
Effects of Bothriochloa ischaemum on the Diversity of Pannonian Sandy Grasslands

 ,
,  ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- Q1: How does the presence and cover of B. ischaemum affect the diversity, naturalness, and ratio of the species of Pannonian sandy grasslands?
- Q2: To what extent does this diversity change depend on site characteristics?
2. Materials and Methods
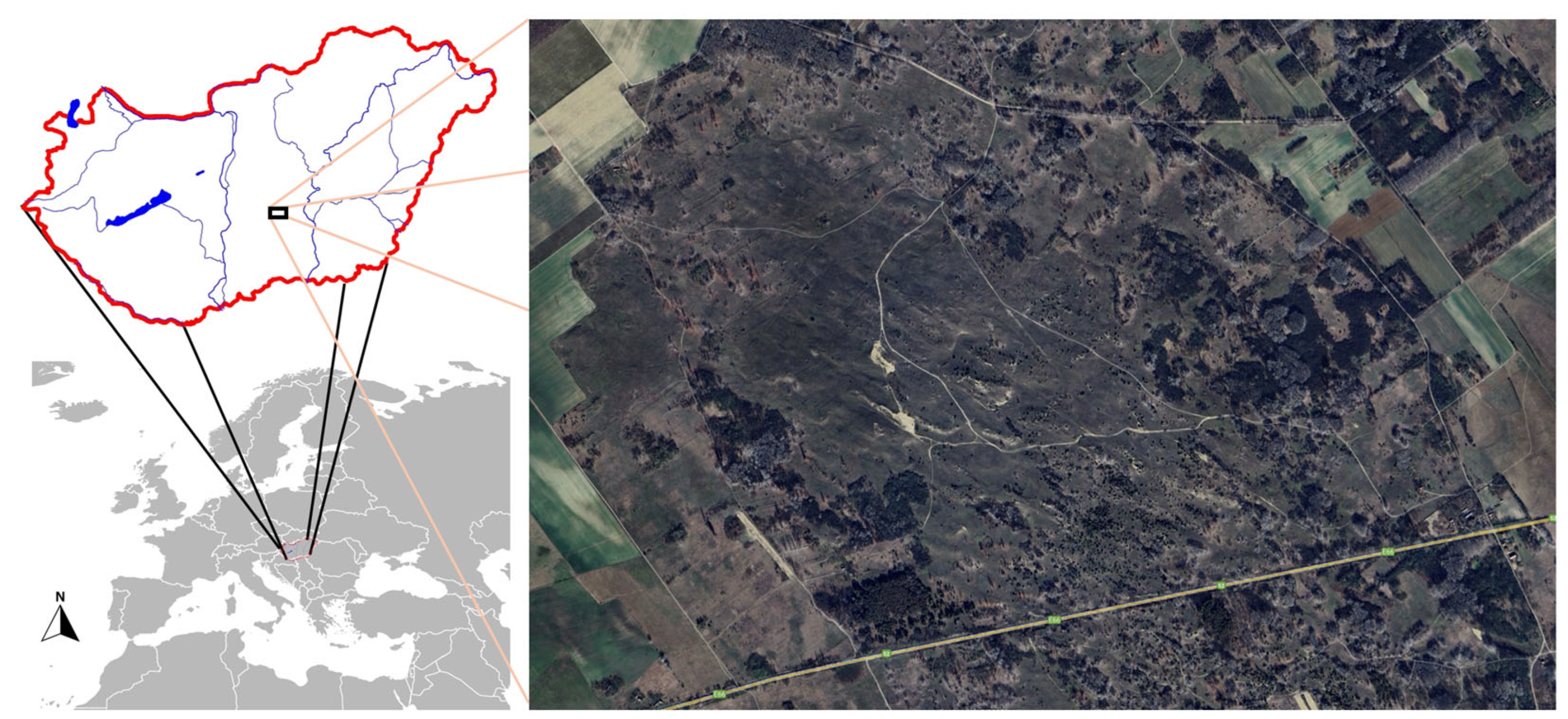
2.1. Study Area
2.2. Coenological Studies
2.3. Data Analysis
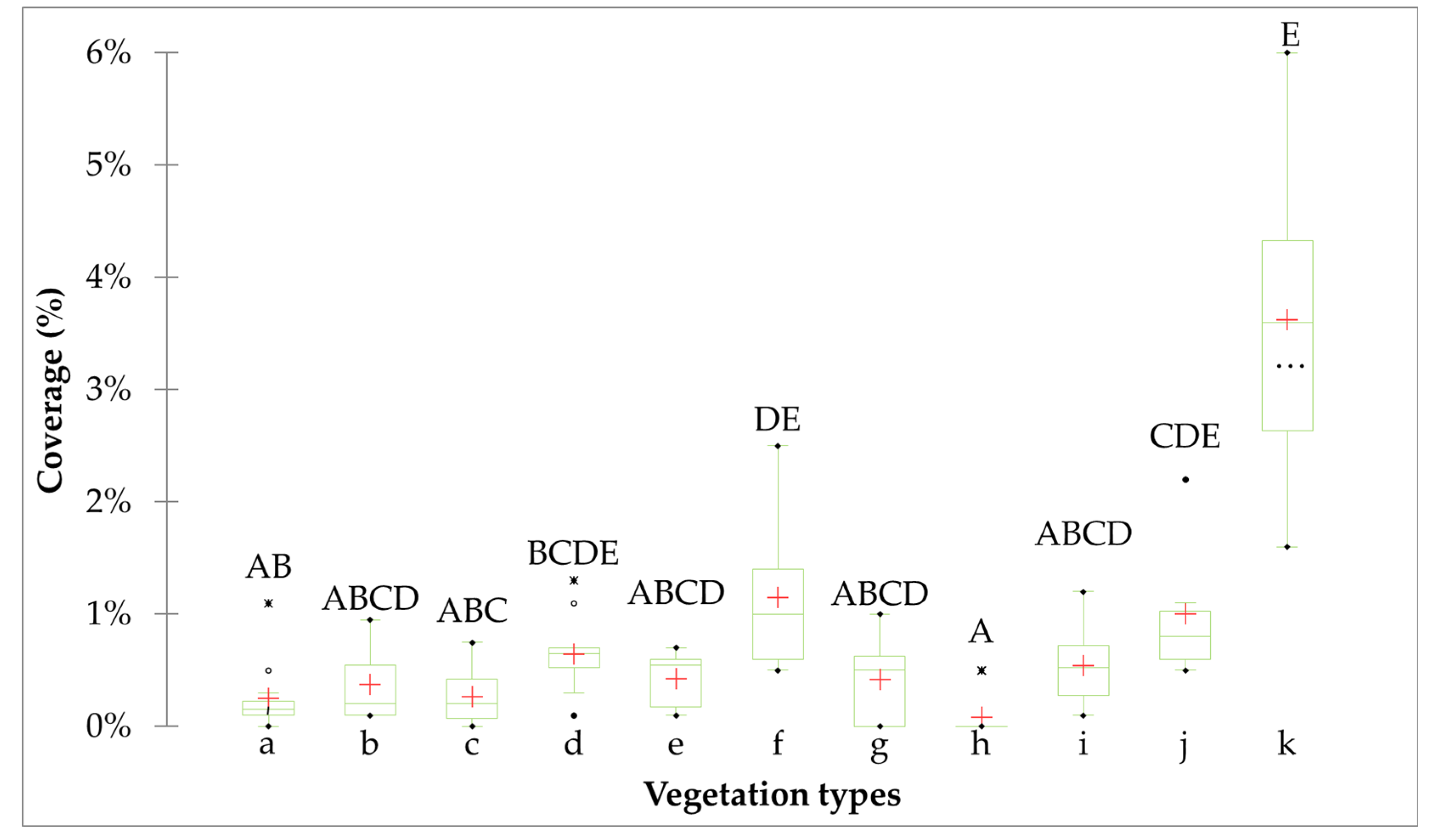
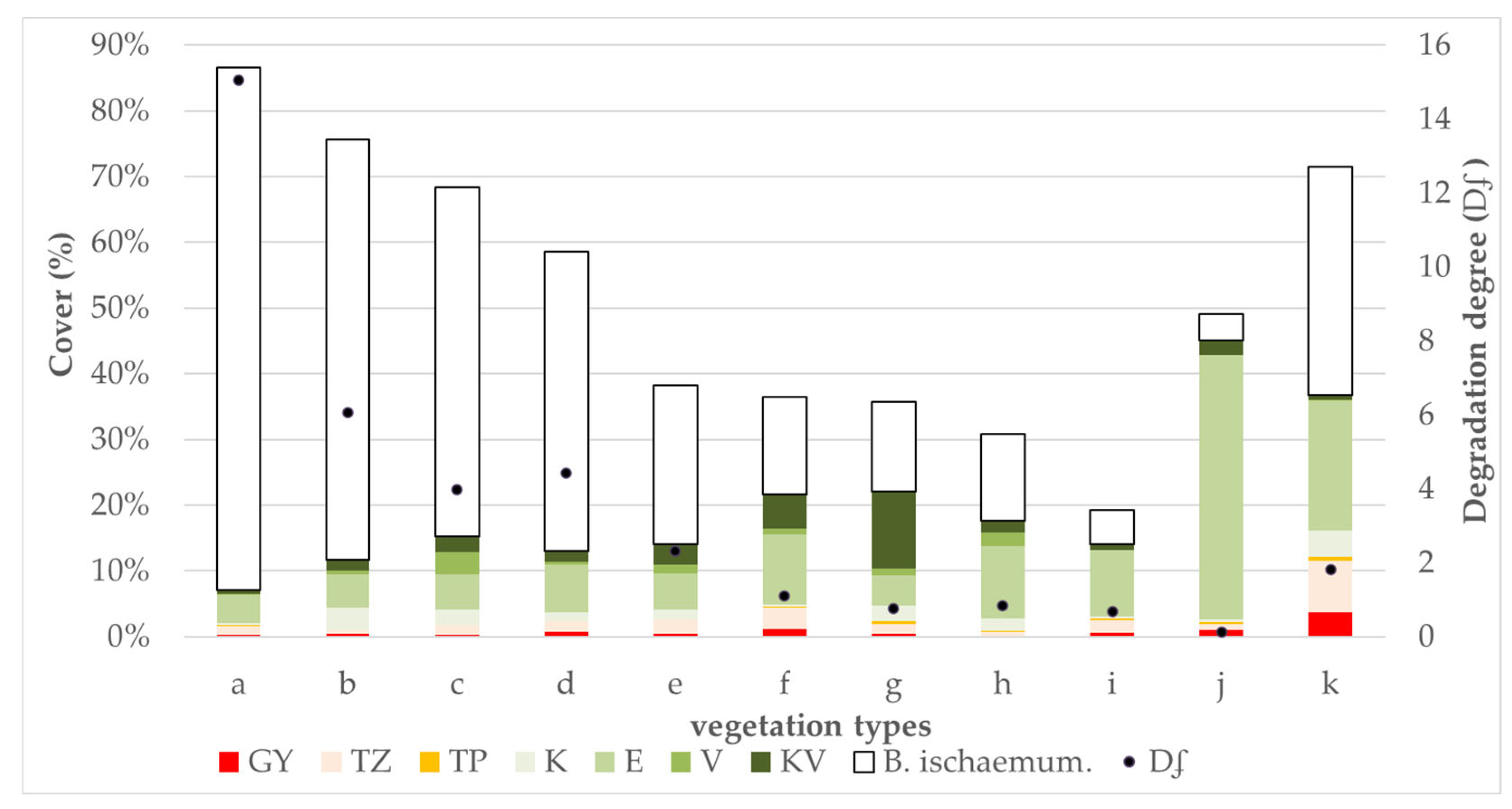
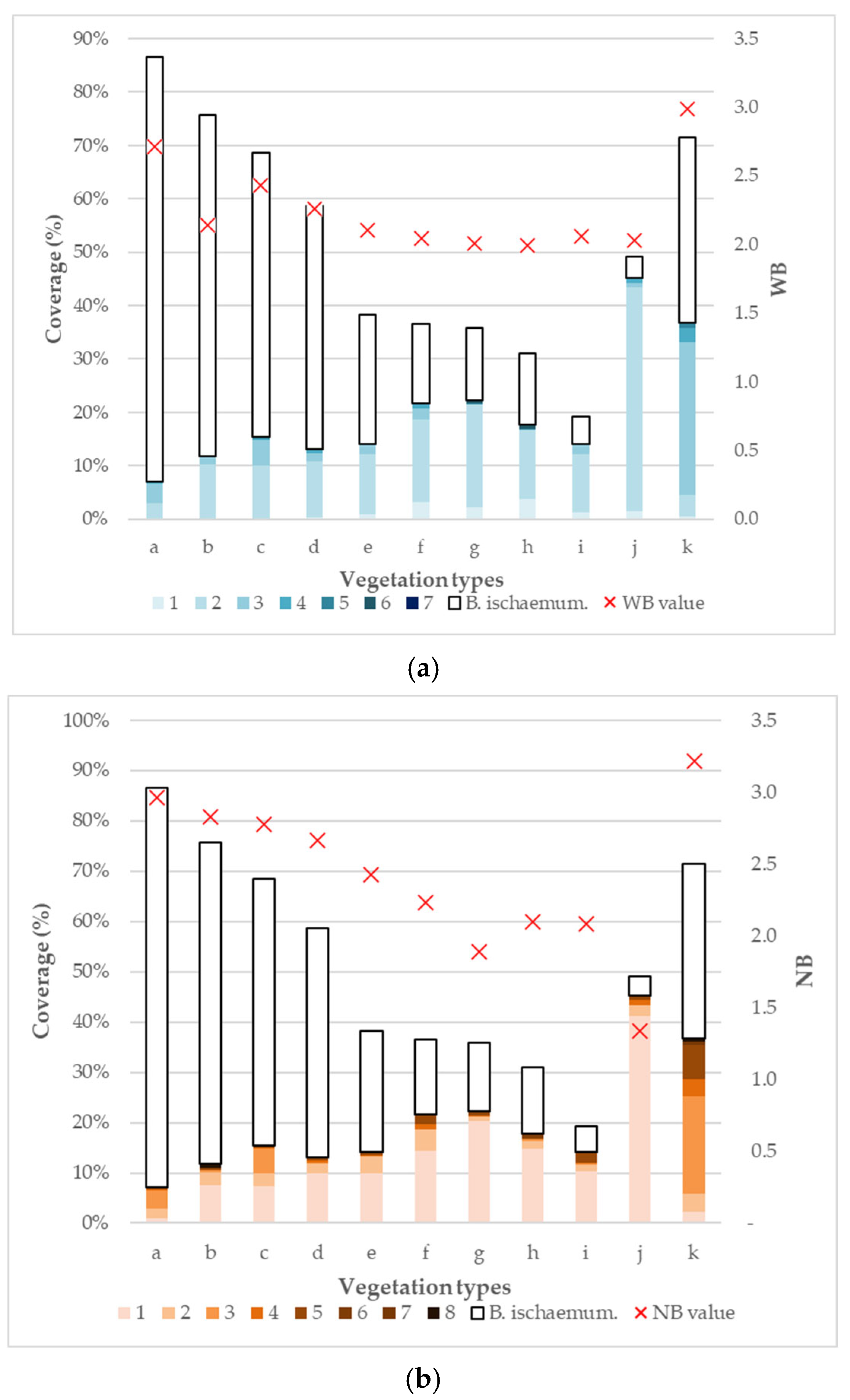
3. Results
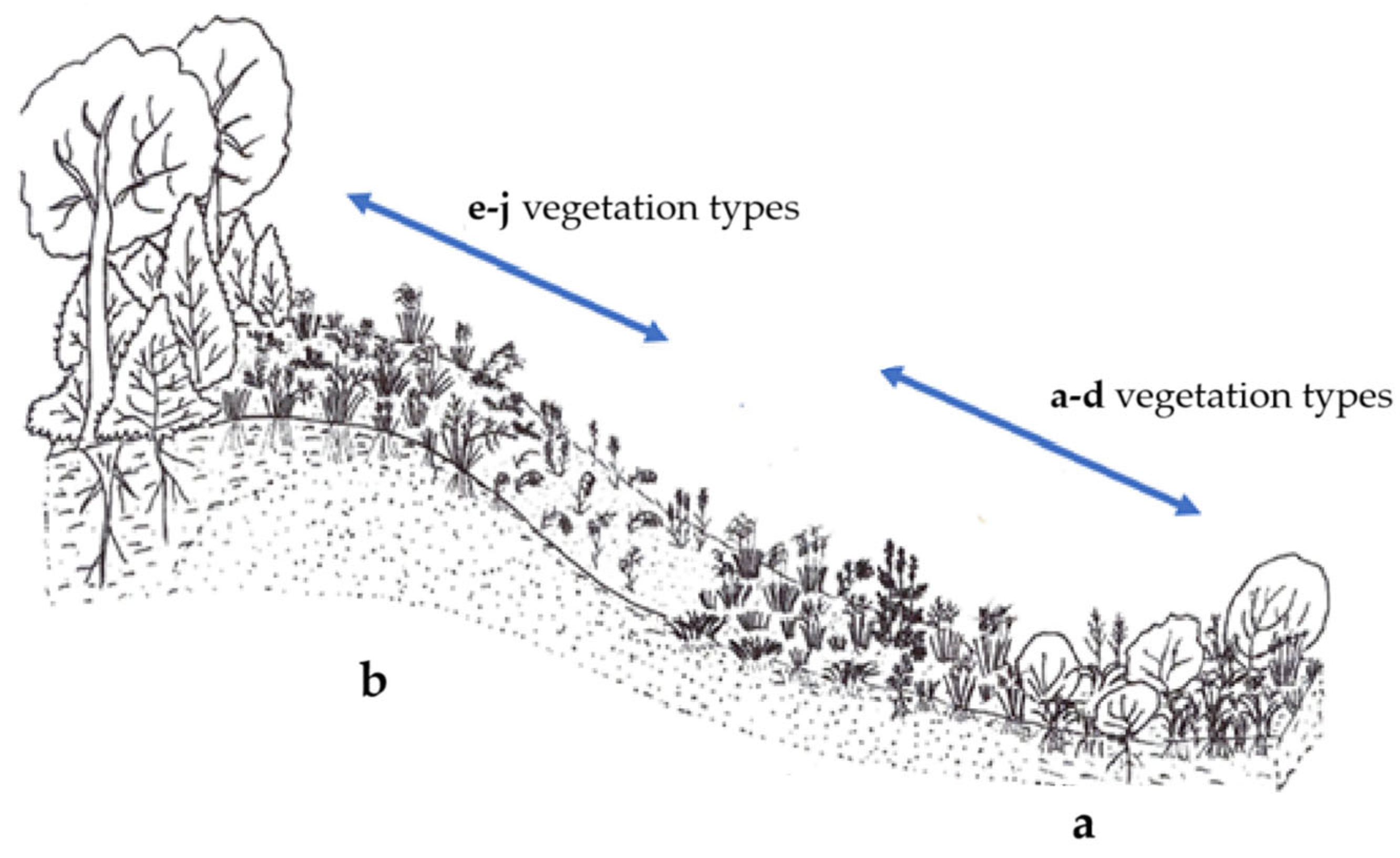
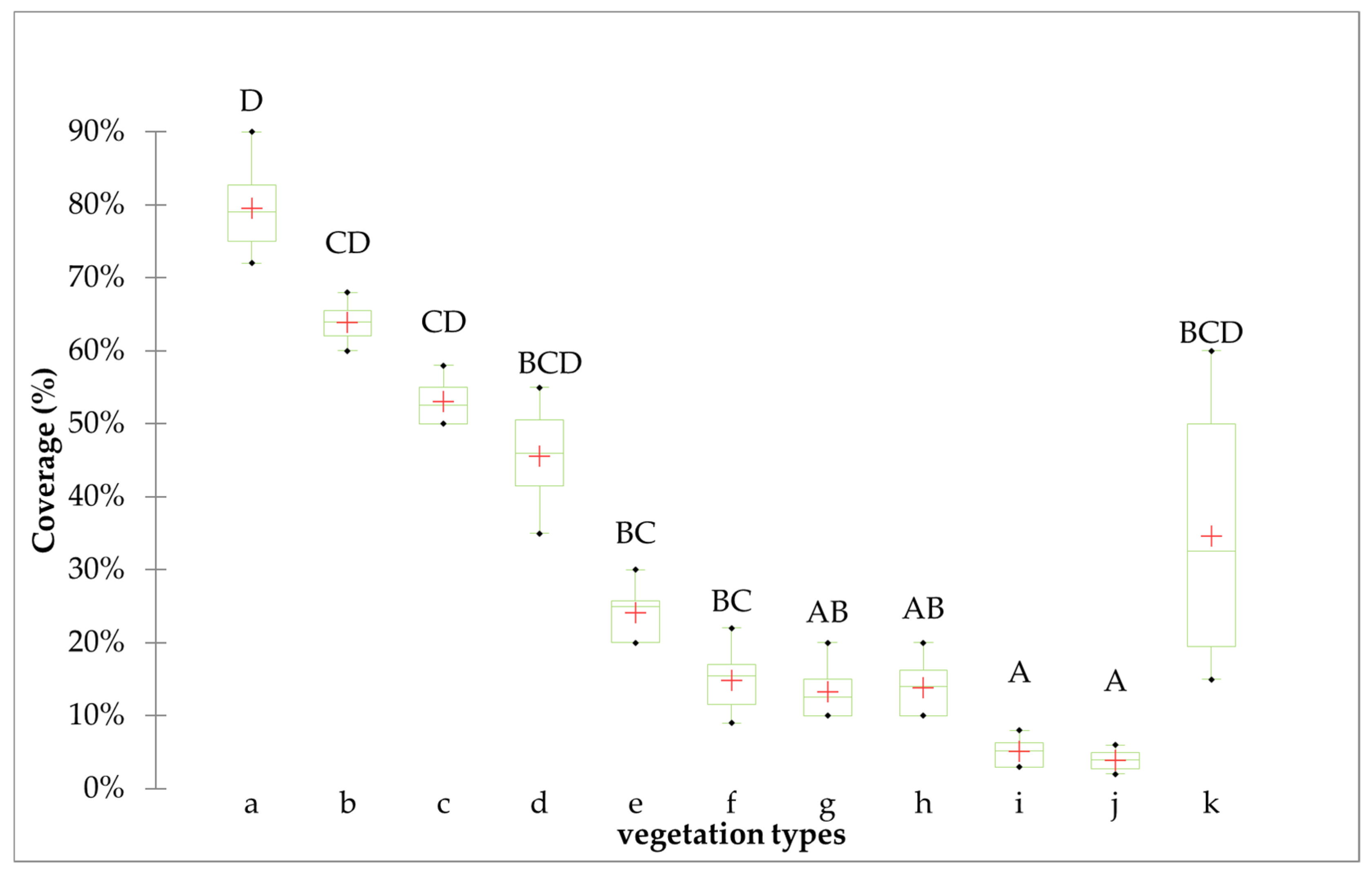
3.1. Characterisation of Each Vegetation Type
3.1.1. Vegetation Types in the Dune Slacks
3.1.2. Vegetation Types on Dune Sides
3.1.3. Vegetation of the Old-Field Area
3.2. Cluster Analysis of Vegetation Types
3.2.1. Relationship Between B. ischaemum and Other Abundant Species
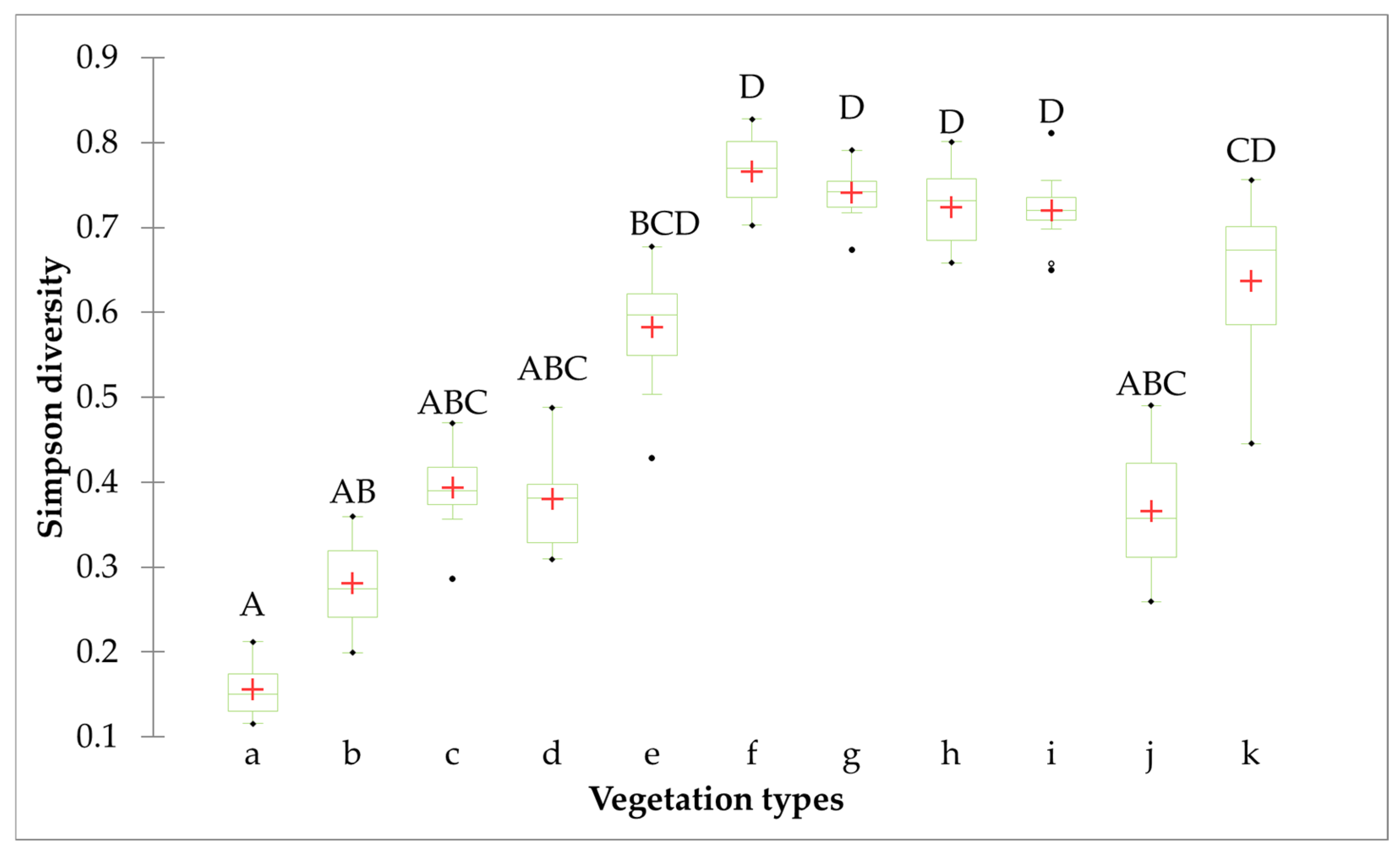
3.2.2. Relationship Between B. ischaemum and Diversity
3.3. Results of Vegetation Types According to Conservation and Relative Ecological Indicators
3.4. Water and Nitrogen Requirement of Vegetation Types
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yelenik, S.G.; Stock, W.D.; Richardson, D.M. Functional Group Identity Does Not Predict Invader Impacts: Differential Effects of Nitrogen-Fixing Exotic Plants on Ecosystem Function. Biol. Invasions 2007, 9, 117–125. [Google Scholar] [CrossRef]
- Vantarová, K.H.; Eliáš, P.; Jiménez-Ruiz, J.; Tokarska-Guzik, B.; Cires, E. Biological Invasions in the Twenty-First Century: A Global Risk. Biologia 2023, 78, 1211–1218. [Google Scholar] [CrossRef]
- Cleland, E.E.; Collins, S.L.; Dickson, T.L.; Farrer, E.C.; Gross, K.L.; Gherardi, L.A.; Hallett, L.M.; Hobbs, R.J.; Hsu, J.S.; Turnbull, L.; et al. Sensitivity of Grassland Plant Community Composition to Spatial vs. Temporal Variation in Precipitation. Ecology 2013, 94, 1687–1696. [Google Scholar] [CrossRef]
- Afzal, M.R.; Naz, M.; Ashraf, W.; Du, D. The Legacy of Plant Invasion: Impacts on Soil Nitrification and Management Implications. Plants 2023, 12, 2980. [Google Scholar] [CrossRef] [PubMed]
- Sitters, J.; Wubs, E.R.J.; Bakker, E.S.; Crowther, T.W.; Adler, P.B.; Bagchi, S.; Bakker, J.D.; Biederman, L.; Borer, E.T.; Cleland, E.E.; et al. Nutrient Availability Controls the Impact of Mammalian Herbivores on Soil Carbon and Nitrogen Pools in Grasslands. Glob. Change Biol. 2020, 26, 2060–2071. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Ibáñez, I. Plant Invasions in the Landscape. Landsc. Ecol. 2011, 26, 461–472. [Google Scholar] [CrossRef]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All Is Not Loss: Plant Biodiversity in the Anthropocene. PLoS ONE 2012, 7, e30535. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, L.; Miiller, N.; Prestes, J.G.; Vitule, J.; Cuthbert, R.N. Impacts and Mechanisms of Biological Invasions in Global Protected Areas. Biol. Invasions 2024, 27, 20. [Google Scholar] [CrossRef]
- Kortz, A.R.; Magurran, A.E. Increases in Local Richness (α-Diversity) Following Invasion Are Offset by Biotic Homogenization in a Biodiversity Hotspot. Biol. Lett. 2019, 15, 20190133. [Google Scholar] [CrossRef]
- Spatz, D.R.; Holmes, N.D.; Will, D.J.; Hein, S.; Carter, Z.T.; Fewster, R.M.; Keitt, B.; Genovesi, P.; Samaniego, A.; Croll, D.A.; et al. The Global Contribution of Invasive Vertebrate Eradication as a Key Island Restoration Tool. Sci. Rep. 2022, 12, 13391. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-Analysis of Their Effects on Species, Communities and Ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Liu, Z.; Song, W.; Chen, X.; Zhang, Z.; Li, B.; van Kleunen, M.; Wu, J. Biodiversity Increases Resistance of Grasslands against Plant Invasions under Multiple Environmental Changes. Nat. Commun. 2024, 15, 4506. [Google Scholar] [CrossRef] [PubMed]
- Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.W.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K.; Blanco, G.; et al. IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2023. [Google Scholar]
- Gruner, D.S.; Bracken, M.E.S.; Berger, S.A.; Eriksson, B.K.; Gamfeldt, L.; Matthiessen, B.; Moorthi, S.; Sommer, U.; Hillebrand, H. Effects of Experimental Warming on Biodiversity Depend on Ecosystem Type and Local Species Composition. Oikos 2017, 126, 8–17. [Google Scholar] [CrossRef]
- Piseddu, F.; Bellocchi, G.; Picon-Cochard, C. Mowing and Warming Effects on Grassland Species Richness and Harvested Biomass: Meta-Analyses. Agron. Sustain. Dev. 2021, 41, 74. [Google Scholar] [CrossRef]
- Korell, L.; Auge, H.; Chase, J.M.; Harpole, W.S.; Knight, T.M. Responses of Plant Diversity to Precipitation Change Are Strongest at Local Spatial Scales and in Drylands. Nat. Commun. 2021, 12, 2489. [Google Scholar] [CrossRef]
- Szigetvári, C. Phytosociological and Edaphyc Aspects of the Invasion by Cleistogenes serotina (L.) Keng in the Kiskunság National Park. Tiscia 2000, 32, 9–17. [Google Scholar]
- Alofs, K.M.; Fowler, N.L. Loss of Native Herbaceous Species Due to Woody Plant Encroachment Facilitates the Establishment of an Invasive Grass. Ecology 2013, 94, 751–760. [Google Scholar] [CrossRef]
- Manzoor, S.A.; Griffiths, G.; Lukac, M. Land Use and Climate Change Interaction Triggers Contrasting Trajectories of Biological Invasion. Ecol. Indic. 2021, 120, 106936. [Google Scholar] [CrossRef]
- de Barros Ruas, R.; Costa, L.M.S.; Bered, F. Urbanization Driving Changes in Plant Species and Communities—A Global View. Glob. Ecol. Conserv. 2022, 38, e02243. [Google Scholar] [CrossRef]
- Juhasz, M.; Bagi, I. The Critical Effect of Drought Stress on the Invading Features of Bird Cherry (Prunus Serotina Ehrh.) during Dispersion Stage on a Sand-Dune Area in Hungary. Cereal Res. Commun. 2009, 37, 553–556. [Google Scholar]
- Szigetvári, C. Distribution and phytosociological relations of two introduced plant species in an open sand grassland area in the Great Hungarian Plain. Acta Bot. Hung. 2002, 44, 163–183. [Google Scholar] [CrossRef]
- Csecserits, A.; Botta-Dukát, Z.; Kröel-Dulay, G.; Lhotsky, B.; Ónodi, G.; Rédei, T.; Szitár, K.; Halassy, M. Tree Plantations Are Hot-Spots of Plant Invasion in a Landscape with Heterogeneous Land-Use. Agric. Ecosyst. Environ. 2016, 226, 88–98. [Google Scholar] [CrossRef]
- Kelemen, A.; Valkó, O.; Kröel-Dulay, G.; Deák, B.; Török, P.; Tóth, K.; Miglécz, T.; Tóthmérész, B. The Invasion of Common Milkweed (Asclepias syriaca) in Sandy Old-Fields—Is It a Threat to the Native Flora? Appl. Veg. Sci. 2016, 19, 218–224. [Google Scholar] [CrossRef]
- Meinhardt, S.; Saláta, D.; Tormáné Kovács, E.; Ábrám, Ö.; Morvai, E.; Szirmai, O.; Czóbel, S. The Multifaceted Botanical Impact of the Invasive Common Milkweed (Asclepias syriaca L.) in a Protected Sandy Grassland in Central Europe. Land 2024, 13, 1594. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Tóth, V.; Hábenczyus, A.A.; Frei, K.; Tóth, B.; Erdős, L.; Török, P.; Bátori, Z. Suppressing the Invasive Common Milkweed (Asclepias syriaca L.) Saves Soil Moisture Reserves. Biol. Invasions 2024, 26, 2791–2799. [Google Scholar] [CrossRef]
- Yan, Y.; Jarvie, S.; Zhang, Q.; Zhang, S.; Han, P.; Liu, Q.; Liu, P. Small Patches Are Hotspots for Biodiversity Conservation in Fragmented Landscapes. Ecol. Indic. 2021, 130, 108086. [Google Scholar] [CrossRef]
- Wani, I.A.; Verma, S.; Gupta, R.; Ganaie, M.M.; Nigam, G.; Shafik, H.M.; Al-Misned, F.A. Nutrient Analysis and Species Diversity of Alpine Grasslands: A Comparative Analysis of Less Studied Biodiversity Hotspots. Sustainability 2022, 14, 887. [Google Scholar] [CrossRef]
- Janišová, M.; Sorescu-Marinković, A.; Aćić, S.; Hubáčková, B.; Magnes, M.; Opravil, Š.; Širka, P. Exploring a Grassland Biodiversity Hotspot in the Serbian Carpathians: Interdisciplinary Perspectives and Conservation Implications. Biol. Conserv. 2024, 299, 110822. [Google Scholar] [CrossRef]
- Borer, E.T.; Risch, A.C. Planning for the Future: Grasslands, Herbivores, and Nature-Based Solutions. J. Ecol. 2024, 112, 2442–2450. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More Important for Ecosystem Services than You Might Think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Malisch, C.S.; Finn, J.A.; Eriksen, J.; Loges, R.; Brophy, C.; Huguenin-Elie, O. The Importance of Multi-Species Grassland Leys to Enhance Ecosystem Services in Crop Rotations. Grass Forage Sci. 2024, 79, 120–134. [Google Scholar] [CrossRef]
- Prangel, E.; Kasari-Toussaint, L.; Neuenkamp, L.; Noreika, N.; Karise, R.; Marja, R.; Ingerpuu, N.; Kupper, T.; Keerberg, L.; Oja, E.; et al. Afforestation and Abandonment of Semi-Natural Grasslands Lead to Biodiversity Loss and a Decline in Ecosystem Services and Functions. J. Appl. Ecol. 2023, 60, 825–836. [Google Scholar] [CrossRef]
- Hábenczyus, A.A.; Tölgyesi, C.; Pál, R.; Kelemen, A.; Aradi, E.; Bátori, Z.; Sonkoly, J.; Tóth, E.; Balogh, N.; Török, P. Increasing Abundance of an Invasive C4 Grass Is Associated with Larger Community Changes Away than at Home. Appl. Veg. Sci. 2022, 25, e12659. [Google Scholar] [CrossRef]
- Explosive Spread of Sand Dropseed (Sporobolus cryptandrus), a C4 Perennial Bunchgrass, Threatens Unique Grasslands in Hungary (Central Europe). NeoBiota 2024, 95, 59–75. [CrossRef]
- Török, P.; Espinoza Ami, F.D.; Szél-Tóth, K.; Díaz Cando, P.; Guallichico Suntaxi, L.R.; McIntosh-Buday, A.; Hábenczyus, A.A.; Törő-Szijgyártó, V.; Kovacsics-Vári, G.; Tölgyesi, C.; et al. Accumulated Soil Seed Bank of the Invasive Sand Dropseed (Sporobolus cryptandrus) Poses a Challenge for Its Suppression. Land Degrad. Dev. 2024, 35, 4105–4120. [Google Scholar] [CrossRef]
- Török, P.; Schmidt, D.; Bátori, Z.; Aradi, E.; Kelemen, A.; Hábenczyus, A.A.; Cando, P.D.; Tölgyesi, C.; Pál, R.W.; Balogh, N.; et al. Invasion of the North American Sand Dropseed (Sporobolus cryptandrus)—A New Pest in Eurasian Sand Areas? Glob. Ecol. Conserv. 2021, 32, e01942. [Google Scholar] [CrossRef]
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural Grassland Management by Mowing of Calamagrostis epigejos in Hungary. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2011, 145, 699–707. [Google Scholar] [CrossRef]
- Házi, J.; Penksza, K.; Barczi, A.; Szentes, S.; Pápay, G. Effects of Long-Term Mowing on Biomass Composition in Pannonian Dry Grasslands. Agronomy 2022, 12, 1107. [Google Scholar] [CrossRef]
- Somodi, I.; Virágh, K.; Podani, J. The Effect of the Expansion of the Clonal Grass Calamagrostis epigejos on the Species Turnover of a Semi-Arid Grassland. Appl. Veg. Sci. 2008, 11, 187–192. [Google Scholar] [CrossRef]
- Błońska, A.; Chmura, D.; Hutniczak, A.; Wilczek, Z.; Jarosz, J.; Besenyei, L.; Woźniak, G. The Plant Species Composition of an Abandoned Meadow as an Element of an Ecosystem Mosaic within an Urban-Industrial Landscape. Sustainability 2022, 14, 11851. [Google Scholar] [CrossRef]
- Házi, J.; Purger, D.; Penksza, K.; Bartha, S. Changes in Species Composition, Diversity, and Biomass of Secondary Dry Grasslands Following Long-Term Mowing: A Case Study in Hungary. Grasses 2024, 3, 130–142. [Google Scholar] [CrossRef]
- Anibaba, Q.A.; Dyderski, M.K.; Woźniak, G.; Jagodziński, A.M. The Inhibitory Tendency of Calamagrostis epigejos and Solidago spp. Depends on the Successional Stage in Postindustrial Vegetation. Land Degrad. Dev. 2025, 36, 121–132. [Google Scholar] [CrossRef]
- Lazu, S.; Titica, G.; Miron, A.; Teleuța, A.; Talmaci, L.; Guțu, A.I.; Dziuba, T. Syntaxonomy of Steppe Semi-Savannoid with Chrysopogon gryllus (L.) Trin. and Bothriochloa ischaemum (L.) Keng. from the Eurasian Area. Rom. J. Grassl. Forage Crops 2021, 23, 13–42. [Google Scholar]
- White, L.M.; Dewald, C.L. Yield and Quality of WW-Iron Master and Caucasian Bluestem Regrowth. J. Range Manag. 1996, 49, 42–45. [Google Scholar] [CrossRef]
- Harmoney, K.R.; Stahlman, P.W.; Hickman, K.R. Herbicide Effect Son Established Yellow Old World Bluestem (Bothriochloa ischaemum). Weed Technol. 2004, 18, 545–550. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Polley, H.W. Effects of Seed Additions and Grazing History on Diversity and Productivity of Subhumid Grasslands. Ecology 2003, 84, 920–931. [Google Scholar] [CrossRef]
- Ruffner, M.E.; McCulley, R.L.; Nelson, J.A.; Barnes, T.G. Ecosystem Function Differs between Old World Bluestem Invaded and Native Coastal Prairie in South Texas. Biol. Invasions 2012, 14, 1483–1500. [Google Scholar] [CrossRef]
- Jiao, J.; Tzanopoulos, J.; Xofis, P.; Bai, W.; Ma, X.; Mitchley, J. Can the Study of Natural Vegetation Succession Assist in the Control of Soil Erosion on Abandoned Croplands on the Loess Plateau, China? Restor. Ecol. 2007, 15, 391–399. [Google Scholar] [CrossRef]
- Zhang, J.-T.; Zhang, G.-L. Ecological Situation and Management of Bothriochloa ischaemum Grasslands in China. Grassl. Sci. 2006, 52, 85–93. [Google Scholar] [CrossRef]
- Lazu, S.; Titica, G.; Talmaci, L. Phytocoenotic Diversity of Grasslands with Bothriochloa ischaemum (L.) Keng in Eurasia. Rom. J. Grassl. Forage Crops 2018, 18, 15–36. [Google Scholar]
- Wang, Z.; Xu, W.; Chen, Z.; Jia, Z.; Huang, J.; Wen, Z.; Chen, Y.; Xu, B. Soil Moisture Availability at Early Growth Stages Strongly Affected Root Growth of Bothriochloa ischaemum When Mixed with Lespedeza Davurica. Front. Plant Sci. 2018, 9, 1050. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Deng, X.; Xu, B.; Palta, J.A.; Chen, Y. Soil Water Availability Changes in Amount and Timing Favor the Growth and Competitiveness of Grass Than a Co-Dominant Legume in Their Mixtures. Front. Plant Sci. 2021, 12, 723839. [Google Scholar] [CrossRef]
- Szentes, S.; Sutyinszki, Z.; Szabó, G.; Zimmermann, Z.; Házi, J.; Wichmann, B.; Hufnágel, L.; Penksza, K.; Bartha, S. Grazed Pannonian Grassland Beta-Diversity Changes Due to C4 Yellow Bluestem. Cent. Eur. J. Biol. 2012, 7, 1055–1065. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in Invasion Biology: Predicting Invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Ortega, Y.K.; Pearson, D.E. Weak Vs. Strong Invaders of Natural Plant Communities: Assessing Invasibility and Impact. Ecol. Appl. 2005, 15, 651–661. [Google Scholar] [CrossRef]
- Allan, E.; Manning, P.; Alt, F.; Binkenstein, J.; Blaser, S.; Blüthgen, N.; Böhm, S.; Grassein, F.; Hölzel, N.; Klaus, V.H.; et al. Land Use Intensification Alters Ecosystem Multifunctionality via Loss of Biodiversity and Changes to Functional Composition. Ecol. Lett. 2015, 18, 834–843. [Google Scholar] [CrossRef]
- Virágh, K.; Fekete, G. Degradation Stages in a Xeroseries: Composition, Similarity, Grouping, Coordination. Acta Bot. Hung. 1984, 30, 427–459. [Google Scholar]
- Zólyomi, B.; Fekete, G. The Pannonian Loess Steppe: Differentiation in Space and Time. Abstr. Bot. 1994, 18, 29–41. [Google Scholar]
- Bartha, S. Composition, Differentiation and Dynamics of Thegrasslands in Theforreststeppebiome. In Slopesteppes, Loesssteppes and Forreststeppemeadows in Hungary; MTA-ÖBKI: Vácrátót, Hungary, 2007; pp. 194–210. [Google Scholar]
- Li, S.; Huang, X.; Shen, J.; Xu, F.; Su, J. Effects of Plant Diversity and Soil Properties on Soil Fungal Community Structure with Secondary Succession in the Pinus yunnanensis Forest. Geoderma 2020, 379, 114646. [Google Scholar] [CrossRef]
- Modelling Competitive Interactions and Plant–Soil Feedback in Vegetation Dynamics | Ricerche Di Matematica. Available online: https://link.springer.com/article/10.1007/s11587-020-00497-6 (accessed on 17 January 2025).
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past Trends, Present State and Future Prospects of Hungarian Forest-Steppes. In Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World; Werger, M.J.A., van Staalduinen, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 209–252. ISBN 978-94-007-3886-7. [Google Scholar]
- Tölgyesi, C.; Körmöczi, L. Structural Changes of a Pannonian Grassland Plant Community in Relation to the Decrease of Water Availability. Acta Bot. Hung. 2012, 54, 413–431. [Google Scholar] [CrossRef]
- Erdős, L.; Kröel-Dulay, G.; Bátori, Z.; Kovács, B.; Németh, C.; Kiss, P.J.; Tölgyesi, C. Habitat Heterogeneity as a Key to High Conservation Value in Forest-Grassland Mosaics. Biol. Conserv. 2018, 226, 72–80. [Google Scholar] [CrossRef]
- Erdős, L.; Krstonošić, D.; Kiss, P.J.; Bátori, Z.; Tölgyesi, C.; Škvorc, Ž. Plant Composition and Diversity at Edges in a Semi-Natural Forest–Grassland Mosaic. Plant Ecol. 2019, 220, 279–292. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Valkó, O.; Deák, B.; Kelemen, A.; Bragina, T.M.; Gallé, R.; Erdős, L.; Bátori, Z. Tree–Herb Co-Existence and Community Assembly in Natural Forest-Steppe Transitions. Plant Ecol. Divers. 2018, 11, 465–477. [Google Scholar] [CrossRef]
- Bátori, Z.; Erdős, L.; Kelemen, A.; Deák, B.; Valkó, O.; Gallé, R.; Bragina, T.M.; Kiss, P.J.; Kröel-Dulay, G.; Tölgyesi, C. Diversity Patterns in Sandy Forest-Steppes: A Comparative Study from the Western and Central Palaearctic. Biodivers. Conserv. 2018, 27, 1011–1030. [Google Scholar] [CrossRef]
- Bakacsy, L.; Szepesi, Á. A Case Study on the Early Stage of Pinus Nigra Invasion and Its Impact on Species Composition and Pattern in Pannonic Sand Grassland. Sci. Rep. 2024, 14, 5125. [Google Scholar] [CrossRef]
- Ónodi, G.; Kertész, M.; Lengyel, A.; Pándi, I.; Somay, L.; Szitár, K.; Kröel-Dulay, G. The Effects of Woody Plant Encroachment and Wildfire on Plant Species Richness and Composition: Temporal Changes in a Forest–Steppe Mosaic. Appl. Veg. Sci. 2021, 24, e12546. [Google Scholar] [CrossRef]
- Molnár, Z. The History of the Landscape of the Sand Hummocks (Strázsa Hill) of Fülöpháza in the Last Two Hundred and Fifty Years. In Dry Sand Vegetation of the Kiskunság; Természetbúvár Alapítvány Kiadó: Budapest, Hungary, 2003; p. 60. [Google Scholar]
- Király, G. Új magyar füvészkönyv: Magyarország hajtásos növényei; Határozókulcsok. [New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key.]; Aggteleki Nemzeti Park Igazgatóság: Aggtelek, Hungary, 2009; ISBN 978-963-87082-9-8. (In Hungarian) [Google Scholar]
- Lumivero XLSTAT Statistical and Data Analysis Solution. Available online: https://www.xlstat.com/ (accessed on 14 October 2024).
- Simon, T. A Magyarországi Edényes Flóra Határozója. [Key to the Vascular Flora of Hungary], 4th revised ed.; Tankönyvkiadó: Budapest, Hungary, 2000. [Google Scholar]
- Borhidi, A. Social Behavior Types, the Naturalness and Relative Ecological Indicator Values of the Highre Plants in the Hungarian Flora. Acta Bot. Acad. Sci. Hung. 1995, 39, 97–181. [Google Scholar]
- Botta-Dukát, Z. Invasion of Alien Species to Hungarian (Semi-)Natural Habitats. Acta Bot. Hung. 2009, 50, 219–227. [Google Scholar] [CrossRef]
- Biró, M.; Révész, A.; Molnár, Z.; Horváth, F.; Czúcz, B. Regional Habitat Pattern of the Danube-Tisza Interfluve in Hungary II. Acta Bot. Hung. 2008, 50, 19–60. [Google Scholar] [CrossRef]
- Wied, J.P.; Perotto-Baldivieso, H.L.; Conkey, A.A.T.; Brennan, L.A.; Mata, J.M. Invasive Grasses in South Texas Rangelands: Historical Perspectives and Future Directions. Invasive Plant Sci. Manag. 2020, 13, 41–58. [Google Scholar] [CrossRef]
- Kovács-Láng, E.; Kröel-Dulay, G.; Kertész, M.; Fekete, G.; Bartha, S.; Mika, J.; Dobi-Wantuch, I.; Rédei, T.; Rajkai, K.; Hahn, I. Changes in the Composition of Sand Grasslands along a Climatic Gradient in Hungary and Implications for Climate Change. Phytocoenologia 2000, 30, 385–407. [Google Scholar] [CrossRef]
- Fiala, K.; Holub, P.; Sedláková, I.; Tůma, I.; Záhora, J.; Tesařová, M. Reasons and Consequences of Expansion of Calamagrostis epigeios in Alluvial Meadows of Landscape Affected by Water Control Measures—A Multidisciplinary Research. Ekológia Bratisl. 2003, 22, 242–253. [Google Scholar]
- Kozak, J.; Ostapowicz, K.; Szablowska-Midor, A.; Widacki, W. Land abandonmentin the Western Beskidy Mountains and its environmental background. Ekológia Bratisl. 2004, 23, 116–126. [Google Scholar]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive Human-Driven Decline of Life on Earth Points to the Need for Transformative Change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef]
- Nadal-Romero, E.; Khorchani, M.; Gaspar, L.; Arnáez, J.; Cammeraat, E.; Navas, A.; Lasanta, T. How Do Land Use and Land Cover Changes after Farmland Abandonment Affect Soil Properties and Soil Nutrients in Mediterranean Mountain Agroecosystems? CATENA 2023, 226, 107062. [Google Scholar] [CrossRef]
- Lindborg, R. Recreating Grasslands in Swedish Rural Landscapes—Effects of Seed Sowing and Management History. Biodivers. Conserv. 2006, 15, 957–969. [Google Scholar] [CrossRef]
- Mendenhall, C.D.; Karp, D.S.; Meyer, C.F.J.; Hadly, E.A.; Daily, G.C. Predicting Biodiversity Change and Averting Collapse in Agricultural Landscapes. Nature 2014, 509, 213–217. [Google Scholar] [CrossRef]
- Liu, D.; Semenchuk, P.; Essl, F.; Lenzner, B.; Moser, D.; Blackburn, T.M.; Cassey, P.; Biancolini, D.; Capinha, C.; Dawson, W.; et al. The Impact of Land Use on Non-Native Species Incidence and Number in Local Assemblages Worldwide. Nat. Commun. 2023, 14, 2090. [Google Scholar] [CrossRef]
- Shivanna, K.R. Climate Change and Its Impact on Biodiversity and Human Welfare. Proc. Indian Natl. Sci. Acad. 2022, 88, 160–171. [Google Scholar] [CrossRef]
- Harlen, J.; Celarier, R.; Richardson, W. Studies on Old World Bluestem II. Okla. Agric. Exp. Stn. Tech. Bull. 1958, 72, 23. [Google Scholar]
- Hickman, K.R.; Farley, G.H.; Channell, R.; Steier, J.E. Effects of Old World Bluestem (Bothriochloa ischaemum) on Food Availability and Avian Community Composition within the Mixed-Grass Prairie. Southwest. Nat. 2006, 51, 524–530. [Google Scholar] [CrossRef]
- Bartha, S.; Szentes, S.; Horváth, A.; Házi, J.; Zimmermann, Z.; Molnár, C.; Dancza, I.; Margóczi, K.; Pál, R.W.; Purger, D.; et al. Impact of Mid-Successional Dominant Species on the Diversity and Progress of Succession in Regenerating Temperate Grasslands. Appl. Veg. Sci. 2014, 17, 201–213. [Google Scholar] [CrossRef]
- Sammon, J.G.; Wilkins, K.T. Effects on Aninvasive Grass (Bothriochloa ischaemum) on a Grassland Rodent Community. Tex. J. Sci. 2005, 57, 371–382. [Google Scholar]
- George, A.D.; O’Connell, T.J.; Hickman, K.R.; Leslie, D.M., Jr. Food Availability in Exotic Grasslands: A Potential Mechanism for Depauperate Breeding Assemblages. Wilson J. Ornithol. 2013, 125, 526–533. [Google Scholar] [CrossRef]
- Greer, M.J.; Wilson, G.W.T.; Hickman, K.R.; Wilson, S.M. Experimental Evidence That Invasive Grasses Use Allelopathic Biochemicals as a Potential Mechanism for Invasion: Chemical Warfare in Nature. Plant Soil 2014, 385, 165–179. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xu, G.C.; Xiao, L.; Ren, Z.P.; Li, Z.B. Growth, Morphological, and Physiological Responses to Drought Stress in Bothriochloa ischaemum. Front. Plant Sci. 2017, 8, 230. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of Allelopathy on Plant Performance: A Meta-Analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant Species Richness: The World Records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, C.; Zhao, N.; Sun, X.; Hou, S.; Wang, P. Physiological Nitrogen Uptake and Utilisation Responses in Two Native Plants from the Qinghai-Tibet Plateau under Different Water and Fertiliser Conditions. Agronomy 2024, 14, 440. [Google Scholar] [CrossRef]
- Ding, W.; Xu, W.; Gao, Z.; Xu, B. Effects of Water and Nitrogen on Growth and Relative Competitive Ability of Introduced versus Native C4 Grass Species in the Semi-Arid Loess Plateau of China. J. Arid Land 2021, 13, 730–743. [Google Scholar] [CrossRef]
- Novak, E.N.; Bertelsen, M.; Davis, D.; Grobert, D.M.; Lyons, K.G.; Martina, J.P.; McCaw, W.M.; O’Toole, M.; Veldman, J.W. Season of Prescribed Fire Determines Grassland Restoration Outcomes after Fire Exclusion and Overgrazing. Ecosphere 2021, 12, e03730. [Google Scholar] [CrossRef]
- Havill, S.; Schwinning, S.; Lyons, K.G. Fire Effects on Invasive and Native Warm-Season Grass Species in a North American Grassland at a Time of Extreme Drought. Appl. Veg. Sci. 2015, 18, 637–649. [Google Scholar] [CrossRef]
- Xu, B.; Gao, Z.; Wang, J.; Xu, W.; Huang, J. Morphological Changes in Roots of Bothriochloa ischaemum Intercropped with Lespedeza Davurica Following Phosphorus Application and Water Stress. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2015, 149, 298–306. [Google Scholar] [CrossRef]
- Winkler, J.; Jeznach, J.; Koda, E.; Sas, W.; Mazur, Ł.; Vaverková, M.D. Promoting Biodiversity: Vegetation in a Model Small Park Located in the Research and Educational Centre. J. Ecol. Eng. 2022, 23, 146–157. [Google Scholar] [CrossRef]
- Long, A.L.; Kettenring, K.M.; Toth, R. Prioritizing Management of the Invasive Grass Common Reed (Phragmites australis) in Great Salt Lake Wetlands. Invasive Plant Sci. Manag. 2017, 10, 155–165. [Google Scholar] [CrossRef]
- Nagy, D.U.; Rauschert, E.S.J.; Callaway, R.M.; Henn, T.; Filep, R.; Pal, R.W. Intense Mowing Management Suppresses Invader, but Shifts Competitive Resistance by a Native to Facilitation. Restor. Ecol. 2022, 30, e13483. [Google Scholar] [CrossRef]
- Szépligeti, M.; Kőrösi, Á.; Szentirmai, I.; Házi, J.; Bartha, D.; Bartha, S. Evaluating Alternative Mowing Regimes for Conservation Management of Central European Mesic Hay Meadows: A Field Experiment. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2018, 152, 90–97. [Google Scholar] [CrossRef]
- Dee, J.R.; Thomas, S.M.; Thompson, S.D.; Palmer, M.W. Long-Term Late Season Mowing Maintains Diversity in Southern US Tallgrass Prairie Invaded by Bothriochloa ischaemum. Appl. Veg. Sci. 2016, 19, 442–453. [Google Scholar] [CrossRef]
- Atchison, J. Experiments in Co-Existence: The Science and Practices of Biocontrol in Invasive Species Management. Environ. Plan. Econ. Space 2015, 47, 1697–1712. [Google Scholar] [CrossRef]
- Duell, E.B.; Hickman, K.R.; Wilson, G.W.T. Inoculation with Native Grassland Soils Improves Native Plant Species Germination in Highly Disturbed Soil. Grassl. Res. 2022, 1, 75–83. [Google Scholar] [CrossRef]
- Duell, E.B.; O’Hare, A.; Wilson, G.W.T. Inoculation with Native Soil Improves Seedling Survival and Reduces Non-Native Reinvasion in a Grassland Restoration. Restor. Ecol. 2023, 31, e13685. [Google Scholar] [CrossRef]
- Mungi, N.A.; Qureshi, Q.; Jhala, Y.V. Distribution, Drivers and Restoration Priorities of Plant Invasions in India. J. Appl. Ecol. 2023, 60, 2400–2412. [Google Scholar] [CrossRef]
- Prangel, E.; Reitalu, T.; Neuenkamp, L.; Kasari-Toussaint, L.; Karise, R.; Tiitsaar, A.; Soon, V.; Kupper, T.; Meriste, M.; Ingerpuu, N.; et al. Restoration of Semi-Natural Grasslands Boosts Biodiversity and Re-Creates Hotspots for Ecosystem Services. Agric. Ecosyst. Environ. 2024, 374, 109139. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szentes, S.; Penksza, K.; Saláta-Falusi, E.; Sipos, L.; Kozma-Bognár, V.; Hoffmann, R.; Wagenhoffer, Z. Effects of Bothriochloa ischaemum on the Diversity of Pannonian Sandy Grasslands. Land 2025, 14, 1107. https://doi.org/10.3390/land14051107
Szentes S, Penksza K, Saláta-Falusi E, Sipos L, Kozma-Bognár V, Hoffmann R, Wagenhoffer Z. Effects of Bothriochloa ischaemum on the Diversity of Pannonian Sandy Grasslands. Land. 2025; 14(5):1107. https://doi.org/10.3390/land14051107
Chicago/Turabian StyleSzentes, Szilárd, Károly Penksza, Eszter Saláta-Falusi, László Sipos, Veronika Kozma-Bognár, Richárd Hoffmann, and Zsombor Wagenhoffer. 2025. "Effects of Bothriochloa ischaemum on the Diversity of Pannonian Sandy Grasslands" Land 14, no. 5: 1107. https://doi.org/10.3390/land14051107
APA StyleSzentes, S., Penksza, K., Saláta-Falusi, E., Sipos, L., Kozma-Bognár, V., Hoffmann, R., & Wagenhoffer, Z. (2025). Effects of Bothriochloa ischaemum on the Diversity of Pannonian Sandy Grasslands. Land, 14(5), 1107. https://doi.org/10.3390/land14051107