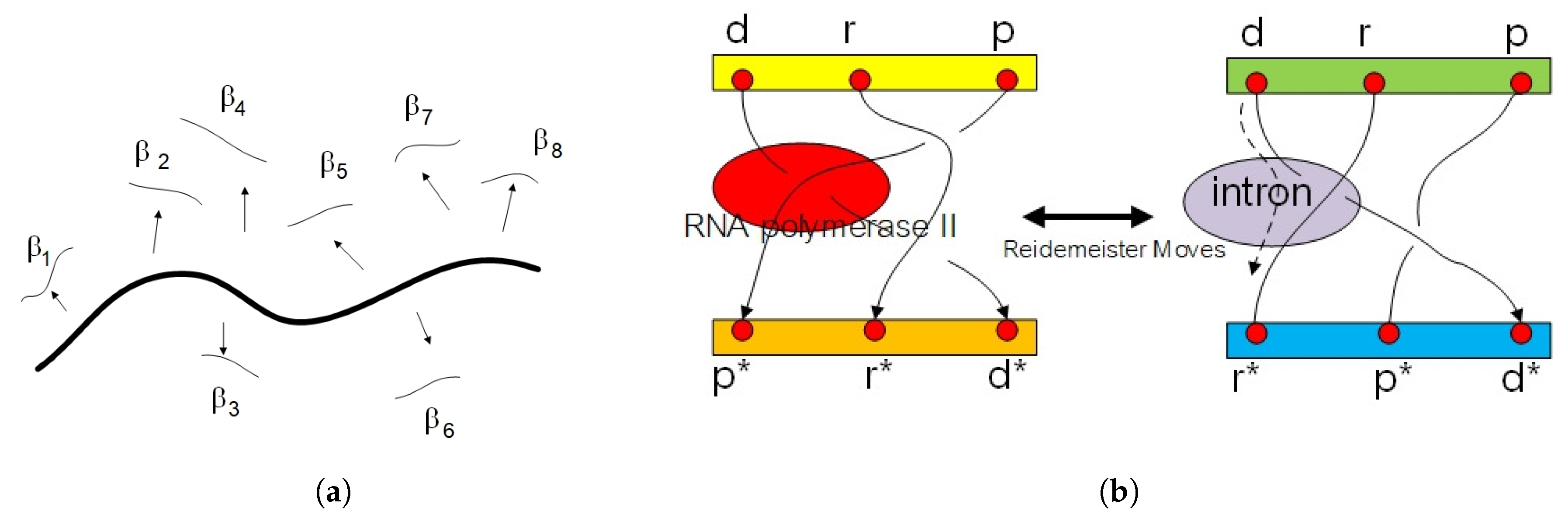
2.2. Circular Artin Braid Group Representation for Spinor Field in Genetic Code
2.3. Classification of the Loop Braid Group in Genetic Code
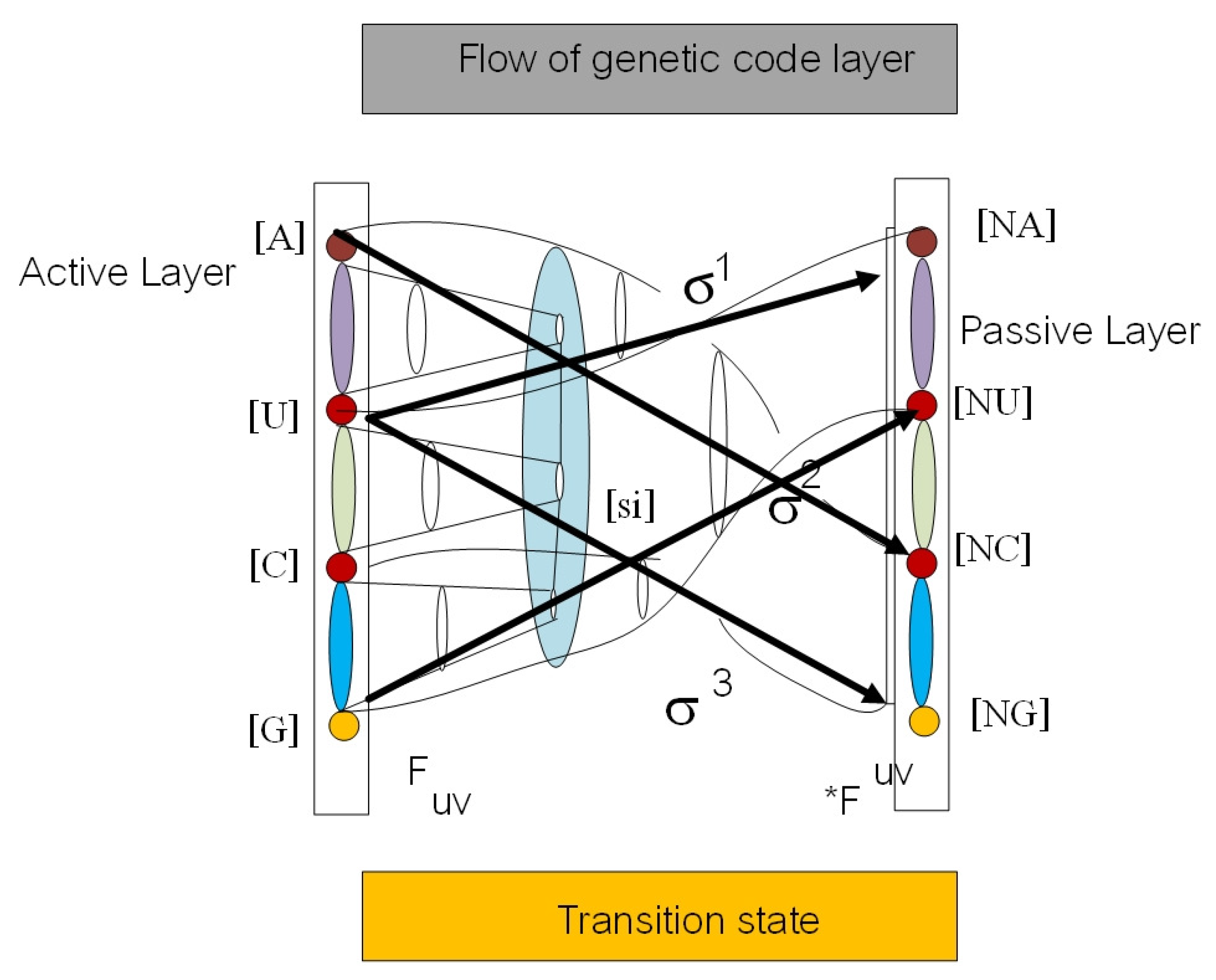
3. Affine Loop Braid Group in Public Key Cryptosystem of Protein Folding
3.1. Public Key Biological Cryptosystem in Protein–Protein Docking
- Preparatory step: A suitable pair of integers are chosen, and a sufficiently complicated -loop braid is selected and published through the gene expression.
- Key agreement: A key is shared through the protein folding by performing the following steps each time a shared key is required:
- Protein A chooses a random secret loop braid and sends to B.
- Protein B chooses a random secret loop braid and sends to A.
- Protein A receives and computes the shared key .
- Protein B receives and computes the shared key .
Since , we haveThus both proteins A and B obtain the same loop braid group element with the same curvature. Therefore they can dock to each other; otherwise the system will not be in equilibrium and the moduli state space for the gene expression with control equation will recursively loop back to send the key again.
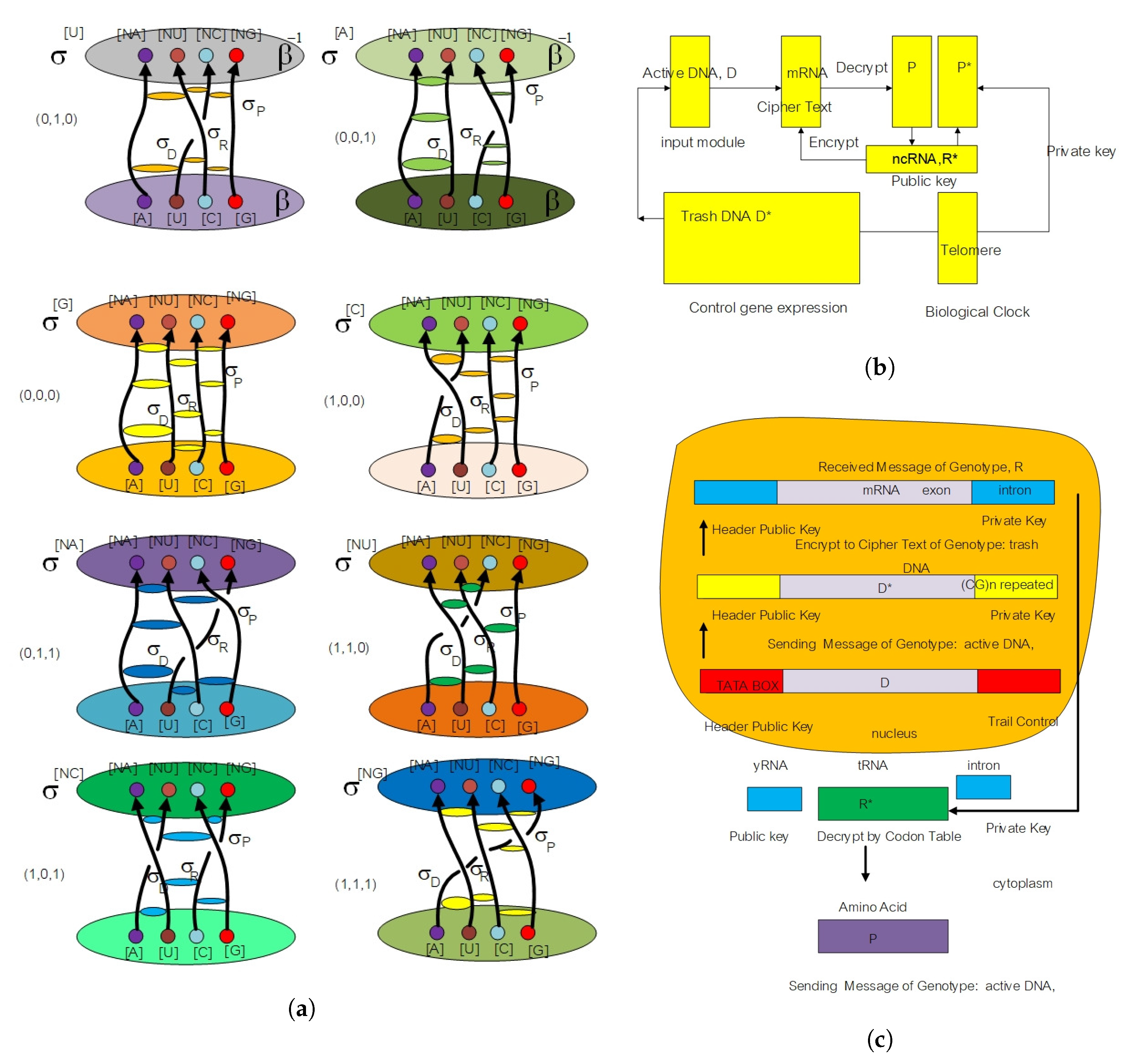
3.2. Moduli State Space Model for Protein–Protein Interaction
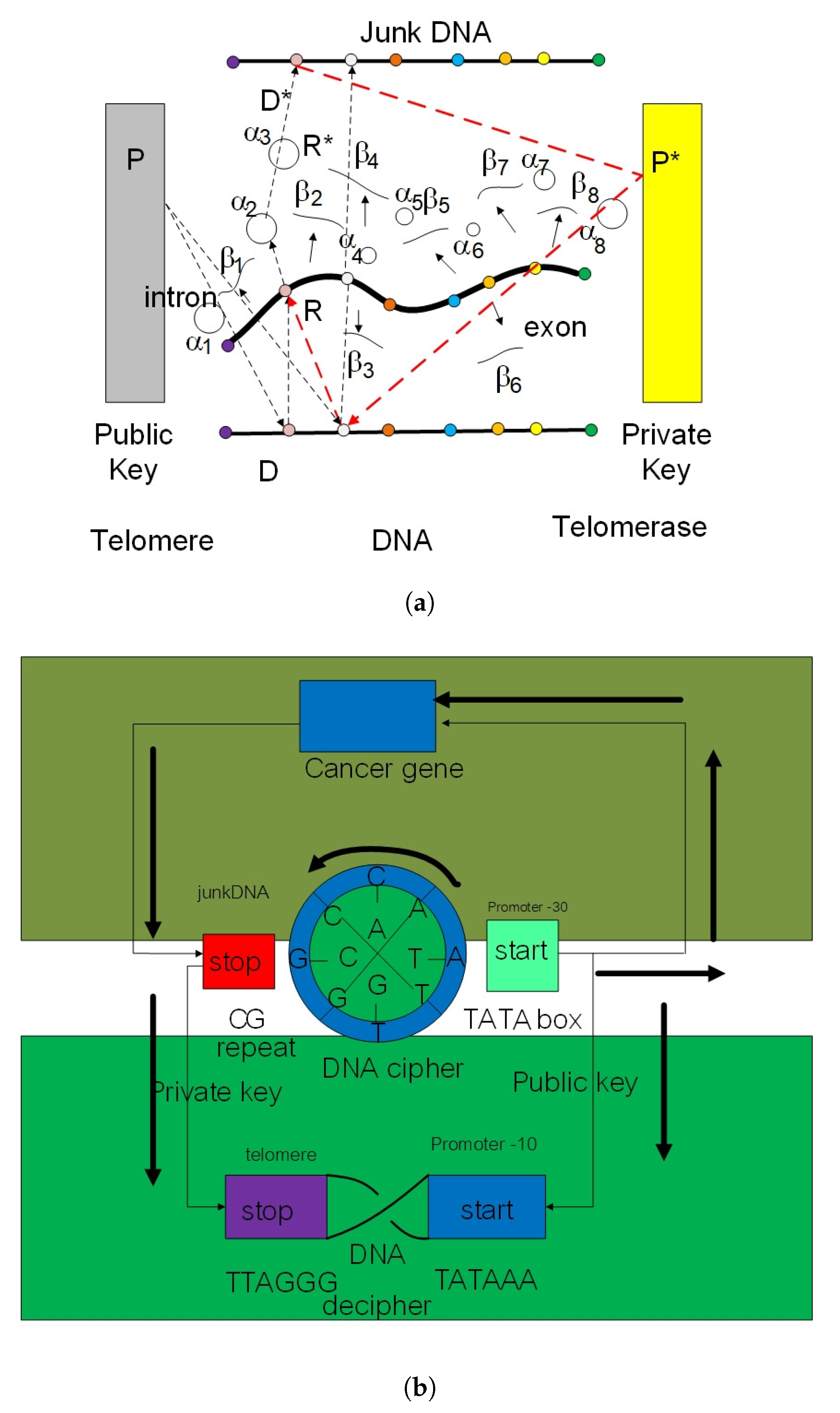
- Key generation module: Protein A and B choose a sufficiently complicated genotype in the loop braid group in genetic code and a hash with a fixed size of 20 amino acids (modulo 20) while transmitting the public key over a two protein docking system. The left protein chooses a genotype with representation in loop braid group in the genetic code . The public key is where .
- Encryption module: Given a biological message authentication code by y-RNA, tRNA, and ncRNA as a hash function with spinor state and the public key , protein B chooses a loop braid group at a rate of random mutation . Biological cipher state is where , .
- Decryption module: Given the biological cipher state and the private key , compute the element .
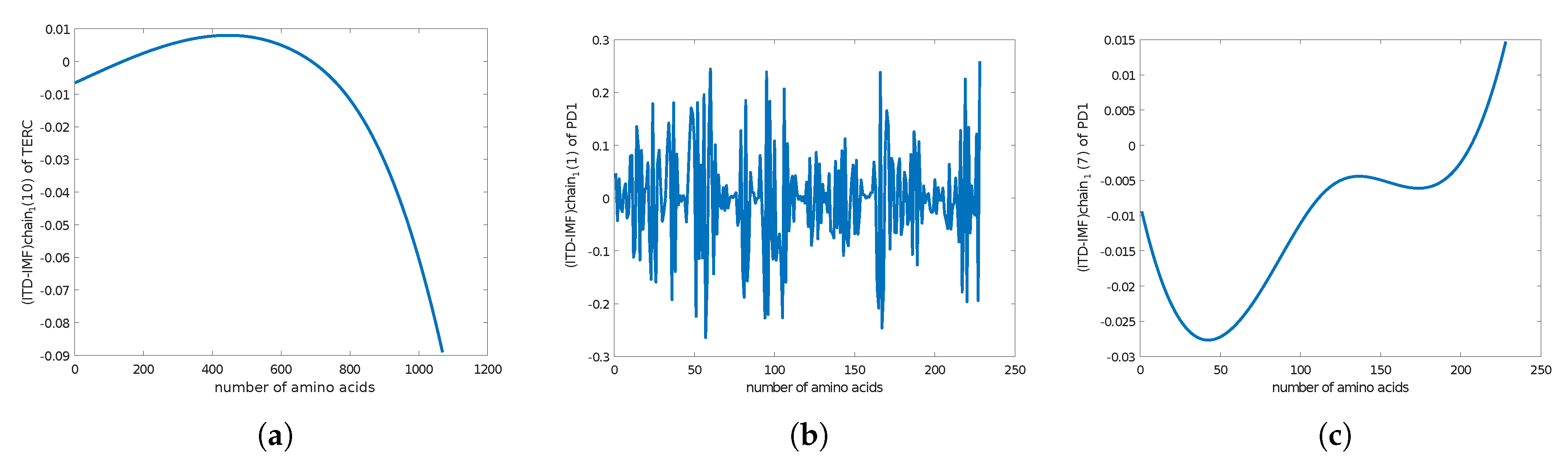
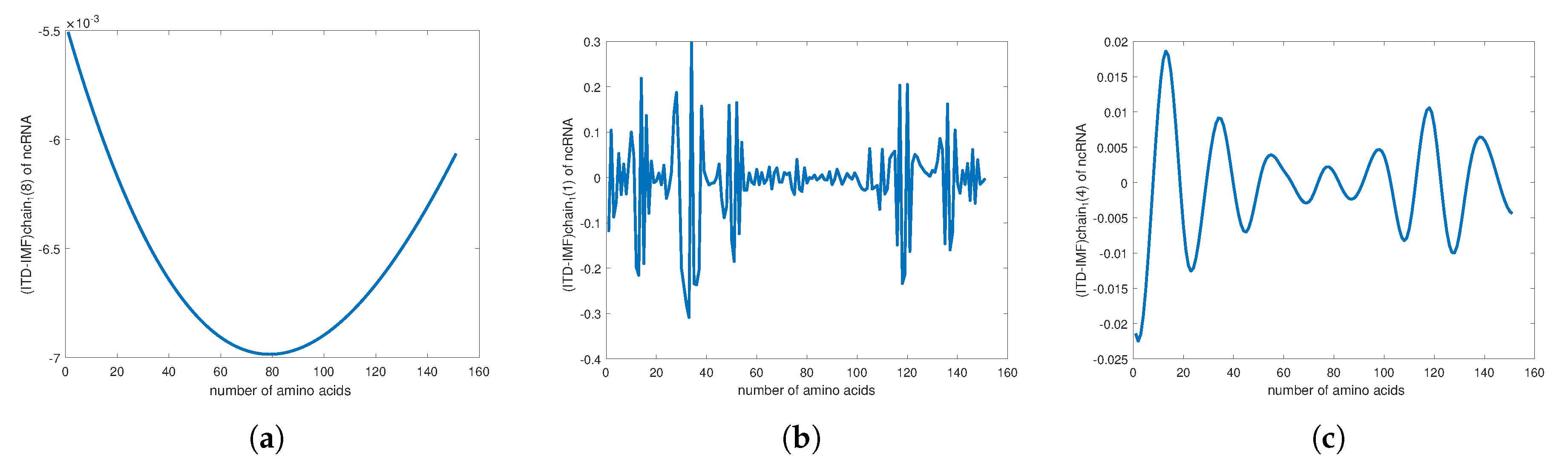
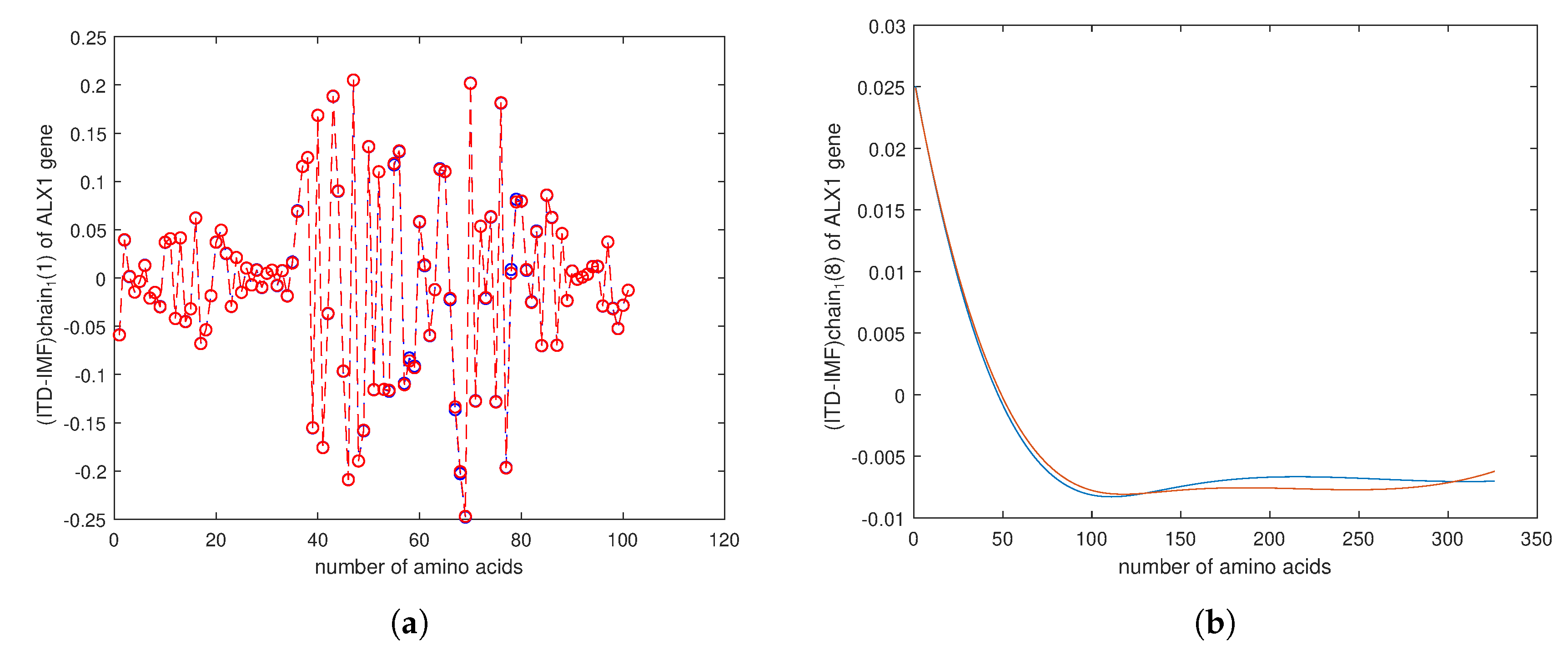
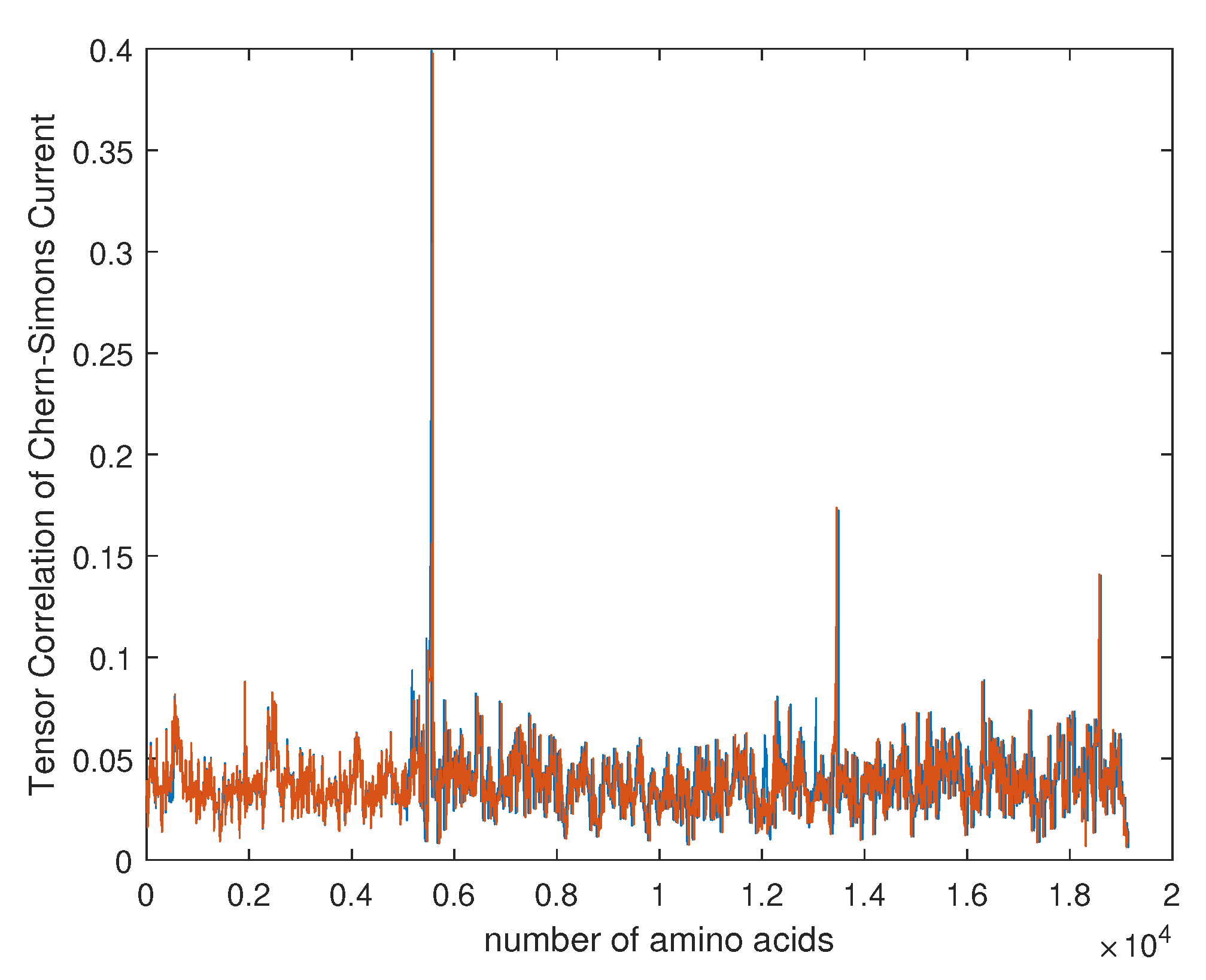
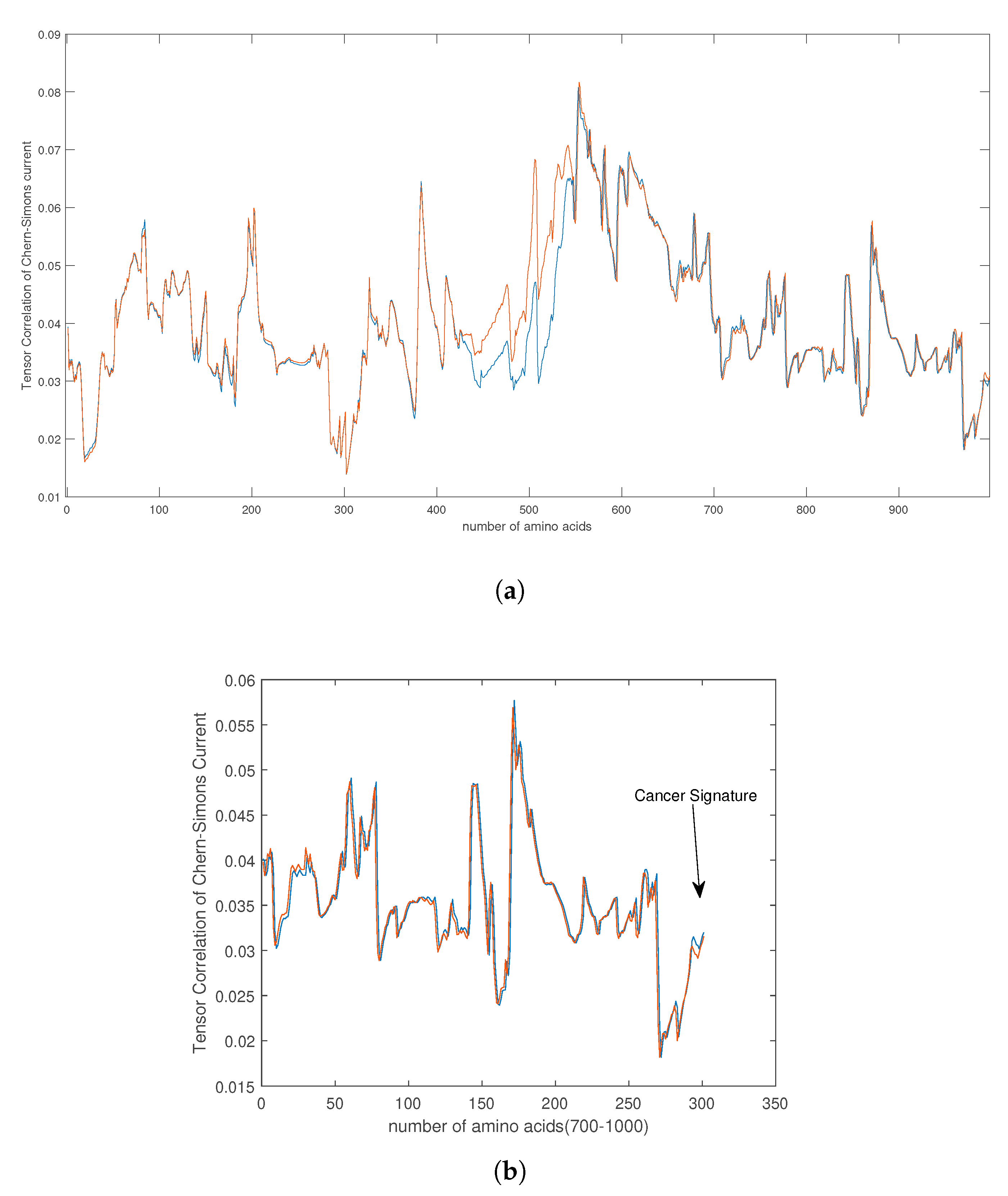
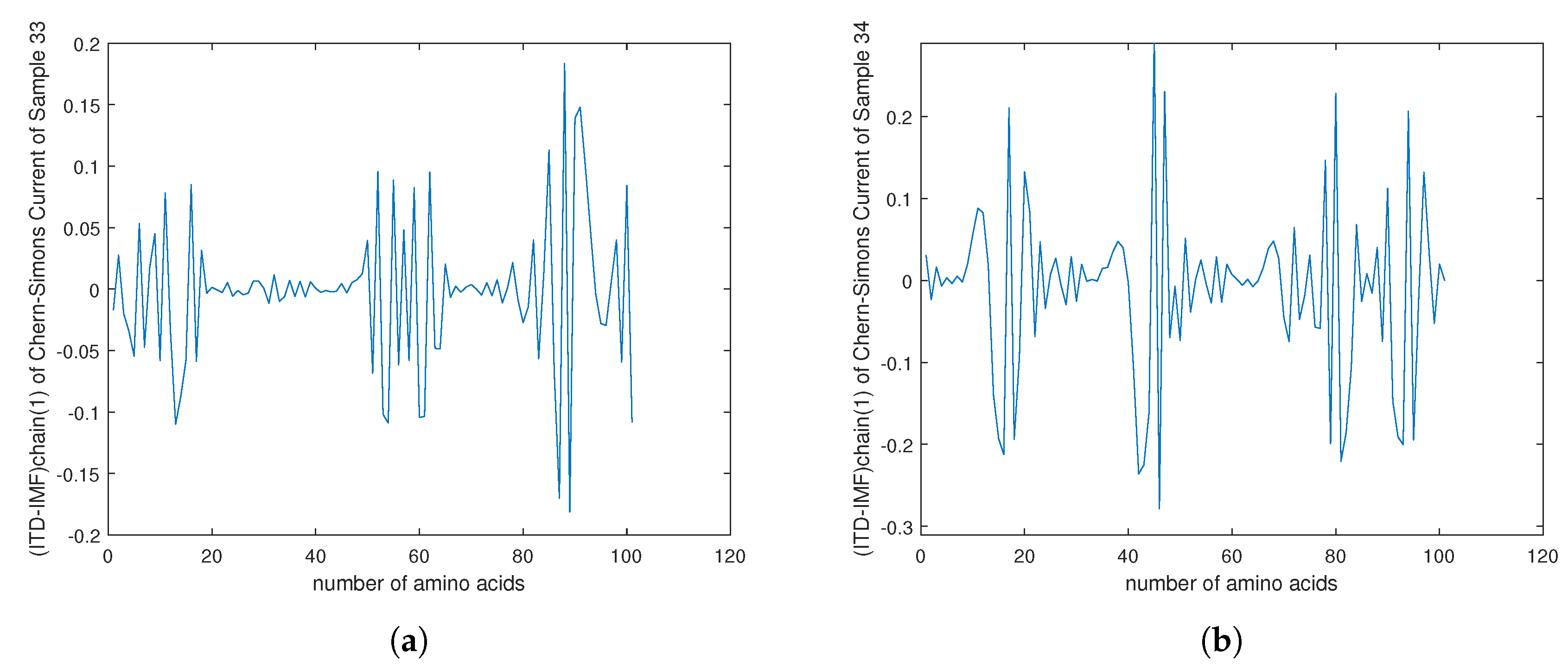
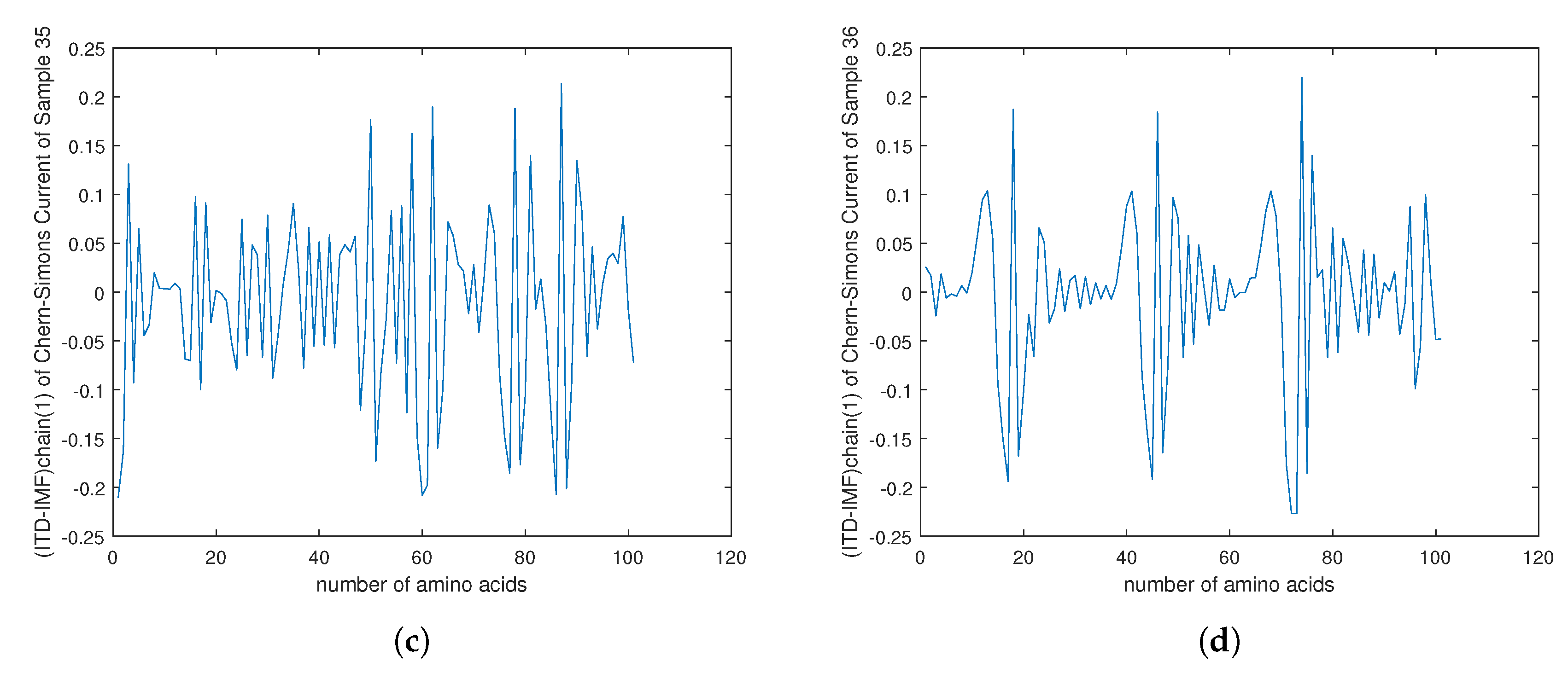
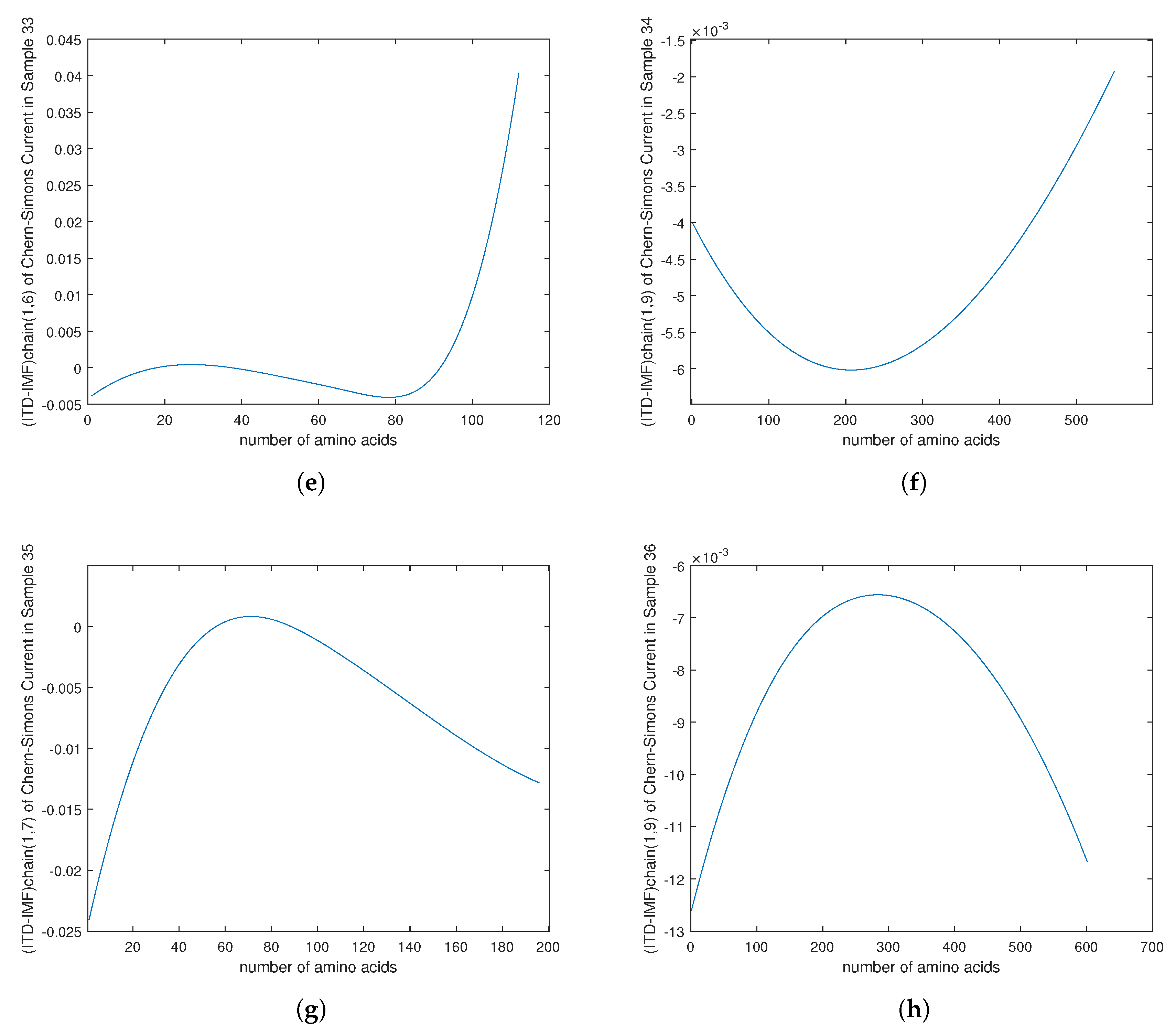
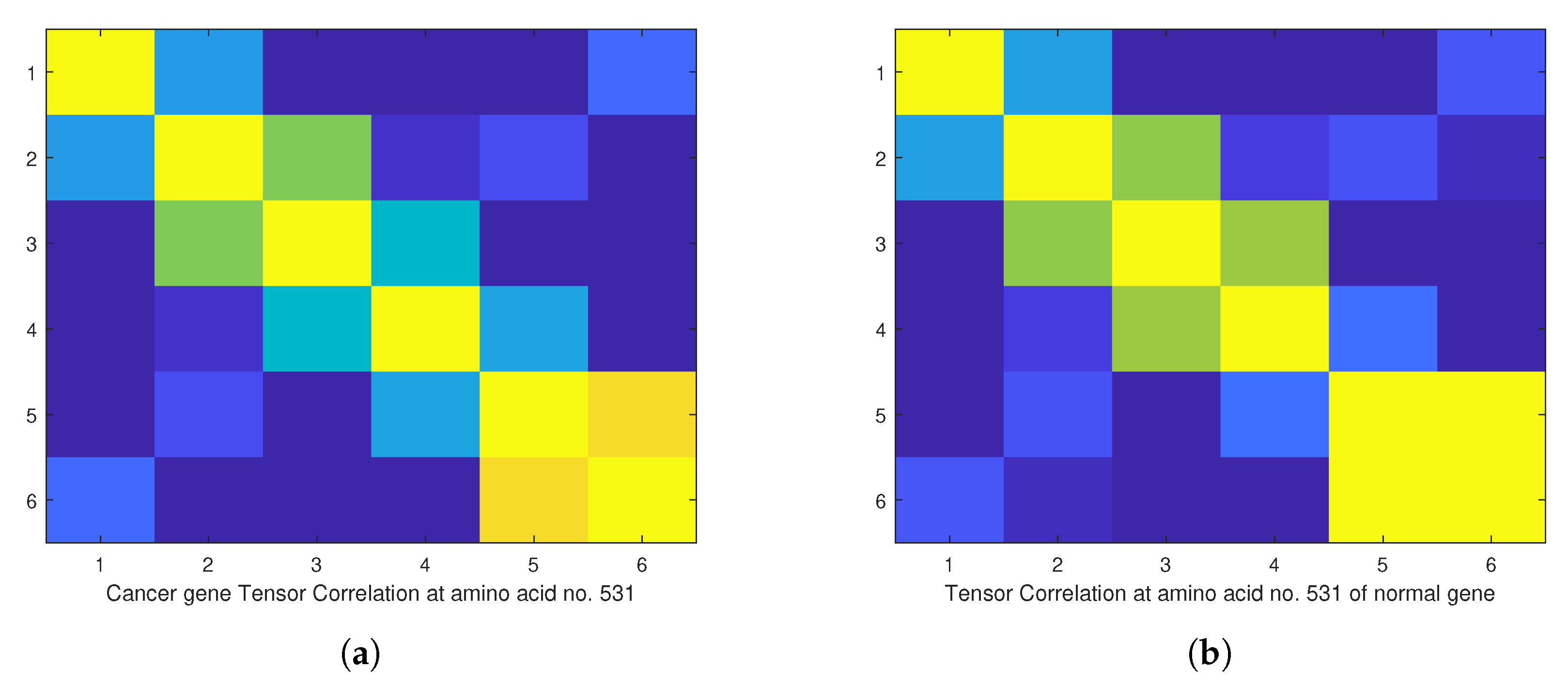
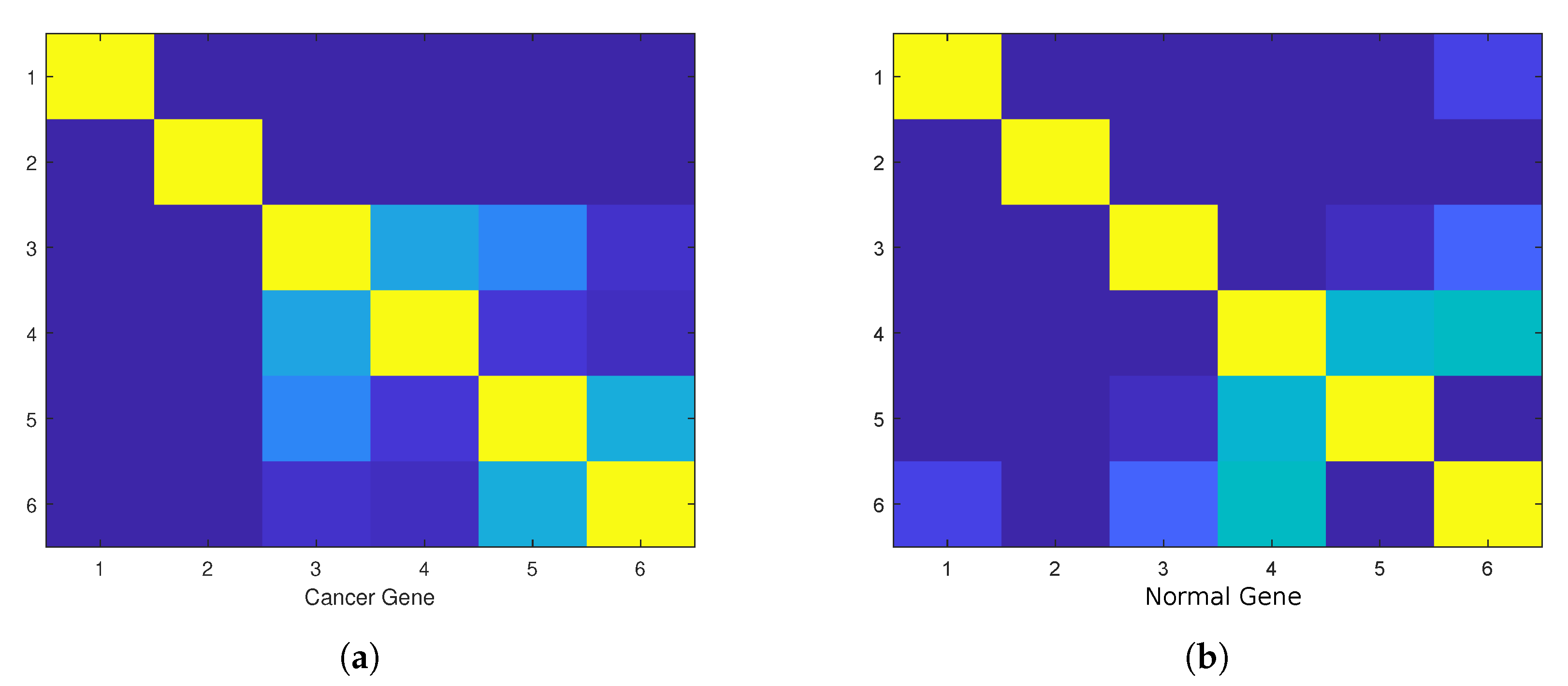
4. Empirical Analysis of Cancer Gene Signature
5. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Path Integral and Quantum Biology Theory
Appendix Largangian of Path Integral Formalism for System of DNA–RNA Transcription

Appendix B. The Source of Protein Folding Structure

Appendix C. Amino Acids Sequence of Noncoding RNA Sequence in Telomerase
Homo sapiens telomerase RNA component (TERC), telomerase RNA
NCBI Reference Sequence: NR_001566.1
FASTA Graphics
Go to:
LOCUS NR_001566 451 bp RNA linear PRI 21-OCT-2018
DEFINITION Homo sapiens telomerase RNA component (TERC), telomerase~RNA.
ncRNA 1..451
/ncRNA_class="telomerase_RNA"
/gene="TERC"
/gene_synonym="DKCA1; hTR; PFBMFT2; SCARNA19; TR; TRC3"
/product="telomerase RNA component"
/db_xref="GeneID:7012"
/db_xref="HGNC:HGNC:11727"
/db_xref="MIM:602322"
exon 1..451
/gene="TERC"
/gene_synonym="DKCA1; hTR; PFBMFT2; SCARNA19; TR; TRC3"
/inference="alignment:Splign:2.1.0"
misc_feature 46..55
/gene="TERC"
/gene_synonym="DKCA1; hTR; PFBMFT2; SCARNA19; TR; TRC3"
/note="template for telomere repeat TTAGGG"
ORIGIN
1 gggttgcgga gggtgggcct gggaggggtg gtggccattt tttgtctaac cctaactgag
61 aagggcgtag gcgccgtgct tttgctcccc gcgcgctgtt tttctcgctg actttcagcg
121 ggcggaaaag cctcggcctg ccgccttcca ccgttcattc tagagcaaac aaaaaatgtc
181 agctgctggc ccgttcgccc ctcccgggga cctgcggcgg gtcgcctgcc cagcccccga
241 accccgcctg gaggccgcgg tcggcccggg gcttctccgg aggcacccac tgccaccgcg
301 aagagttggg ctctgtcagc cgcgggtctc tcgggggcga gggcgaggtt caggcctttc
361 aggccgcagg aagaggaacg gagcgagtcc ccgcgcgcgg cgcgattccc tgagctgtgg
421 gacgtgcacc caggactcgg ctcacacatg~c
programmed cell death protein 1 precursor [Homo sapiens]
ORIGIN
1 mqipqapwpv vwavlqlgwr pgwfldspdr pwnpptfspa llvvtegdna tftcsfsnts
61 esfvlnwyrm spsnqtdkla afpedrsqpg qdcrfrvtql pngrdfhmsv vrarrndsgt
121 ylcgaislap kaqikeslra elrvterrae vptahpspsp rpagqfqtlv vgvvggllgs
181 lvllvwvlav icsraargti garrtgqplk edpsavpvfs vdygeldfqw rektpeppvp
241 cvpeqteyat ivfpsgmgts sparrgsadg prsaqplrpe~dghcswpl
Appendix D. Amino Acide Sequence of Telomoerase
/product="telomerase reverse transcriptase isoform~2"
Region 460..594
/region_name="telomerase_RBD"
/note="telomerase ribonucleoprotein complex - RNA binding
domain; smart00975"
/db_xref="CDD:214948"
Region 618..>729
/region_name="RT_like"
/note="RT_like: Reverse transcriptase (RT, RNA-dependent
DNA polymerase)_like family. An~RT gene is usually
indicative of a mobile element such as a retrotransposon
or retrovirus. RTs occur in a variety of mobile elements,
including retrotransposons; cl02808"
/db_xref="CDD:295487"
Region 825..>884
/region_name="TERT"
/note="TERT: telomerase reverse transcriptase (TERT).
telomerase is a ribonucleoprotein (RNP) that synthesizes
telomeric DNA repeats. The~telomerase RNA subunit provides
the template for synthesis of these repeats. The~catalytic
subunit of RNP is known as...; cd01648"
/db_xref="CDD:238826"
Site 834
/site_type="other"
/note="putative nucleic acid binding site [nucleotide
binding]"
/db_xref="CDD:238826"
CDS 1..1069
/gene="TERT"
ORIGIN
1 mpraprcrav rsllrshyre vlplatfvrr lgpqgwrlvq rgdpaafral vaqclvcvpw
61 darpppaaps frqvsclkel varvlqrlce rgaknvlafg falldgargg ppeafttsvr
121 sylpntvtda lrgsgawgll lrrvgddvlv hllarcalfv lvapscayqv cgpplyqlga
181 atqarpppha sgprrrlgce rawnhsvrea gvplglpapg arrrggsasr slplpkrprr
241 gaapepertp vgqgswahpg rtrgpsdrgf cvvsparpae eatslegals gtrhshpsvg
301 rqhhagppst srpprpwdtp cppvyaetkh flyssgdkeq lrpsfllssl rpsltgarrl
361 vetiflgsrp wmpgtprrlp rlpqrywqmr plflellgnh aqcpygvllk thcplraavt
421 paagvcarek pqgsvaapee edtdprrlvq llrqhsspwq vygfvraclr rlvppglwgs
481 rhnerrflrn tkkfislgkh aklslqeltw kmsvrdcawl rrspgvgcvp aaehrlreei
541 lakflhwlms vyvvellrsf fyvtettfqk nrlffyrksv wsklqsigir qhlkrvqlre
601 lseaevrqhr earpalltsr lrfipkpdgl rpivnmdyvv gartfrrekr aerltsrvka
661 lfsvlnyera rrpgllgasv lglddihraw rtfvlrvraq dpppelyfvk vdvtgaydti
721 pqdrltevia siikpqntyc vrryavvqka ahghvrkafk shvstltdlq pymrqfvahl
781 qetsplrdav vieqssslne assglfdvfl rfmchhavri rgksyvqcqg ipqgsilstl
841 lcslcygdme nklfagirrd glllrlvddf llvtphltha ktflsyarts irasltfnrg
901 fkagrnmrrk lfgvlrlkch slfldlqvns lqtvctniyk illlqayrfh acvlqlpfhq
961 qvwknptffl rvisdtaslc ysilkaknag mslgakgaag plpseavqwl chqafllklt
1021 rhrvtyvpll gslrtaqtql srklpgttlt aleaaanpal~psdfktild
Appendix E. Cipher Text for Encryption in ALX1 Gene
-Encryption-
Ciphertext:
2984 3181 2990 22 2639 3181 1680 2990
2112 22 1680 2639 180 180 2639 1680 1993 2639
403 2990 260 2984 1091 2112 1091 1091 180 22
3181 3192 2526 2984 3181 3544 22 403 1993 3181
2639 2990 260 2639 1680 2112 2639 2112 1091 1680
1675 2526 1221 2112 2990 1091 180 22 180 2275
2112 3181 3192 3192 2526 2275 22 3181 2275 3544
2639 180 1675 1221 403 2639 2639 2526 1993 260
1091 3756 3544 1680 2526 3181 1091 1221 180 22
3192 3544 3181 22 1993 2275 2112 2984 403 1993
1675 1993 2639 22 2275 2984 2639 180 2526 1680
1091 2984 1221 3181 1680 1091 3181 22 403 3181
22 1091 403 1680 1675 403 2639 1993 2526 2639
2639 2639 1680 1680 2275 2275 3192 2275 3544 3544
2990 3544 2639 22 1221 22 3181 3181 22 3181
1680 2526 2990 1221 1680 3544 3192 260 180 403
2526 260 2526 2275 3181 1221 22 2112 22 2275
3544 3181 22 3544 3181 2112 2275 2526 1221 2526
1956 2990 1221 1993 2275 2275 2112 1680 1956 2275
1680 2275 3181 2275 260 1091 1221 3756 1221 1221
2112 1680 2639 3192 2990 2112 2112 3544 260 403
3756 2639 2526 22 180 2275 3544 403 2639 260
180 1221 3756 1221 1993 1993 22 1956 2112 1091
1993 2112 2639 1091 1091 2639 2526 2526 3544 2639
1675 2984 22 180 2275 403 3544 2639 2639 1675
2984 3544 180 260 2639 3192 2639 180 2275 3544
403 2639 2639 260 3544 1091 2990 2639 1993 3192
1221 1993 1221 2990 2639 3192 2526 180 22 1993
1993 2990 2990 3544 403 2639 22 22 3544 1091
2112 3544 1993 1091 3192 2112 2990 3181 3544 1680
180 3181 2990 3181 2275 2275 2639 2639 2639 3756
2112 2526 22 2275 2984 1680 2112 1680 3181 3192
3544 2112 1993 3756 2639 1956 2112~2984
encrypt message in nucleotide sequence:
Restored Message: ’MEFLSEKFALKSPPSKNSDFYMGAGGPLEHVMET
LDNESFYSKASAGKCVQAFGPLPRAEHHVRLERTSPCQDSSVNYGITKVEGQPLHTELNR
AMDNCNSLRMSPVKGMQEKGELDELGDKCDSNVSSSKKRRHRTTFTSLQLEELEKVFQKTH
YPDVYVREQLALRTELTEARVQVWFQNRRAKWRKRERYGQIQQAKSHFAATYDISVLPRTDS
YPQIQNNLWAGNASGGSVVTSCMLPRDTSSCMTPYSHSPRTDSSYTGFSNHQNQFSHVPLNNF
FTDSLLTGATNGHAFETKPEFERRSSSIAVLRMKAKEHTANISWAM’
-Key Pair-
Modulus: 3953
Public Exponent: 5
Private Exponent: 2297
-Signing-
Signature: 2338 2587 1861 1967 974 2587 2926
1861 3376 1967 2926 974 3839 3839 974 2926 2172 974
3686 1861 3292 2338 1882 3376 1882 1882 3839 1967 2587
276 942 2338 2587 2401 1967 3686 2172 2587 974 1861
3292 974 2926 3376 974 3376 1882 2926 2412 942 2318
3376 1861 1882 3839 1967 3839 952 3376 2587 276 276
942 952 1967 2587 952 2401 974 3839 2412 2318 3686
974 974 942 2172 3292 1882 457 2401 2926 942 2587
1882 2318 3839 1967 276 2401 2587 1967 2172 952 3376
2338 3686 2172 2412 2172 974 1967 952 2338 974 3839
942 2926 1882 2338 2318 2587 2926 1882 2587 1967 3686
2587 1967 1882 3686 2926 2412 3686 974 2172 942 974
974 974 2926 2926 952 952 276 952 2401 2401 1861
2401 974 1967 2318 1967 2587 2587 1967 2587 2926 942
1861 2318 2926 2401 276 3292 3839 3686 942 3292 942
952 2587 2318 1967 3376 1967 952 2401 2587 1967 2401
2587 3376 952 942 2318 942 3733 1861 2318 2172 952
952 3376 2926 3733 952 2926 952 2587 952 3292 1882
2318 457 2318 2318 3376 2926 974 276 1861 3376 3376
2401 3292 3686 457 974 942 1967 3839 952 2401 3686
974 3292 3839 2318 457 2318 2172 2172 1967 3733 3376
1882 2172 3376 974 1882 1882 974 942 942 2401 974
2412 2338 1967 3839 952 3686 2401 974 974 2412 2338
2401 3839 3292 974 276 974 3839 952 2401 3686 974
974 3292 2401 1882 1861 974 2172 276 2318 2172 2318
1861 974 276 942 3839 1967 2172 2172 1861 1861 2401
3686 974 1967 1967 2401 1882 3376 2401 2172 1882 276
3376 1861 2587 2401 2926 3839 2587 1861 2587 952 952
974 974 974 457 3376 942 1967 952 2338 2926 3376
2926 2587 276 2401 3376 2172 457 974 3733 3376~2338
References
- Anshel, I.; Anshel, M.; Fisher, B.; Goldfeld, D. New key agreement protocols in braid group cryptography. Lect. Notes Comput. Sci. 2001, 2020, 13–27. [Google Scholar]
- Birman, J.; Ko, K.; Lee, S. A New Approach to the Word and Conjugacy Problems in the Braid Groups. Adv. Math. 1998, 139, 322–353. [Google Scholar] [CrossRef]
- Anshel, I.; Anshel, M.; Goldfeld, D. An algebraic method for public-key cryptography. Math. Res. Lett. 1999, 6, 287–291. [Google Scholar] [CrossRef]
- Capozziello, S.; Pincak, R.; Kanjamapornkul, K.; Saridakis, E. The Chern-Simons Current in Systems of DNA-RNA Transcriptions. Ann. Der Phys. 2018, 530. [Google Scholar] [CrossRef]
- Capozziello, S.; Pincak, R. The Chern–Simons current in time series of knots and links in proteins. Ann. Phys. 2018, 393, 413–446. [Google Scholar] [CrossRef]
- Pincak, R.; Kaniamapornkul, K.; Bartoš, E. Forecasting Laurent Polynomial in the Chern–Simons current of V3 Loop Time Series. Annalen der Physik 2019, 1800375. [Google Scholar] [CrossRef]
- Srinivas, N.; Rachakonda, S.; Kumar, R. Telomeres and Telomere Length: A General Overview. Cancers 2020, 12, 558. [Google Scholar] [CrossRef]
- Trentin, E. Asymptotic Convergence of Soft-Constrained Neural Networks for Density Estimation. Mathematics 2020, 8, 572. [Google Scholar] [CrossRef]
- Bilson-Thompson, S.; Hackett, J.; Kauffman, L.; Wan, Y. Emergent braided matter of quantum geometry. Symmetry Integr. Geom. Methods Appl. 2012, 8, 014. [Google Scholar] [CrossRef]
- Kanjamapornkul, K.; Pinčák, R. Kolmogorov space in time series data. Math. Methods Appl. Sci. 2016, 4463–4483. [Google Scholar] [CrossRef]
- Lundholm, D.; Seiringer, R. Fermionic behavior of ideal anyons. Lett. Math. Phys. 2018, 108, 2523–2541. [Google Scholar] [CrossRef] [PubMed]
- Field, B.; Simula, T. Introduction to topological quantum computation with non-Abelian anyons. Quantum Sci. Technol. 2018, 3, 045004. [Google Scholar] [CrossRef]
- Rusca, D.; Boaron, A.; Grünenfelder, F.; Martin, A.; Zbinden, H. Finite-key analysis for the 1-decoy state QKD protocol. Appl. Phys. Lett. 2018, 112, 171104. [Google Scholar] [CrossRef]
- Bacco, D.; Vagniluca, I.; Da Lio, B.; Biagi, N.; Della Frera, A.; Calonico, D.; Toninelli, C.; Cataliotti, F.; Bellini, M.; Oxenløwe, L.; et al. Field trial of a three-state quantum key distribution scheme in the Florence metropolitan area. EPJ Quantum Technol. 2019, 6. [Google Scholar] [CrossRef]
- Armas, P.; Calcaterra, N. G-quadruplex in animal development: Contribution to gene expression and genomic heterogeneity. Mech. Dev. 2018, 154, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Damiani, C. A journey through loop braid groups. Expo. Math. 2017, 35, 252–285. [Google Scholar] [CrossRef]
- Gresnigt, N. Braids, normed division algebras, and Standard Model symmetries. Phys. Lett. Sect. B Nucl. Elem. Part. High-Energy Phys. 2018, 783, 212–221. [Google Scholar] [CrossRef]
- Noble, D. Central dogma or central debate? Physiology 2018, 33, 246–249. [Google Scholar] [CrossRef]
- Kanjamapornkul, K.; Pinčák, R.; Bartoš, E. The study of Thai stock market across the 2008 financial crisis. Physica 2016, A462, 117–133. [Google Scholar] [CrossRef]
- Kanjamapornkul, K.; Pinčák, R.; Bartoš, E. Cohomology theory for financial time series. Physica 2020, A546, 122212. [Google Scholar] [CrossRef]
- Pinčák, R.; Kanjamapornkul, K.; Bartoš, E. Cohomology theory for biological time series. Math. Methods Appl. Sci. 2020, 43, 552–579. [Google Scholar] [CrossRef]
- Ayeni, B.; Pfeifer, R.; Brennen, G. Phase transitions on a ladder of braided non-Abelian anyons. Phys. Rev. B 2018, 98, 045432. [Google Scholar] [CrossRef]
- Saad, N. A ribonucleopeptide world at the origin of life. J. Syst. Evol. 2018, 56, 1–13. [Google Scholar] [CrossRef]
- Carey, N. Junk DNA. A Journey through the Dark Matter of the Genome; Icon Books Ltd.: London, UK, 2015. [Google Scholar]
- Harley, C.; Futcher, A.; Greider, C. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef]
- Ivancich, M.; Schrank, Z.; Wojdyla, L.; Leviskas, B.; Kuckovic, A.; Sanjali, A.; Puri, N. Treating cancer by targeting telomeres and telomerase. Antioxidants 2017, 6, 15. [Google Scholar] [CrossRef]
- Jafri, M.; Ansari, S.; Alqahtani, M.; Shay, J. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef]
- Bilson-Thompson, S. Braided topology and the emergence of matter. J. Phys. Conf. Ser. 2012, 360. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, 3156. [Google Scholar] [CrossRef]
- Nakamura, T.; Cech, T. Reversing time: Origin of telomerase. Cell 1998, 92, 587–590. [Google Scholar] [CrossRef]
- Wigner, E. On the quantum correction for thermodynamic equilibrium. Phys. Rev. 1932, 40, 749–759. [Google Scholar] [CrossRef]
- Christian, T.; Sakaguchi, R.; Perlinska, A.; Lahoud, G.; Ito, T.; Taylor, E.; Yokoyama, S.; Sulkowska, J.; Hou, Y.M. Methyl transfer by substrate signaling from a knotted protein fold. Nat. Struct. Mol. Biol. 2016, 23, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Gene ID: 7012. TERC Telomerase RNA Component. Bethesda (MD): National Library of Medicine (US). National Center for Biotechnology Information; 2004-[cited 2018]. Available online: https://www.ncbi.nlm.nih.gov/gene/?term=7012 (accessed on 14 June 2020).
- Tate, J.G. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [PubMed]
- Pinčák, R.; Kanjamapornkul, K.; Bartoš, E. A theoretical investigation on the predictability of genetic patterns. Chem. Phys. 2020, 535. [Google Scholar] [CrossRef]
- Pinčák, R.; Bartoš, E. Chemical evolution of protein folding in amino acids. Chem. Phys. 2020, 537, 110856. [Google Scholar] [CrossRef]
- Feynman, R.P.; Hibbs, A.R.; Styer, D.F. Quantum Mechanics and Path Integrals; Dover Books on Physics; Dover Publications: New York, NY, USA, 2010. [Google Scholar]
- Lim, A.P.C. Chern-Simons Path Integrals in S2 × S1. Mathematics 2015, 3, 843–879. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein No. (Sample No.) | Gene Name | Public Key (Normal) | Public Key (Cancer) | Private Key (Normal) | Private Key (Cancer) | Modulus (Normal) | Modulus (Cancer) | Size |
|---|---|---|---|---|---|---|---|---|
| ALX1 | 7 | 5 | 43 | 2297 | 341 | 3953 | 326 | |
| ANO2(1) | 3 | 7 | 467 | 1003 | 781 | 2449 | 998 | |
| ANO2(2) | − | 5 | − | 437 | − | 2291 | 998 | |
| ANO2(3) | − | 11 | − | 191 | − | 2201 | 999 | |
| ATAD2 | 5 | 5 | 749 | 389 | 1027 | 703 | 1390 | |
| C20orf103 | 3 | 5 | 1531 | 2165 | 2407 | 5561 | 280 | |
| C6orf170 | 11 | 3 | 611 | 387 | 6887 | 649 | 1257 | |
| CTNNA2 | 3 | 3 | 2515 | 1203 | 3901 | 1909 | 905 | |
| DENND1B | 5 | 3 | 1037 | 2347 | 1387 | 3649 | 396 | |
| ERBB3(1) | 5 | 5 | 5117 | 845 | 6557 | 1139 | 1342 | |
| ERBB3(2) | − | 11 | − | 2291 | − | 4331 | 1342 | |
| ESYT2 | 3 | 3 | 3171 | 323 | 4897 | 529 | 893 | |
| F2RL1 | 5 | 5 | 1037 | 1037 | 1369 | 2701 | 397 | |
| FAF1 | 5 | 7 | 1373 | 823 | 7031 | 1037 | 650 | |
| GPR75 | 7 | 5 | 343 | 1373 | 1271 | 1817 | 540 | |
| IDH1 | 5 | 3 | 53 | 2843 | 299 | 4399 | 414 | |
| ISX | 3 | 3 | 2707 | 4811 | 4189 | 7387 | 245 | |
| MAGEB5 | 3 | 5 | 2707 | 749 | 4189 | 1343 | 275 | |
| MAML3(1) | 5 | 11 | 1109 | 611 | 1943 | 6887 | 1138 | |
| MAML3(2) | − | 7 | − | 2263 | − | 2747 | 1138 | |
| PTPN5(1) | 5 | 3 | 2765 | 675 | 3589 | 1081 | 565 | |
| PTPN5(2) | − | 3 | − | 299 | − | 493 | 565 | |
| Q6ZVS6 HUMAN | 3 | 3 | 763 | 4107 | 1219 | 6319 | 177 | |
| RAD23A | 3 | 5 | 1227 | 2297 | 1927 | 3953 | 363 | |
| RGS7 | 5 | 7 | 1217 | 2263 | 6241 | 5429 | 495 | |
| SETD4 | 5 | 7 | 281 | 103 | 1501 | 209 | 440 | |
| SGCD | 11 | 5 | 191 | 1997 | 2201 | 5141 | 290 | |
| SMAD1 | 3 | 3 | 547 | 1531 | 913 | 2407 | 465 | |
| SRCAP | 5 | 5 | 317 | 1901 | 1691 | 3293 | 3230 | |
| SULF1 | 11 | 5 | 1031 | 845 | 1349 | 1157 | 871 | |
| THOC4 | 3 | 3 | 851 | 675 | 1357 | 1081 | 257 | |
| UGT3A2 | 5 | 7 | 1037 | 2263 | 1843 | 2759 | 523 | |
| XAGE3 | 3 | 5 | 1307 | 1037 | 2059 | 1369 | 111 | |
| ZNF253 | − | 5 | − | 1373 | − | 3551 | 549 | |
| ZNF90(1) | − | 3 | − | 491 | − | 799 | 195 | |
| ZNF90(2) | − | 7 | − | 823 | − | 1517 | 601 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capozziello, S.; Pinčák, R.; Bartoš, E. A Supersymmetry and Quantum Cryptosystem with Path Integral Approach in Biology. Symmetry 2020, 12, 1214. https://doi.org/10.3390/sym12081214
Capozziello S, Pinčák R, Bartoš E. A Supersymmetry and Quantum Cryptosystem with Path Integral Approach in Biology. Symmetry. 2020; 12(8):1214. https://doi.org/10.3390/sym12081214
Chicago/Turabian StyleCapozziello, Salvatore, Richard Pinčák, and Erik Bartoš. 2020. "A Supersymmetry and Quantum Cryptosystem with Path Integral Approach in Biology" Symmetry 12, no. 8: 1214. https://doi.org/10.3390/sym12081214
APA StyleCapozziello, S., Pinčák, R., & Bartoš, E. (2020). A Supersymmetry and Quantum Cryptosystem with Path Integral Approach in Biology. Symmetry, 12(8), 1214. https://doi.org/10.3390/sym12081214