Four Decades of Land-Cover Change on the Kenai Peninsula, Alaska: Detecting Disturbance-Influenced Vegetation Shifts Using Landsat Legacy Data

 ,
,
Abstract
1. Introduction
2. Materials and Methods
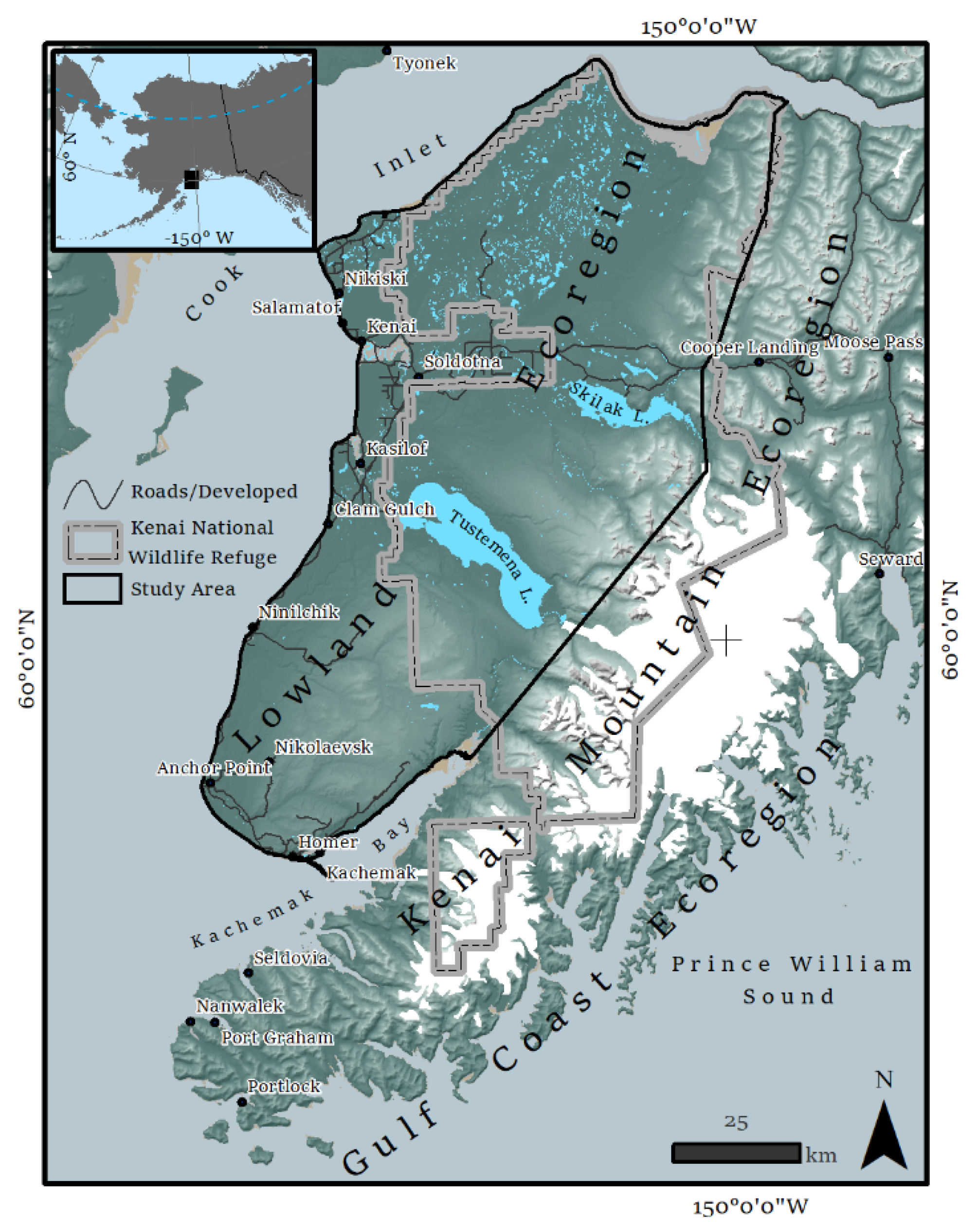
2.1. Study Area
2.2. Approach
2.3. Vegetation Classification
2.4. Image Acquisition
2.5. Model Development
2.6. Land-Cover Assessment and Change Detection
3. Results
3.1. Classification Accuracy
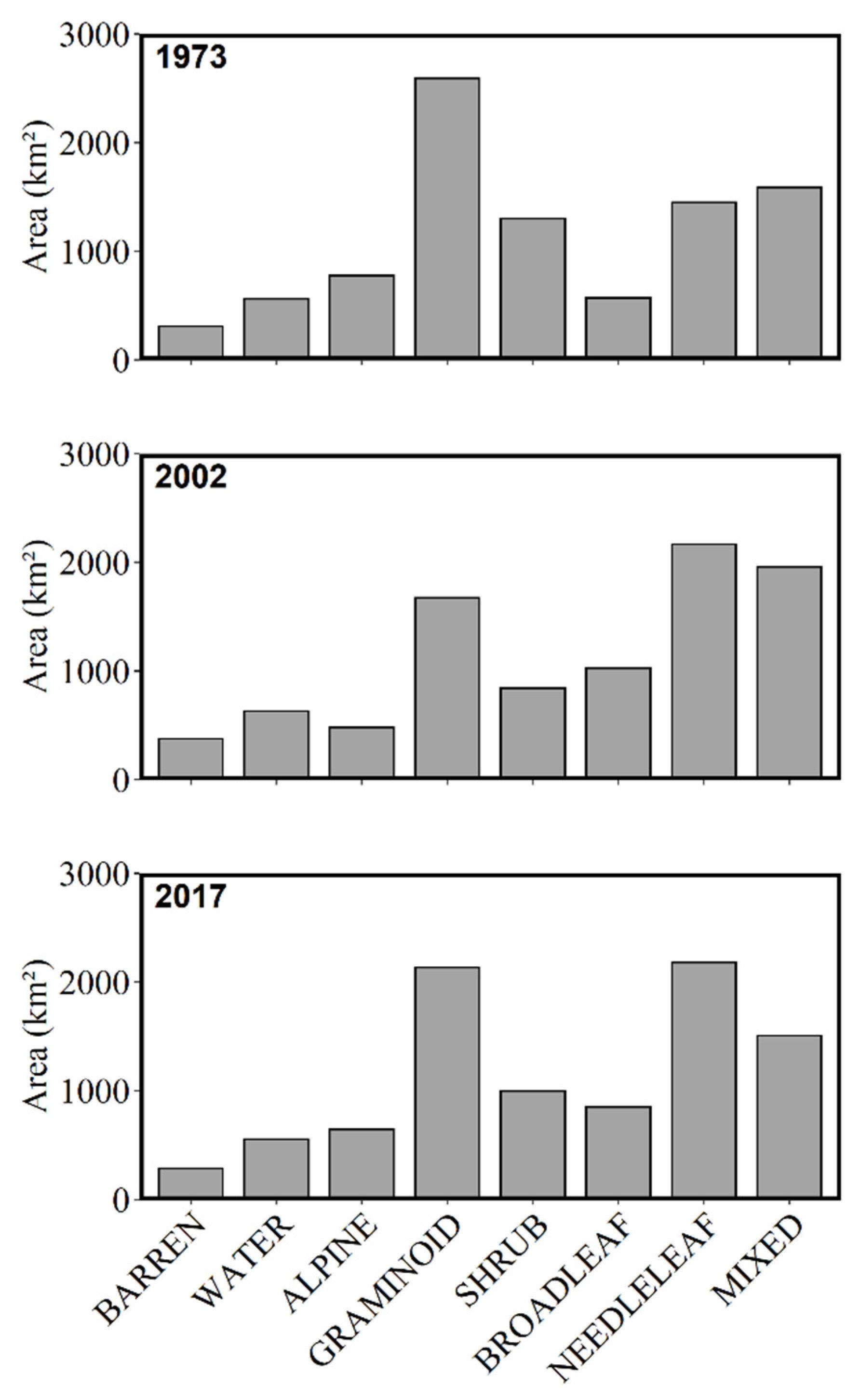
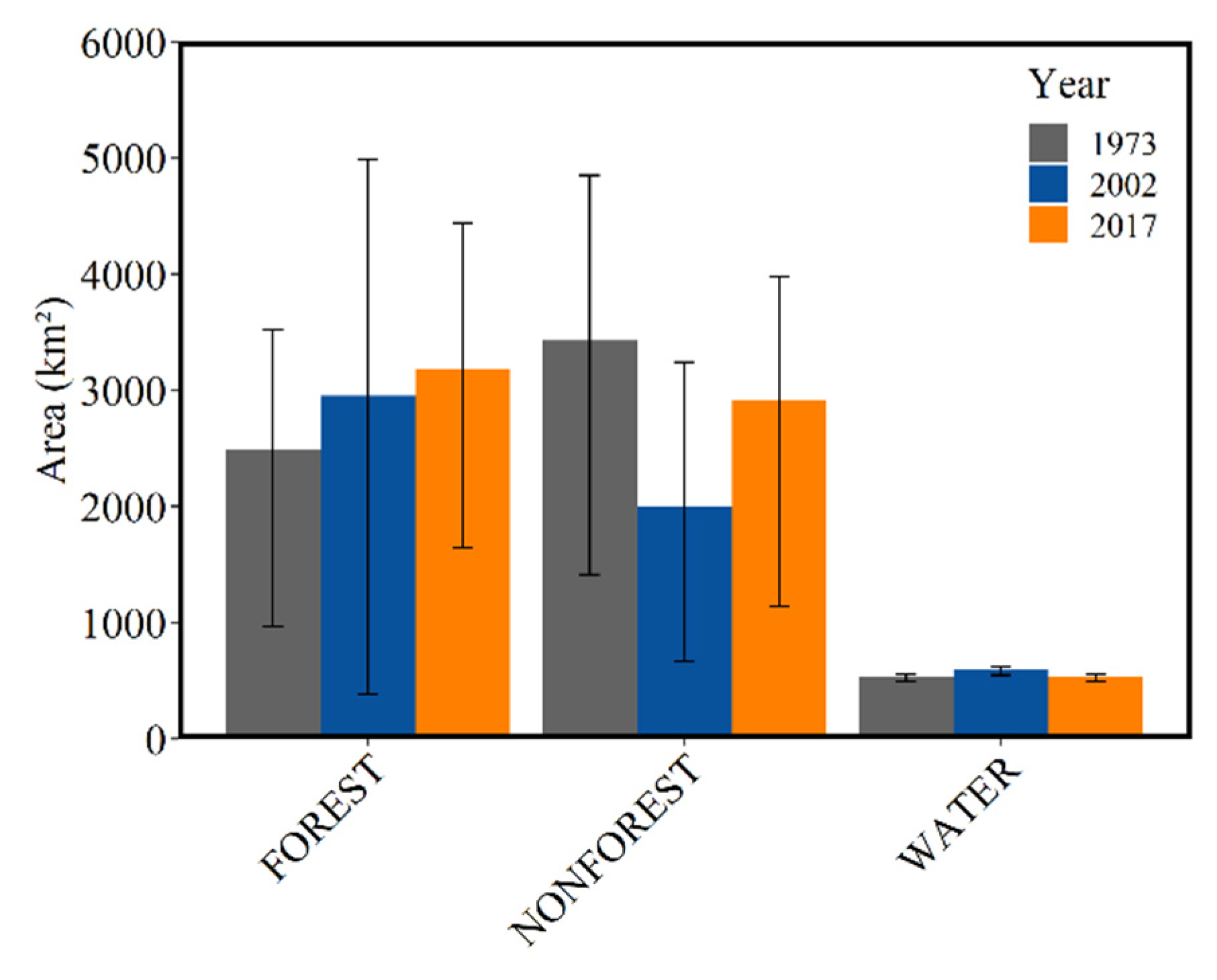
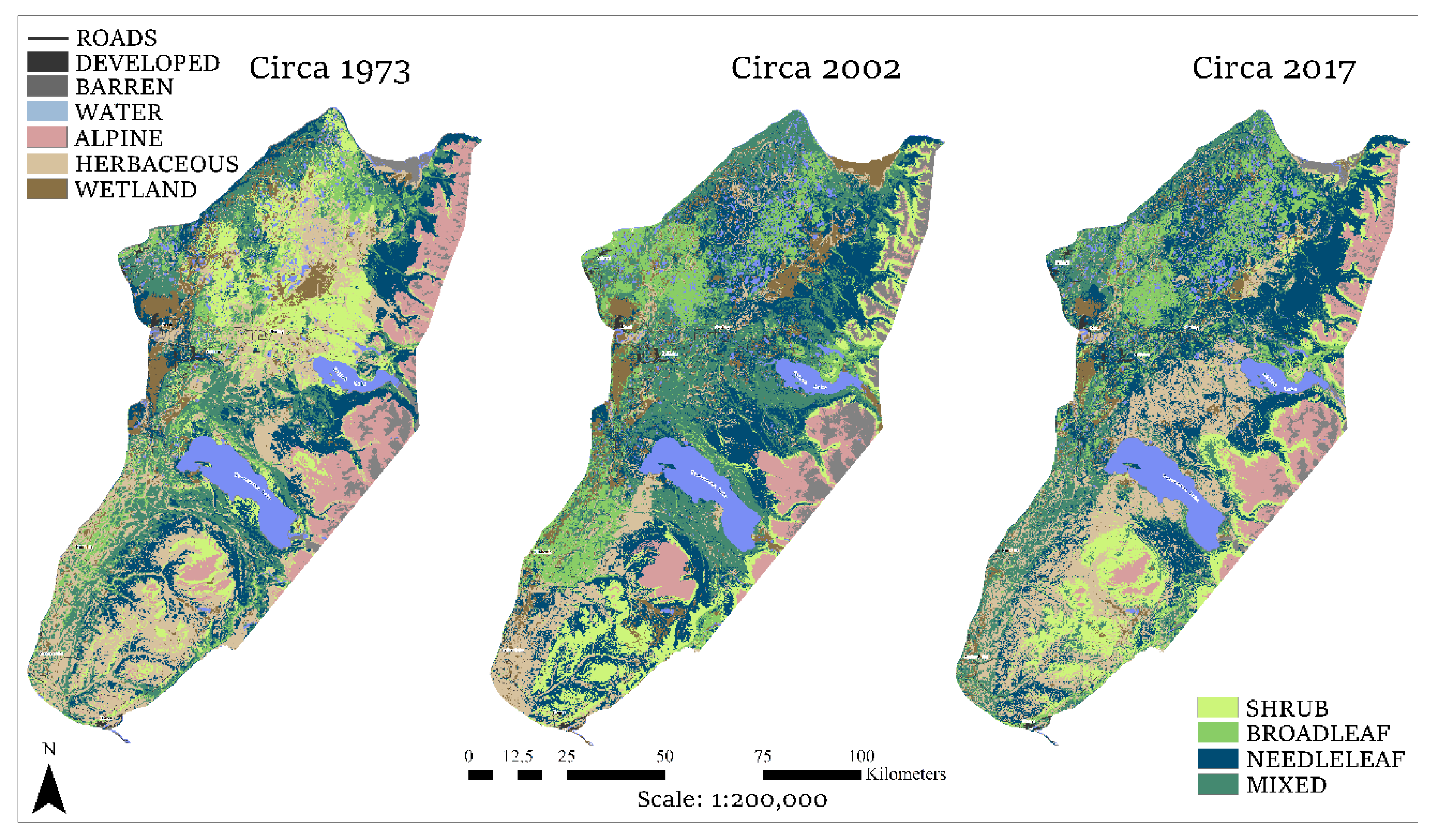
3.2. Land Cover by Era
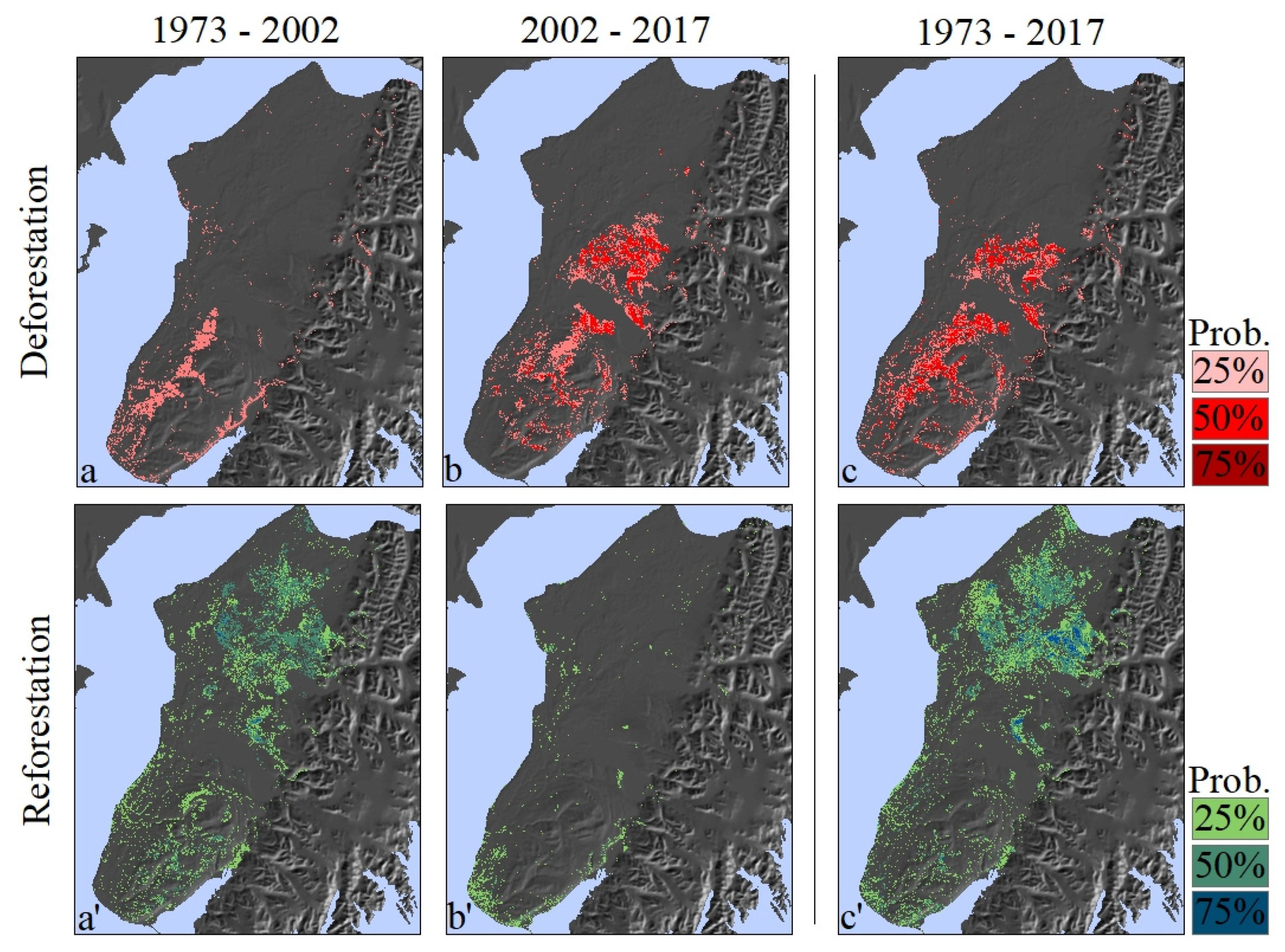
3.3. Change Detection: Deforestation
3.3.1. 1973–2002
3.3.2. 2002–2017
3.3.3. 1973–2017
3.4. Change Detection: Reforestation
3.4.1. 1973–2002
3.4.2. 2002–2017
3.4.3. 1973–2017
4. Discussion
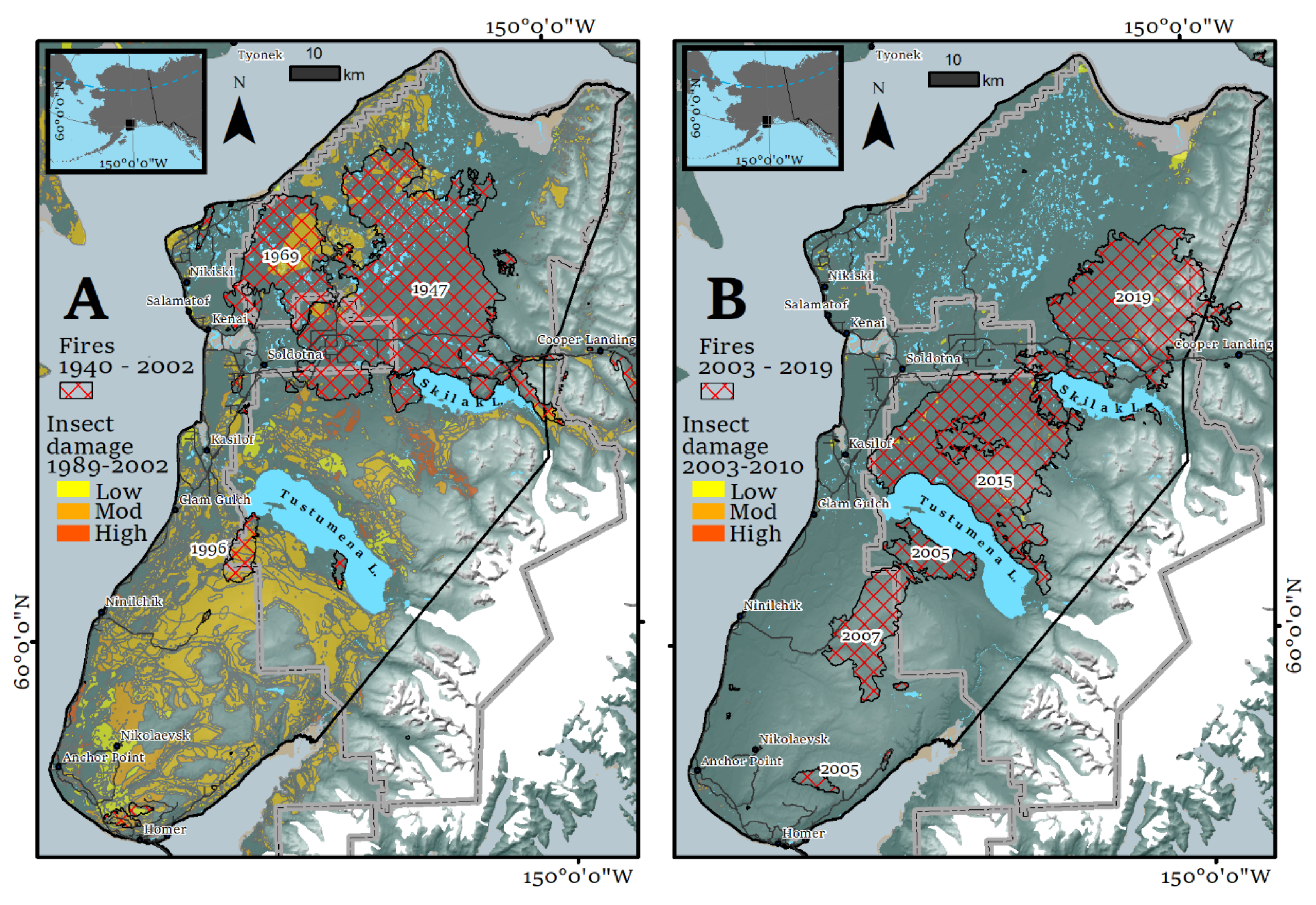
4.1. Four Decades of Land-Cover change on the Kenai Peninsula
4.2. Assumptions and Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Supporting Data
References
- Spence, J.R. The new boreal forestry: Adjusting timber management to accommodate biodiversity. Trends Ecol. Evol. 2001, 14, 591–593. [Google Scholar] [CrossRef]
- Walker, X.J.; Baltzer, J.L.; Cumming, S.G.; Day, N.J.; Ebert, C.; Goetz, S.; Johnstone, J.F.; Potter, S.; Rogers, B.M.; Schuur, E.A.G.; et al. Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature 2019, 572, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.P.; Flannigan, M.D.; Maynard, D.G.; Thompson, I.D.; Volney, W.I.A. An introduction to Canada’s boreal zone: Ecosystem processes, health, sustainability, and environmental issues. Environ. Rev. 2013, 21, 207–226. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Warkentin, I.G. Global estimates of boreal forest carbon stocks and flux. Glob. Planet. Chang. 2015, 128, 24–30. [Google Scholar] [CrossRef]
- Gerardo, M.; Katila, P.; Galloway, G.; Alfaro, R.I.; Markku, K.; Lobovikov, M.; Varjo, J. Forests and Society—Responding to Global Drivers of Change; IUFRO World Series; IUFRO: Vienna, Austria, 2010; Volume 25, ISBN 978-3-901347-93-1. [Google Scholar]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shividenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef]
- Yarie, J.; Billings, S. Carbon balance of the taiga forest within Alaska: Present and future. Can. J. Res. 2002, 32, 757–767. [Google Scholar] [CrossRef]
- Hayes, D.J.; McGuire, A.D.; Kicklighter, D.W.; Gurney, K.R.; Burnside, T.J.; Melillo, J.M. Is the northern high-latitude land-based CO2 sink weakening? Glob. Biogeochem. Cycles 2011, 25, 3. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Woodcock, C.E.; Friedl, M.A. Canadian boreal forest greening and browning trends: An analysis of biogeographic patterns and the relative roles of disturbance versus climate drivers. Environ. Res. Lett. 2018, 13, 014007. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Fastie, C.L. Recent changes in treeline forest distribution and structure in interior Alaska. Écoscience 2003, 10, 176–185. [Google Scholar] [CrossRef]
- Petersen, B.; Stuart, D. Explanations of a changing landscape: A critical examination of the British Columbia bark beetle epidemic. Environ. Plan. Econ. Space 2013, 46, 598–613. [Google Scholar] [CrossRef]
- Peng, C.; Ma, Z.; Lei, X.; Zhu, Q.; Chen, H.; Wang, W.; Liu, S.; Li, W.; Fang, X.; Zhou, X. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Soja, A.J.; Tchebakova, N.M.; French, N.H.F.; Flannigan, M.D.; Shugart, H.H.; Stocks, B.J.; Sukhinin, A.I.; Parfenova, E.I.; Chapin, F.S., III; Stackhouse, P.W., Jr. Climate-induced boreal forest change: Predictions versus current observations. Glob. Planet. Chang. 2007, 56, 274–296. [Google Scholar] [CrossRef]
- Kelly, R.; Chipman, M.L.; Higuera, P.E.; Stefanova, I.; Brubaker, L.B.; Hu, F.S. Recent burning of boreal forests exceeds fire regime limits of the past 10,000 years. Proc. Natl. Acad. Sci. USA 2013, 110, 13055–13060. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar] [CrossRef]
- Schaphoff, S.; Reyer, C.P.O.; Schepaschenko, D.; Gerten, D.; Shvidenko, A. Tamm review: Observed and projected climate change impacts on Russia’s forests and its carbon balance. Ecol. Manag. 2016, 361, 432–444. [Google Scholar] [CrossRef]
- Young, A.M.; Higuera, P.E.; Duffy, P.A.; Hu, F.S. Climatic thresholds shape northern high-latitude fire regimes and imply vulnerability to future climate change. Ecography 2017, 40, 606–617. [Google Scholar] [CrossRef]
- Safranyik, L.; Carroll, A.L.; Regniere, J.; Langor, D.W.; Riel, W.G.; Shore, T.L.; Peter, B.; Cooke, B.J.; Nealis, V.G.; Taylor, S.W. Potential for range expansion of mountain pine beetle into the boreal forest of north America. Can. Entomol. 2010, 142, 415–442. [Google Scholar] [CrossRef]
- Anielski, M.; Wilson, S. Counting Canada’s Natural Capital: Assessing the Real Value of Canada’s Boreal Ecosystems; The Pembina Institute and the Canadian Boreal Initiative: Ottawa, ON, Canada, 2009. [Google Scholar]
- Berg, E.E.; Hillman, K.M.; Dial, R.; DeRuwe, A. Recent woody invasion of wetlands on the Kenai Peninsula Lowlands, south-central Alaska: A major regime shift after 18 000 years of wet Sphagnum–sedge peat recruitment. Can. J. Res. 2009, 39, 2033–2046. [Google Scholar] [CrossRef]
- Boucher, T.V.; Mead, B.R. Vegetation change and forest regeneration on the Kenai Peninsula, Alaska following a spruce beetle outbreak, 1987–2000. Ecol. Manag. 2006, 227, 233–246. [Google Scholar] [CrossRef]
- Klein, E.; Berg, E.E.; Dial, R. Wetland drying and succession across the Kenai Peninsula Lowlands, south-central Alaska. Can. J. Res. 2005, 35, 1931–1941. [Google Scholar] [CrossRef]
- Berg, E.E.; David Henry, J.; Fastie, C.L.; De Volder, A.D.; Matsuoka, S.M. Spruce beetle outbreaks on the Kenai Peninsula, Alaska, and Kluane National Park and reserve, Yukon territory: Relationship to summer temperatures and regional differences in disturbance regimes. Ecol. Manag. 2006, 227, 219–232. [Google Scholar] [CrossRef]
- Berg, E.E.; Anderson, R.S. Fire history of white and Lutz spruce forests on the Kenai Peninsula, Alaska, over the last two millennia as determined from soil charcoal. Ecol. Manag. 2006, 227, 275–283. [Google Scholar] [CrossRef]
- Jain, T.; Barrett, T.M. The western Kenai Peninsula: An opportunity to study fire and its effects on soils and trees. In Forests of Southeast and South-Central Alaska, 2004–2008: Five-Year Forest Inventory and Analysis Report; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2011; pp. 70–74. [Google Scholar]
- Jones, B.M.; Baughman, C.A.; Romanovsky, V.E.; Parsekian, A.D.; Jones, M.C.; Grosse, G.; Berg, E.E. The presence and degradation of residual permafrost plateaus on the western Kenai Peninsula Lowlands, southcentral Alaska. Cryosphere 2016, 10, 2673–2692. [Google Scholar] [CrossRef]
- Lowell, E.C.; Willits, S.A. Lumber recovery from beetle-killed spruce trees, Kenai Peninsula, Alaska. West. J. Appl. 1998, 13, 54–59. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Chapin, F.S., III; Hollingsworth, T.N.; Mack, M.C.; Romanovsky, V.E.; Turetsky, M.R. Fire, climate change, and forest resilience in interior Alaska. Can. J. Res. 2010, 40, 1302–1312. [Google Scholar] [CrossRef]
- USDA. Chugach National Forest Land Management Plan. R10-MB. 828c; United States Department of Agriculture, Forest Service, Chugach National Forest: Anchorage, AL, USA, 2019.
- Oliver, S.G. Kenai Peninsula Tourism Industry Expected To Grow In 2017. Available online: https://www.kbbi.org (accessed on 6 June 2018).
- King, R.B. Land cover mapping principles: A return to interpretation fundamentals. Int. J. Remote Sens. 2010, 3525–3545. [Google Scholar] [CrossRef]
- Wallace, C.S.A.; Webb, R.H.; Thomas, K.A. Estimation of perennial vegetation cover distribution in the mojave desert using MODIS-EVI data. Giscience Remote Sens. 2008, 45, 167–187. [Google Scholar] [CrossRef]
- Jones, M.C.; Peteet, D.M.; Kurdyla, D.; Guilderson, T. Climate and vegetation history from a 14,000-year peatland record, Kenai Peninsula, Alaska. Quat. Res. 2009, 72, 207–217. [Google Scholar] [CrossRef]
- Allen, L.H. One Hundred Centuries of Native Life on the Kenai Peninsula; Kenai Peninsula Historical Association: Soldotna, AK, USA, 2002. [Google Scholar]
- Bangs, E.; Spraker, T.; Bailey, T.; Berns, V. Effects of Increased Human Populations on Wildlife Resources of the Kenai Peninsula, Alaska; US Fish & Wildlife Publications: Lincoln, NE, USA, 1982.
- Goodman, L.F.; Hungate, B.A. Managing forests infested by spruce beetles in south-central Alaska: Effects on nitrogen availability, understory biomass, and spruce regeneration. Ecol. Manag. 2006, 227, 267–274. [Google Scholar] [CrossRef]
- Anderson, R.S.; Hallett, D.J.; Berg, E.; Jass, R.B.; Toney, J.L.; de Fontaine, C.S.; DeVolder, A. Holocene development of Boreal forests and fire regimes on the Kenai Lowlands of Alaska. Holocene 2006, 16, 791–803. [Google Scholar] [CrossRef]
- Hrobak, J.; Schmunk, G. The Alaska Large Fire Database [Data set]. 2019. Available online: https://www.frames.gov/catalog/10465 (accessed on 6 June 2018).
- Werner, R.A.; Holsten, E.H.; Matsuoka, S.M.; Burnside, R.E. Spruce beetles and forest ecosystems in south-central Alaska: A review of 30 years of research. Ecol. Manag. 2006, 227, 195–206. [Google Scholar] [CrossRef]
- Yang, L.; Jin, S.; Danielson, P.; Homer, C.; Gass, L.; Bender, S.M.; Case, A.; Costello, C.; Dewitz, J.; Fry, J.; et al. A new generation of the United States National land cover database: Requirements, research priorities, design, and implementation strategies. ISPRS J. Photogramm. Remote Sens. 2018, 146, 108–123. [Google Scholar] [CrossRef]
- ArcMap; Environmental Systems Research Institute (ESRI): Redlands, CA, USA, 2019.
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Wulder, M.A.; Franklin, S.E. Remote Sensing of Forest Environments: Concepts and Case Studies; Kluwer Academic Publishers: Boston, MA, USA, 2003. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Alaska 2 Arc-second Digital Elevation Models (DEMs)—USGS National Map 3DEP Downloadable Data Collection [Data Set]. 2017. Available online: https://catalog.data.gov/dataset/national-elevation-dataset-ned-alaska-2-arc-second-downloadable-data-collection-national-geosp (accessed on 6 June 2018).
- Kenai Peninsula Vegetation Map. Available online: https://usfs.maps.arcgis.com/apps/MapSeries/index.html?appid=4e21c25d5eac421babaef3222004cccf (accessed on 6 June 2018).
- Breiman, L. Random Forests. Mach. Leaning 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Baughman, C.A.; Loehman, R.A.; Saperstein, L. Land cover estimates for the Kenai Peninsula lowlands; 1973, 2002, and 2017. US Geol. Surv. Data Release 2020. [Google Scholar] [CrossRef]
- Jones, B.M. Land-cover change on the southern Kenai Peninsula lowlands, Alaska using USGS land cover trends methodology. J. Geogr. Reg. Plan. 2008, 1, 68–71. [Google Scholar]
- Dial, R.; Berg, E.; Timm, K.; McMahon, A.; Geck, J. Changes in the alpine forest-tundra ecotone commensurate with recent warming in southcentral Alaska: Evidence from orthophotos and field plots. J. Geophys. Res. 2007, 112, G4. [Google Scholar] [CrossRef]
- Flint, C.G. Community perspectives on spruce beetle impacts on the Kenai Peninsula, Alaska. Ecol. Manag. 2006, 227, 207–218. [Google Scholar] [CrossRef]
- Reynolds, K.M.; Hard, J.S. Risk and hazard of spruce beetle attack in unmanaged stands on the Kenai Peninsula, Alaska, under epidemic conditions. Ecol. Manag. 1991, 43, 137–151. [Google Scholar] [CrossRef]
- De Volder, A. Fire and Climate History of Lowland Black Spruce Forests, Kenai National Wildlife Refuge, Alaska. Master’s Thesis, Northern Arizona University, Flagstaff, AZ, USA, 1999. [Google Scholar]
- DeLong, H.B.; Lieffers, V.J.; Blenis, P.V. Microsite effects on first-year establishment and overwinter survival of white spruce in aspen-dominated boreal mixedwoods. Can. J. Res. 1997, 27, 1452–1457. [Google Scholar] [CrossRef]
- Zasada, J.C.; Norum, R.A.; Veldhuizen, R.M.; Teutsch, C. Artifical regeneration of trees and tall shrubs in experimentally burned upland black spruce/feather moss stands in Alaska. Can. J. Res. 1983, 13, 903–913. [Google Scholar] [CrossRef]
- von Ow, F.; Joyce, P.; Keane, M. Factors affecting the establishment of natural regeneration of Sitka spruce (Picea sitchensis (Bong.) Carr.) in Ireland. Ir. For. 1996, 53, 2–18. [Google Scholar]
- Viereck, L.A. Wildfire in the taiga of Alaska. J. Quat. Res. 1973, 3, 465–495. [Google Scholar] [CrossRef]
- Carter, T.C.; Chapin, F.S., III. Differential effects of competition or microenvironment on boreal tree seedling establishment after fire. Ecology 2000, 84, 1086–1099. [Google Scholar] [CrossRef]
- Fastie, C.L.; Lloyd, A.H.; Doak, P. Fire history and post-fire forest development in an upland watershed of interior Alaska. J. Geophys. Res. 2002, 108, 8150. [Google Scholar] [CrossRef]
- Boggs, K.; Sturdy, M.; Rinella, D.J.; Rinella, M.J. White spruce regeneration following a major spruce beetle outbreak in forests on the Kenai Peninsula, Alaska. Ecol. Manag. 2008, 255, 3571–3579. [Google Scholar] [CrossRef]
- Kasischke, E.S.; Verbyla, D.L.; Rupp, T.S.; McGuire, A.D.; Murphy, K.A.; Jandt, R.; Barnes, J.L.; Hoy, E.E.; Duffy, P.A.; Calef, M.; et al. Alaska’s changing fire regime—Implications for the vulnerability of its boreal forests. Can. J. Res. 2010, 40, 1313–1324. [Google Scholar] [CrossRef]
- Serreze, M.C.; Barry, R.G. Processes and impacts of Arctic amplification: A research synthesis. Glob. Planet. Chang. 2011, 77, 85–96. [Google Scholar] [CrossRef]
- Sherriff, R.L.; Berg, E.E.; Miller, A.E. Climate variability and spruce beetle (Dendroctonus rufipennis) outbreaks in south-central and southwest Alaska. Ecology 2011, 92, 1459–1470. [Google Scholar] [CrossRef]
- Fauria, M.M.; Johnson, E.A. Climate and wildfires in the North American boreal forest. Philos. Trans. R. Soc. B 2008, 363, 2317–2329. [Google Scholar] [CrossRef]
- Hess, K.A.; Cullen, C.; Cobian-Iniguez, J.; Ramthun, J.S.; Lenske, V.; Magness, D.R.; Bolten, J.D.; Foster, A.C.; Spruce, J. Satellite-based assessment of grassland conversion and related fire disturbance in the Kenai Peninsula, Alaska. Remote Sens. 2019, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Kuusinen, N.; Kolari, P.; Levula, J.; Porcar-Castell, A.; Stenberg, P.; Berninger, F. Seasonal variation in boreal pine forest albedo and effects of canopy snow on forest reflectance. Agric. Meteorol. 2012, 164, 53–60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scene Identifier | Landsat | Sensor | Tier | Date | Band/Index | Season |
|---|---|---|---|---|---|---|
| LM10740171973229AAA05 | 1 | MSS | T2 | 8/17/1973 | TVI | Summer |
| LM10740181973229AAA05 | 1 | MSS | T2 | 8/17/1973 | TVI | Summer |
| LM10750171974063FAK04 | 1 | MSS | T2 | 3/4/1974 | B7 | Winter |
| LM10750181974063FAK04 | 1 | MSS | T2 | 3/4/1974 | B7 | Winter |
| LM10760171975041AAA02 | 1 | MSS | T2 | 2/10/1975 | B7 | Winter |
| LM10760181975041AAA02 | 1 | MSS | T2 | 2/10/1975 | B7 | Winter |
| LM10740181975093FAK08 | 1 | MSS | T2 | 4/3/1975 | B7 | Winter |
| LE70690172002211EDC00 | 7 | ETM+ | T1 | 7/30/2002 | EVI * | Summer |
| LE70690182002211EDC00 | 7 | ETM+ | T1 | 7/30/2002 | EVI * | Summer |
| LE70690172002051EDC00 | 7 | ETM+ | T1 | 2/20/2002 | B1 | Winter |
| LE70690182002051EDC00 | 7 | ETM+ | T1 | 2/20/2002 | B1 | Winter |
| LC80690172017084LGN00 | 8 | OLI | T1 | 8/29/2016 | EVI * | Summer |
| LC80690182017084LGN00 | 8 | OLI | T1 | 8/29/2016 | EVI * | Summer |
| LC80690172017084LGN00 | 8 | OLI | T1 | 3/25/2017 | B2 | Winter |
| LC80690182017084LGN00 | 8 | OLI | T1 | 3/25/2017 | B2 | Winter |
| Spectral Characteristics | Interpretation | Class ID |
|---|---|---|
| Intermediate EVI/TVI values in summer, low reflectance in blue or IR bands during winter | Closed canopy forest with significant conifer component concealing snow in winter | Needleleaf |
| High EVI/TVI values in summer, low reflectance in blue or IR bands during winter | Closed canopy forest with significant hardwood component partially concealing snow in winter | Broadleaf |
| Intermediate EVI/TVI values in summer, low reflectance in blue or IR bands during winter | Closed canopy forest with mixed species partially concealing snow in winter | Mixed |
| Intermediate EVI/TVI values in summer, intermediate reflectance in blue or IR bands during winter | Sparser leafy vegetation standing above snow in winter | Shrub |
| Intermediate EVI/TVI values in summer, high reflectance in blue or IR bands during winter | Short mixed vegetation becoming completely snow-covered in winter | Herbaceous |
| Intermediate EVI/TVI values in summer, high reflectance in blue or IR bands during winter | Short hydrophytic vegetation becoming completely snow-covered in winter | Wetland |
| Intermediate EVI/TVI values in summer, high reflectance in blue or IR bands during winter | Short and/or sparse vegetation becoming completely snow-covered in winter | Alpine |
| Low EVI/TVI values in summer, maximum reflectance in blue or IR bands during winter | Non- to minimally vegetated surfaces becoming completely snow-covered in winter | Barren |
| Negative EVI/TVI values in summer, maximum reflectance in blue or IR bands during winter | Standing water with no visible vegetation becoming completely snow-covered in winter | Water |
| 1973 | Referenced Forest | Referenced Non-forest | Total | UA | Commission |
| Classified Forest | 1135 | 65 | 1200 | 94.6% | 5.4% |
| Classified Non-forest | 68 | 2332 | 2400 | 97.2% | 2.8% |
| Total | 1203 | 2397 | 3600 | ||
| PA | 94.3% | 97.3% | |||
| Omission | 5.7% | 2.7% | |||
| 2002 | Referenced Forest | Referenced Non-forest | Total | UA | Commission |
| Classified Forest | 1028 | 172 | 1200 | 85.7% | 14.3% |
| Classified Non-forest | 163 | 2237 | 2400 | 93.2% | 6.8% |
| Total | 1191 | 2409 | 3600 | ||
| PA | 86.3% | 92.9% | |||
| Omission | 13.7% | 7.1% | |||
| 2017 | Referenced Forest | Referenced Non-forest | Total | UA | Commission |
| Classified Forest | 1171 | 29 | 1200 | 97.6% | 2.4% |
| Classified Non-forest | 26 | 2374 | 2400 | 98.9% | 1.1% |
| Total | 1197 | 2403 | 3600 | ||
| PA | 97.8% | 98.8% | |||
| Omission | 2.2% | 1.2% |
| 1973 Reference Conditions | ||||||||||||
| OOB Est. of Error: | 14.5% | |||||||||||
| Classifications | Needle | Mixed | Broad | Shrub | Herb | Wet. | Alpine | Barren | Water | Total | UA | Commission |
| Needleleaf | 324 | 51 | 1 | 20 | 3 | 1 | 0 | 0 | 0 | 400 | 81.0% | 19.0% |
| Mixed | 44 | 314 | 22 | 13 | 5 | 2 | 0 | 0 | 0 | 400 | 78.5% | 21.5% |
| Broadleaf | 2 | 33 | 344 | 3 | 18 | 0 | 0 | 0 | 0 | 400 | 86.0% | 14.0% |
| Shrub | 13 | 9 | 2 | 327 | 26 | 3 | 20 | 0 | 0 | 400 | 81.8% | 18.3% |
| Herbaceous | 5 | 6 | 20 | 32 | 308 | 26 | 3 | 0 | 0 | 400 | 77.0% | 23.0% |
| Wetland | 5 | 5 | 0 | 12 | 21 | 353 | 3 | 0 | 1 | 400 | 88.3% | 11.8% |
| Alpine | 1 | 0 | 0 | 14 | 5 | 0 | 337 | 43 | 0 | 400 | 84.3% | 15.8% |
| Barren | 0 | 0 | 0 | 0 | 0 | 0 | 21 | 378 | 1 | 400 | 94.5% | 5.5% |
| Water | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 2 | 393 | 400 | 98.3% | 1.8% |
| Total | 395 | 418 | 390 | 422 | 387 | 386 | 384 | 423 | 395 | 3600 | ||
| PA | 34.5% | 15.1% | 42.9% | 18.5% | 14.4% | 30.4% | 83.6% | 15.5% | 8.9% | |||
| Omission | 18.0% | 24.9% | 11.8% | 22.5% | 20.4% | 8.5% | 12.2% | 10.6% | 0.5% | |||
| 2002 Reference Conditions | ||||||||||||
| OOB Est. of Error: | 30.67% | |||||||||||
| Classifications | Needle | Mixed | Broad | Shrub | Herb | Wet. | Alpine | Barren | Water | Total | UA | Commission |
| Needleleaf | 211 | 78 | 23 | 26 | 42 | 14 | 3 | 1 | 2 | 400 | 52.8% | 47.3% |
| Mixed | 81 | 223 | 68 | 8 | 9 | 6 | 0 | 0 | 5 | 400 | 55.8% | 44.3% |
| Broadleaf | 28 | 66 | 250 | 12 | 33 | 6 | 0 | 0 | 5 | 400 | 62.5% | 37.5% |
| Shrub | 17 | 7 | 31 | 249 | 37 | 12 | 28 | 19 | 0 | 400 | 62.3% | 37.8% |
| Herbaceous | 36 | 9 | 19 | 33 | 220 | 81 | 2 | 0 | 0 | 400 | 55.0% | 45.0% |
| Wetland | 18 | 4 | 13 | 6 | 57 | 284 | 3 | 0 | 15 | 400 | 71.0% | 29.0% |
| Alpine | 0 | 0 | 0 | 23 | 1 | 0 | 312 | 64 | 0 | 400 | 78.0% | 22.0% |
| Barren | 0 | 0 | 0 | 9 | 0 | 0 | 17 | 374 | 0 | 400 | 93.5% | 6.5% |
| Water | 5 | 2 | 2 | 0 | 0 | 18 | 0 | 0 | 373 | 400 | 93.5% | 6.8% |
| Total | 396 | 389 | 406 | 366 | 399 | 421 | 365 | 458 | 400 | 3600 | ||
| PA | 53.3% | 57.3% | 61.6% | 68.0% | 55.1% | 67.5% | 85.5% | 81.7% | 93.3% | |||
| Omission | 46.7% | 42.7% | 38.4% | 32.0% | 44.9% | 32.5% | 14.5% | 18.3% | 6.8% | |||
| 2017 Reference Conditions | ||||||||||||
| OOB Est. of Error: | 10.06% | |||||||||||
| Classifications | Needle | Mixed | Broad | Shrub | Herb | Wet. | Alpine | Barren | Water | Total | UA | Commission |
| Needleleaf | 354 | 31 | 7 | 2 | 4 | 0 | 2 | 0 | 0 | 400 | 88.2% | 11.5% |
| Mixed | 22 | 329 | 38 | 5 | 6 | 0 | 0 | 0 | 0 | 400 | 82.3% | 17.8% |
| Broadleaf | 12 | 31 | 347 | 7 | 3 | 0 | 0 | 0 | 0 | 400 | 86.8% | 13.3% |
| Shrub | 6 | 3 | 7 | 345 | 20 | 5 | 13 | 1 | 0 | 400 | 86.3% | 13.8% |
| Herbaceous | 5 | 1 | 0 | 7 | 338 | 42 | 5 | 2 | 0 | 400 | 84.5% | 15.5% |
| Wetland | 1 | 2 | 1 | 2 | 18 | 376 | 0 | 0 | 0 | 400 | 94.0% | 6.0% |
| Alpine | 0 | 0 | 0 | 7 | 0 | 0 | 367 | 26 | 0 | 400 | 91.8% | 8.3% |
| Barren | 0 | 0 | 0 | 0 | 4 | 1 | 12 | 383 | 0 | 400 | 95.8% | 4.3% |
| Water | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 399 | 400 | 99.8% | 0.3% |
| Total | 400 | 397 | 400 | 375 | 393 | 424 | 399 | 413 | 399 | 3600 | ||
| PA | 34.5% | 15.1% | 42.9% | 18.5% | 14.4% | 30.4% | 83.6% | 15.5% | 8.9% | |||
| Omission | 11.5% | 17.1% | 13.3% | 8.0% | 14.0% | 11.3% | 8.0% | 7.3% | 0.0% | |||
| 1973 | 2002 | 2017 | ||||
|---|---|---|---|---|---|---|
| % of Classified Area | km2 | % of Classified Area | km2 | % of Classified Area | km2 | |
| Alpine | 10.2% | 660 | 7.2% | 404 | 9.0% | 598 |
| Barren | 3.9% | 251 | 5.9% | 330 | 3.4% | 224 |
| Broadleaf | 6.3% | 407 | 9.9% | 558 | 9.6% | 634 |
| Herbaceous | 20.8% | 1340 | 6.3% | 356 | 17.9% | 1185 |
| Mixed | 16.5% | 1064 | 20.4% | 1148 | 13.7% | 907 |
| Needleleaf | 15.9% | 1024 | 22.2% | 1250 | 24.7% | 1642 |
| Wetland | 5.7% | 367 | 7.8% | 440 | 3.9% | 256 |
| Shrub | 12.6% | 812 | 9.9% | 558 | 9.9% | 657 |
| Water | 8.2% | 532 | 10.5% | 595 | 8.0% | 533 |
| Total | 100% | 6457 | 100% | 5638 | 100% | 6637 |
| Deforestation | ||||||||
| 1973–2002 | 2002–2017 | 1973–2017 | ||||||
| Forest to Graminoid | Forest to Shrub | Forest to Water | Forest to Graminoid | Forest to Shrub | Forest to Barren | Forest to Graminoid | Forest to Shrub | Forest to Barren |
| 182 km2 81% a | 28 km2 17% a | 2 km2 2% a | 319 km2 85% a | 55 km2 15% a | <1 km2 0.07% a | 396 km2 87% a | 58 km2 13% a | 1 km2 0.3% a |
| Reforestation | ||||||||
| 1973–2002 | 2002–2017 | 1973–2017 | ||||||
| Graminoid to Forest | Shrub to Forest | Water to Forest | Shrub to Forest | Graminoid to Forest | Alpine to Forest | Graminoid to Forest | Shrub to Forest | Alpine to Forest |
| 373 km2 54% b | 321 km2 46% b | 0.8 km2 0.1% b | 27 km2 50% b | 26 km2 49% b | 2.4 km2 0.5% b | 463 km2 52% b | 416 km2 47% b | 2.6 km2 0.29% b |
| Afforestation | ||||||||
| 1973–2002 | 2002–2017 | 1973–2017 | ||||||
| --- | --- | --- | --- | --- | --- | Wetland to Forest | Wetland to Shrub | Shrub to Forest |
| --- | --- | --- | --- | --- | --- | 0.6 km2 0.2% b | 2.6 km2 1.5% c | 0.9 km2 0.3% b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baughman, C.A.; Loehman, R.A.; Magness, D.R.; Saperstein, L.B.; Sherriff, R.L. Four Decades of Land-Cover Change on the Kenai Peninsula, Alaska: Detecting Disturbance-Influenced Vegetation Shifts Using Landsat Legacy Data. Land 2020, 9, 382. https://doi.org/10.3390/land9100382
Baughman CA, Loehman RA, Magness DR, Saperstein LB, Sherriff RL. Four Decades of Land-Cover Change on the Kenai Peninsula, Alaska: Detecting Disturbance-Influenced Vegetation Shifts Using Landsat Legacy Data. Land. 2020; 9(10):382. https://doi.org/10.3390/land9100382
Chicago/Turabian StyleBaughman, Carson A., Rachel A. Loehman, Dawn R. Magness, Lisa B. Saperstein, and Rosemary L. Sherriff. 2020. "Four Decades of Land-Cover Change on the Kenai Peninsula, Alaska: Detecting Disturbance-Influenced Vegetation Shifts Using Landsat Legacy Data" Land 9, no. 10: 382. https://doi.org/10.3390/land9100382
APA StyleBaughman, C. A., Loehman, R. A., Magness, D. R., Saperstein, L. B., & Sherriff, R. L. (2020). Four Decades of Land-Cover Change on the Kenai Peninsula, Alaska: Detecting Disturbance-Influenced Vegetation Shifts Using Landsat Legacy Data. Land, 9(10), 382. https://doi.org/10.3390/land9100382