Reading-Network in Developmental Dyslexia before and after Visual Training

Abstract
1. Introduction
2. Materials and Methods
2.1. Research Design
2.2. Children
2.3. Experimental Paradigm
2.4. EEG Recording and Signal Pre-Processing
2.5. Functional Connectivity Analysis
2.6. Minimum Spanning Tree
2.7. Statistical Analysis
3. Results
3.1. Behavior Results
3.2. Global Measures of MST
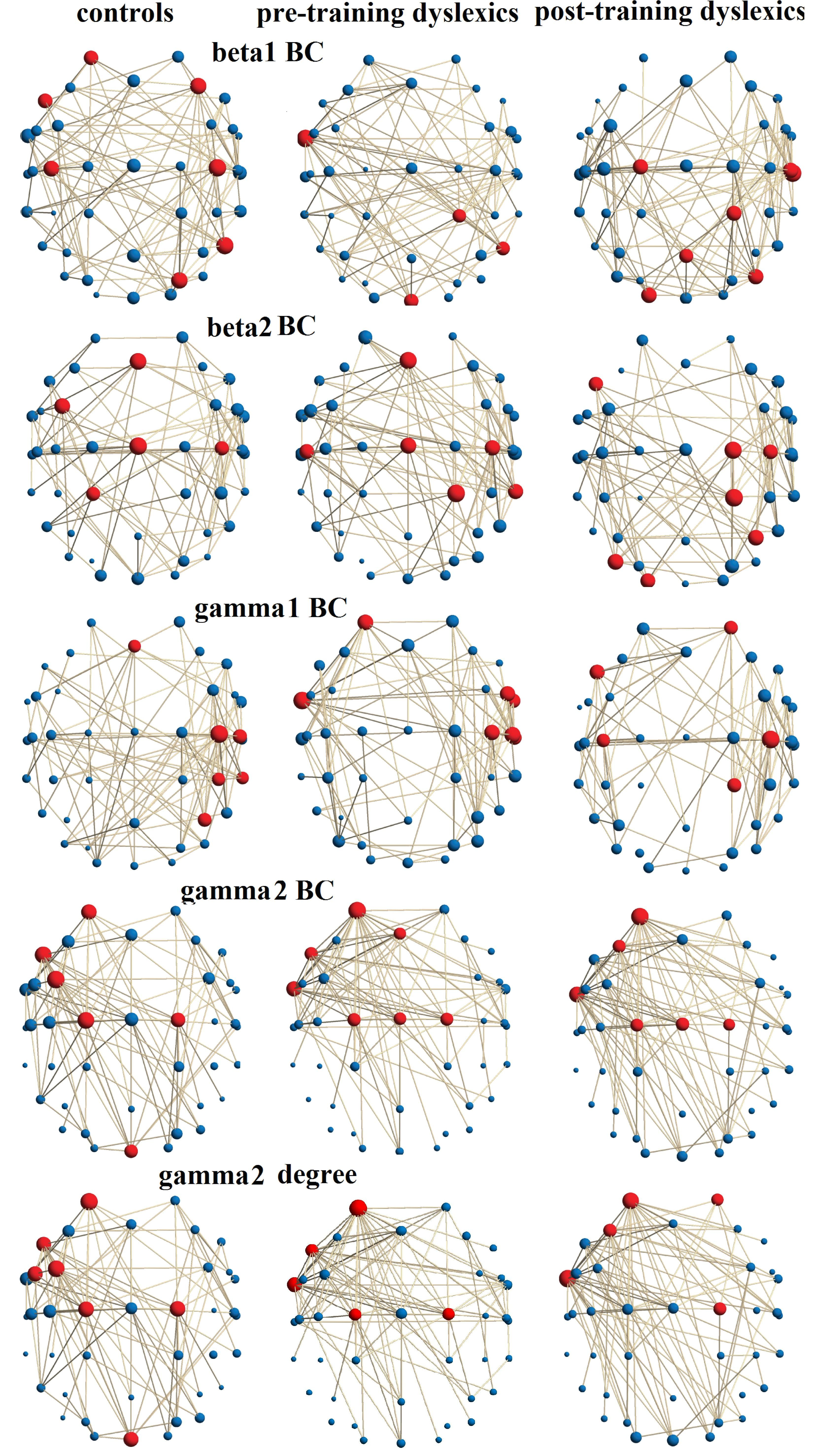
3.3. Distribution of Connectivity Hubs
4. Discussion
4.1. Altered Global Topology in Developmental Dyslexia
4.2. Distribution of the Connectivity Hubs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dehaene, S.; Cohen, L.; Morais, J.; Kolinsky, R. Illiterate to literate: Behavioural and cerebral changes induced by reading acquisition. Nat. Rev. Neurosci. 2015, 16, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Castles, A.; Rastle, K.; Nation, K. Ending the Reading Wars: Reading Acquisition from Novice to Expert. Psychol. Sci. Public Interes. 2018, 19, 5–51. [Google Scholar] [CrossRef] [PubMed]
- Share, D.L. Phonological recoding and self-teaching: Sine qua non of reading acquisition. Cognition 1995, 55, 151–218. [Google Scholar] [CrossRef]
- Landerl, K.; Ramus, F.; Moll, K.; Lyytinen, H.; Leppänen, P.H.T.; Lohvansuu, K.; O’Donovan, M.; Williams, J.; Bartling, J.; Bruder, J.; et al. Predictors of developmental dyslexia in European orthographies with varying complexity. J. Child Psychol. Psychiatry 2012, 54, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.; Pugh, K.R.; Hoeft, F. Neural Noise Hypothesis of Developmental Dyslexia. Trends Cogn. Sci. 2017, 21, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.S.; Bullmore, E. Small-World Brain Networks. Neuroscience 2006, 12, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Tewarie, P.; Van Dellen, E.; Hillebrand, A.; Stam, C.J. The minimum spanning tree: An unbiased method for brain network analysis. NeuroImage 2015, 104, 177–188. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Sowman, P.F.; Brock, J.; Etchell, A.C.; Stam, C.J.; Hillebrand, A. Increased segregation of functional networks in developing brains. NeuroImage 2019, 200, 607–620. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- González, G.F.; Van Der Molen, M.; Žarić, G.; Bonte, M.; Tijms, J.; Blomert, L.; Stam, C.J.; Van der Molen, M.W. Graph analysis of EEG resting state functional networks in dyslexic readers. Clin. Neurophysiol. 2016, 127, 3165–3175. [Google Scholar]
- Boets, B.; Vandermosten, M.; Cornelissen, P.; Wouters, J.; Ghesquière, P. Coherent Motion Sensitivity and Reading Development in the Transition from Prereading to Reading Stage. Child Dev. 2011, 82, 854–869. [Google Scholar] [CrossRef]
- Demb, J.B.; Boynton, G.M.; Best, M.; Heeger, D.J. Psychophysical evidence for a magnocellular pathway deficit in dyslexia. Vis. Res. 1998, 38(11), 1555–1559. [Google Scholar] [CrossRef]
- Lalova, J.; Dushanova, J.; Kalonkina, A.; Tsokov, S. Application of specialised psychometric tests and training practices in children with developmental dyslexia. Psychol. Res. 2019, 22, 271–283. [Google Scholar]
- Cornelissen, P.L.; Hansen, P.C.; Hutton, J.L.; Evangelinou, V.; Stein, J.F. Magnocellular visual function and children’s single word reading. Vision Res. 1998, 38, 471–482. [Google Scholar] [CrossRef]
- Stein, J.F. Dyslexia: The Role of Vision and Visual Attention. Curr. Dev. Disord. Rep. 2014, 1, 267–280. [Google Scholar] [CrossRef]
- Sperling, A.J.; Lu, Z.-L.; Manis, F.R.; Seidenberg, M.S. Deficits in perceptual noise exclusion in developmental dyslexia. Nat. Neurosci. 2005, 8, 862–863. [Google Scholar] [CrossRef]
- Wilmer, J.B.; Richardson, A.; Chen, Y.; Stein, J.F. Two Visual Motion Processing Deficits in Developmental Dyslexia Associated with Different Reading Skills Deficits. J. Cogn. Neurosci. 2004, 16, 528–540. [Google Scholar] [CrossRef]
- Goodale, M.A.; Westwood, D.A. An evolving view of duplex vision: Separate but interacting cortical pathways for perception and action. Curr. Opin. Neurobiol. 2004, 14, 203–211. [Google Scholar] [CrossRef]
- Lawton, T. Improving magnocellular function in the dorsal stream remediates reading deficits. Optom. Vis. Dev. 2011, 42, 142–154. [Google Scholar]
- Chouake, T.; Levy, T.; Javitt, D.C.; Lavidor, M. Magnocellular training improves visual word recognition. Front. Hum. Neurosci. 2012, 6, 14. [Google Scholar] [CrossRef]
- Qian, Y.; Bi, H.-Y. The effect of magnocellular-based visual-motor intervention on Chinese children with developmental dyslexia. Front. Psychol. 2015, 6. [Google Scholar] [CrossRef]
- Lawton, T.; Shelley-Tremblay, J. Training on Movement Figure-Ground Discrimination Remediates Low-Level Visual Timing Deficits in the Dorsal Stream, Improving High-Level Cognitive Functioning, Including Attention, Reading Fluency, and Working Memory. Front. Hum. Neurosci. 2017, 11, 236. [Google Scholar] [CrossRef]
- Ebrahimi, L.; Pouretemad, H.; Khatibi, A.; Stein, J. Magnocellular Based Visual Motion Training Improves Reading in Persian. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Lawton, T. Improving Dorsal Stream Function in Dyslexics by Training Figure/Ground Motion Discrimination Improves Attention, Reading Fluency, and Working Memory. Front. Hum. Neurosci. 2016, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Habib, M. The neurological basis of developmental dyslexia: An overview and working hypothesis. Brain 2000, 123, 2373–2399. [Google Scholar] [CrossRef]
- Raven, J.; Raven, J.C.; Court, J.H. Manual for Raven’s Progressive Matrices and Vocabulary Scales; Section 2: The coloured progressive, matrices; Oxford Psychologists Press: Oxford, UK; The Psychological Corporation: San Antonio, TX, USA, 1998. [Google Scholar]
- Sartori, G.; Remo, J.; Tressoldi, P.E. Updated and revised edition for the evaluation of dyslexia. In DDE-2, Battery for the Developmental Dyslexia and Evolutionary Disorders-2, 1995; Giunti O.S.: Florence, Italy, 2007. [Google Scholar]
- Benassi, M.; Simonelli, L.; Giovagnoli, S.; Bolzani, R. Coherence motion perception in developmental dyslexia: A meta-analysis of behavioral studies. Dyslexia 2010, 16, 341–357. [Google Scholar] [CrossRef]
- Joshi, M.R.; Falkenberg, H.K. Development of radial optic flow pattern sensitivity at different speeds. Vis. Res. 2015, 110, 68–75. [Google Scholar] [CrossRef]
- Pammer, K.; Wheatley, C. Isolating the M(y)-cell response in dyslexia using the spatial frequency doubling illusion. Vis. Res. 2001, 41, 2139–2147. [Google Scholar] [CrossRef]
- Lalova, J.; Dushanova, J.; Kalonkina, A.; Tsokov, S.; Hristov, I.; Totev, T.; Stefanova, M. Vision and visual attention of children with developmental dyslexia. Psychol. Res. 2018, 21, 247–261. [Google Scholar]
- Ross-Sheehy, S.; Oakes, L.M.; Luck, S.J. Exogenous attention influences visual short-term memory in infants. Dev. Sci. 2011, 14, 490–501. [Google Scholar] [CrossRef]
- Dushanova, J.; Christov, M. Auditory event-related brain potentials for an early discrimination between normal and pathological brain aging. Neural Regen. Res. 2013, 8, 1390–1399. [Google Scholar]
- Stam, C.J.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef]
- Kruskal, J.B. On the shortest spanning subtree of a graph and the traveling salesman problem. Proc. Am. Math. Soc. 1956, 7, 48. [Google Scholar] [CrossRef]
- Smith, K.; Abásolo, D.; Escudero, J. Accounting for the complex hierarchical topology of EEG phase-based functional connectivity in network binarisation. PLoS ONE 2017, 12, e0186164. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.T.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Stam, C.; Tewarie, P.; Van Dellen, E.; Van Straaten, E.; Hillebrand, A.; Van Mieghem, P. The trees and the forest: Characterization of complex brain networks with minimum spanning trees. Int. J. Psychophysiol. 2014, 92, 129–138. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Koessler, L.; Maillard, L.; Benhadid, A.; Vignal, J.; Felblinger, J.; Vespignani, H.; Braun, M. Automated cortical projection of EEG sensors: Anatomical correlation via the international 10–10 system. NeuroImage 2009, 46, 64–72. [Google Scholar] [CrossRef]
- Giacometti, P.; Perdue, K.L.; Diamond, S.G. Algorithm to find high density EEG scalp coordinates and analysis of their correspondence to structural and functional regions of the brain. J. Neurosci. Methods 2014, 229, 84–96. [Google Scholar] [CrossRef]
- Liu, K.; Shi, L.; Chen, F.; Waye, M.M.-Y.; Lim, C.K.; Cheng, P.-W.; Luk, S.S.; Mok, V.C.T.; Chu, W.C.W.; Wang, D. Altered topological organization of brain structural network in Chinese children with developmental dyslexia. Neurosci. Lett. 2015, 589, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Hagmann, P.; Sporns, O.; Madan, N.; Cammoun, L.; Pienaar, R.; Wedeen, V.J.; Meuli, R.; Thiran, J.P.; Grant, P.E. White matter maturation reshapes structural connectivity in the late developing human brain. Proc. Natl. Acad. Sci. USA 2010, 107, 19067–19072. [Google Scholar] [CrossRef]
- Vourkas, M.; Micheloyannis, S.; Simos, P.G.; Rezaie, R.; Fletcher, J.M.; Cirino, P.T.; Papanicolaou, A.C. Dynamic task-specific brain network connectivity in children with severe reading difficulties. Neurosci. Lett. 2011, 488, 123–128. [Google Scholar] [CrossRef]
- Chan, M.Y.; Park, D.C.; Savalia, N.K.; Petersen, S.E.; Wig, G.S. Decreased segregation of brain systems across the healthy adult lifespan. Proc. Natl. Acad. Sci. USA 2014, 111, E4997–E5006. [Google Scholar] [CrossRef]
- Nicholson, A. (Ed.) Brain Health Across the Life Span: Proceedings of a Workshop. National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Board on Population Health and Public Health Practice; National Academies Press: Washington, DC, USA, 2020. [Google Scholar]
- Bassett, D.S.; Wymbs, N.F.; Rombach, M.P.; Porter, M.A.; Mucha, P.J.; Grafton, S.T. Task-Based Core-Periphery Organization of Human Brain Dynamics. PLoS Comput. Biol. 2013, 9, e1003171. [Google Scholar] [CrossRef]
- Bassett, D.S.; Yang, M.; Wymbs, N.F. Grafton ST Learning-induced autonomy of sensorimotor systems. Nat. Neurosci. 2015, 18, 744–751. [Google Scholar] [CrossRef]
- Levy, J.; Pernet, C.; Treserras, S.; Boulanouar, K.; Aubry, F.; Démonet, J.F.; Celsis, P. Testing for the Dual-Route Cascade Reading Model in the Brain: An fMRI Effective Connectivity Account of an Efficient Reading Style. PLoS ONE 2009, 4, e6675. [Google Scholar] [CrossRef]
- Stoodley, C.J.; Stein, J.F. Cerebellar Function in Developmental Dyslexia. Cerebellum 2012, 12, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Kujala, J.; Pammer, K.; Cornelissen, P.; Roebroeck, A.; Formisano, E.; Salmelin, R. Phase coupling in a cerebro-cerebellar network at 8–13 hz during reading. Cerebral Cortex 2007, 17, 1476–1485. [Google Scholar] [CrossRef]
- Richlan, F. Developmental dyslexia: Dysfunction of a left hemisphere reading network. Front. Hum. Neurosci. 2012, 6, 120. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.C.; Church, J.A.; Power, J.D.; Miezin, F.M.; Petersen, S.E.; Schlaggar, B.L. Functional network architecture of reading-related regions across development. Brain Lang. 2013, 125, 231–243. [Google Scholar] [CrossRef]
- Murdaugh, D.L.; Maximo, J.O.; Kana, R.K. Changes in intrinsic connectivity of the brain’s reading network following intervention in children with autism. Hum Brain Mapp. 2015, 36, 2965–2979. [Google Scholar] [CrossRef] [PubMed]
- Richlan, F.; Kronbichler, M.; Wimmer, H. Functional abnormalities in the dyslexic brain: A quantitative meta-analysis of neuroimaging studies. Hum. Brain Mapp. 2009, 30, 3299–3308. [Google Scholar] [CrossRef] [PubMed]
- Schulz, E.; Maurer, U.; van der Mark, S.; Bucher, K.; Brem, S.; Martin, E.; Brandeis, D. Impaired semantic processing during sentence reading inchildren with dyslexia: Combined fMRI and ERP evidence. Neuroimage 2008, 41, 153–168. [Google Scholar] [CrossRef]
- Mahe, G.; Pont, C.; Zesiger, P.; Laganaro, M. The electrophysiological correlates of developmental dyslexia: New insights from lexical decision and reading aloud in adults. Neuropsychology 2018, 121, 19–27. [Google Scholar] [CrossRef]
- Maurer, U.; Brem, S.; Kranz, F.; Bucher, K.; Benz, R.; Halder, P.; Steinhausen, H.-C.; Brandeis, D. Coarse neural tuning for print peaks when children learn to read. NeuroImage 2006, 33, 749–758. [Google Scholar] [CrossRef]
- Horowitz-Kraus, T.; Hutton, J.S. Brain connectivity in children is increased by the time they spend reading books and decreased by the length of exposure to screen-based media. Acta Paediatr. 2017, 107, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Pugh, K.R.; Landi, N.; Preston, J.L.; Mencl, W.E.; Austin, A.C.; Sibley, D.; Fulbright, R.K.; Seidenberg, M.S.; Grigorenko, E.L.; Constable, R.T.; et al. The relationship between phonological and auditory processing and brain organization in beginning readers. Brain Lang. 2013, 125, 173–183. [Google Scholar] [CrossRef]
- Bohland, J.W.; Tourville, J.A.; Guenther, F.H. Neural bases of speech production. In The Routledge Handbook of Phonetics; Routledge: London, UK, 2019; pp. 126–163. [Google Scholar]
- Friederici, A.D. The neural basis for human syntax: Broca’s area and beyond. Curr. Opin. Behav. Sci. 2018, 21, 88–92. [Google Scholar] [CrossRef]
- Vandermosten, M.; Boets, B.; Wouters, J.; Ghesquière, P. A qualitative and quantitative review of diffusion tensor imaging studies in reading and dyslexia. Neurosci. Biobehav. Rev. 2012, 36, 1532–1552. [Google Scholar] [CrossRef]
- Dehaene, S.; Cohen, L. The unique role of the visual word form area in reading. Trends Cogn. Sci. 2011, 15, 254–262. [Google Scholar] [CrossRef]
- Behrmann, M.; Plaut, D.C. A vision of graded hemispheric specialization. Ann. N. Y. Acad. Sci. 2015, 1359, 30–46. [Google Scholar] [CrossRef]
- Kershner, J.R. Neuroscience and education: Cerebral lateralization of networks and oscillations in dyslexia. Laterality 2019, 25, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Weems, S.A.; Zaidel, E. The relationship between reading ability and lateralized lexical decision. Brain Cogn. 2004, 55, 507–515. [Google Scholar] [CrossRef]
- Brownell, H. Chapter 10—Right Hemisphere Contributions to Understanding Lexical Connotation and Metaphor. In Language and the Brain, Representation and Processing, Foundations of Neuropsychology; Academic Press: Cambridge, MA, USA, 2000; pp. 185–201. [Google Scholar]
- Schlaggar, B.L.; McCandliss, B.D. Development of Neural Systems for Reading. Annu. Rev. Neurosci. 2007, 30, 475–503. [Google Scholar] [CrossRef]
- Dehaene-Lambertz, G.; Monzalvo, K.; Dehaene, S. The emergence of the visual word form: Longitudinal evolution of category-specific ventral visual areas during reading acquisition. PLoS Biol. 2018, 16, e2004103. [Google Scholar] [CrossRef]
- Lallier, M.; Tainturier, M.-J.; Dering, B.; Donnadieu, S.; Valdois, S.; Thierry, G. Behavioral and ERP evidence for amodal sluggish attentional shifting in developmental dyslexia. Neuropsychology 2010, 48, 4125–4135. [Google Scholar] [CrossRef]
- Harrar, V.; Tammam, J.; Pérez-Bellido, A.; Pitt, A.; Stein, J.; Spence, C. Multisensory Integration and Attention in Developmental Dyslexia. Curr. Biol. 2014, 24, 531–535. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Metrics | Controls | Pre-Training Dys | Post-Training Dys | Con/Pre-Training Dys | Con/Post-Training Dys | Pre-/Post-Training Dys | |||
| Mean ± s.e. | Mean ± s.e. | Mean ± s.e. | p | χ2 | p | χ2 | p | χ2 | |
| δ | |||||||||
| D | 0.263 ± 0.006 | 0.271 ± 0.003 | 0.276 ± 0.007 | 0.174 | 0.184 | 0.105 | 2.62 | 0.53 | 0.37 |
| LF | 0.618 ± 0.009 | 0.623 ± 0.005 | 0.608 ± 0.011 | 0.945 | 0.004 | 0.451 | 0.56 | 0.38 | 0.76 |
| TH | 0.413 ± 0.006 | 0.427 ± 0.003 | 0.419 ± 0.008 | 0.034 | 4.476 | 0.204 | 1.63 | 0.50 | 0.45 |
| K | 3.934 ± 0.133 | 4.004 ± 0.099 | 3.798 ± 0.145 | 0.968 | 0.001 | 0.379 | 0.72 | 0.23 | 1.32 |
| θ | |||||||||
| D | 0.337 ± 0.004 | 0.311 ± 0.003 | 0.317 ± 0.004 | <0.0001 | 21.22 | 0.0025 | 9.12 | 0.17 | 1.85 |
| LF | 0.531 ± 0.004 | 0.569 ± 0.004 | 0.555 ± 0.005 | <0.0001 | 32.26 | 0.0014 | 10.14 | 0.067 | 3.34 |
| TH | 0.394 ± 0.004 | 0.409 ± 0.003 | 0.404 ± 0.004 | 0.004 | 8.13 | 0.036 | 4.36 | 0.60 | 0.27 |
| K | 3.019 ± 0.042 | 3.321 ± 0.041 | 3.195 ± 0.044 | <0.0001 | 32.43 | 0.002 | 9.48 | 0.058 | 3.58 |
| α | |||||||||
| D | 0.332 ± 0.003 | 0.316 ± 0.003 | 0.324 ± 0.004 | 0.001 | 10.35 | 0.088 | 2.89 | 0.27 | 1.16 |
| LF | 0.523 ± 0.004 | 0.552 ± 0.003 | 0.538 ± 0.004 | <0.0001 | 26.34 | 0.035 | 4.42 | 0.011 | 6.44 |
| TH | 0.391 ± 0.003 | 0.402 ± 0.003 | 0.398 ± 0.003 | 0.026 | 4.93 | 0.275 | 1.18 | 0.333 | 0.93 |
| K | 2.943 ± 0.025 | 3.155 ± 0.031 | 3.041 ± 0.034 | <0.0001 | 21.40 | 0.077 | 3.12 | 0.017 | 5.66 |
| β1 | |||||||||
| D | 0.333 ± 0.003 | 0.318 ± 0.002 | 0.333 ± 0.004 | 0.0002 | 14.13 | 0.762 | 0.09 | 0.0012 | 10.4 |
| LF | 0.507 ± 0.003 | 0.539 ± 0.003 | 0.519 ± 0.004 | <0.0001 | 41.57 | 0.044 | 4.03 | <0.0001 | 16.11 |
| TH | 0.374 ± 0.003 | 0.393 ± 0.003 | 0.387 ± 0.003 | <0.0001 | 19.97 | 0.011 | 6.41 | 0.11 | 2.51 |
| K | 2.854 ± 0.021 | 3.056 ± 0.021 | 2.925 ± 0.023 | <0.0001 | 46.11 | 0.024 | 5.08 | <0.0001 | 15.84 |
| β2 | |||||||||
| D | 0.331 ± 0.003 | 0.315 ± 0.002 | 0.328 ± 0.003 | 0.0006 | 11.68 | 0.591 | 0.28 | 0.006 | 7.48 |
| LF | 0.506 ± 0.003 | 0.542 ± 0.003 | 0.517 ± 0.004 | <0.0001 | 47.08 | 0.057 | 3.60 | <0.0001 | 20.11 |
| TH | 0.377 ± 0.003 | 0.396 ± 0.003 | 0.385 ± 0.003 | <0.0001 | 20.20 | 0.061 | 3.50 | 0.0238 | 5.01 |
| K | 2.829 ± 0.019 | 3.068 ± 0.024 | 2.898 ± 0.022 | <0.0001 | 59.41 | 0.0238 | 5.10 | <0.0001 | 24.22 |
| γ1 | |||||||||
| D | 0.329 ± 0.003 | 0.301 ± 0.003 | 0.315 ± 0.003 | <0.0001 | 41.11 | 0.003 | 8.04 | 0.0025 | 9.07 |
| LF | 0.517 ± 0.004 | 0.566 ± 0.004 | 0.529 ± 0.004 | <0.0001 | 70.91 | 0.085 | 2.95 | <0.0001 | 39.51 |
| TH | 0.384 ± 0.003 | 0.409 ± 0.003 | 0.387 ± 0.003 | <0.0001 | 29.44 | 0.635 | 0.22 | <0.0001 | 22.65 |
| K | 2.866 ± 0.019 | 3.263 ± 0.033 | 2.958 ± 0.027 | <0.0001 | 89.93 | 0.020 | 5.33 | <0.0001 | 46.60 |
| γ2 | |||||||||
| D | 0.280 ± 0.002 | 0.262 ± 0.003 | 0.267 ± 0.003 | <0.0001 | 24.28 | 0.001 | 10.62 | 0.188 | 1.72 |
| LF | 0.596 ± 0.003 | 0.636 ± 0.003 | 0.623 ± 0.004 | <0.0001 | 60.79 | <0.0001 | 29.80 | 0.055 | 3.66 |
| TH | 0.422 ± 0.003 | 0.441 ± 0.002 | 0.434 ± 0.003 | <0.0001 | 18.32 | 0.009 | 6.72 | 0.207 | 1.59 |
| K | 3.427 ± 0.037 | 3.903 ± 0.048 | 3.708 ± 0.053 | <0.0001 | 56.67 | <0.0001 | 24.11 | 0.037 | 4.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taskov, T.; Dushanova, J. Reading-Network in Developmental Dyslexia before and after Visual Training. Symmetry 2020, 12, 1842. https://doi.org/10.3390/sym12111842
Taskov T, Dushanova J. Reading-Network in Developmental Dyslexia before and after Visual Training. Symmetry. 2020; 12(11):1842. https://doi.org/10.3390/sym12111842
Chicago/Turabian StyleTaskov, Tihomir, and Juliana Dushanova. 2020. "Reading-Network in Developmental Dyslexia before and after Visual Training" Symmetry 12, no. 11: 1842. https://doi.org/10.3390/sym12111842
APA StyleTaskov, T., & Dushanova, J. (2020). Reading-Network in Developmental Dyslexia before and after Visual Training. Symmetry, 12(11), 1842. https://doi.org/10.3390/sym12111842