1. Introduction
The extended use of biofuels is widely promoted to decrease the carbon (C) emissions from fossil fuels, i.e., to fulfil the requirements of the EU directive (2009/28/EC) on renewable energies and to achieve zero net emissions of greenhouse gases in Sweden by the year 2045 [
1,
2]. In 2017, biofuels alone contributed 25% [
3] of Sweden’s total energy supply. Most of the energy supply from biofuels are based on ‘classical’ forest products (pulp industry fuels, wood fuel and sawmill by-products) but logging residues and tree stumps have also been used [
4]. However, the contribution to energy production based on agroforestry (i.e., energy crops) is expected to increase to meet the requirements of the EU directive. Besides ‘classical’ biofuel crops like rapeseed, sugar beets or oil seeds, fast-growing tree species (willow (
Salix spp.), poplar and hybrid aspen) are increasingly used as energy crops [
5], either for direct combustion or for the production of liquid fuels by ‘second generation’ bioethanol from lignocellulose. Energy crops are currently using 3% of arable land in Sweden [
5].
Salix trees have been reported to grow on 12,000 ha in Sweden in 2014 [
6] but the potential use is estimated at 300,000 ha [
5]. The advantages of willow are the high energy content (more than twice that of oat), the ability to clean up from soils waste water treatment products and cadmium, and the greater increase in the C stock in soil and mulch compared to with annual crops, as willow is grown for 4 years before cutting [
7,
8].
Willow has been used intensively as energy crop since the 1990s, and varieties have been propagated to increase both biomass production and resistance against weeds and pests [
9]. Depending on the climate conditions, different varieties are suitable. For instance, at higher latitudes, such as in the middle and northern parts of Sweden, varieties need to be more resistant to frost, whereas at southern latitudes, trees can suffer from heat damage. There exist no official data on the distribution of the varieties, but studies have shown that Tora has been successfully grown in Sweden, as it gives 40–50% higher yield and has a better resistance to rust compared to older varieties such as L 78183, Orm and Rapp [
9]. Other varieties, e.g., Sven and Inger, are suitable to be grown in Sweden, and Inger is also suitable for soils with a low soil water capacity [
10]. As
Salix is easy to propagate, new varieties are continuously propagated by commercial companies that aim at increasing biomass yield and tolerance against insects, plant pests and weeds. Additionally, a change of growing conditions due to climate change might imply that older varieties should be replaced with newer ones, which have been specifically propagated to cope better with drought.
While
Salix plantations may be a good option for energy crop production, a large-scale land-use change towards
Salix might have severe impacts on atmospheric chemistry and local air quality. Areas used for short-rotation coppices (SRC) to increase the production of biofuels are most converted from traditional agricultural crops. In contrast to agricultural crops,
Salix species are regarded as high-emitters of biogenic volatile organic compounds (BVOCs) [
11] that are very reactive and can contribute to the production of ozone (O
3) and secondary organic aerosols (SOAs) [
12,
13,
14,
15,
16,
17].
Salix species have been shown to emit large amounts of isoprene, with standardized emission rates ranging from 12.5 to 115.0 µg g
dw−1 h
−1 [
11,
18,
19,
20,
21,
22]. Monoterpene (MT) emissions from some
Salix spp. have also been reported [
11,
18,
23], but data for the quantification of compounds other than isoprene and monoterpenes (MTs) are scarce [
22].
The large variation of published standardized emission rates for
Salix spp. indicates an influence of genetic disposition on the BVOC production and emission [
24], which has been observed for other species as well [
25,
26]. Consequently, commercial propagation methods to find better varieties that provide higher biomass yields, increased resistance against plant pests and enhanced competitiveness against weeds might also affect the production and emission of BVOCs.
Here, we analyze leaf-scale BVOC emissions from several varieties of willow that were growing either in field trails or commercially on SRC plantations. We aim to identify the compound spectrum emitted by these varieties, and provide standardized emission rates that can be used in emission inventories and distributed vegetation models to assess the impact of willow plantations on regional air quality.
4. Discussion
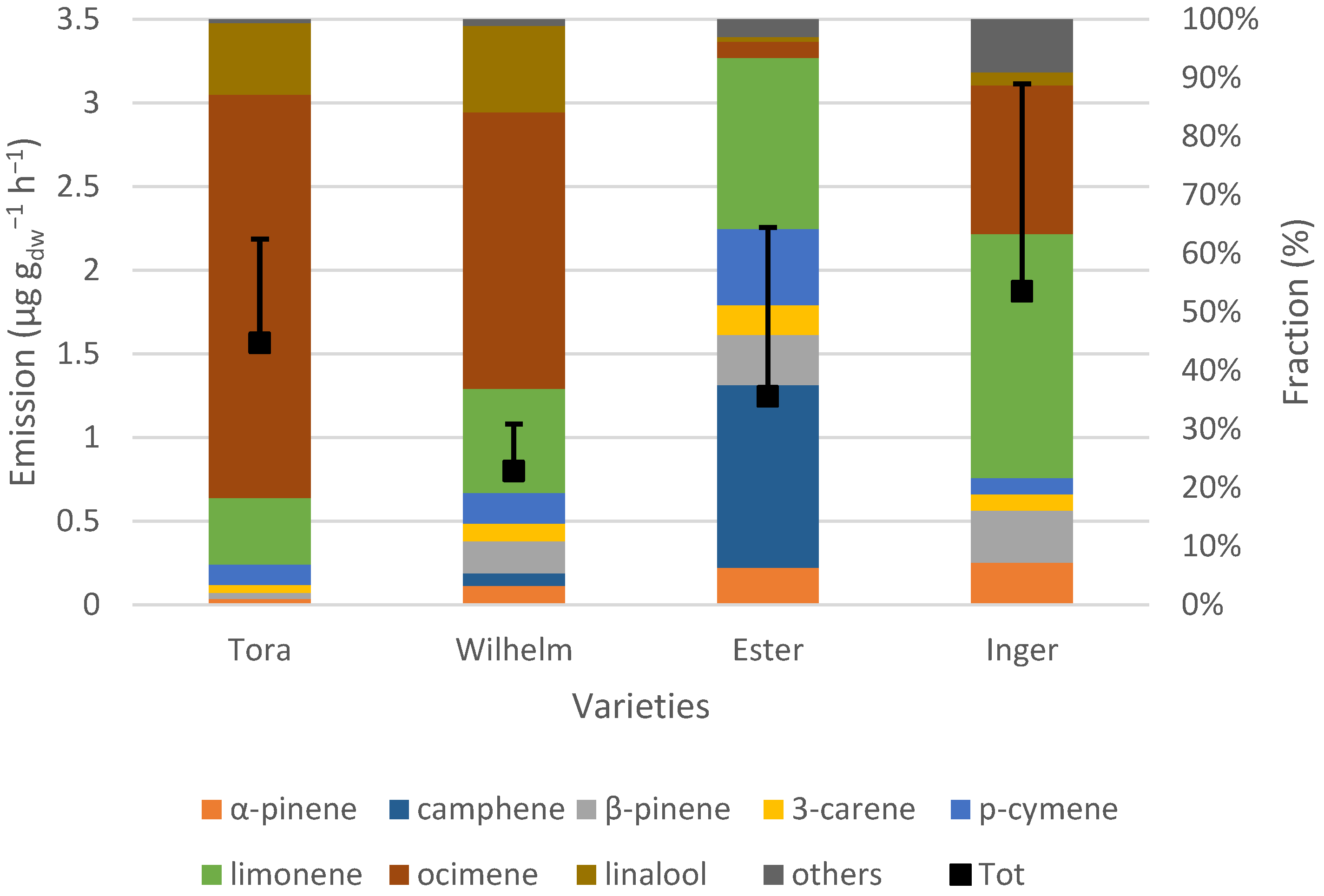
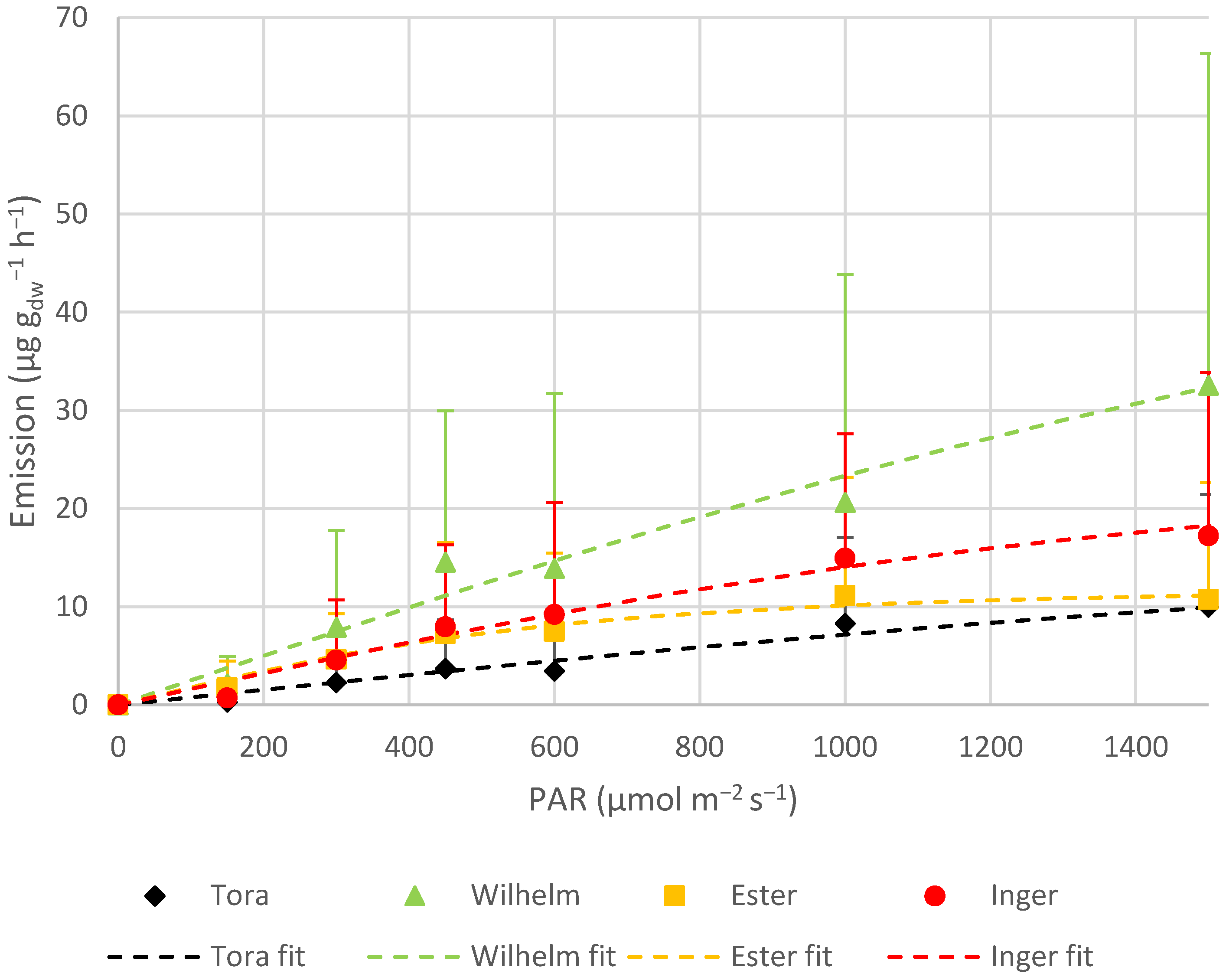
Wilhelm was the variety that emitted the highest rate of terpenoids. Most of this emission (circa 90%) came from isoprene. In fact, Wilhelm emitted over three times more isoprene than Tora and almost twice as much as Ester and Inger. However, when comparing MTs and SQTs, Wilhelm had the lowest emissions. The average emissions of isoprene and SQTs were almost the same for Ester and Inger. The pathways of producing BVOCs have been studied and disentangled to a certain extent. Even if it is not fully understood, studies have shown that there is some linkage between the productions of these compounds [
36,
37]. The originating substrates responsible for the end products (e.g., isoprene, MTs and SQTs) are shared and divided into the separate pathways, which could be one of the explanations why Wilhelm emits lower amounts of MTs and SQTs, but more isoprene. The average T and PAR values within the chambers were almost the same for the varieties, indicating that the different emission rates among the varieties are related to other differences in the environment, or genetic variation. Genetic diversity was concluded by van Meeningen et al. [
30] to be more important than, e.g., local growing conditions, when studying spruce BVOC emissions. Hence, for one specific species, the BVOC emission rates can differ among the varieties or clones. This difference is not always accounted for in models and should not be discarded when improving modelling for upscaling BVOC emissions.
The A was similar for Tora, Wilhelm and Inger (circa 13.5 µmol CO
2 m
−2 s
−1), reflecting that they are equally good at biomass production in the prevailing conditions in this study. Ester had circa 25% lower A, showing less productivity than the others. Despite the lower A, Ester showed a better ability to utilize water for producing biomass when photosynthesis occurred. The values of WUE related to Ester were up to twice as large compared to the others for some PAR values, which means that Ester lost less than 40% of the water. Therefore, Ester is more suitable for hot and dry climates and it outcompetes the other varieties in regions warmer and drier than southern Sweden. The maximum A for Salix trees has been reported to range from 20 to 30 µmol CO
2 m
−2 s
−1 [
38]. The varieties in this study had, in general, lower A, but they were able to assimilate more than 20 µmol CO
2 m
−2 s
−1 when PAR reached 1000 or 1500 µmol m
−2 s
−1.
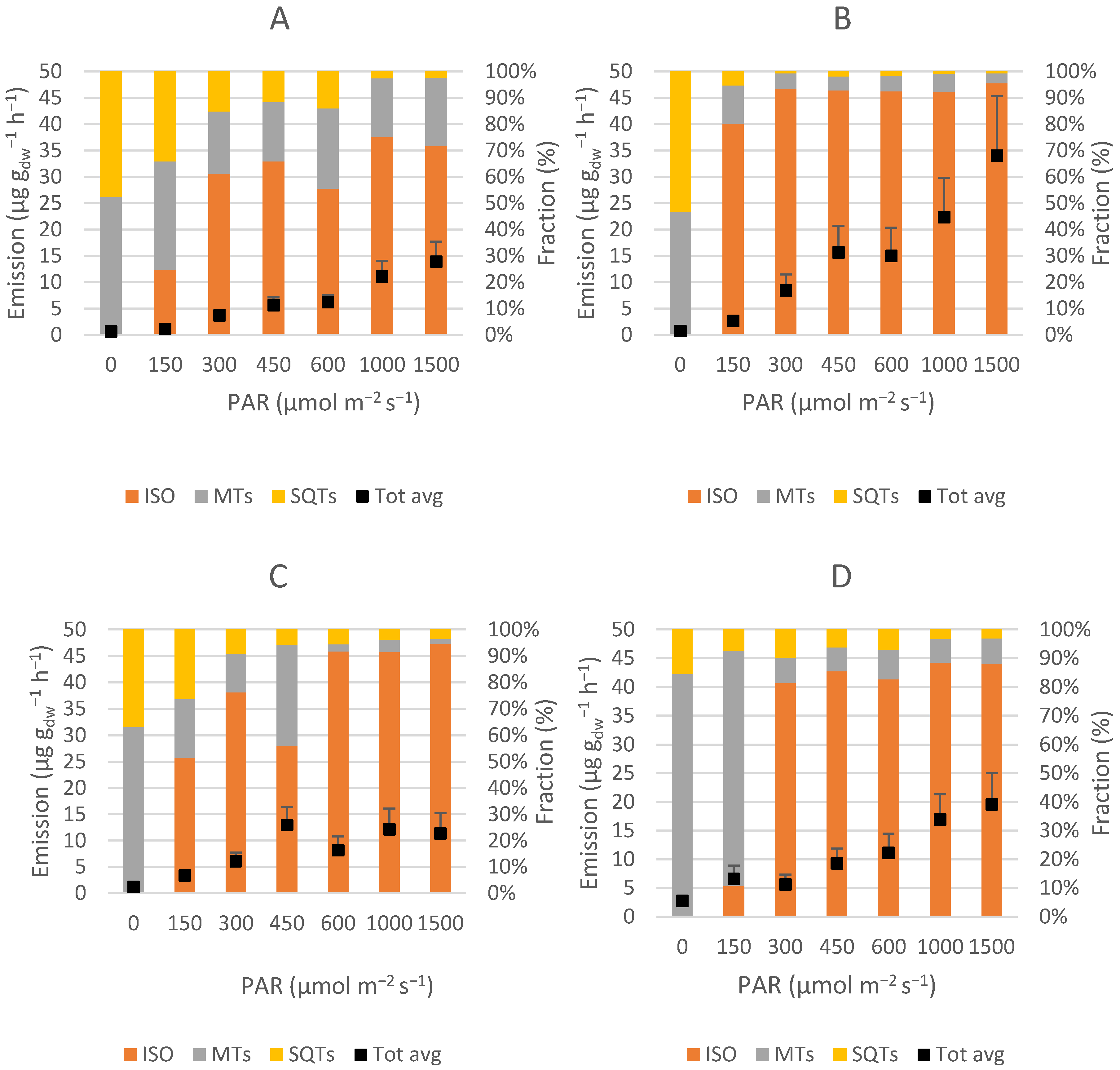
As expected [
31,
34,
39,
40,
41], isoprene increased with increasing PAR levels. Studies have also shown that the emission rates of isoprene have a hyperbolic relationship with PAR [
34,
40,
42,
43]. Tora, Ester and Inger showed a similar trend, where the emission rates levelled out for the higher light levels. Ester was the only variety that peaked at 1000 μmol m
−2 s
−1. Since no obvious damages could be seen on the leaves, this result indicates that the leaves belonging to Ester were saturated at lower light levels and could not utilize and respond to the highest PAR level like the other varieties. On the other hand, isoprene emission from Wilhelm continued to increase and showed no trend towards levelling out. Even though Wilhelm and Tora share similar ancestors from the breeding program, Tora is closer to Ester and Inger when it comes to isoprene emission. The photolysis of BVOCs and NO
x can lead to the production of O
3 and PAN [
44,
45], which are harmful for humans and vegetation at high concentrations [
46,
47,
48,
49]. Isoprene has been shown to be able to increase O
3 and PAN [
44,
50], which makes Wilhelm less preferable in high-NO
x environments compared to the other varieties. A major part of land cover in Sweden is boreal forest, whereof most is spruce (
Picea abies) and pine (
Pinus sylvestris). Isoprene emission from these species is much lower compared to that from
Salix [
11,
30]. In the Southern part of Sweden, the common land cover is farmland. Commercial crops growing on agricultural areas in Sweden, such as wheat, also emit significantly lower rates of isoprene [
11,
23]. Hence, a land cover change from the traditional species to
Salix plantations could alter the regional atmospheric chemistry leading to, e.g., increased levels of O
3. However, isoprene-emitting plants seem to tolerate O
3 better than other non-isoprene emitting plants, and in this sense, varieties such as Wilhelm may be more resistant if growing in areas with high prevailing O
3 concentrations [
51,
52,
53].
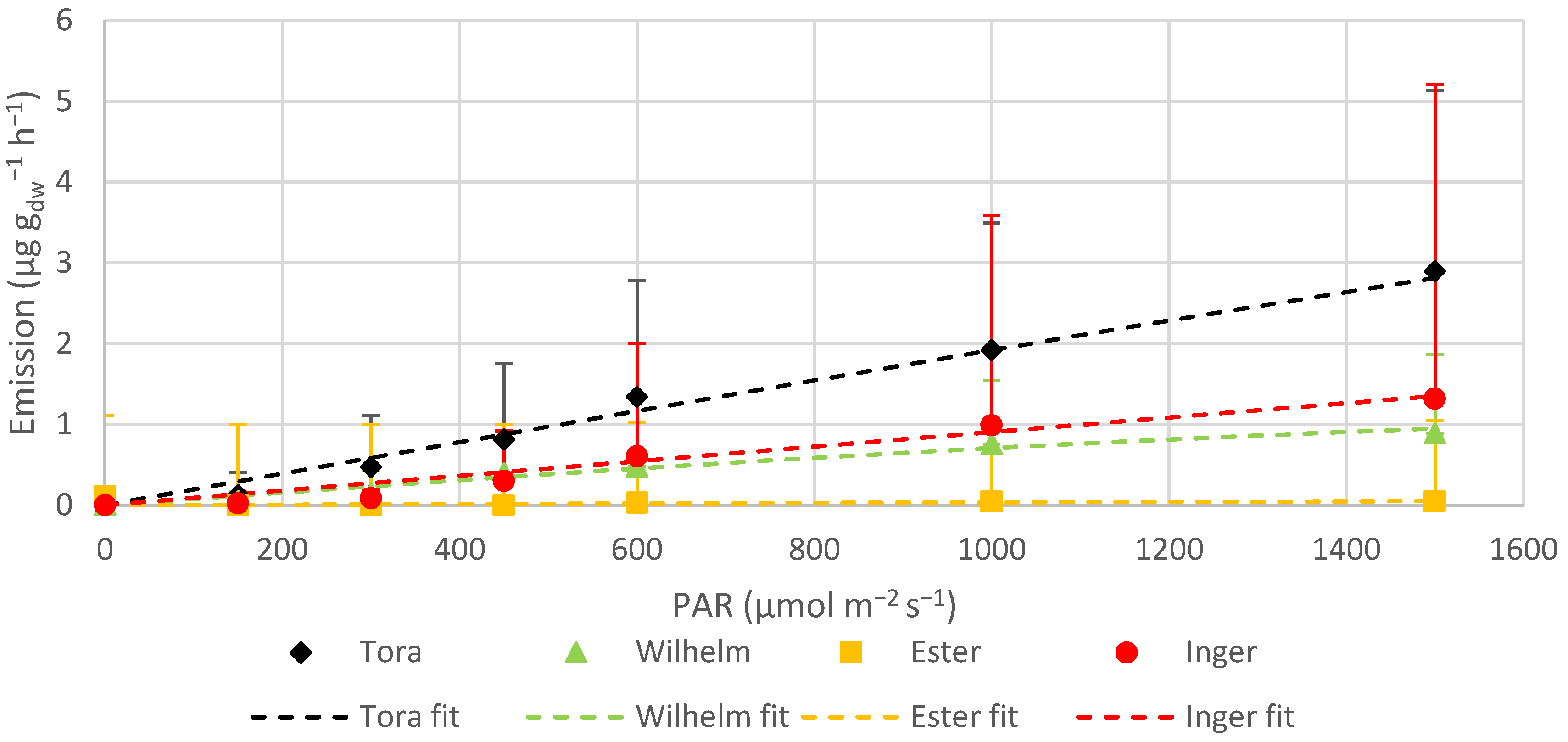
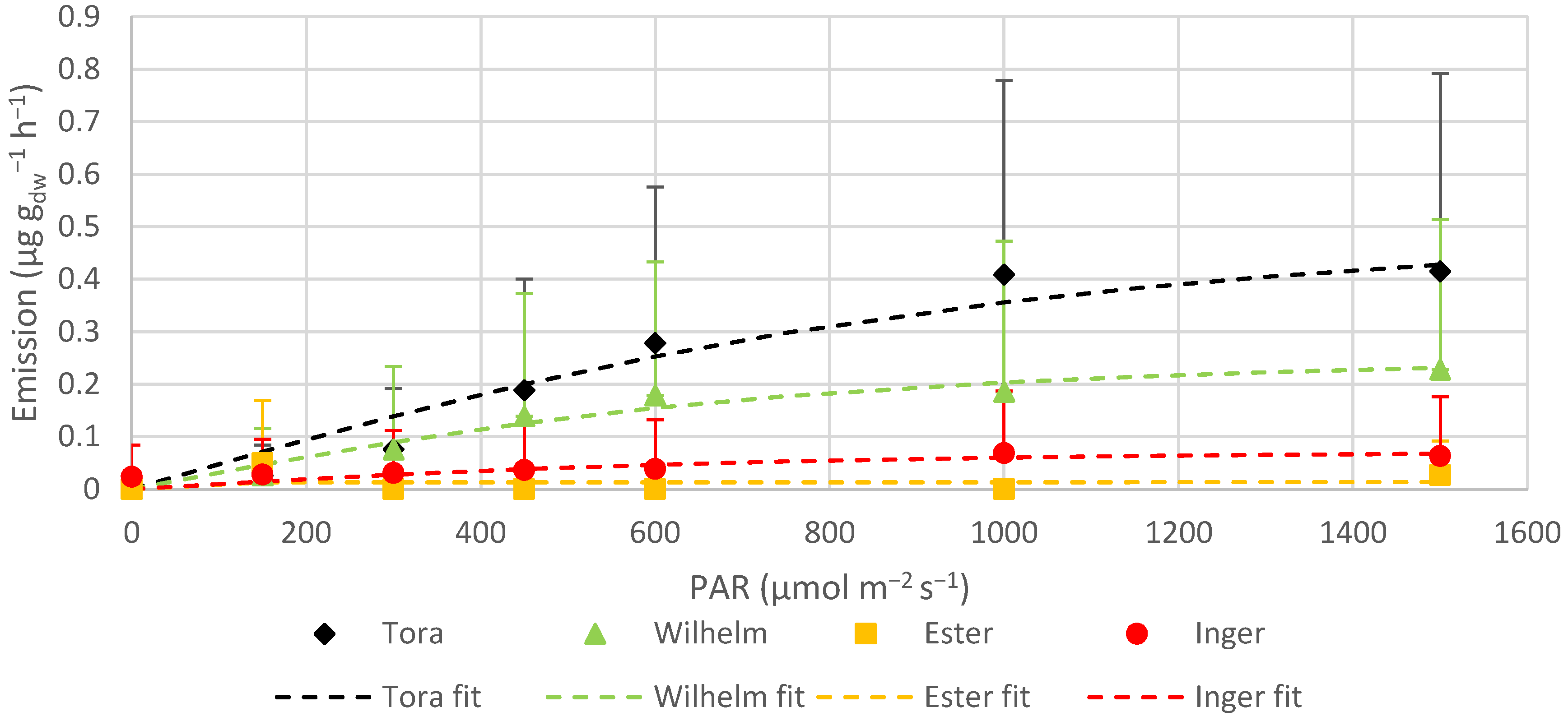
The monoterpene ocimene was emitted by all varieties, but at different rates. For Tora, Wilhelm and Inger, ocimene contributed circa 25–69% of the total MT emissions, whilst it was a minor compound for Ester. Ocimene and linalool were the only MTs which showed light dependency in Tora, Wilhelm and Inger, but not in Ester, likely due to being emitted only in very low amounts. In Tora and Inger, ocimene emission did not show any clear indication of leveling out, even at the highest measured PAR values. Wilhelm, on the other hand, did not increase the emission of ocimene much after 1000 μmol m
2 s
−1, and linalool seemed to level out for Tora and Wilhelm when PAR was above 1000 μmol m
2 s
−1. To date, no study has reported a light dependent relationship for MT emissions from willow trees, because the focus of most studies has been on isoprene. Monoterpenes can be important for generating secondary organic aerosols [
54,
55,
56,
57]. Since Ester was the only variety that did not increase MT emissions with increasing light, this variety might be more suitable near urban regions with more solar irradiance to avoid impaired air quality.
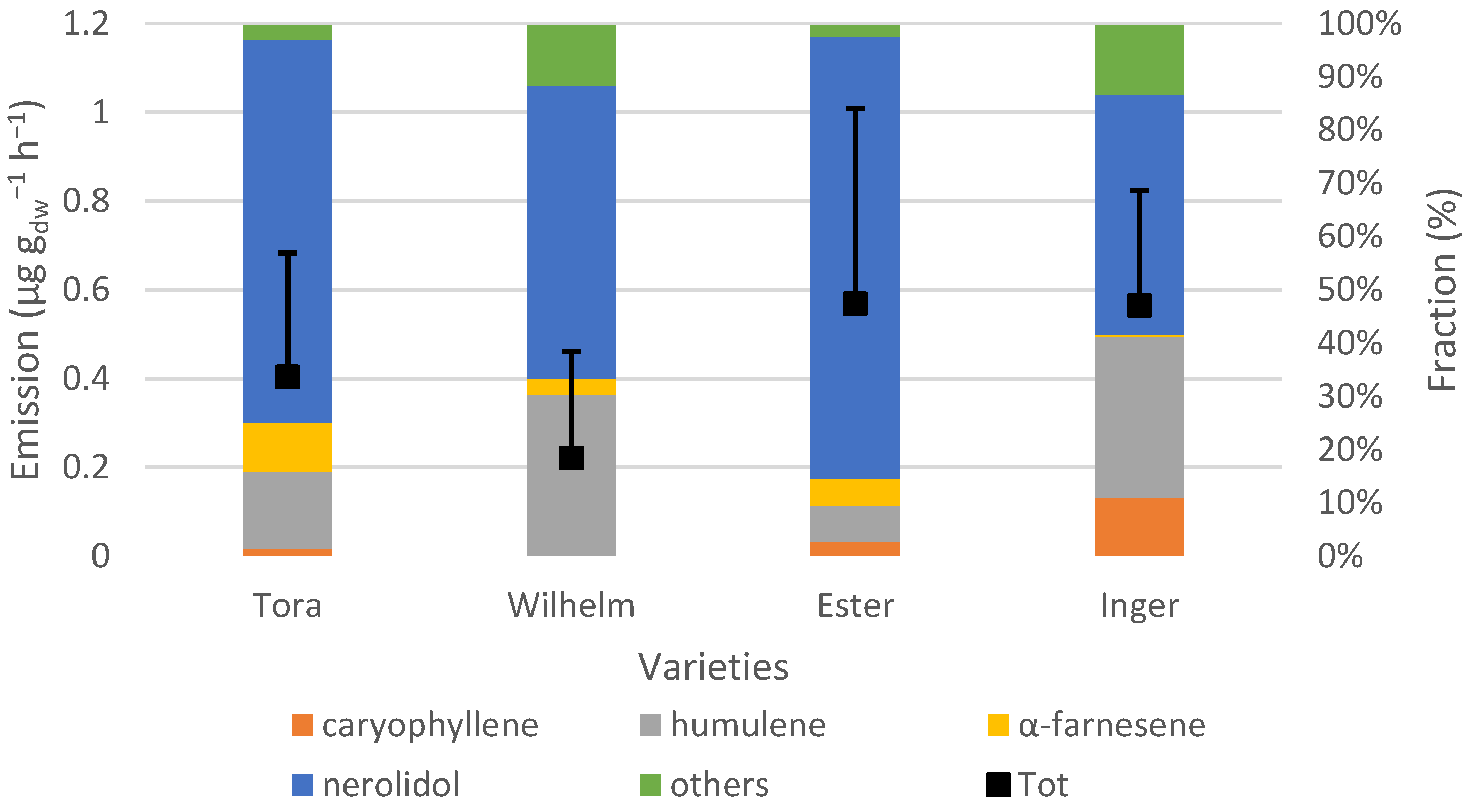
Nerolidol was the most dominant SQT and, together with humulene, constituted 75% or more of the total SQT emissions. Ester and Inger emitted approximately the same amounts of SQTs. Both of these varieties are female hybrids suitable for warm climates, and Ester also originates from Inger, which probably explains the similarities. However, the fractions of the emitted BVOCs differed. For example, no camphene was emitted by Inger, while camphene contributed almost one third of the MT emissions of Ester. In addition, Inger was the only variety that had a clearly increased emission rate of caryophyllene when light availability increased.
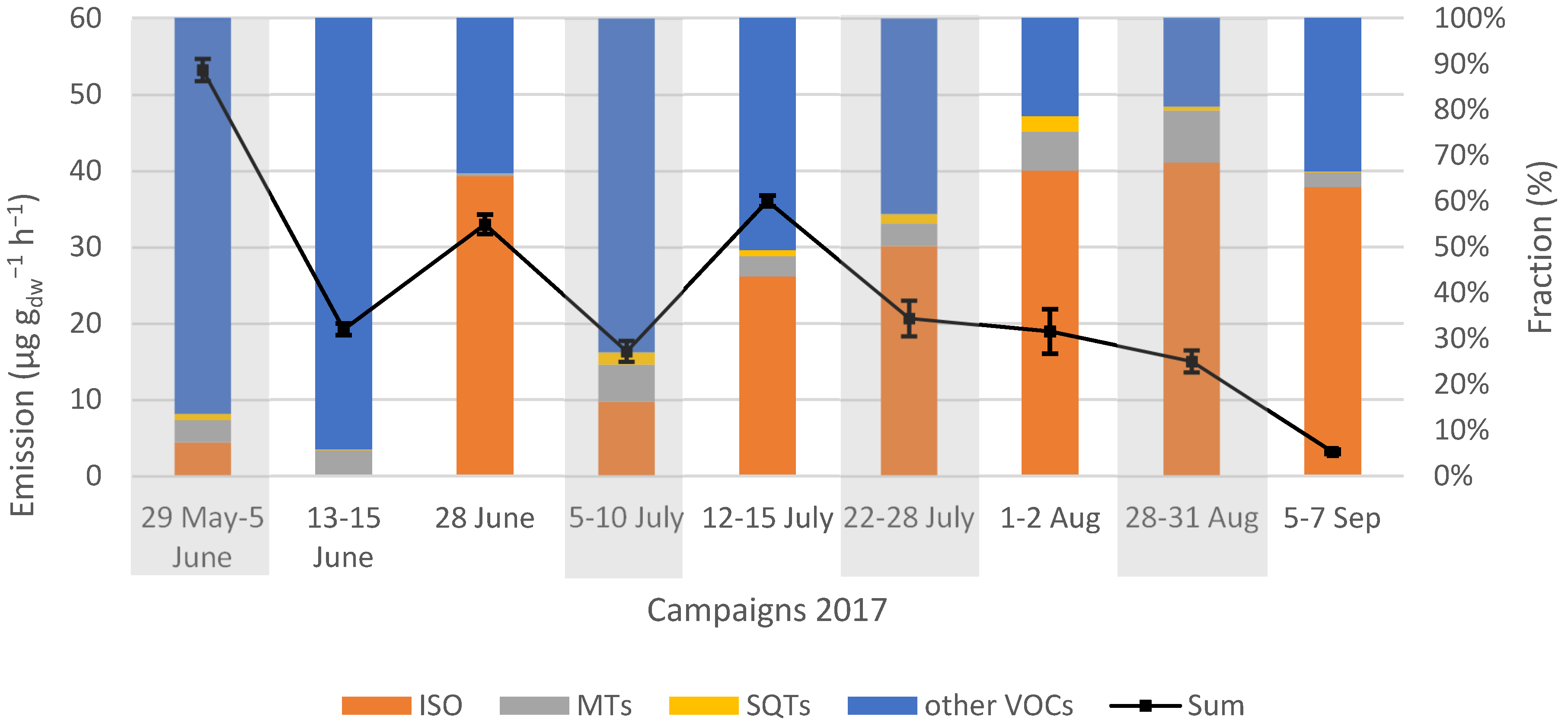
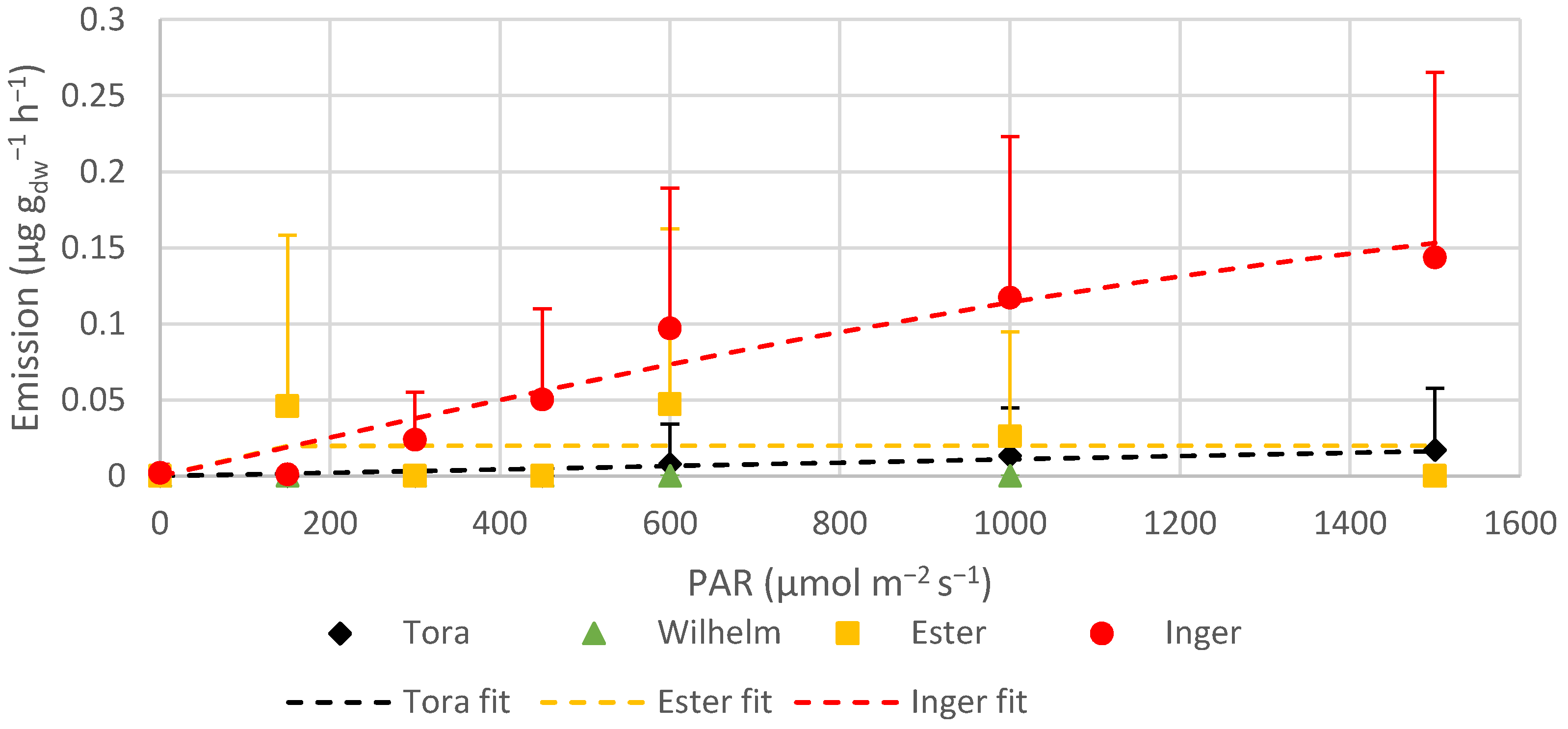
Saplings emitted approximately 3–19 times more other VOCs than the trees that were 1-year-old. Younger plants are more vulnerable than mature ones, and one way to strengthen their survival could be to emit more BVOCs [
58]. Tora, on plot 3, which had the same growing season as the saplings, emitted lower rates of other VOCs than the one year old Tora on plot 1, but higher than the saplings belonging to Tora on plot 2. The root system on plot 3 was established in 2003, which can be one reason why they differed in comparison to the saplings, since they had already a developed root system and trunk. The ratio between other VOCs and isoprene emission changed according to the aging of the trees. At the beginning of the season, the fraction of other VOCs exceeded the fraction of isoprene, but at the end of the season, the opposite was seen.
Compounds other than isoprene and MTs are rarely reported in studies on Salix trees, and only low emission rates have been observed for these compounds in the few studies that have [
22]. However, the results of this study show that they should not be discarded, at least not for saplings. Hexanal, which was the most emitted other VOC, has been reported as an important compound in abiotic and biotic stress [
59,
60,
61]. Irrespectively, the reason why leaf beetles attacked all of the other varieties but not Ester is unclear. No unique compound emitted by Ester was found. Benzaldehyde and xylenes have been reported as stress-induced compounds in trees [
62]. Even though the emissions of compounds such as benzaldehyde, furfural, p-cymene, camphene and nerolidol were higher from Ester compared to from the other varieties, the major contributions to these emission rates were observed from the saplings belonging to Ester and not from the 1-year-old trees. Therefore, one suggestion why the insects avoid Ester could be that this variety has compounds or other substances stored within their leaves that are not emitted unless the surface layer is broken, making Ester less attractive for leaf beetles.
The average standardized isoprene emission for the whole season (33.21 ± 53.43 µg g
dw−1 h
−1) is in line with other studies that have measured emissions from Salix trees [
11,
23]. It is hard to make a straightforward comparison since the methods, soil, adaptation to local growing conditions, age and different clones are likely to affect the emissions, and all these pieces of information are seldom presented in studies. Wild growing Salix species will also probably have different emission rates compared to commercial managed species. According to Morrison et al. [
23], standardized isoprene emission from Salix trees can be more than 100 µg g
dw−1 h
−1 but many emission rates range from 20 to 50 µg g
dw−1 h
−1.
The standardized average MT emission rate was 4.40 ± 2.05 µg g
dw−1 h
−1. The time of the year has been shown to influence the emission rate, and other studies have reported that Salix trees are prone to emit higher concentrations of MTs when they recently have had their bud break [
23,
63]. The study done by Ghelardini et al. [
64] showed that the day of bud burst for Salix can vary between seasons, and differ for different varieties [
65]. For the trees studied in Ghelardini et al. [
64], it took up to 260 degree days of T > 0 °C since the first of March to have a bud burst. This value was reached by the middle of April for plots 1 and 2, and by the end of April for plots 3 and 4, when counting degree days in the same way as in their study. The first campaign in this study was started by the end of May for plots 1 and 2, and in the middle of June for plots 3 and 4, which makes it unlikely that the observed emissions included any enhanced emissions of MTs close to the bud break. Besides, saplings planted on plots 2 and 3 had developed their leaves before they were put in the ground, and therefore, no elevated MT emissions were expected from them due to the changing processes during bud break and leaf development.
The sesquiterpenes were the group that contributed least to the total BVOC emissions. The standardized emissions were 2.51 ± 2.03 µg g
dw−1 h
−1. Sesquiterpenes are, in general, less studied when measuring emissions from Salix. Toome et al. [
66] observed emissions of α-copaene, (E,E)-α-farnesene and α-murolene from rust-infected leaves, but not from control leaves. Emissions of α-copaene and α-farnesene have also been seen for wild growing Salix species [
67]. α-farnesene was emitted from all varieties in this study, but no visible sign of rust was seen from the measured leaves.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}