
Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring

 ,
,  , and
, and 
Abstract
1. Introduction
2. Methods
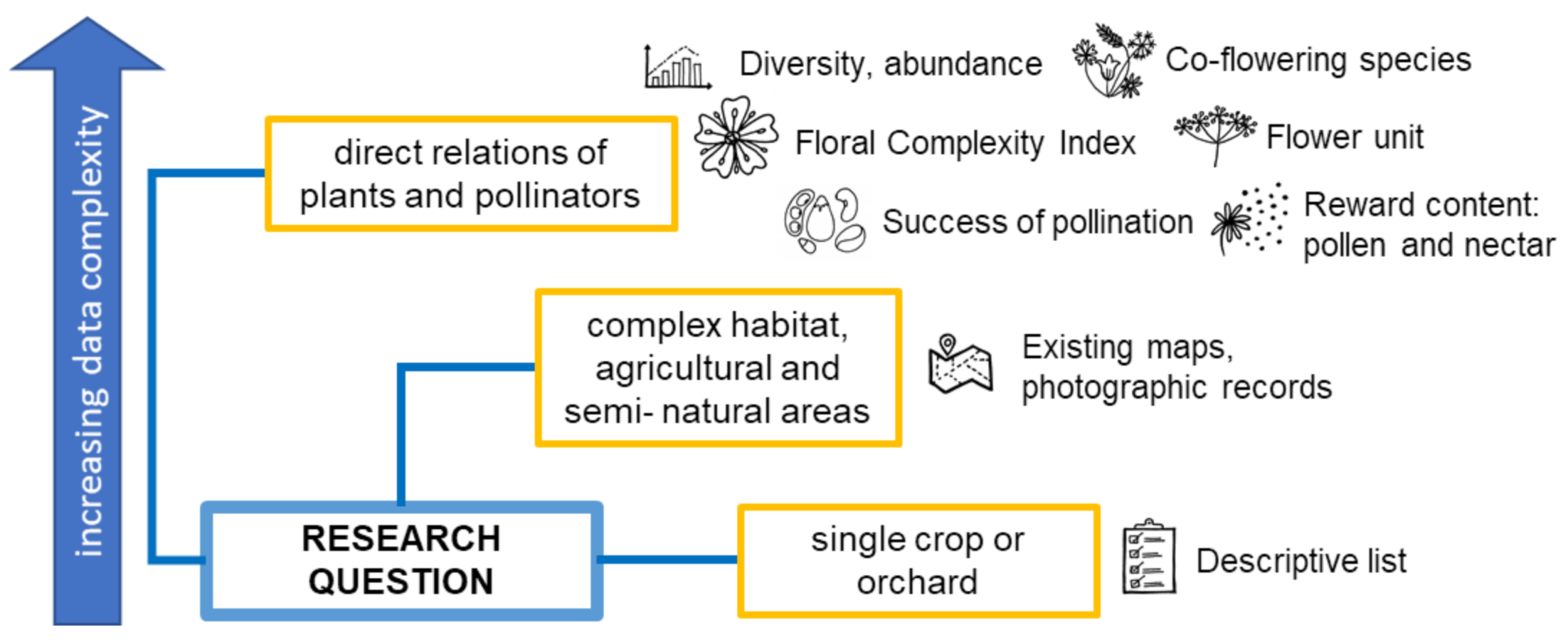
3. Leading Role of the Research Question Perspective
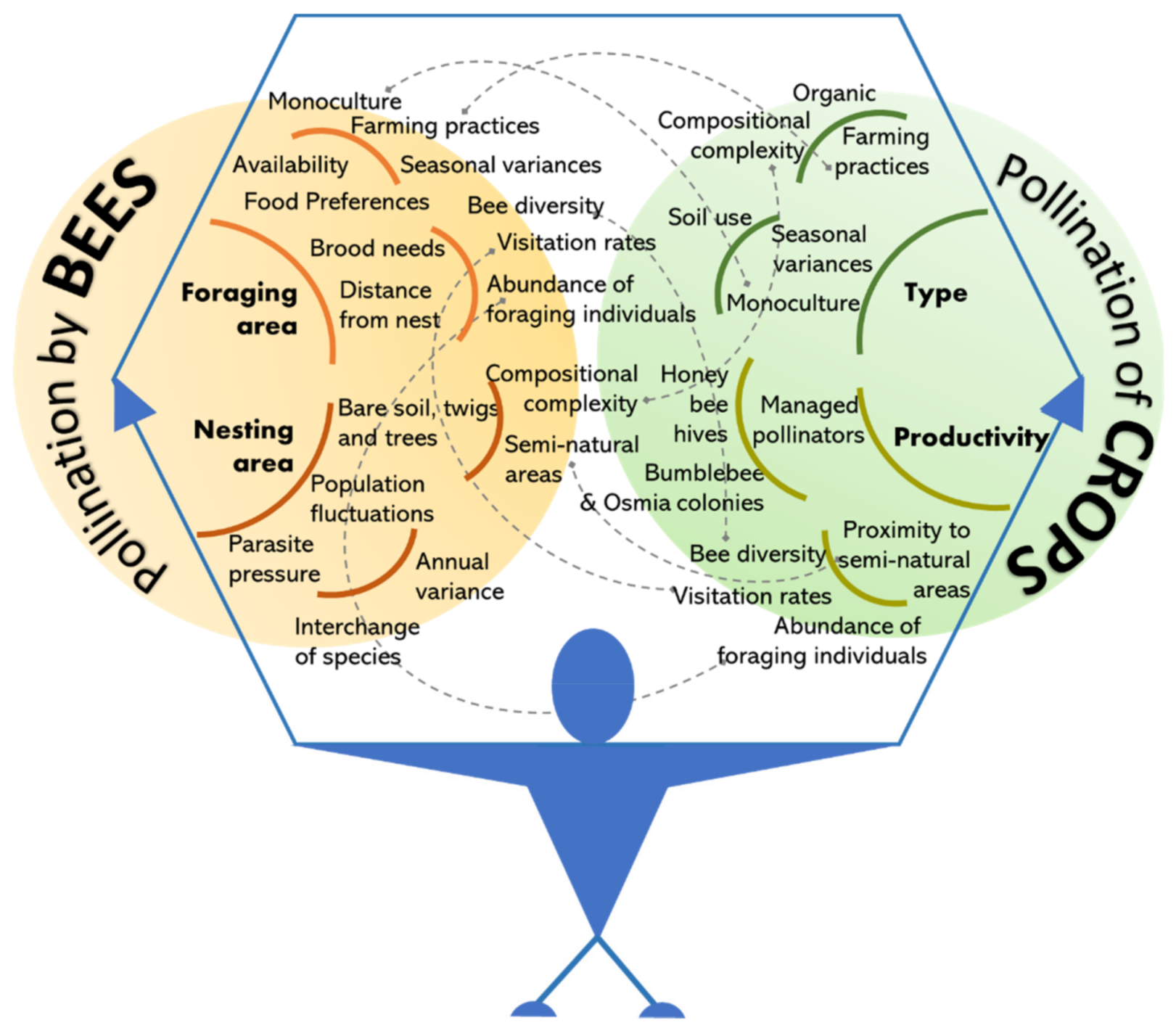
3.1. Investigating Pollination by Bees
3.2. Investigating Pollination of Crops/Orchards
3.3. Integrating Pollinators and Plants
4. Addressing the Increasing Power of Landscape Analysis
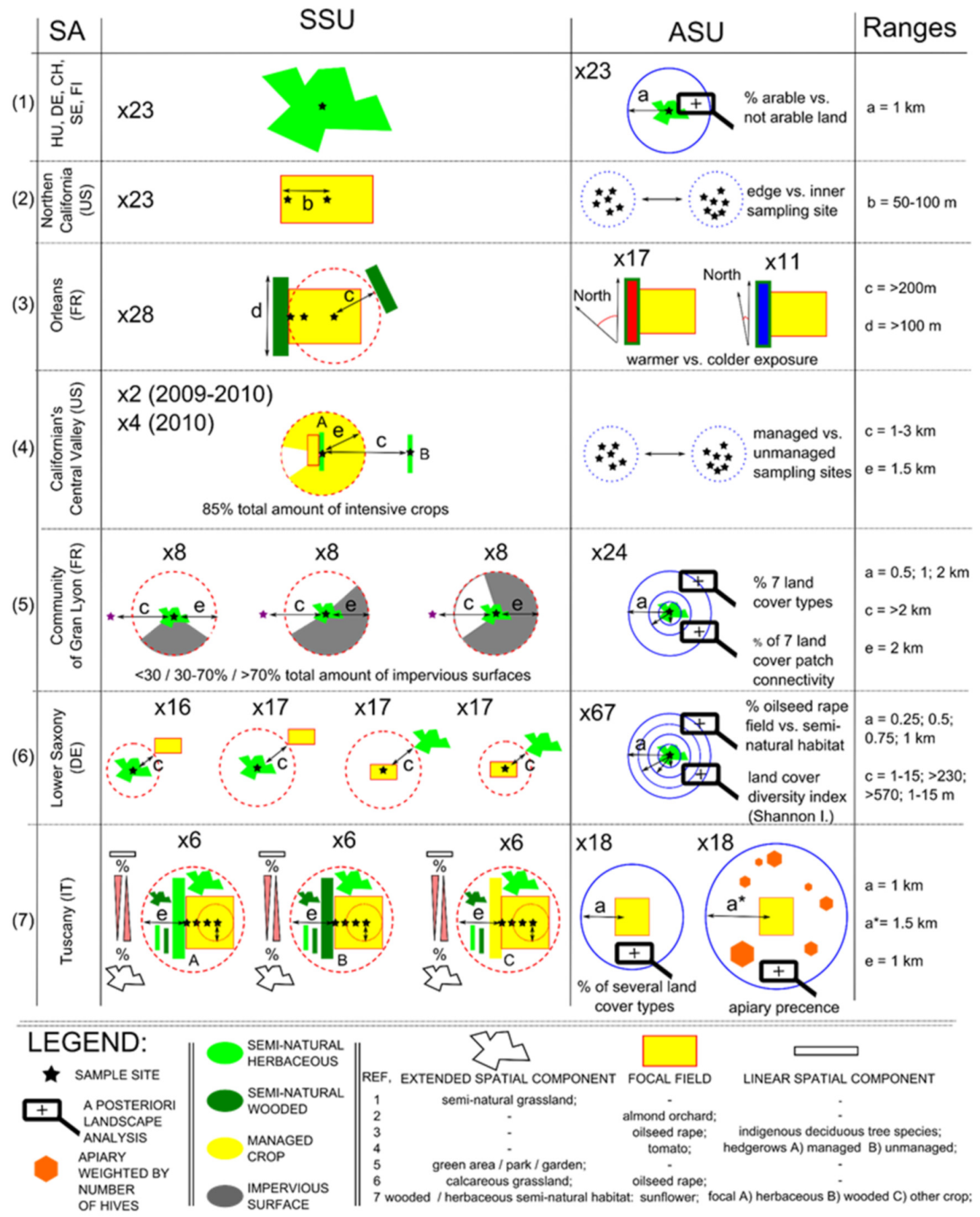
4.1. Study Area [SA] and Specific Spatial Unit [SSU]
4.2. Advanced Spatial Units [ASUs]
4.3. Predictive Power
5. Addressing the Neglected Vegetation
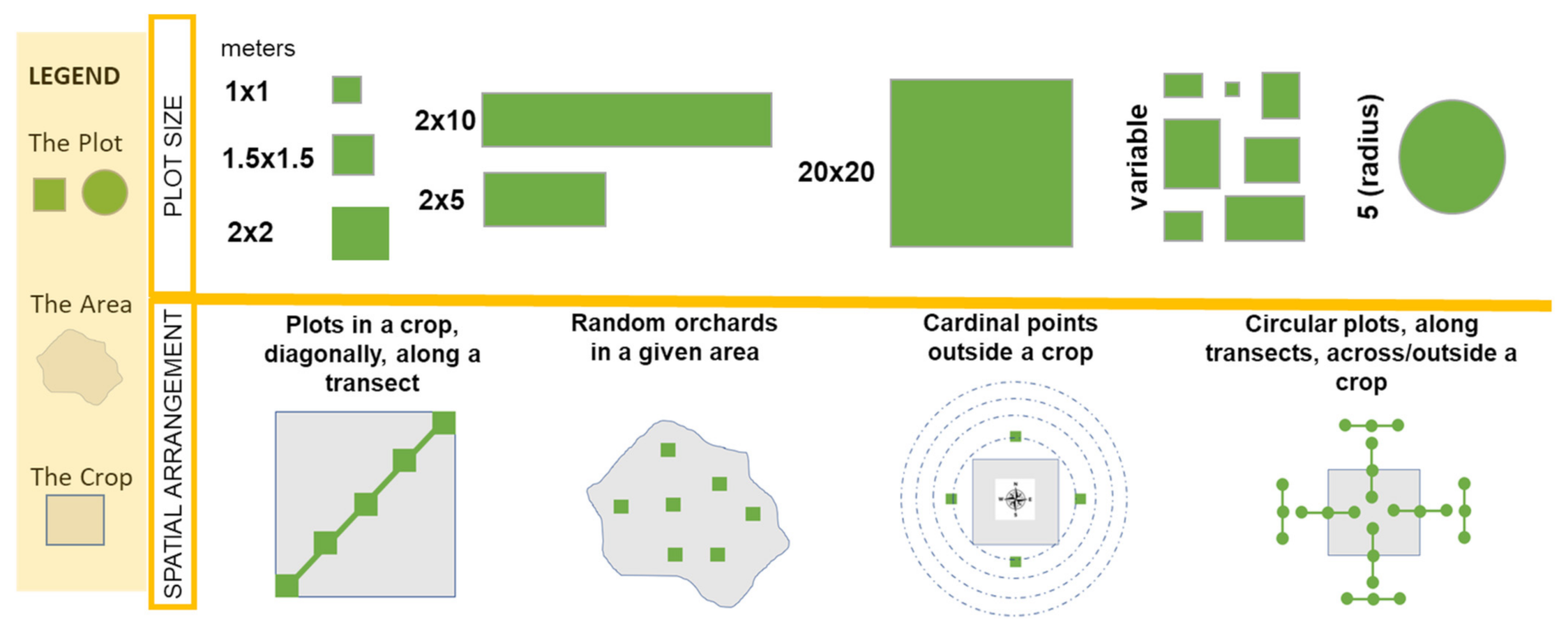
5.1. Plots for Pollination Studies
5.2. Flower Traits
5.3. Pollination Success
6. Addressing the Key Players: Pollinators
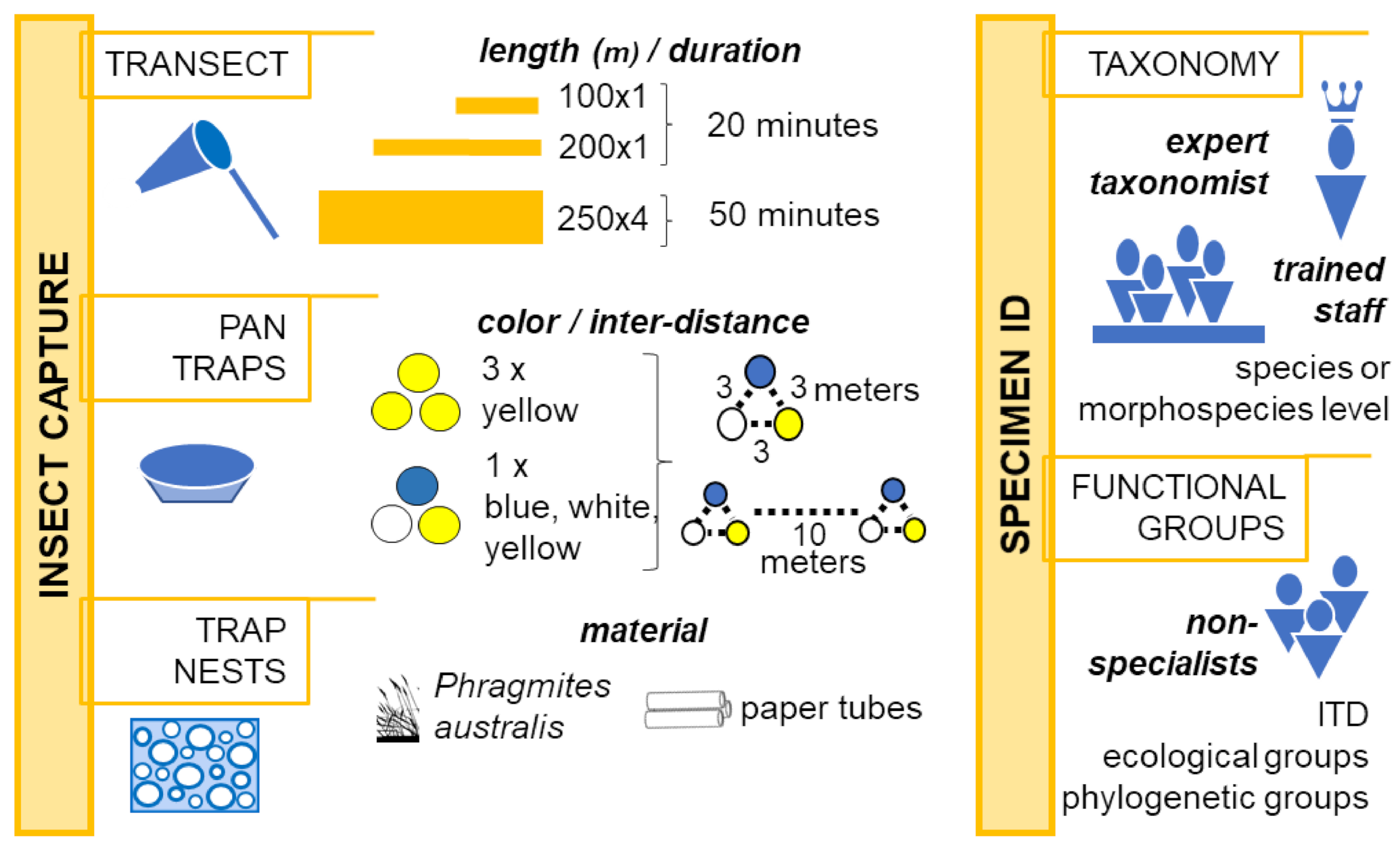
6.1. Capture
6.2. A Bottleneck: The Taxonomy
6.3. Plant-Pollinator Interactions
7. Discussion and Conclusions: Emerging Key Points
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Comes, H.P. The Mediterranean region—A hotspot for plant biogeographic research. New Phytol. 2004, 164, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Rojas, J.; Pinto-Correia, T.; Napoleone, C. Farm and land system dynamics in the Mediterranean: Integrating different spatial-temporal scales and management approaches. Land Use Policy 2019, 88, 104082. [Google Scholar] [CrossRef]
- Lomba, A.; Alves, P.; Jongman, R.H.G.; Mccracken, D.I. Reconciling nature conservation and traditional farming practices: A spatially explicit framework to assess the extent of High Nature Value farmlands in the European countryside. Ecol. Evol. 2015, 5, 1031–1044. [Google Scholar] [CrossRef]
- Dernini, S.; Berry, E.M.; Serra-Majem, L.; La Vecchia, C.; Capone, R.; Medina, F.X.; Aranceta-Bartrina, J.; Belahsen, R.; Burlingame, B.; Calabrese, G.; et al. Med Diet 4.0: The Mediterranean diet with four sustainable benefits. Public Health Nutr. 2017, 20, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Gallai, N.; Garibaldi, L.A.; Kuhlmann, M.; Klein, A.M. Economic gain, stability of pollination and bee diversity decrease from southern to northern Europe. Basic Appl. Ecol. 2013, 14, 461–471. [Google Scholar] [CrossRef]
- Scherber, C.; Beduschi, T.; Tscharntke, T. Novel approaches to sampling pollinators in whole landscapes: A lesson for landscape-wide biodiversity monitoring. Landsc. Ecol. 2019, 34. [Google Scholar] [CrossRef]
- Tamburini, G.; Bommarco, R.; Kleijn, D.; van der Putten, W.H.; Marini, L. Pollination contribution to crop yield is often context-dependent: A review of experimental evidence. Agric. Ecosyst. Environ. 2019, 280, 16–23. [Google Scholar] [CrossRef]
- Willis, K.J.; Jeffers, E.S.; Tovar, C.; Long, P.R.; Caithness, N.; Smit, M.G.D.; Hagemann, R.; Collin-Hansen, C.; Weissenberger, J. Determining the ecological value of landscapes beyond protected areas. Biol. Conserv. 2012, 147, 3–12. [Google Scholar] [CrossRef]
- Oguro, M.; Taki, H.; Konuma, A.; Uno, M.; Nakashizuka, T. Importance of national or regional specificity in the relationship between pollinator dependence and production stability. Sustain. Sci. 2019, 14, 139–146. [Google Scholar] [CrossRef]
- Mayer, C.; Adler, L.; Armbruster, S.; Dafni, A.; Eardley, C.; Huang, S.-Q.; Kevan, P.G.; Ollerton, J.; Packer, L.; Ssymank, A.; et al. Pollination ecology in the 21st Century: Key questions for future research. J. Pollinat. Ecol. 2011, 3. [Google Scholar] [CrossRef]
- Holzschuh, A.; Dormann, C.F.; Tscharntke, T.; Steffan-Dewenter, I. Mass-flowering crops enhance wild bee abundance. Oecologia 2013, 172, 477–484. [Google Scholar] [CrossRef]
- Brittain, C.; Kremen, C.; Klein, A.M. Biodiversity buffers pollination from changes in environmental conditions. Glob. Chang. Biol. 2013, 19, 540–547. [Google Scholar] [CrossRef]
- Coudrain, V.; Rittiner, S.; Herzog, F.; Tinner, W.; Entling, M.H. Landscape distribution of food and nesting sites affect larval diet and nest size, but not abundance of Osmia bicornis. Insect Sci. 2016, 23, 746–753. [Google Scholar] [CrossRef]
- Rosanigo, M.P.; Marrero, H.J.; Torretta, J.P. Limiting resources on the reproductive success of a cavity-nesting bee species in a grassland agroecosystem. J. Apic. Res. 2020, 59, 583–591. [Google Scholar] [CrossRef]
- Beduschi, T.; Kormann, U.G.; Tscharntke, T.; Scherber, C. Spatial community turnover of pollinators is relaxed by semi-natural habitats, but not by mass-flowering crops in agricultural landscapes. Biol. Conserv. 2018, 221, 59–66. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Münzenberg, U.; Bürger, C.; Thies, C.; Tscharntke, T. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 2002, 83. [Google Scholar] [CrossRef]
- Grass, I.; Berens, D.G.; Farwig, N. Natural habitat loss and exotic plants reduce the functional diversity of flower visitors in a heterogeneous subtropical landscape. Funct. Ecol. 2014, 28, 1117–1126. [Google Scholar] [CrossRef]
- Martins, K.T.; Gonzalez, A.; Lechowicz, M.J. Pollination services are mediated by bee functional diversity and landscape context. Agric. Ecosyst. Environ. 2015, 200. [Google Scholar] [CrossRef]
- Bailey, S.; Requier, F.; Nusillard, B.; Roberts, S.P.M.; Potts, S.G.; Bouget, C. Distance from forest edge affects bee pollinators in oilseed rape fields. Ecol. Evol. 2014, 4, 370–380. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Garratt, M.P.D.; Powney, G.D.; Shaw, R.F.; Osborne, J.L.; Soroka, J.; Lindström, S.A.M.; Stanley, D.; Ouvrard, P.; Edwards, M.E.; et al. Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nat. Commun. 2019, 10, 1481. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, C.C.; Koh, I.; Richardson, L.L.; Beauchemin, A.; Ricketts, T.H. Farm and landscape factors interact to affect the supply of pollination services. Agric. Ecosyst. Environ. 2017, 250, 113–122. [Google Scholar] [CrossRef]
- Shuler, R.E.; Roulston, T.H.; Farris, G.E. Farming practices influence wild pollinator populations on squash and pumpkin. J. Econ. Entomol. 2005, 98, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Kratschmer, S.; Pachinger, B.; Schwantzer, M.; Paredes, D.; Guernion, M.; Burel, F.; Nicolai, A.; Strauss, P.; Bauer, T.; Kriechbaum, M.; et al. Tillage intensity or landscape features: What matters most for wild bee diversity in vineyards? Agric. Ecosyst. Environ. 2018, 266, 142–152. [Google Scholar] [CrossRef]
- Clough, Y.; Ekroos, J.; Báldi, A.; Batáry, P.; Bommarco, R.; Gross, N.; Holzschuh, A.; Hopfenmüller, S.; Knop, E.; Kuussaari, M.; et al. Density of insect-pollinated grassland plants decreases with increasing surrounding land-use intensity. Ecol. Lett. 2014, 17, 1168–1177. [Google Scholar] [CrossRef]
- Albor, C.; García-Franco, J.G.; Parra-Tabla, V.; Díaz-Castelazo, C.; Arceo-Gómez, G. Taxonomic and functional diversity of the co-flowering community differentially affect Cakile edentula pollination at different spatial scales. J. Ecol. 2019, 107, 2167–2181. [Google Scholar] [CrossRef]
- Fründ, J.; Linsenmair, K.E.; Blüthgen, N. Pollinator diversity and specialization in relation to flower diversity. Oikos 2010, 119, 1581–1590. [Google Scholar] [CrossRef]
- Uyttenbroeck, R.; Piqueray, J.; Hatt, S.; Mahy, G.; Monty, A. Increasing plant functional diversity is not the key for supporting pollinators in wildflower strips. Agric. Ecosyst. Environ. 2017, 249, 144–155. [Google Scholar] [CrossRef]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing abundance, increasing diversity and changing structure of the wild bee community (hymenoptera: Anthophila) along an urbanization gradient. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Hopfenmüller, S.; Steffan-Dewenter, I.; Holzschuh, A. Trait-specific responses of wild bee communities to landscape composition, configuration and local factors. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2014, 189, 164–170. [Google Scholar] [CrossRef]
- Franzén, M.; Schweiger, O.; Betzholtz, P.E. Species-area relationships are controlled by species traits. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Riedinger, V.; Renner, M.; Rundlöf, M.; Steffan-Dewenter, I.; Holzschuh, A. Early mass-flowering crops mitigate pollinator dilution in late-flowering crops. Landsc. Ecol. 2014, 29, 425–435. [Google Scholar] [CrossRef]
- Ekroos, J.; Rundlöf, M.; Smith, H.G. Trait-dependent responses of flower-visiting insects to distance to semi-natural grasslands and landscape heterogeneity. Landsc. Ecol. 2013, 28, 1283–1292. [Google Scholar] [CrossRef]
- Bartual, A.M.; Bocci, G.; Marini, S.; Moonen, A.C. Local and landscape factors affect sunflower pollination in a Mediterranean agroecosystem. PLoS ONE 2018, 13, e0203990. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Kleijn, D.; Tscharntke, T. Diversity of flower-visiting bees in cereal fields: Effects of farming system, landscape composition and regional context. J. Appl. Ecol. 2007, 44, 41–49. [Google Scholar] [CrossRef]
- Senapathi, D.; Carvalheiro, L.G.; Biesmeijer, J.C.; Dodson, C.A.; Evans, R.L.; McKerchar, M.; Morton, D.R.; Moss, E.D.; Roberts, S.P.M.; Kunin, W.E.; et al. The impact of over 80 years of land cover changes on bee and wasp pollinator communities in England. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Keil, P.; Schweiger, O.; Kühn, I.; Kunin, W.E.; Kuussaari, M.; Settele, J.; Henle, K.; Brotons, L.; Pe’er, G.; Lengyel, S.; et al. Patterns of beta diversity in Europe: The role of climate, land cover and distance across scales. J. Biogeogr. 2012, 39, 1473–1486. [Google Scholar] [CrossRef]
- Polce, C.; Termansen, M.; Aguirre-Gutiérrez, J.; Boatman, N.D.; Budge, G.E.; Crowe, A.; Garratt, M.P.; Pietravalle, S.; Potts, S.G.; Ramirez, J.A.; et al. Species Distribution Models for Crop Pollination: A Modelling Framework Applied to Great Britain. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Tiedeken, E.J.; Stout, J.C. Insect-flower interaction network structure is resilient to a temporary pulse of floral resources from invasive Rhododendron ponticum. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Bartomeus, I.; Dietzsch, A.C.; Petanidou, T.; Steffan-Dewenter, I.; Stout, J.C.; Tscheulin, T. Invasive plant integration into native plant-pollinator networks across Europe. Proc. Biol. Sci. 2009, 276, 3887–3893. [Google Scholar] [CrossRef] [PubMed]
- Dengler, J.; Chytrý, M.; Ewald, J. Phytosociology. In International Encyclopedia of Geography: People, the Earth, Environment and Technology; Jörgensen, S.E., Fath, B.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 2767–2779, Chapter in Encyclopedia of, ecology. [Google Scholar] [CrossRef]
- Ouvrard, P.; Jacquemart, A.L. Review of methods to investigate pollinator dependency in oilseed rape (Brassica napus). F. Crop. Res. 2019, 231, 18–29. [Google Scholar] [CrossRef]
- Mallinger, R.E.; Bradshaw, J.; Varenhorst, A.J.; Prasifka, J.R. Native Solitary Bees Provide Economically Significant Pollination Services to Confection Sunflowers (Helianthus annuus L.) (Asterales: Asteraceae) Grown Across the Northern Great Plains. J. Econ. Entomol. 2019, 112, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.W.; Stanley-Stahr, C.; Bammer, M.; Daniels, J.C.; Ellis, J.D. Contribution of bees and other pollinators to watermelon (Citrullus lanatus Thunb.) pollination. J. Apic. Res. 2019, 58, 597–603. [Google Scholar] [CrossRef]
- Pisanty, G.; Afik, O.; Wajnberg, E.; Mandelik, Y. Watermelon pollinators exhibit complementarity in both visitation rate and single-visit pollination efficiency. J. Appl. Ecol. 2016, 53, 360–370. [Google Scholar] [CrossRef]
- Giovanetti, M. Do bees like olive? A preliminary analysis of honey bee behaviour on flowers of the wind-pollinated species Olea europea. Acta Hortic. 2018, 1199, 121–126. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape effects on crop pollination services: Are there general patterns? Ecol. Lett. 2008, 11. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Bennett, A.B.; Lovell, S. Landscape and local site variables differentially influence pollinators and pollination services in urban agricultural sites. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Frankie, G.W.; Thorp, R.W.; Hernandez, J.; Rizzardi, M.; Pawelek, J.C.; Witt, S.L.; Coville, R.; Wojcik, V.A. Native bees are a rich natural resource in urban California gardens. Calif. Agric. 2009, 63, 113–120. [Google Scholar] [CrossRef]
- Geslin, B.; Le Féon, V.; Folschweiller, M.; Flacher, F.; Carmignac, D.; Motard, E.; Perret, S.; Dajoz, I. The proportion of impervious surfaces at the landscape scale structures wild bee assemblages in a densely populated region. Ecol. Evol. 2016, 6. [Google Scholar] [CrossRef]
- Lerman, S.B.; Milam, J. Bee Fauna and Floral Abundance Within Lawn-Dominated Suburban Yards in Springfield, MA. Ann. Entomol. Soc. Am. 2016, 109. [Google Scholar] [CrossRef]
- Jha, S.; Kremen, C. Resource diversity and landscape-level homogeneity drive native bee foraging. Proc. Natl. Acad. Sci. USA 2013, 110. [Google Scholar] [CrossRef]
- Ebeling, A.; Klein, A.M.; Weisser, W.W.; Tscharntke, T. Multitrophic effects of experimental changes in plant diversity on cavity-nesting bees, wasps, and their parasitoids. Oecologia 2012, 169. [Google Scholar] [CrossRef]
- Nielsen, A.; Steffan-Dewenter, I.; Westphal, C.; Messinger, O.; Potts, S.G.; Roberts, S.P.M.; Settele, J.; Szentgyörgyi, H.; Vaissière, B.E.; Vaitis, M.; et al. Assessing bee species richness in two Mediterranean communities: Importance of habitat type and sampling techniques. Ecol. Res. 2011, 26, 969–983. [Google Scholar] [CrossRef]
- Ramos–Jiliberto, R.; Moisset de Espanés, P.; Franco-Cisterna, M.; Petanidou, T.; Vázquez, D.P. Phenology determines the robustness of plant–pollinator networks. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Fantinato, E.; Del Vecchio, S.; Giovanetti, M.; Acosta, A.T.R.; Buffa, G. New insights into plants co-existence in species-rich communities: The pollination interaction perspective. J. Veg. Sci. 2018, 29. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E.S. Evaluating the dependence of urban pollinators on ornamental, non-native, and ‘weedy’ floral resources. Urban Ecosyst. 2019, 22, 293–302. [Google Scholar] [CrossRef]
- Pellissier, L.; Pottier, J.; Vittoz, P.; Dubuis, A.; Guisan, A. Spatial pattern of floral morphology: Possible insight into the effects of pollinators on plant distributions. Oikos 2010, 119, 1805–1813. [Google Scholar] [CrossRef]
- Stefanaki, A.; Kantsa, A.; Tscheulin, T.; Charitonidou, M.; Petanidou, T. Lessons from red data books: Plant vulnerability increases with floral complexity. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Takkis, K.; Tscheulin, T.; Petanidou, T. Differential effects of climate warming on the nectar secretion of early-and late-flowering mediterranean plants. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, L.G.; Biesmeijer, J.C.; Benadi, G.; Fründ, J.; Stang, M.; Bartomeus, I.; Kaiser-Bunbury, C.N.; Baude, M.; Gomes, S.I.F.; Merckx, V.; et al. The potential for indirect effects between co-flowering plants via shared pollinators depends on resource abundance, accessibility and relatedness. Ecol. Lett. 2014, 17, 1389–1399. [Google Scholar] [CrossRef]
- Yamamoto, M.; Da Silva, C.I.; Augusto, S.C.; Barbosa, A.A.A.; Oliveira, P.E. The role of bee diversity in pollination and fruit set of yellow passion fruit (Passiflora edulis forma flavicarpa, Passifloraceae) crop in Central Brazil. Apidologie 2012, 43. [Google Scholar] [CrossRef]
- Jayasinghe, U.J.M.S.R.; Silva, T.H.S.E.; Karunaratne, W.A.I.P. Buzzing Wild Bee Visits Enhance Seed Set in Eggplant, Solanum melongena. Psyche (London) 2017, 2017. [Google Scholar] [CrossRef]
- Duan, Y.W.; Ren, H.; Li, T.; Wang, L.L.; Zhang, Z.Q.; Tu, Y.L.; Yang, Y.P. A century of pollination success revealed by herbarium specimens of seed pods. New Phytol. 2019, 224, 1512–1517. [Google Scholar] [CrossRef]
- Ballantyne, G.; Baldock, K.C.R.; Rendell, L.; Willmer, P.G. Pollinator importance networks illustrate the crucial value of bees in a highly speciose plant community. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- Jauker, F.; Bondarenko, B.; Becker, H.C.; Steffan-Dewenter, I. Pollination efficiency of wild bees and hoverflies provided to oilseed rape. Agric. For. Entomol. 2012, 14. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48. [Google Scholar] [CrossRef]
- Petanidou, T.; Kallimanis, A.S.; Tzanopoulos, J.; Sgardelis, S.P.; Pantis, J.D. Long-term observation of a pollination network: Fluctuation in species and interactions, relative invariance of network structure and implications for estimates of specialization. Ecol. Lett. 2008, 11. [Google Scholar] [CrossRef]
- Uhl, P.; Brühl, C.A. The Impact of Pesticides on Flower-Visiting Insects: A Review with Regard to European Risk Assessment. Environ. Toxicol. Chem. 2019, 38, 2355–2370. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef]
- Potts, S.G.; Petanidou, T.; Roberts, S.; O’Toole, C.; Hulbert, A.; Willmer, P. Plant-pollinator biodiversity and pollination services in a complex Mediterranean landscape. Biol. Conserv. 2006, 129, 519–529. [Google Scholar] [CrossRef]
- Ropars, L.; Affre, L.; Schurr, L.; Flacher, F.; Genoud, D.; Mutillod, C.; Geslin, B. Land cover composition, local plant community composition and honeybee colony density affect wild bee species assemblages in a Mediterranean biodiversity hot-spot. Acta Oecologica 2020, 104. [Google Scholar] [CrossRef]
- McCravy, K.W. A review of sampling and monitoring methods for beneficial arthropods in agroecosystems. Insects 2018, 9. [Google Scholar] [CrossRef]
- Sheffield, C.S.; Pindar, A.; Packer, L.; Kevan, P.G. The potential of cleptoparasitic bees as indicator taxa for assessing bee communities. Apidologie 2013, 44. [Google Scholar] [CrossRef]
- Gervais, A.; Fournier, V.; Sheffield, C.S.; Chagnon, M. Assessing wild bee biodiversity in cranberry agroenvironments: Influence of natural habitats. J. Econ. Entomol. 2017, 110. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P. To enrich or not to enrich? Are there any benefits of using multiple colors of pan traps when sampling aculeate Hymenoptera? J. Insect Conserv. 2014, 18, 1123–1136. [Google Scholar] [CrossRef]
- Lazarina, M.; Devalez, J.; Neokosmidis, L.; Sgardelis, S.P.; Kallimanis, A.S.; Tscheulin, T.; Tsalkatis, P.; Kourtidou, M.; Mizerakis, V.; Nakas, G.; et al. Moderate fire severity is best for the diversity of most of the pollinator guilds in Mediterranean pine forests. Ecology 2019, 100. [Google Scholar] [CrossRef]
- Droege, S.; Tepedino, V.J.; Lebuhn, G.; Link, W.; Minckley, R.L.; Chen, Q.; Conrad, C. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Broussard, M.; Rao, S.; Stephen, W.P.; White, L. Native bees, Honeybees, and pollination in Oregon cranberries. HortScience 2011, 46, 885–888. [Google Scholar] [CrossRef]
- Hall, M.A.; Reboud, E.L. High sampling effectiveness for non-bee flower visitors using vane traps in both open and wooded habitats. Austral Entomol. 2019, 58. [Google Scholar] [CrossRef]
- Popic, T.J.; Davila, Y.C.; Wardle, G.M. Evaluation of Common Methods for Sampling Invertebrate Pollinator Assemblages: Net Sampling Out-Perform Pan Traps. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Rhoades, P.; Griswold, T.; Waits, L.; Bosque-Pérez, N.A.; Kennedy, C.M.; Eigenbrode, S.D. Sampling technique affects detection of habitat factors influencing wild bee communities. J. Insect Conserv. 2017, 21, 703–714. [Google Scholar] [CrossRef]
- Wilson, J.S.; Jahner, J.P.; Starley, L.; Calvin, C.L.; Ikerd, H.; Griswold, T. Sampling bee communities using pan traps: Alternative methods increase sample size. J. Insect Conserv. 2016, 20, 919–922. [Google Scholar] [CrossRef]
- Gezon, Z.J.; Wyman, E.S.; Ascher, J.S.; Inouye, D.W.; Irwin, R.E. The effect of repeated, lethal sampling on wild bee abundance and diversity. Methods Ecol. Evol. 2015, 6, 1044–1054. [Google Scholar] [CrossRef]
- Wägele, H.; Klussmann-Kolb, A.; Kuhlmann, M.; Haszprunar, G.; Lindberg, D.; Koch, A.; Wägele, J.W. The taxonomist—An endangered race. A practical proposal for its survival. Front. Zool. 2011, 8, 25. [Google Scholar] [CrossRef]
- Cane, J.H. Estimation of Bee Size Using Intertegular Span (Apoidea). J. Kansas Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Normandin, É.; Vereecken, N.J.; Buddle, C.M.; Fournier, V. Taxonomic and functional trait diversity of wild bees in different urban settings. PeerJ 2017, 2017. [Google Scholar] [CrossRef]
- De Palma, A.; Kuhlmann, M.; Roberts, S.P.M.; Potts, S.G.; Börger, L.; Hudson, L.N.; Lysenko, I.; Newbold, T.; Purvis, A. Ecological traits affect the sensitivity of bees to land-use pressures in European agricultural landscapes. J. Appl. Ecol. 2015, 52. [Google Scholar] [CrossRef]
- Ekroos, J.; Kuussaari, M. Landscape context affects the relationship between local and landscape species richness of butterflies in semi-natural habitats. Ecography 2012, 35. [Google Scholar] [CrossRef]
- Giovanetti, M.; Giuliani, C.; Boff, S.; Fico, G.; Lupi, D. A botanic garden as a tool to combine public perception of nature and life-science investigations on native/exotic plants interactions with local pollinators. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Stewart, A.B.; Waitayachart, P. Year-round temporal stability of a tropical, urban plant-pollinator network. PLoS ONE 2020, 15, 1–12. [Google Scholar] [CrossRef]
- MacIvor, J.S. Cavity-nest boxes for solitary bees: A century of design and research. Apidologie 2017, 48, 311–327. [Google Scholar] [CrossRef]
- Loyola, R.D.; Martins, R.P. Trap-nest occupation by solitary wasps and bees (Hymenoptera: Aculeata) in a forest urban remanent. Neotrop. Entomol. 2006, 35, 41–48. [Google Scholar] [CrossRef]
- Pereira-Peixoto, M.H.; Pufal, G.; Martins, C.F.; Klein, A.M. Spillover of trap-nesting bees and wasps in an urban–rural interface. J. Insect Conserv. 2014, 18, 815–826. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Cabral, J.M.; Packer, L. Pollen specialization by solitary bees in an urban landscape. Urban Ecosyst. 2014, 17, 139–147. [Google Scholar] [CrossRef]
- Berger, G.; Kaechele, H.; Pfeffer, H. The greening of the European common agricultural policy by linking the European-wide obligation of set-aside with voluntary agri-environmental measures on a regional scale. Environ. Sci. Policy 2006, 9. [Google Scholar] [CrossRef]
- Russo, L.; Miller, A.D.; Tooker, J.; Bjornstad, O.N.; Shea, K. Quantitative evolutionary patterns in bipartite networks: Vicariance, phylogenetic tracking or diffuse co-evolution? Methods Ecol. Evol. 2018, 9, 761–772. [Google Scholar] [CrossRef]
- Litman, J.R.; Danforth, B.N.; Eardley, C.D.; Praz, C.J. Why do leafcutter bees cut leaves? New insights into the early evolution of bees. Proc. R. Soc. B Biol. Sci. 2011, 278, 3593–3600. [Google Scholar] [CrossRef] [PubMed]
- Angelella, G.M.; McCullough, C.T.; O’Rourke, M.E. Honey bee hives decrease wild bee abundance, species richness, and fruit count on farms regardless of wildflower strips. Sci. Rep. 2021, 11, 3202. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M. Gradual replacement of wild bees by honeybees in flowers of the Mediterranean Basin over the last 50 years. Proc. R. Soc. B Biol. Sci. 2020, 287, 16–20. [Google Scholar] [CrossRef]
- Baños-Picón, L.; Torres, F.; Tormos, J.; Gayubo, S.F.; Asís, J.D. Comparison of two Mediterranean crop systems: Polycrop favours trap-nesting solitary bees over monocrop. Basic Appl. Ecol. 2013, 14, 255–262. [Google Scholar] [CrossRef]
- Bartholomée, O.; Aullo, A.; Becquet, J.; Vannier, C.; Lavorel, S. Pollinator presence in orchards depends on landscape-scale habitats more than in-field flower resources. Agric. Ecosyst. Environ. 2020, 293, 106806. [Google Scholar] [CrossRef]
- Balzan, M.V.; Caruana, J.; Zammit, A. Assessing the capacity and flow of ecosystem services in multifunctional landscapes: Evidence of a rural-urban gradient in a Mediterranean small island state. Land Use Policy 2018, 75, 711–725. [Google Scholar] [CrossRef]
- Dainese, M.; Montecchiari, S.; Sitzia, T.; Sigura, M.; Marini, L. High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. J. Appl. Ecol. 2017, 54, 380–388. [Google Scholar] [CrossRef]
- Bassa, M.; Chamorro, L.; José-María, L.; Blanco-Moreno, J.M.; Sans, F.X. Factors affecting plant species richness in field boundaries in the Mediterranean region. Biodivers. Conserv. 2012, 21, 1101–1114. [Google Scholar] [CrossRef]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Dorchin, A.; Dafni, A.; Izhaki, I.; Sapir, Y.; Vereecken, N.J. Patterns and drivers of wild bee community assembly in a Mediterranean IUCN important plant area. Biodivers. Conserv. 2018, 27, 695–717. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conceptual Framework | the Research Question Perspective | Increasing Power of Landscape Analysis | the Neglected Vegetation | Pollinators |
| usually has a one-way workflow | needs to select the appropriate spatial unit | is a key feature of the landscape and vital for pollinators | managed and wild bees are the best-known pollinators | |
| Research Approaches | goal: pollination of crops (managed pollinators, soil use, field margins) | SA: the study area | crop or orchards + spontaneous flora | insect capture in the field |
| goal: pollination by bees (nesting + foraging areas) | SSU: the specific spatial unit containing the field site(s) | complex habitat or direct plant-pollinator relations | specimen identification in the lab | |
| similar data irrespective of goal | ASU: advanced spatial unit for a posteriori analysis | plots + buffers flower traits | walking transects, pan traps and/or observation plots | |
| Current/Future Expectations | assembling previous results irrespective of how they were gathered | predicting pollinators based on landscape features | standardizing plot size and spatial arrangement | increasing taxonomic identification and behavioral observations |
| including the evolutionary history of pollinators and plants | including landscape analysis as a future pre-requisite for monitoring studies | reminding the precise relationship between a plant species and its flower visitors | correlating functional traits, taxonomy and landscape features |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovanetti, M.; Albertazzi, S.; Flaminio, S.; Ranalli, R.; Bortolotti, L.; Quaranta, M. Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring. Land 2021, 10, 540. https://doi.org/10.3390/land10050540
Giovanetti M, Albertazzi S, Flaminio S, Ranalli R, Bortolotti L, Quaranta M. Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring. Land. 2021; 10(5):540. https://doi.org/10.3390/land10050540
Chicago/Turabian StyleGiovanetti, Manuela, Sergio Albertazzi, Simone Flaminio, Rosa Ranalli, Laura Bortolotti, and Marino Quaranta. 2021. "Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring" Land 10, no. 5: 540. https://doi.org/10.3390/land10050540
APA StyleGiovanetti, M., Albertazzi, S., Flaminio, S., Ranalli, R., Bortolotti, L., & Quaranta, M. (2021). Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring. Land, 10(5), 540. https://doi.org/10.3390/land10050540