The Diversity, Multiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Clearance
2.2. Study Site and Sample Collection
2.3. DNA Extraction
2.4. Msp2 Genotyping
2.5. Microsatellite Analysis
2.6. Data Analyses
3. Results
3.1. Demographics of Study Participants
3.2. Parasite Diversity and Multiplicity of Infection (MOI)
3.2.1. Msp2 Estimates of MOI
3.2.2. Msp2 Estimates of Genetic Diversity
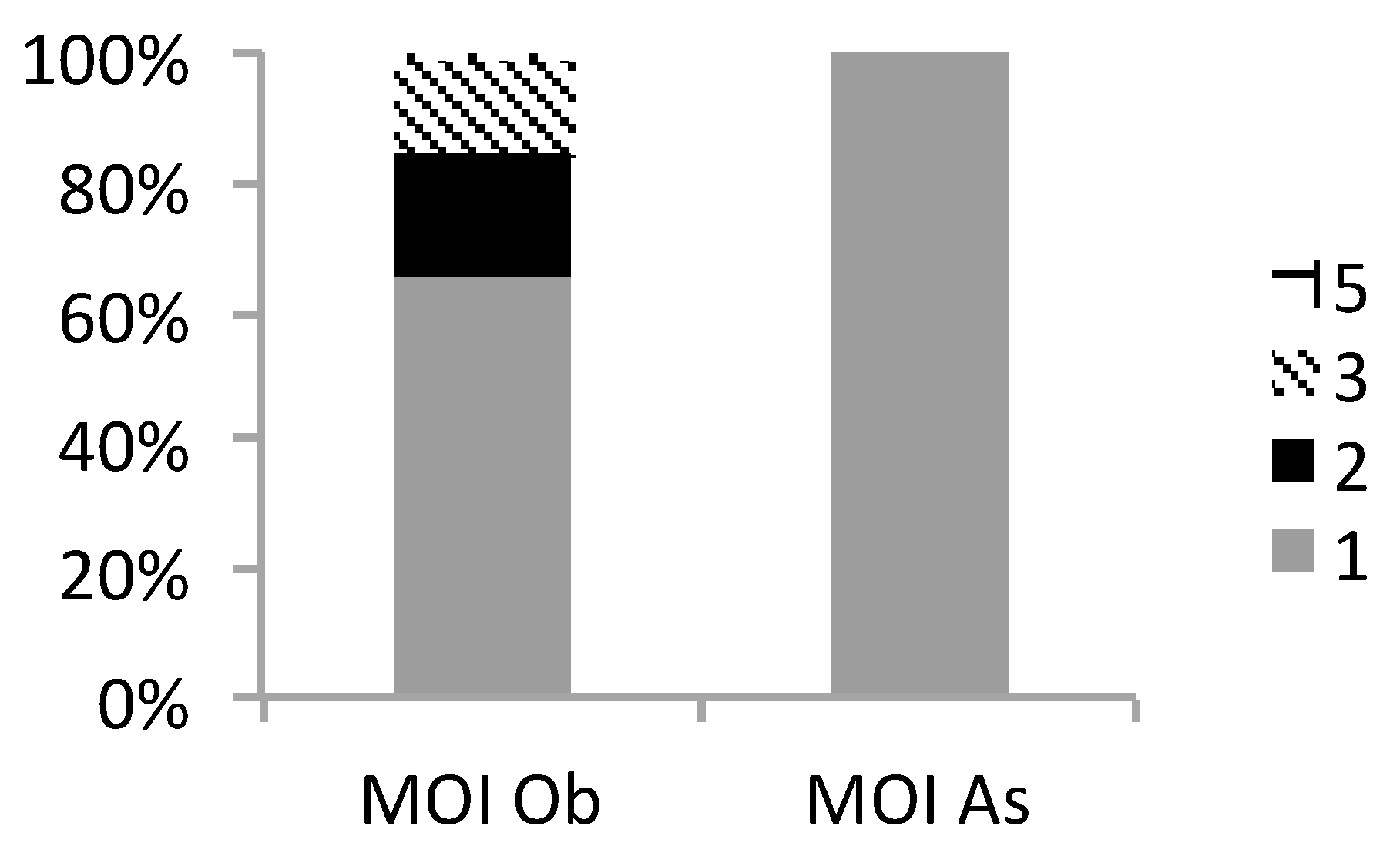
3.2.3. MOI Determined by Microsatellite (MS) Analysis
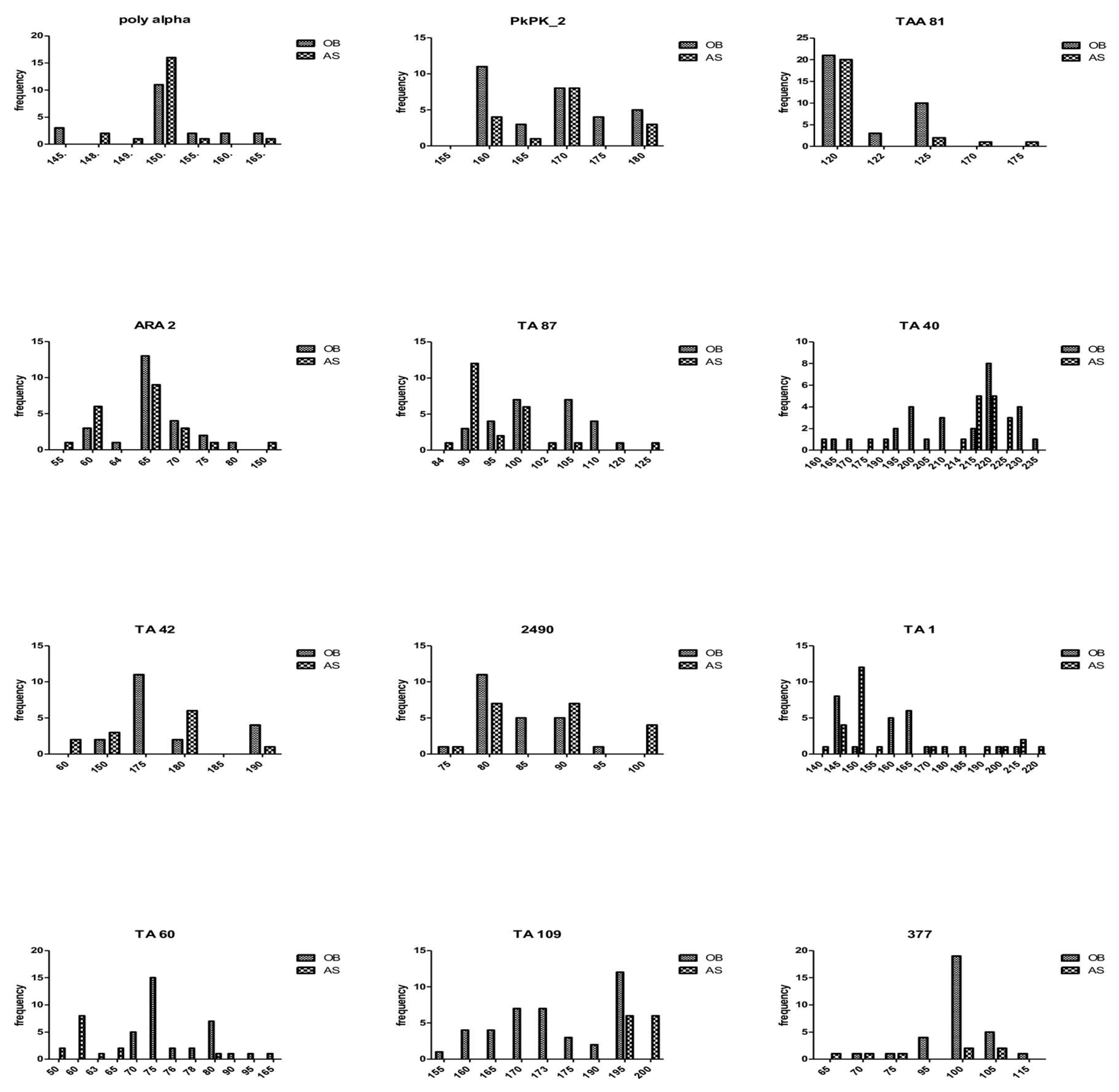
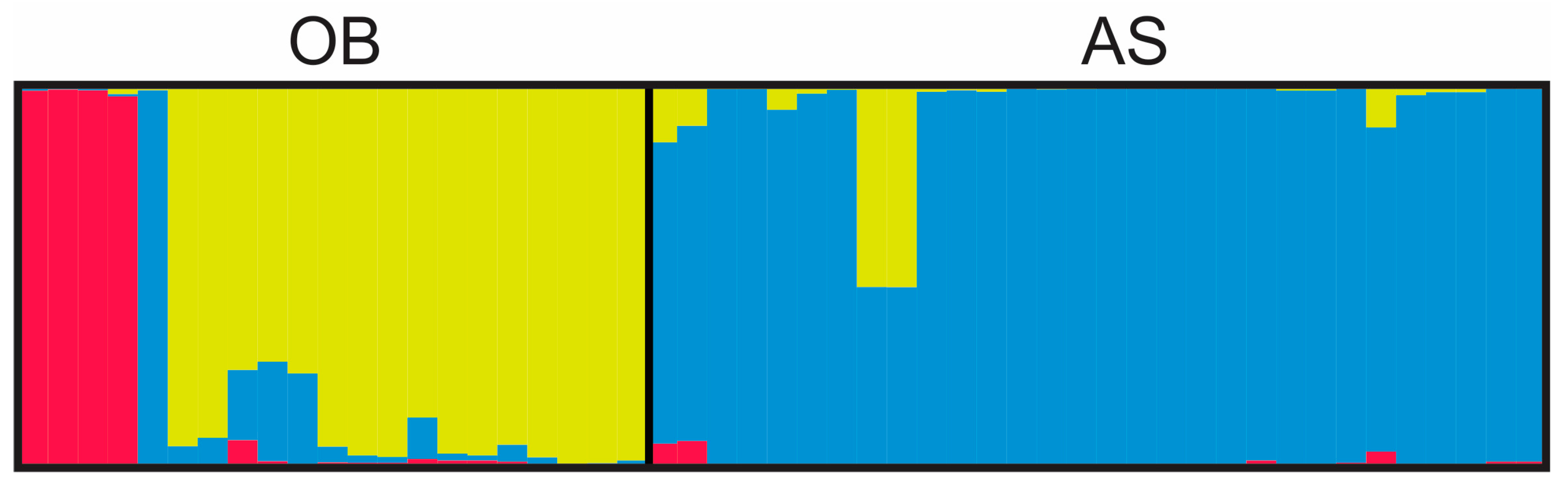
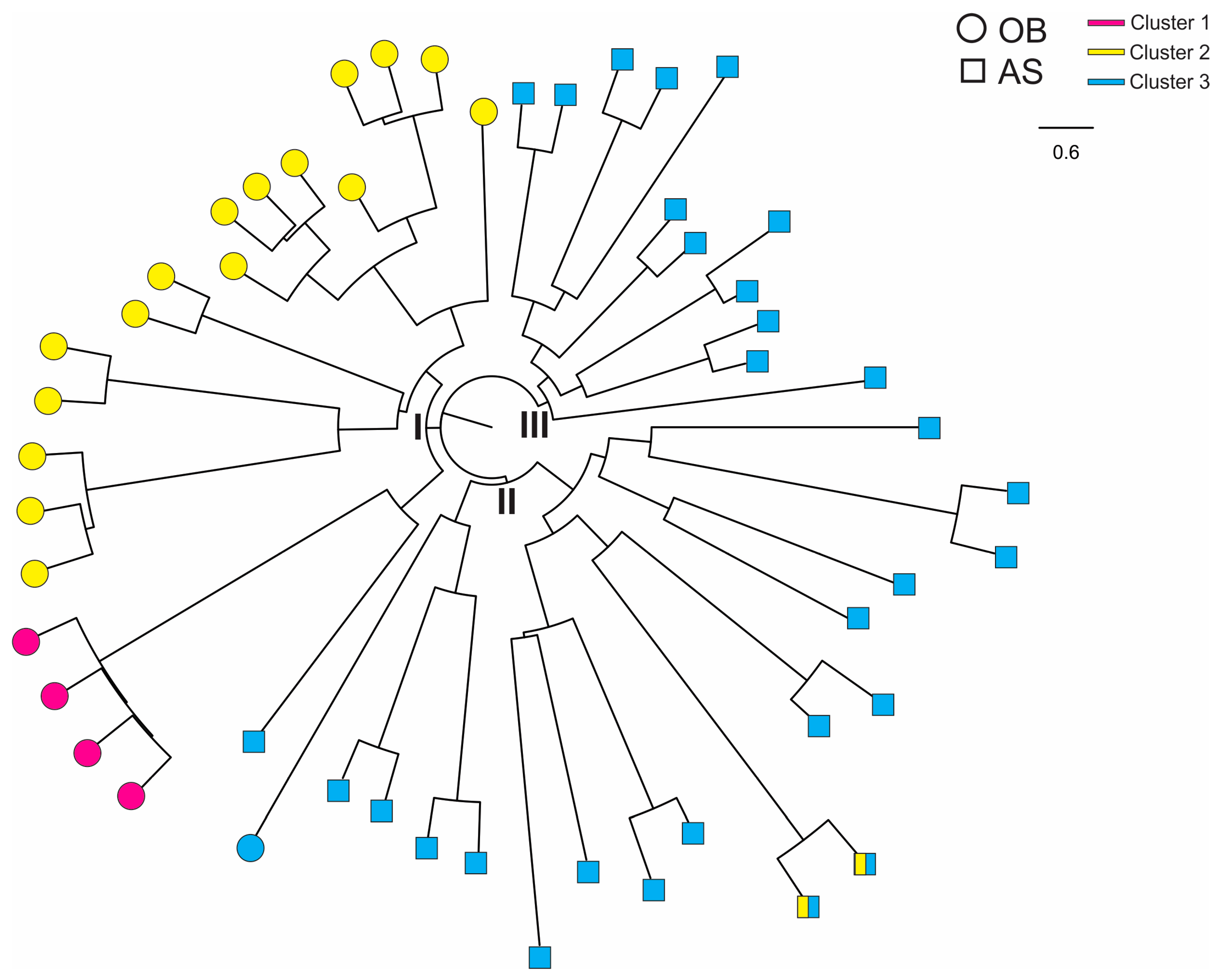
3.3. Genetic Diversity Estimated by Microsatellite Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GNA Ghana Health Service Marks World Malaria Day; Accra, Ghana. 2017. Available online: https://www.afro.who.int/news/ghana-commemorates-world-malaria-day (accessed on 16 February 2019).
- MOH. National Malaria Control Programme, World Malaria Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- GHS. 2016 Annual Report; Ghana Health Service: Accra, Ghana, 2017; pp. 1–129. [Google Scholar]
- Ingasia, L.A.; Cheruiyot, J.; Okoth, S.A.; Andagalu, B.; Kamau, E. Genetic variability and population structure of Plasmodium falciparum parasite populations from different malaria ecological regions of Kenya. Infect. Genet. Evolut. 2016, 39, 372–380. [Google Scholar] [CrossRef]
- Tadesse, F.G.; Slater, H.C.; Chali, W.; Teelen, K.; Lanke, K.; Belachew, M.; Menberu, T.; Shumie, G.; Shitaye, G.; Okell, L.C. The relative contribution of symptomatic and asymptomatic Plasmodium vivax and Plasmodium falciparum infections to the infectious reservoir in a low-endemic setting in Ethiopia. Clin. Infect. Dis. 2018, 66, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Laishram, D.D.; Sutton, P.L.; Nanda, N.; Sharma, V.L.; Sobti, R.C.; Carlton, J.M.; Joshi, H. The complexities of malaria disease manifestations with a focus on asymptomatic malaria. Malar. J. 2012, 11, 29. [Google Scholar] [CrossRef]
- Lindblade, K.A.; Steinhardt, L.; Samuels, A.; Kachur, S.P.; Slutsker, L. The silent threat: Asymptomatic parasitemia and malaria transmission. Expert Rev. Anti-Infect. Ther. 2013, 11, 623–639. [Google Scholar] [CrossRef] [PubMed]
- Alves, F.P.; Gil, L.H.; Marrelli, M.T.; Ribolla, P.E.; Camargo, E.P.; Da Silva, L.H. Asymptomatic carriers of Plasmodium spp. as infection source for malaria vector mosquitoes in the Brazilian Amazon. J. Med. Entomol. 2005, 42, 777–779. [Google Scholar] [CrossRef]
- Menard, D.; Dondorp, A. Antimalarial Drug Resistance: A Threat to Malaria Elimination. Cold Spring Harb. Perspect. Med. 2017, 7, a025619. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, J.; Zhao, Y.; Liu, Q.; He, Y.; Zhang, J.; Yang, Z.; Fan, Q.; Wang, Q.; Cui, L.; et al. Risk factors for asymptomatic malaria infections from seasonal cross-sectional surveys along the China–Myanmar border. Malar. J. 2018, 17, 1–13. [Google Scholar] [CrossRef]
- Babiker, H.; Ranford-Cartwright, L.C.; Currie, D.; Charlwood, J.; Billingsley, P.; Teuscher, T.; Walliker, D. Random mating in a natural population of the malaria parasite Plasmodium falciparum. Parasitology 1994, 109, 413–421. [Google Scholar] [CrossRef]
- McKenzie, F.E.; Ferreira, M.U.; Baird, J.K.; Snounou, G.; Bossert, W.H. Meiotic recombination, cross-reactivity, and persistence in Plasmodium falciparum. Evolution 2001, 55, 1299–1307. [Google Scholar] [CrossRef]
- Blasco, B.; Leroy, D.; Fidock, D.A. Antimalarial drug resistance: Linking Plasmodium falciparum parasite biology to the clinic. Nat. Med. 2017, 23, 917. [Google Scholar] [CrossRef]
- Felger, I.; Irion, A.; Steiger, S.; Beck, H.-P. 2. Genotypes of Merozoite Surface protein 2 of Plasmodium falciparum in Tanzania. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 3–9. [Google Scholar] [CrossRef]
- Raj, D.; Das, A.; Supakar, P. Genetic diversity in the merozoite surface protein 1 gene of P. falciparum in different malaria-endemic localities. Am. J. Trop. Med. Hyg. 2004, 71, 285–289. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Takala, S.L.; Coulibaly, D.; Thera, M.A.; Dicko, A.; Smith, D.L.; Guindo, A.B.; Kone, A.K.; Traore, K.; Ouattara, A.; Djimde, A.A.; et al. Dynamics of polymorphism in a malaria vaccine antigen at a vaccine-testing site in Mali. PLoS Med. 2007, 4, e93. [Google Scholar] [CrossRef] [PubMed]
- Neafsey, D.E.; Juraska, M.; Bedford, T.; Benkeser, D.; Valim, C.; Griggs, A.; Lievens, M.; Abdulla, S.; Adjei, S.; Agbenyega, T.; et al. Genetic Diversity and Protective Efficacy of the RTS,S/AS01 Malaria Vaccine. N. Engl. J. Med. 2015, 373, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Muleba, M.; Stevenson, J.C.; Pringle, J.C.; Norris, D.E. Beyond the entomological inoculation rate: Characterizing multiple blood feeding behavior and Plasmodium falciparum multiplicity of infection in Anopheles mosquitoes in northern Zambia. Parasites Vectors 2017, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Amoah, L.; Nuvor, S.; Obboh, E.; Acquah, F.; Asare, K.; Singh, S.; Boampong, J.; Theisen, M.; Williamson, K. Natural antibody responses to Plasmodium falciparum MSP3 and GLURP (R0) antigens are associated with low parasite densities in malaria patients living in the Central Region of Ghana. Parasites Vectors 2017, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, L.C.; Mideo, N.; Drew, D.R.; Schneider, P.; Colegrave, N.; Reece, S.E. Competition and the evolution of reproductive restraint in malaria parasites. Am. Nat. 2011, 177, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Josling, G.A.; Llinás, M. Sexual development in Plasmodium parasites: Knowing when it’s time to commit. Nat. Rev. Microbiol. 2015, 13, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Oyebola, M.K.; Idowu, E.T.; Nyang, H.; Olukosi, Y.A.; Otubanjo, O.A.; Nwakanma, D.C.; Awolola, S.T.; Amambua-Ngwa, A. Microsatellite markers reveal low levels of population sub-structuring of Plasmodium falciparum in southwestern Nigeria. Malar. J. 2014, 13, 493. [Google Scholar] [CrossRef]
- Ferdig, M.T.; Su, X.Z. Microsatellite markers and genetic mapping in Plasmodium falciparum. Parasitol. Today 2000, 16, 307–312. [Google Scholar] [CrossRef]
- Anderson, T.J.; Haubold, B.; Williams, J.T.; Estrada-Franco, J.G.; Richardson, L.; Mollinedo, R.; Bockarie, M.; Mokili, J.; Mharakurwa, S.; French, N. Microsatellite markers reveal a spectrum of population structures in the malaria parasite Plasmodium falciparum. Mol. Biol. Evol. 2000, 17, 1467–1482. [Google Scholar] [CrossRef] [PubMed]
- Barry, A.E.; Schultz, L.; Senn, N.; Nale, J.; Kiniboro, B.; Siba, P.M.; Mueller, I.; Reeder, J.C. High levels of genetic diversity of Plasmodium falciparum populations in Papua New Guinea despite variable infection prevalence. Am. J. Trop. Med. Hyg. 2013, 88, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Nabet, C.; Doumbo, S.; Jeddi, F.; Konaté, S.; Manciulli, T.; Fofana, B.; L’Ollivier, C.; Camara, A.; Moore, S.; Ranque, S. Genetic diversity of Plasmodium falciparum in human malaria cases in Mali. Malar. J. 2016, 15, 353. [Google Scholar] [CrossRef] [PubMed]
- Razak, M.R.M.A.; Sastu, U.R.; Norahmad, N.A.; Abdul-Karim, A.; Muhammad, A.; Muniandy, P.K.; Jelip, J.; Rundi, C.; Imwong, M.; Mudin, R.N. Genetic diversity of Plasmodium falciparum populations in malaria declining areas of Sabah, East Malaysia. PLoS ONE 2016, 11, e0152415. [Google Scholar]
- Felger, I.; Tavul, L.; Kabintik, S.; Marshall, V.; Genton, B.; Alpers, M.; Beck, H.-P. Plasmodium falciparum: extensive polymorphism in merozoite surface antigen 2 alleles in an area with endemic malaria in Papua New Guinea. Exp. Parasitol. 1994, 79, 106–116. [Google Scholar] [CrossRef]
- Baniecki, M.L.; Faust, A.L.; Schaffner, S.F.; Park, D.J.; Galinsky, K.; Daniels, R.F.; Hamilton, E.; Ferreira, M.U.; Karunaweera, N.D.; Serre, D. Development of a single nucleotide polymorphism barcode to genotype Plasmodium vivax infections. PLoS Negl. Trop. Dis. 2015, 9, e0003539. [Google Scholar] [CrossRef]
- Mwingira, F.; Nkwengulila, G.; Schoepflin, S.; Sumari, D.; Beck, H.-P.; Snounou, G.; Felger, I.; Olliaro, P.; Mugittu, K. Plasmodium falciparum msp1, msp2 and glurp allele frequency and diversity in sub-Saharan Africa. Malar. J. 2011, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.J.; Su, X.Z.; Bockarie, M.; Lagog, M.; Day, K.P. Twelve microsatellite markers for characterization of Plasmodium falciparum from finger-prick blood samples. Parasitology 1999, 119, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.A.; Lal, A.A.; Ayala, F.J. Genetic Polymorphism and natural selection in the malaria parasites Plasmodium falciparum. Genetics 1998, 149, 189–202. [Google Scholar]
- Hughes, A.L. Positive selection and interallelic recombination at the merozoite surface antigen-1 (MSA-1) locus of Plasmodium falciparum. Mol. Biol. Evol. 1992, 9, 381–393. [Google Scholar]
- Greenhouse, B.; Myrick, A.; Dokomajilar, C.; Woo, J.M.; Carlson, E.J.; Rosenthal, P.J.; Dorsey, G. Validation of microsatellite markers for use in genotyping polyclonal Plasmodium falciparum infections. Am. J. Trop. Med. Hyg. 2006, 75, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Amoah, L.E.; Opong, A.; Ayanful-Torgby, R.; Abankwa, J.; Acquah, F.K. Prevalence of G6PD deficiency and Plasmodium falciparum parasites in asymptomatic school children living in southern Ghana. Malar. J. 2016, 15, 388. [Google Scholar] [CrossRef] [PubMed]
- Adu, B.; Dodoo, D.; Adukpo, S.; Hedley, P.L.; Arthur, F.K.; Gerds, T.A.; Larsen, S.O.; Christiansen, M.; Theisen, M. Fc γ receptor IIIB (FcgammaRIIIB) polymorphisms are associated with clinical malaria in Ghanaian children. PLoS ONE 2012, 7, e46197. [Google Scholar] [CrossRef] [PubMed]
- WHO. Giemsa Staining of Malaria Blood Films. In malaria microscopy standard operating procedure–MM-SOP-07A. 2016. Available online: https://apps.who.int/iris/handle/10665/274382 (accessed on 16 February 2019).
- Adjah, J.; Fiadzoe, B.; Ayanful-Torgby, R.; Amoah, L.E. Seasonal variations in Plasmodium falciparum genetic diversity and multiplicity of infection in asymptomatic children living in southern Ghana. Bmc Infect. Dis. 2018, 18, 432. [Google Scholar] [CrossRef] [PubMed]
- Ayanful-Torgby, R.; Oppong, A.; Abankwa, J.; Acquah, F.; Williamson, K.C.; Amoah, L.E. Plasmodium falciparum genotype and gametocyte prevalence in children with uncomplicated malaria in coastal Ghana. Malar. J. 2016, 15, 592. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.; Mindaye, T.; Belayneh, M.; Kassa, M.; Assefa, A.; Tadesse, M.; Woyessa, A.; Mengesha, T.; Kebede, A. Genetic diversity of Plasmodium falciparum isolates based on MSP-1 and MSP-2 genes from Kolla-Shele area, Arbaminch Zuria District, southwest Ethiopia. Malar. J. 2015, 14, 73. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2007, 1, 47–50. [Google Scholar] [CrossRef]
- Meirmans, P.G.; Van Tienderen, P.H. GenoType and GenoDive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Note 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Makarenkov, V. T-REX: Reconstructing and visualizing phylogenetic trees and reticulation networks. Bioinformatics 2001, 17, 664–668. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Note 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Agyeman-Budu, A.; Brown, C.; Adjei, G.; Adams, M.; Dosoo, D.; Dery, D.; Wilson, M.; Asante, K.P.; Greenwood, B.; Owusu-Agyei, S. Trends in multiplicity of Plasmodium falciparum infections among asymptomatic residents in the middle belt of Ghana. Malar. J. 2013, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Duah, N.O.; Matrevi, S.A.; Quashie, N.B.; Abuaku, B.; Koram, K.A. Genetic diversity of Plasmodium falciparum isolates from uncomplicated malaria cases in Ghana over a decade. Parasit Vectors 2016, 9, 416. [Google Scholar] [CrossRef] [PubMed]
- Bushman, M.; Morton, L.; Duah, N.; Quashie, N.; Abuaku, B.; Koram, K.A.; Dimbu, P.R.; Plucinski, M.; Gutman, J.; Lyaruu, P.; et al. Within-host competition and drug resistance in the human malaria parasite Plasmodium falciparum. Proc. Biol. Sci. 2016, 283, 20153038. [Google Scholar] [CrossRef]
- Rorick, M.M.; Artzy-Randrup, Y.; Ruybal-Pesantez, S.; Tiedje, K.E.; Rask, T.S.; Oduro, A.; Ghansah, A.; Koram, K.; Day, K.P.; Pascual, M. Signatures of competition and strain structure within the major blood-stage antigen of Plasmodium falciparum in a local community in Ghana. Ecol. Evol. 2018, 8, 3574–3588. [Google Scholar] [CrossRef] [PubMed]
- Kusi, K.A.; Manu, E.A.; Manful Gwira, T.; Kyei-Baafour, E.; Dickson, E.K.; Amponsah, J.A.; Remarque, E.J.; Faber, B.W.; Kocken, C.H.M.; Dodoo, D.; et al. Variations in the quality of malaria-specific antibodies with transmission intensity in a seasonal malaria transmission area of Northern Ghana. PLoS ONE 2017, 12, e0185303. [Google Scholar] [CrossRef] [PubMed]
- Mara, S.E.; Silué, K.D.; Raso, G.; N’Guetta, S.P.; N’Goran, E.K.; Tanner, M.; Utzinger, J.; Ding, X.C. Genetic diversity of Plasmodium falciparum among school-aged children from the Man region, western Côte d’Ivoire. Malar. J. 2013, 12, 419. [Google Scholar] [CrossRef]
- Mayor, A.; Saute, F.; Aponte, J.J.; Almeda, J.; Gómez-Olivé, F.X.; Dgedge, M.; Alonso, P.L. Plasmodium falciparum multiple infections in Mozambique, its relation to other malariological indices and to prospective risk of malaria morbidity. Trop. Med. Int. Health 2003, 8, 3–11. [Google Scholar] [CrossRef]
- Smith, T.; Beck, H.-P.; Kitua, A.; Mwankusye, S.; Felger, I.; Fraser-Hurt, N.; Irion, A.; Alonso, P.; Teuscher, T.; Tanner, M. 4. Age dependence of the multiplicity of Plasmodium falciparum infections and of other malariological indices in an area of high endemicity. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 15–20. [Google Scholar] [CrossRef]
- Mobegi, V.A.; Loua, K.M.; Ahouidi, A.D.; Satoguina, J.; Nwakanma, D.C.; Amambua-Ngwa, A.; Conway, D.J. Population genetic structure of Plasmodium falciparum across a region of diverse endemicity in West Africa. Malar. J. 2012, 11, 223. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.A.; Lopez-Perez, M.; Vallejo, A.F.; Herrera, S.; Arévalo-Herrera, M.; Escalante, A.A. Multiplicity of infection and disease severity in Plasmodium vivax. PLoS Negl. Trop. Dis. 2016, 10, e0004355. [Google Scholar] [CrossRef] [PubMed]
- Beck, S.; Mockenhaupt, F.P.; Bienzle, U.; Eggelte, T.A.; Thompson, W.; Stark, K. Multiplicity of Plasmodium falciparum infection in pregnancy. Am. J. Trop. Med. Hyg. 2001, 65, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Nicastri, E.; Paglia, M.G.; Severini, C.; Ghirga, P.; Bevilacqua, N.; Narciso, P. Plasmodium falciparum multiple infections, disease severity and host characteristics in malaria affected travellers returning from Africa. Travel Med. Infect. Dis. 2008, 6, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Rout, R.; Mohapatra, B.N.; Kar, S.K.; Ranjit, M. Genetic complexity and transmissibility of Plasmodium falciparum parasites causing severe malaria in central-east coast India. Trop. Biomed. 2009, 26, 165–172. [Google Scholar]
- Congpuong, K.; Sukaram, R.; Prompan, Y.; Dornae, A. Genetic diversity of the msp-1, msp-2, and glurp genes of P lasmodium falciparum isolates along the Thai-Myanmar borders. Asian Pac. J. Trop. Biomed. 2014, 4, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Kidima, W.; Nkwengulila, G. Plasmodium falciparum msp2 genotypes and multiplicity of infections among children under five years with uncomplicated malaria in Kibaha, Tanzania. J. Parasitol. Res. 2015, 2015. [Google Scholar]
- Gupta, V.; Dorsey, G.; Hubbard, A.E.; Rosenthal, P.J.; Greenhouse, B. Gel versus capillary electrophoresis genotyping for categorizing treatment outcomes in two anti-malarial trials in Uganda. Malar. J. 2010, 9, 19. [Google Scholar] [CrossRef]
- Conway, D.J.; Machado, R.L.; Singh, B.; Dessert, P.; Mikes, Z.S.; Povoa, M.M.; Oduola, A.M.; Roper, C. Extreme geographical fixation of variation in the Plasmodium falciparum gamete surface protein gene Pfs48/45 compared with microsatellite loci. Mol. Biochem. Parasitol. 2001, 115, 145–156. [Google Scholar] [CrossRef]
- Su, X.-Z.; Wellems, T.E. Toward a High-Resolution Plasmodium falciparum Linkage Map: Polymorphic Markers from Hundreds of Simple Sequence Repeats. Genomics 1996, 33, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Fola, A.A.; Harrison, G.A.; Hazairin, M.H.; Barnadas, C.; Hetzel, M.W.; Iga, J.; Siba, P.M.; Mueller, I.; Barry, A.E. Higher complexity of infection and genetic diversity of Plasmodium vivax than Plasmodium falciparum across all malaria transmission zones of Papua New Guinea. Am. J. Trop. Med. Hyg. 2017, 96, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Peyerl-Hoffmann, G.; Jelinek, T.; Kilian, A.; Kabagambe, G.; Metzger, W.; Von Sonnenburg, F. Genetic diversity of Plasmodium falciparum and its relationship to parasite density in an area with different malaria endemicities in West Uganda. Trop. Med. Int. Health 2001, 6, 607–613. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Obom (80) | Asutsuare (80) | p Value |
|---|---|---|---|
| Age (yrs) Mean (SEM) | 17.6 (1.1) | 19.6 (1.2) | |
| Min–Max | 6–45 | 3–66 | |
| Males N (%) | 41 (52%) | 43 (54%) | |
| Microscopy N (%) | 27 (33.75%) | 3 (3.75) | 0.001 |
| Positive, N (%) | 27 (33.75%) | 3 (3.75) | <0.001 |
| Min–Max | 32–5080 | 16–400 | |
| Geometric mean PD (95% Cl) | 318.8 (180.6–562.5) | * |
| Marker | F (n) | Range (bp) | MOI (GM) | |
|---|---|---|---|---|
| Obom (61) | ||||
| 3D7 | 68 (6) | 200–600 | 1.37 | |
| FC27 | 24 (4) | 300–700 | ||
| Asutsuare (7) | ||||
| 3D7 | 5 (3) | 300–500 | 1 | |
| FC27 | 2 (2) | 300–400 | ||
| Dataset | Study Site | Samples (n) | Effective Alleles (n) | Heterozygosity |
|---|---|---|---|---|
| Genotype | ||||
| AS | 21 * | 2.961 | 0.67 | |
| OB | 30 * | 3.384 | 0.694 | |
| Binary | ||||
| OB | 35 | 1.233 | 0.158 | |
| AS | 25 | 1.196 | 0.135 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abukari, Z.; Okonu, R.; Nyarko, S.B.; Lo, A.C.; Dieng, C.C.; Salifu, S.P.; Gyan, B.A.; Lo, E.; Amoah, L.E. The Diversity, Multiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana. Genes 2019, 10, 434. https://doi.org/10.3390/genes10060434
Abukari Z, Okonu R, Nyarko SB, Lo AC, Dieng CC, Salifu SP, Gyan BA, Lo E, Amoah LE. The Diversity, Multiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana. Genes. 2019; 10(6):434. https://doi.org/10.3390/genes10060434
Chicago/Turabian StyleAbukari, Zakaria, Ruth Okonu, Samuel B. Nyarko, Aminata C. Lo, Cheikh C. Dieng, Samson P. Salifu, Ben A. Gyan, Eugenia Lo, and Linda E. Amoah. 2019. "The Diversity, Multiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana" Genes 10, no. 6: 434. https://doi.org/10.3390/genes10060434
APA StyleAbukari, Z., Okonu, R., Nyarko, S. B., Lo, A. C., Dieng, C. C., Salifu, S. P., Gyan, B. A., Lo, E., & Amoah, L. E. (2019). The Diversity, Multiplicity of Infection and Population Structure of P. falciparum Parasites Circulating in Asymptomatic Carriers Living in High and Low Malaria Transmission Settings of Ghana. Genes, 10(6), 434. https://doi.org/10.3390/genes10060434