Abstract
This study investigated bacterial transport in a two-dimensional (2-D) tank to evaluate the bacterial behavior of Legionella pneumophila as compared to Escherichia coli under saturated flow to simulate aquifer conditions. The experiments were performed in a 2-D tank packed with 3700 in3 (60,632 cm3) of commercially available bagged play sand under saturated conditions. The tank was disinfected by backwashing with 10% chlorine solution and subsequently neutralized by backwashing with tap water containing sodium thiosulphate (Na2S2O3) to ensure no chlorine residual. Bacterial transport was measured using samples collected from ports located at vertical transport distances of 5, 15 and 25 inches (12.7, 38.1 and 63.5 cm, respectively) below the sand surface along two vertical sections in the tank. An influent concentration of 105 CFU/mL was used for bacterial cells and the vertical fluid transport rate was 10.3 in/day (26.2 cm/day), controlled using a peristaltic pump at the bottom outlet. Legionella breakthroughs were recorded at 8, 22 and 35 h for the ports on the right side and 9, 24 and 36 h for the ports on the left side, at 5, 15 and 25 inch depths, respectively. At the same depths, E. coli breakthroughs were recorded at 5, 17 and 30 h for the ports on the right side and 7, 19 and 31 h for the ports on the left sides. The delay in Legionella transport compared to E. coli is homologous to Legionella’s pleomorphic nature. This study provides evidence of the mobility of both E. coli and Legionella in saturated aquifer conditions at a scale more representative of actual aquifer conditions. This study also provides a substantive basis for the premise that cell characteristics affect transport characteristics under those conditions.
1. Introduction
Worldwide, many cities are heavily dependent on groundwater for their water supplies, with more than one billion people living in such locales. In particular, most large cities in arid and semiarid regions are highly dependent on extracting groundwater to meet their rising water demand. This practice is not restricted to arid regions; some large cities in humid regions (e.g., Tokyo, Osaka, Taipei, Manila, and Jakarta) also depend on large extractions of groundwater as part of their water supply [1]. In the United States (U.S.), over 150 million people are estimated to directly depend on groundwater for their water supply [2].
Consumption of untreated groundwater has been associated with increased risk of infection by Escherichia coli O157:H7 [3]. Microbiological and water quality data for 30 public water-supply wells in Worcester and Wicomico Counties, Maryland, showed bacterial presence in all wells at some point during a year-long sampling campaign [4]. In a nationwide study in the U.S., 15% of groundwater wells tested positive for a broad range of bacteria, including coliforms, enterococci, and Clostridium [5]. A study investigating microbial contamination in groundwater wells found that 29.6% (347/1174) of wells tested positive for fecal indicator bacteria (total coliform, E. coli, or fecal coliform) [6]. Meta-analysis of data from 12 international groundwater studies of 718 public drinking-water systems located in a range of hydrogeological settings found that 36%, 12%, 15%, 52% and 26% of wells positive for total coliform, E. coli, Enterococci, aerobic spores, and anaerobic spores, respectively [7].
Depleting groundwater aquifers are naturally and/or artificially replenished by infiltration and percolation, which may transport microbial and chemical contaminants. Pathogen transport in soil aquifers has been extensively studied using different groups of microbial surrogates. However, the relevance of using surrogate organism transport data for predicting the transport of pathogens, e.g., Legionella, is uncertain [8]. The factors contributing to the transport of pathogens to groundwater aquifers include hydrogeology and characteristics of microbial cells such as survival, size, shape and mobility [6]. The significance of these factors is further complicated by transient flow and transport conditions, including seasonal, annual and long-term variability in recharge as well as transient interactions between climatic factors [9]. In addition, disparities between longitudinal and lateral dispersion in aquifers and transport of contaminants have been reported [10,11]. In the context of the discussion above, the unique features of Legionella warrant investigation of its transport properties in aquifers.
Legionella is ubiquitous in water systems, and Legionella pneumophila is responsible for the majority of the waterborne disease outbreaks (drinking water and non-recreational) in the U.S. [12]. Legionella has a high proclivity for growing in reclaimed water [13] and is not only present in reclaimed water, but also surface water [14]. For example, Legionella was detected in 60% of the samples (3/5) collected from recharge basin samples in California [15,16]. In Arizona, which is among the leading states practicing groundwater recharge, the presence of Legionella is increasing the risk of exposure and public health concerns.
Legionella is a unique bacterium with high lipopolysaccharide content in the cell membrane [17] and a pleomorphic nature, which can potentially impact its fate and transport. Under normal conditions, Legionella is a Gram-negative bacterium measuring 2 to 20 μm, but it can transform both its size and shape under different environmental conditions. For example, Legionella can transform into a facultative intracellular stage, wherein it requires a host such as an amoeba, which serves as a protective shell for Legionella cells. This plasticity of Legionella cells warrants in-depth study of their transport through different types of aquifers.
The objective of this study was to investigate and compare the transport characteristics of Legionella (human opportunistic pathogen) and E. coli (bacterial surrogate) in a two-dimensional packed porous media tank under saturated conditions. These bacteria were selected based on the public health significance of Legionella and immense value of E. coli as a surrogate of bacterial pathogens (based on the availability of an extensive amount of historical data). This approach more closely resembled aquifer conditions with intrinsic, multidimensional heterogeneities in comparison to more common one-dimensional column testing.
2. Materials and Methods
2.1. Preparation of Bacterial Stocks
Legionella pneumophila (ATCC® 33153 TM) and E. coli (ATCC® 25922 TM) were obtained from the American Type Culture Collection (ATCC®, Rockville, MD, USA) and propagated using the methods recommended by the ATCC. The pure cultures of frozen stocks (stored at −80 °C) were thawed at 37 °C. A sterile inoculation loop was used to streak Legionella and E. coli onto their selective media: buffered charcoal yeast extract (BCYE, Thermo Fisher Scientific) for Legionella and brilliant green agar (Sigma Aldrich, St Louis, MO, USA) for E. coli. Legionella was incubated at 37 °C for 96 h before harvesting by flooding the uniform bacterial lawn with 1X PBS buffer containing 10% glycine and scraping the colonies using a cell scraper. E. coli was incubated at 37 °C overnight before transferring an isolated colony to 15 mL tryptic soy broth (TSB) (Sigma Aldrich) and incubating at 37 °C for 24 h.
2.2. Configuration, Preparation and Operation of the 2-D Tank
The tank used for this study was a 72 inch (182.88 cm) tall, 24 inch (60.96 cm) wide, 4 inch (10.16 cm) deep, rigid stainless steel frame that held 72 inch (182.88 cm) tall, 24 inch (60.96 cm) wide, 0.5 inch (1.27 cm) thick clear acrylic panels, front and back. The stainless-steel side panels and base of the tank were ported to allow flow control as desired. For this study, bilateral ports at 42 inches (106.7 cm) from the base of the tank were used for influent feed control and a French drain at the base of the tank provided uniform drainage to a single port for outlet flow control. Lateral bracing across the acrylic panels was used to prevent the panels from bowing under the pressure of a packed, saturated tank. The front acrylic face of the tank was fitted with two vertical series of sampling ports, 18 inches (45.72 cm) apart and centered across the tank. Vertical spacing of the ports in each column was on 2 inch (5.08 cm) centers (Figure 1) through the packed zone of the tank.
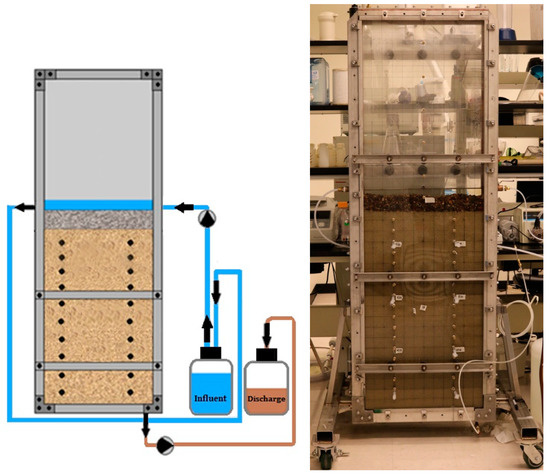
Figure 1.
A schematic (not to scale) and a photo of the 2-D tank packed with Sakrete© play sand media with sampling ports. Note: A 3 inch gravel pack is not visible at the base of the sand pack but is present to provide uniform effluent drainage across the tank.
The tank was packed with approximately 3750 in3 (61,451 cm3) of dry Sakrete© play sand (7-64661-15650-5). Sieve analysis for the product is shown in Table 1. The product presented as a fairly heterogeneous mix of filter media consisting of particles ranging from10 to 80 mesh (>2.0 mm to <0.18 mm). Porosity of the product was 31%. The total depth of sand pack was 35 inches (88.9 cm). Packing the tank involved the introduction and packing of sequential layers of dry sand to consolidate the sand pack and minimize settling during testing. Since play sand is not a uniform sieve grade, this packing technique resulted in a heterogeneous pack with some visually apparent, isolated areas of horizontal layering of fines and/or coarse materials. The character of the sand and the heterogeneous nature of the pack were considered advantageous as being more representative of natural conditions.

Table 1.
Sieve analysis for Sakrete© play sand.
The top of the sand pack was covered with a 3 inch (7.62 cm) gravel pack to preserve the integrity of the packed sand surface while allowing a uniform influent across the lateral profile of the tank. The base of the tank was packed with a 3 inch (7.62 cm) gravel pack to create a French drain to provide uniform drainage across the base of the tank to a single port for outlet flow control.
Prior to use, the tank was disinfected with a 10% (by volume) bleach solution followed by 4 flushes with dechlorinated tap water (containing 600 mg/L sodium thiosulfate). During each tank fill, water was delivered through the base of the tank at a rate of 3.6 mL/min to minimize air entrapment in the saturated media.
Tank flow was vertically downward for both experiments. Vertical transport during tank operation was managed via an influent pool at the packed media surface and a flow controlled outlet at the base of the tank. Tank flow was outlet controlled with a peristaltic pump operated at a flowrate of 3.5 mL/min, providing a fluid transport velocity for the tank of 10.3 in/day (26.2 cm/day) if undisturbed, uniform flow is assumed. However, the process of sampling created focused points of water withdrawal, creating disturbances in the uniform flow field at each sampling point and increasing the flow. While it was not possible to specifically define the flow field disturbance, the overall increase in flowrate during the period of tank operation can be defined and was 3.7 mL/min for a fluid transport velocity for the tank of 10.9 ft/day (27.7 cm/day) for the period of operation.
To maintain a uniform head and influent across the horizontal aspect of the tank, a 1.5 inch (3.8 cm) pool of water was maintained above the gravel surface. To ensure the delivery of fresh, dechlorinated, microbially enriched media, a peristaltic pump was used to deliver 15 mL/min from a carboy containing the influent water spiked with 105 colony forming units (CFU) per mL of Legionella and E. coli, the suspension of which was maintained by continuous stirring on a magnetic stir plate. Excess influent was allowed to passively drain from the tank at the same vertical level as the influent.
2.3. Sample Collection, Sample Processing and Assay Methods
Along with the influent (from the carboy), tank sampling was performed at ports 5 inches (12.7 cm), 15 inches (38.1 cm) and 25 inches (6.35 cm) below the sand surface along both the right (R) and left (L) columns of sampling ports to determine the mobility of the bacteria in-line with flow and to determine migration/transport times. Since the tank had both a left and right sampling array, sample locations were identified by their depth beneath the sand surface and whether that port was along the R or L sampling array (Figure 1). Hence, a sample taken from the right array at the 15 inch depth was denoted as 15R and from the left array as 15 L.
Preliminary data regarding tank flow velocity, collected during basic tank operation, were used to estimate expected breakthrough times for each sampling depth to guide sample collection. Sampling for each interval was on an hourly basis until breakthrough was achieved at both ports. Samples were collected by placing an 18G needle directly into the tank media through the port septa. Prior to sample collection, 5–10 mL of water was flushed, followed by the collection of a 10 mL sample. For this study, the first detection of target bacteria at a sampling port was considered the breakthrough point.
2.3.1. Legionella Analysis by Spread Plate Method
Samples for Legionella were analyzed by spread plate method using the BCYE media containing antibiotics Polymyxin B (100 units/mL), Vancomycin (5 µg/mL), Cycloheximide (80 µg/mL) and L-cysteine HCl (0.4 g/L). The samples were collected at specified time intervals, and 0.1 mL from each sample was transferred onto a petri dish and uniformly spread throughout. The petri dish was incubated at 37 °C for 96 h. Legionella on the BCYE media appeared as gray-white colonies.
2.3.2. E. coli Analysis by Spread Plate Technique
E. coli samples were analyzed using the spread plate method on selective media (Brilliance media, Oxoid CM1046 or Brilliant media, Sigma Aldrich 27815). Each sample was collected in a 15 mL tube and 0.1 mL was transferred onto a petri dish containing selective agar. A flame sterilized spreader (first dipped in Ethanol and flamed) was used to evenly spread the inoculum throughout the petri dish, which was then incubated at 37 °C for 24 h. The E. coli colonies appeared pink on the Brilliant media.
3. Results and Discussion
3.1. E. coli Transport Experimental Results
E. coli transport was studied by collecting hourly samples at the 5 inch ports between 4 and 9 h, the 15 inch ports between 16 and 20 h, and the 25 inch ports between 28 and 32 h. In order to accurately capture breakthrough, sampling began 1 h prior to the expected breakthrough time, and in all cases, those samples were negative for the target cells. The accuracy of the reported breakthrough times was within one hour. All experiments were repeated 5 times and variations in breakthrough concentrations and times at the respective ports were observed. Breakthrough transport times for E. coli are shown in Table 2. Based on the intervals 0 to 5 inches, 0 to 15 inches, and 0 to 25 inches, the number of pore volumes for breakthrough at 5, 15, and 25 inches was 0.46, 0.52, and 0.53, respectively.
Table 2.
Breakthrough times for E. coli.
Breakthrough concentrations for E. coli were also tracked at all of the ports, with sampling for several hours after initial breakthrough. Concentration vs. time is shown in Figure 2. Note that concentration vs. time showed increasing concentration with time after breakthrough followed by a decline. This behavior is consistent with a previous study in saturated porous media, which showed E. coli concentrations initially increasing after breakthrough followed by a decline [18].
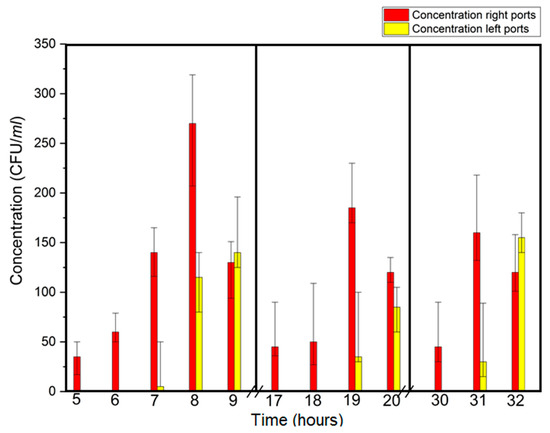
Figure 2.
Breakthrough time (±1 h) and concentration of E. coli as a function of time in three time periods.
3.2. Legionella Transport Experimental Results
Following E. coli tests, Legionella transport was studied by collecting hourly samples at the 5, 15, and 25 inch ports between 5 and 11 h, 17 and 26 h, and 28 and 38 h, respectively. As with E. coli testing, Legionella sampling began 1 h prior to the expected breakthrough time, and in all cases, those samples were negative for target cells. Again, the accuracy of the reported breakthrough times was within one hour and all experiments were repeated 5 times and variations in breakthrough concentrations and times at the respective ports were observed. Breakthrough transport times for Legionella are shown in Table 3. Based on the intervals 0 to 5 inches, 0 to 15 inches, and 0 to 25 inches, the number of pore volumes for breakthrough at 5, 15, and 25 inches was 0.54, 0.54, and 0.53, respectively.
Table 3.
Breakthrough times for Legionella.
Breakthrough concentrations vs. time for E. coli and Legionella are shown in Figure 2 and Figure 3, respectively. These graphics indicate that, for each time period tested Legionella concentrations continuously increased with time, unlike the decrease observed for E. coli. This might be due to the difference in the behavior of E. coli and Legionella transport in the plume. In plume form, bacteria move in high concentrations, which allows greater cell-to-cell interaction. Bacterial clumping is a well known phenomenon which can occur in culture or in plume environments. Some of the factors that can influence bacterial clumping are: a) number of autoagglutinins (agglutinating proteins) on the cell surface and the presence divalent cation such as Ca2+. It is known that E. coli cell can have up to eight different autoagglutinins compared to only one type on Legionella cells [19]. The difference in the number of surface-active agglutinating proteins might account for the difference in retention because of cations in the filter media.
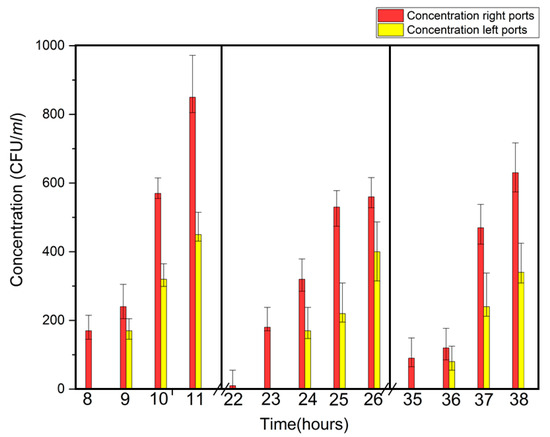
Figure 3.
Breakthrough time (±1 h) and concentration of Legionella as a function of time in three time periods.
3.3. Discussion of Transport Characteristics
3.3.1. Tank Characteristics
As indicated previously, the tank was packed with play sand and the heterogeneities associated with the pack were visually apparent as isolated areas of horizontal layering of fines or coarser sand. While no dye or chemical tracer tests were performed to determine overall flow characteristics, previous tank experience would indicate that while such packing idiosyncrasies introduce some irregularity to flow profile, no significant vertical flow heterogeneities would exist laterally across the tank. Via calculations based on the controlled flowrate and volume and porosity of the sand pack, the overall tank velocity assuming uniform flow across the tank was 10.3 in/day (26.2 cm/day), but sampling created an increase in tank velocity to 10.9 ft/day (27.7 cm/day) for the period of operation. Using the increased flowrate, fluid travel times from the top of the sand pack to the 5, 15, and 25 inch sampling ports were 11, 33, and 55 h, respectively, with an overall travel velocity of 0.45 in/h (1.14 cm/h; see Table 4). Travel time between the 5 and 15 inch (or 15 and 25 inch) ports was 22 h.
Table 4.
Travel times for fluid vs. E. coli vs. Legionella.
Of additional note regarding tank characteristics is the apparent lag in the microbial transport rate for left array versus right array ports. This is most likely related to the tank sand pack or sample timing issue. The lag of up to 2 h from 0 to 5 inches for E. coli was not much different than the cumulative lag of 2 h from 0 to 15 inches for Legionella (±1 h for both), and might simply be an artifact associated with sample timing. This is compelling when one considers the overall differential in travel time from 0 to 25 inches of only one hour. However, since the lag is consistently noted on the left array, there could be some issue associated with left side tank flow. Given that the transport lag predominates in the top 5 inches of travel, the most plausible explanations include the following:
- The surface of the sand pack was irregular and was only 4 inches from the 5 inch port on the right side. That one inch difference would yield a 2.3 h differential in transit time.
- The influent pool was filled from the right side of the tank with outflow on the left, with a residence time was 253 min. This ponding could have resulted in preferential deposition of microbes on the right side of the tank, ultimately affecting transit times to the 5 inch port.
In either case, this idiosyncrasy is irrelevant towards the overall purpose of this study, which was to investigate microbial transport characteristics.
3.3.2. Microbial Transport Characteristics
In vertically downward, saturated flow conditions in sand media, both E. coli and Legionella showed significant mobility and were readily transported across the 25 in (63.5 cm) zone of observation. Most notably, both E. coli and Legionella showed transport rates that were greater than the apparent fluid velocity (see Table 4). While preferential flow and/or mobility of the species could be argued, the consistency of travel times between the right and left arrays and from one port to the next reduces the likelihood of this explanation. In any case, both E. coli and Legionella showed significant mobility in simulated aquifer conditions, which is a major point of significance for this study.
Legionella transport characteristics differed from those of E. coli, with Legionella exhibiting retarded breakthrough times for the 0 to 5 and 5 to 15 inch zones. While the E. coli rate of transport was relatively consistent from 0 to 25 inches at 0.80 to 0.83 in/h (2.03 to 2.11 cm/h), Legionella showed increasing rates of transport at 0.59 in/h (1.50 cm/h) from 0 to 5 inches, 0.69 in/h (1.75 cm/h) from 5 to 15 inches, and 0.80 in/h (2.03 cm/h) from 15 to 25 inches. Several factors can influence the transport of bacteria through saturated porous media. Attachment of bacterial cells to media surfaces is influenced by cell surface electrostatic charge and hydrophobic interaction with the media, size, and the presence of surface structures such as flagella, fimbriae, and extracellular lipopolysaccharides [20]. Each of these parameters adds complexity to building a deeper understanding of bacterial transport through the subsurface, such that microbial subsurface transport is not fully understood [21]. For example, electrostatic charges are influenced by not only the microorganisms’ surface (e.g., the presence and configuration of proteins, and lipopolysaccharides), but also the granular media characteristics and the water matrix itself. Specifically, the pH and the ionic strength of the solution affect the surface charge of the bacterial cell and soil particle, thereby dictating electrostatic interactions. Direct assessment of surface charges was beyond the scope of this study. However, reported that the electrophoretic mobility of some Legionella pneumophila serogroups varies with solution pH while other serogroups remain constant between pH 6 and 9 [22]. These pH values encompass common water ranges, including the water tested in this study).
Different types of lipopolysaccharides (LPSs) are located on the outside of bacterial cells and they are considered a key factor in cell attachment to mineral surfaces and microbially induced precipitation/dissolution reactions. The E. coli LPSs are composed of three different components: (i) hydrophobic lipid A anchored in the outer membrane, (ii) phosphorylated, nonrepetitive hetero-oligosaccharide known as the core oligosaccharide, and (iii) polysaccharide that extends from the cell surface and forms the O antigen detected in serotyping [23]. Alternately, LPSs on Legionella pneumophila cells are composed of a very hydrophobic lipid A acylated by long chain fatty acids and an O-antigen-specific chain consisting of homopolymeric legionaminic acid [24]. The variation in transport profiles between E. coli and Legionella might be due to the significant differences in the LPSs on cell surfaces resulting in reversible or irreversible attachment of the Legionella cells to the tank media. There has not been any study investigating a direct link between bacterial cell transport in porous media and the different types of lipopolysaccharides on bacterial cell membranes.
In addition, filamentation in response to different environmental stresses has been observed for numerous bacterial species [25,26,27]. Under stress and nutrient deficient conditions, Legionella tends to become long and filamentous [28]. The size and shape of the bacteria are known to be determining factors in their transport under saturated conditions [29,30]. Therefore, it is hypothesized that the pleomorphic nature of Legionella might be a factor in its slower transport characteristics as compared to E. coli, which does not exhibit pleomorphic characteristics.
Regarding Legionella’s apparent increase in transport for all zones, this could simply be a function of filtration or size selection, with preferential transport rates favoring those microbes with smaller profiles or less filamentous development.
4. Conclusions
While it is accepted that E. coli is mobile in saturated aquifer environments, the scale of this study provided a basis for the premise that Legionella is also mobile in saturated aquifer conditions. In addition, data from this study suggested that microbial cell types and characteristics in conjunction with aquifer characteristics might have impacted the transport of those pathogens. Legionella’s pleomorphic nature and/or the differences in the LPSs on cell surfaces, which may result in reversible or irreversible attachment of the Legionella cells to the media of the tank, may both have affected its transport properties. Further, vadose zone conditions are typically heterogeneous, ranging from the micro to the macro scale, which can lead to preferential transport affecting dispersion in both the lateral and vertical directions.
Future studies should investigate the extent of Legionella contamination in groundwater impacted directly or indirectly by aquifer recharge practices and Legionella’s transport characteristic in the vadose zone. An assessment of the overall data indicated parallel trends in transport of E. coli and Legionella under the experimental conditions studied. Considering the delay in Legionella transport, historical data for E. coli may serve as a useful, reasonable prediction of Legionella’s transport under recharge conditions.
Author Contributions
I.M., J.A.: investigation; A.A., B.K.M., P.D.: formal analysis; M.A.: project administration. All authors contributed equally at all the stages of this study. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Science Foundation (NSF) Water and Environmental Technology (WET) Center at Arizona State University (award number 1361815).
Acknowledgments
All opinions expressed in this paper are the authors’ and do not necessarily reflect the policies and views of NSF.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shen, Y.; Oki, T. Groundwater Resources in Urban Water Management. In Groundwater Problems in Urban Areas; Takizawa, S., Ed.; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Hynds, P.D.; Thomas, M.K.; Pintar, K.D.M. Contamination of groundwater systems in the US and Canada by enteric pathogens, 1990–2013: A review and pooled-analysis. PLoS ONE 2014, 9, e93301. [Google Scholar] [CrossRef] [PubMed]
- Slutsker, L.; Ries, A.A.; Maloney, K.; Wells, J.G.; Greene, K.D.; Griffin, P.M. A nationwide case-control study of Escherichia coli O157:H7 infection in the United States. J. Infect. Dis. 1998, 177, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.S.; Klohe, C.A.; Battigelli, D.A. Occurrence and Distribution of Enteric Viruses in Shallow Ground Water and Factors Affecting Well Vulnerability to Microbiological Contamination in Worcester and Wicomico Counties, Maryland; USGS and Maryland Department of the Environment: Baltimore, MD, USA, 2001.
- Abbaszadegan, M.; LeChevalier, M.; Gerba, C.P. Occurrence of viruses in US Groundwaters. JAWWA 2003, 95, 107–120. [Google Scholar] [CrossRef]
- Embrey, S.S.; Runkle, D.L. Microbial Quality of the Nation’s Ground-Water Resources, 1993–2004: U.S. Geological Survey Scientific-Investigations Report 2006-5290; USGS: Reston, VA, USA, 2006; 34p.
- Fout, G.S.; Borchardt, M.A.; Kieke, B.A.; Karim, M.R. Human virus and microbial indicator occurrence in public-supply groundwater systems: Meta-analysis of 12 international studies. Hydrogeol. J. 2017, 25, 903–919. [Google Scholar] [CrossRef] [PubMed]
- McBurnett, L.R.; Holt, N.T.; Alum, A.; Abbaszadegan, M. Legionella- A threat to groundwater: Pathogen transport in recharge basin. Sci. Total Environ. 2018, 621, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Arora, B.; Spycher, N.F.; Steefel, C.I.; Molins, S.; Bill, M.; Conrad, M.E.; Dong, W.; Faybishenko, B.; Tokunaga, T.K.; Wan, J.; et al. Influence of hydrological, biogeochemical and temperature transients on subsurface carbon fluxes in a flood plain environment. Biogeochemistry 2016, 127, 367–396. [Google Scholar] [CrossRef]
- Frind, E.O.; Germain, D. Simulation of contaminant plumes with large dispersive contrast: Evaluation of alternating direction Galerkin models. Water Resour. Res. 1986, 22, 1857–1873. [Google Scholar] [CrossRef]
- Erickson, A.J.; Weiss, P.T.; Gulliver, J.S. Optimizing Stormwater Treatment Practices, A Handbook of Assessment and Maintenance; Springer: New York, NY, USA, 2013. [Google Scholar]
- Brunkard, J.M.; Ailes, E.; Roberts, V.A.; Hill, V.; Hilborn, E.D.; Craun, G.F.; Rajasingham, A.; Kahler, A.; Garrison, L.; Hicks, L.; et al. Surveillance for waterborne diseases outbreaks associated with drinking water—United States, 2007–2008. CDC Surveill. Summ. 2011, 60, 38–68. [Google Scholar]
- Jjemba, P.K.; Weinrich, L.A.; Cheng, W.; Giraldo, E.; LeChevallier, M.W. Regrowth of potential opportunistic pathogens and algae in reclaimed-water distribution systems. Appl. Environ. Microbiol. 2010, 76, 4169–4178. [Google Scholar] [CrossRef]
- Palmer, C.J.; Bonilla, G.F.; Roll, B.; Paszko-Kolva, C.; Sangermano, L.R.; Fujioka, R.S. Detection of Legionella species in reclaimed water and air with the EnviroAmp Legionella PCR kit and direct fluorescent antibody staining. Appl. Environ. Microbiol. 1995, 61, 407–412. [Google Scholar] [CrossRef]
- Palmer, C.J.; Tsai, Y.L.; Bonilla, G.F.; Sangermano, L.R.; Paszko-Kolva, C.; Roll, B.; Fujioka, R.S. Detection of Legionella species in sewage and ocean water in California and Hawaii. In Proceedings of the Annual Conference of Water and Environment Federations, Water and Environment Federations, Alexandria, VA, USA, 2–7 October 1993; pp. 123–131. [Google Scholar]
- Palmer, C.J.; Tsai, Y.; Paszko-Kolva, C.; Mayer, C.; Sangermano, L. Detection of Legionella species in sewage and ocean water by polymerase chain reaction, direct fluorescent-antibody, and plate culture methods. Appl. Environ. Microbiol. 1993, 59, 3618–3624. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.J.; Steigerwalt, A.G.; McDade, J.E. Classification of the Legionnaires’ disease bacterium: Legionella pneumophila, genus novum, species nova, of the family Legionellaceae, familia nova. Ann. Intern. Med. 1979, 90, 656–658. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Bradford, S.A.; Torkzaban, S.; Ye, X.; Vanderzalm, J.; Du, X.; Page, D. Unraveling the complexities of the velocity dependency of E. coli retention and release parameters in saturated porous media. Sci. Total Environ. 2017, 603–604, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Gilbert, P.; Evans, D.J.; Evans, E.; Duguid, I.G.; Brown, M.R.W. Surface characteristics and adhesion of Escherichia coli and Staphylococcus epidermis. J. Appl. Bacteriol. 1991, 71, 72–77. [Google Scholar]
- Dowd, S.E.; Pillai, S.D.; Wang, S.Y.; Corapcioglu, M.Y. Delineating the specific influence of virus isoelectric point and size on virus adsorption and transport through sandy soils. Appl. Environ. Microbiol. 1998, 64, 405–410. [Google Scholar] [CrossRef]
- Buse, H.Y.; Hoelle, J.M.; Muhlen, C.; Lytle, D.A. Electrophoretic mobility of Legionella seropgrousp 1 to 4. FEMS Microbiol. Lett. 2018, 365, 1–7. [Google Scholar] [CrossRef]
- Amor, K.; Heinrichs, D.E.; Frirdich, E.; Ziebell, K.; Johnson, R.P.; Whitfield, C. Distribution of core oligosaccharide types in lipopolysaccharides from Escherichia coli. Infect. Immun. 2000, 6, 1116–1124. [Google Scholar] [CrossRef]
- Lück, C.; Helbig, J.H. Characterization of Legionella Lipopolysaccharide. In Legionella. Methods in Molecular Biology (Methods and Protocols); Buchrieser, C., Hilbi, H., Eds.; Humana Press: Totowa, NJ, USA, 2013; Volume 954. [Google Scholar]
- Michel, B. After 30 years of study, the bacterial SOS response still surprises us. PLoS Biol. 2005, 3, e255. [Google Scholar] [CrossRef]
- Miller, C.; Thomsen, L.E.; Gaggero, C.; Mosseri, R.; Ingmer, H.; Cohen, S.N. SOS response induction by beta-lactams and bacterial defense against antibiotic lethality. Science 2004, 305, 1629–1631. [Google Scholar] [CrossRef]
- Radman, M. SOS repair hypothesis: Phenomenology of an inducible DNA repair which is accompanied by mutagenesis. Basic Life Sci. 1975, 5, 355–367. [Google Scholar]
- Greub, G.; Raoult, D. Morphology of Legionella pneumophila according to their location within Hartmanella vermiformis. Res. Microbiol. 2003, 154, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Johnson, W.P.; Shaifeian, P.; Ryu, H.; Alum, A.; Abbaszadegan, M.; Hubbs, S.A.; Williams, T.R. Riverbank Filtration: Comparison of Pilot Scale Transport with Theory. Environ. Sci. Technol. 2009, 43, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Firouzi, A.F.; Homaee, M.; Klumpp, E.; Kasteel, R.; Tappe, W. Bacteria transport and retention in intact calcareous soil columns under saturated flow conditions. J. Hydrol. Hydromech. 2015, 63, 102–109. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).