Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals

 , , , , ,
, , , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Characterization of the Study Object
2.2. Cytogenetics
2.3. Histological Analysis
2.4. RNA Extraction and cDNA Synthesis
2.5. Quantitative PCR in Real-Time (qRT-PCR)
2.6. Illumina Sequencing and Coverage Analysis of Sex-Associated Genes
3. Results
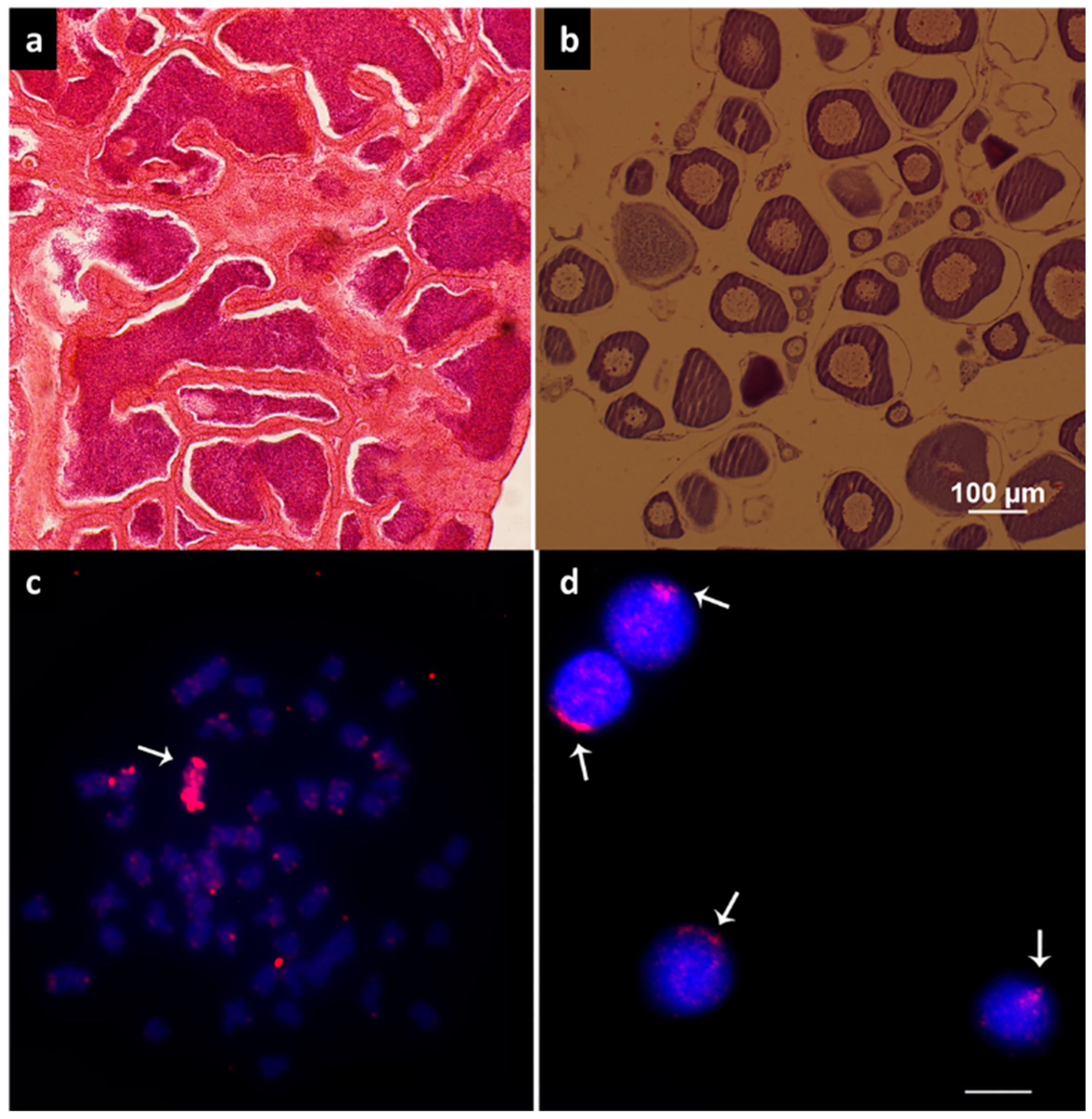
3.1. Histological Analysis and Cytogenetics
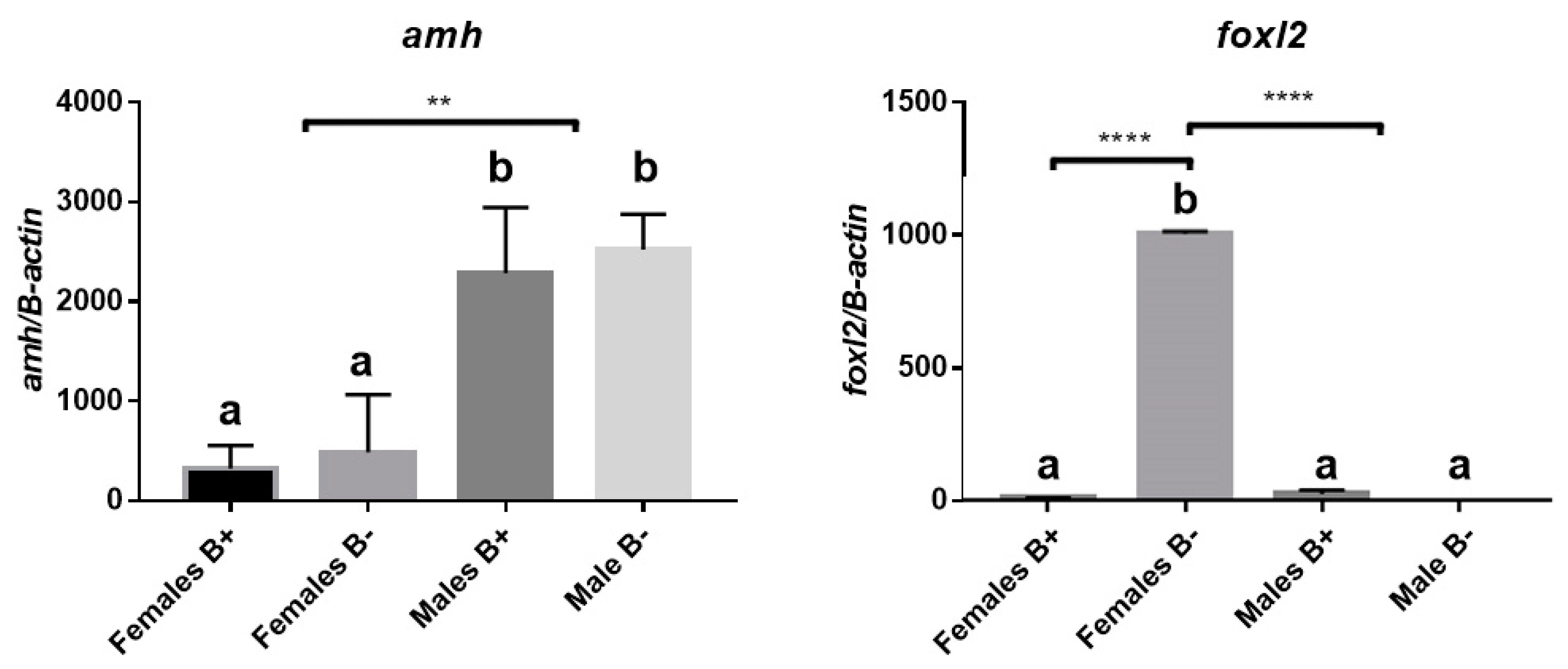
3.2. Analysis of Gene Expression of Amh and Foxl2a
3.3. Number of Gene Copies between Individuals with and without B Chromosomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trukhina, A.V.; Lukina, N.A.; Wackerow-Kouzova, N.D.; Smirnov, A.F. The variety of vertebrate mechanisms of sex determination. BioMed Res. Int. 2013, 2013, 587460. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Martínez, P.; Viñas, A.M.; Sánchez, L.; Díaz, N.; Ribas, L.; Piferrer, F. Genetic architecture of sex determination in fish: Applications to sex ratio control in aquaculture. Front. Genet. 2014, 5, 340. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Myosho, T.; Otake, H.; Masuyama, H.; Matsuda, M.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Tracing the emergence of a novel sex-determining gene in medaka, Oryzias luzonensis. Genetics 2012, 191, 163–170. [Google Scholar] [CrossRef]
- Yano, A.; Nicol, B.; Jouanno, E.; Quillet, E.; Fostier, A.; Guyomard, R.; Guiguen, Y. The sexually dimorphic on the Y-chromosome gene (sdY) is a conserved male-specific Y-chromosome sequence in many salmonids. Evol. Appl. 2013, 6, 486–496. [Google Scholar] [CrossRef]
- Hattori, R.S.; Murai, Y.; Oura, M.; Masuda, S.; Majhi, S.K.; Sakamoto, T.; Fernandino, J.I.; Somoza, G.M.; Yokota, M.; Strussmann, C.A. A Y-linked anti-Mullerian hormone duplication takes over a critical role in sex determination. Proc. Natl. Acad. Sci. USA 2012, 109, 2955–2959. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Zhang, Y.; Sarida, M.; Hattori, R.S.; Strüssmann, C.A. Coexistence of Genotypic and Temperature-Dependent Sex Determination in Pejerrey Odontesthes bonariensis. PLoS ONE 2014, 9, e102574. [Google Scholar] [CrossRef]
- Bej, D.K.; Miyoshi, K.; Hattori, R.S.; Strüssmann, C.A.; Yamamoto, Y. A Duplicated, Truncated amh Gene Is Involved in Male Sex Determination in an Old World Silverside. G3 Genes Genomes Genet. 2017, 7, 2489–2495. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yoshinaga, N.; Yazawa, T.; Gen, K.; Kitano, T. Cortisol Is Involved in Temperature-Dependent Sex Determination in the Japanese Flounder. Endocrinology 2010, 151, 3900–3908. [Google Scholar] [CrossRef]
- Haugen, T.; Almeida, F.F.; Andersson, E.; Bogerd, J.; Male, R.; Skaar, K.S.; Schulz, R.W.; Sørhus, E.; Wijgerde, T.; Taranger, G.L. Sex differentiation in Atlantic cod (Gadus morhua L.): Morphological and gene expression studies. Reprod. Biol. Endocrinol. 2012, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Mankiewicz, J.L.; Godwin, J.; Holler, B.L.; Turner, P.M.; Murashige, R.; Shamey, R.; Daniels, H.V.; Borski, R.J. Masculinizing Effect of Background Color and Cortisol in a Flatfish with Environmental Sex-Determination. Integr. Comp. Biol. 2013, 53, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, K.R. In search of determinants: Gene expression during gonadal sex differentiation. J. Fish Biol. 2010, 76, 1879–1902. [Google Scholar] [CrossRef] [PubMed]
- Eshel, O.; Shirak, A.; Dor, L.; Band, M.; Zak, T.; Markovich-Gordon, M.; Chalifa-Caspi, V.; Feldmesser, E.; Weller, J.I.; Seroussi, E.; et al. Identification of male-specific amh duplication, sexually differentially expressed genes and microRNAs at early embryonic development of Nile tilapia (Oreochromis niloticus). BMC Genom. 2014, 15, 774. [Google Scholar] [CrossRef] [PubMed]
- Pannetier, M. FOXL2 activates P450 aromatase gene transcription: Towards a better characterization of the early steps of mammalian ovarian development. J. Mol. Endocrinol. 2006, 36, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Bertho, S.; Pasquier, J.; Pan, Q.; Le Trionnaire, G.; Bobe, J.; Postlethwait, J.H.; Pailhoux, E.; Schartl, M.; Herpin, A.; Guiguen, Y. Foxl2 and Its Relatives Are Evolutionary Conserved Players in Gonadal Sex Differentiation. Sex. Dev. 2016, 10, 111–129. [Google Scholar] [CrossRef]
- Zhang, X.; Li, M.; Ma, H.; Liu, X.; Shi, H.; Li, M.; Wang, D. Mutation of foxl2 or cyp19a1a results in female to male sex reversal in XX Nile tilapia. Endocrinology 2017, 158, 2634–2647. [Google Scholar] [CrossRef]
- Caulier, M.; Brion, F.; Chadili, E.; Turies, C.; Piccini, B.; Porcher, J.M.; Guiguen, Y.; Hinfray, N. Localization of steroidogenic enzymes and Foxl2a in the gonads of mature zebrafish (Danio rerio). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 188, 96–106. [Google Scholar] [CrossRef]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef]
- Gery, J. Characoids of the World; TFH Publications: Neptune, NJ, USA, 1977. [Google Scholar]
- Eschemeyer, W. Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 16 March 2018).
- Tenorio, R.C.C.D.O.; Vitorino, C.D.A.; Souza, I.L.; Oliveira, C.; Venere, P.C. Comparative cytogenetics in Astyanax (Characiformes: Characidae) with focus on the cytotaxonomy of the group. Neotrop. Ichthyol. 2013, 11, 553–564. [Google Scholar] [CrossRef]
- Salvador, L.B.; Moreira-Filho, O. B chromosomes in Astyanax scabripinnis (Pisces, Characidae). Heredity. 1992, 69, 50–56. [Google Scholar] [CrossRef]
- Moreira-Filho, O.; Galetti, P.M.; Bertollo, L.A.C. B chromosomes in the fish Astyanax scabripinnis (Characidae, Tetragonopterinae): An overview in natural populations. Cytogenet. Genome Res. 2004, 106, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M. B chromosomes. In The Evolution of the Genome; Gregory, T.R., Ed.; Elsevier Academic Press: London, UK, 2005; pp. 223–286. ISBN 0123014638. [Google Scholar]
- Banaei-Moghaddam, A.M.; Martis, M.M.; Macas, J.; Gundlach, H.; Himmelbach, A.; Altschmied, L.; Mayer, K.F.X.; Houben, A. Genes on B chromosomes: Old questions revisited with new tools. Biochim. Biophys. Acta 2015, 1849, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Camacho, J.P.M.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Martins, C. The modern view of b chromosomes under the impact of high scale omics analyses. Cells 2019, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Domínguez, B.; Ruiz-Ruano, F.J.; Cabrero, J.; Corral, J.M.; López-León, M.D.; Sharbel, T.F.; Camacho, J.P.M. Protein-coding genes in B chromosomes of the grasshopper Eyprepocnemis plorans. Sci. Rep. 2017, 7, 45200. [Google Scholar] [CrossRef]
- Makunin, A.I.; Dementyeva, P.V.; Graphodatsky, A.S.; Volobouev, V.T.; Kukekova, A.V.; Trifonov, V.A. Genes on B chromosomes of vertebrates. Mol. Cytogenet. 2014, 7, 99. [Google Scholar] [CrossRef]
- Castro, J.P.; Hattori, R.S.; Yoshinaga, T.T.; Silva, D.M.Z.D.A.; Foresti, F.; Santos, M.H.; Almeida, M.C.; Artoni, R.F. Differential Expression of dmrt1 in Astyanax scabripinnis (Teleostei, Characidade) Is Correlated with B Chromosome Occurrence. Zebrafish 2018, 16, 182–188. [Google Scholar] [CrossRef]
- Vicente, V.E.; Moreira-Filho, O.; Camacho, J.P. Sex-ratio distortion associated with the presence of a B chromosome in Astyanax scabripinnis (Teleostei, Characidae). Cytogenet. Cell Genet. 1996, 74, 70–75. [Google Scholar] [CrossRef]
- Silva, D.M.Z.D.A.; Pansonato-Alves, J.C.; Utsunomia, R.; Araya-Jaime, C.; Ruiz-Ruano, F.J.; Daniel, S.N.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Porto-Foresti, F.; et al. Delimiting the Origin of a B Chromosome by FISH Mapping, Chromosome Painting and DNA Sequence Analysis in Astyanax paranae (Teleostei, Characiformes). PLoS ONE 2014, 9, e94896. [Google Scholar] [CrossRef]
- Porto-Foresti, F.; Oliveira, C.; Maistro, E.L.; Foresti, F. Estimated frequency of B-chromosomes and population density of Astyanax scabripinnis paranae in a small stream. Braz. J. Genet. 1997, 20. [Google Scholar] [CrossRef]
- Cornelio, D.; Castro, J.P.; Santos, M.H.; Vicari, M.R.; de Almeida, M.C.; Moreira-Filho, O.; Camacho, J.P.M.; Artoni, R.F. Hermaphroditism can compensate for the sex ratio in the Astyanax scabripinnis species complex (Teleostei: Characidae): Expanding the B chromosome study model. Rev. Fish Biol. Fish. 2017, 27, 681–689. [Google Scholar] [CrossRef]
- Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. Cytotaxonomic considerations on Hoplias lacerdae (Pisces, Erythrinidae). Braz. J. Genet. 1978, 1, 103–120. [Google Scholar]
- Vicari, M.R.; de Pistune Mello, H.F.; Castro, J.P.; de Almeida, M.C.; Bertollo, L.A.C.; Moreira-Filho, O.; Camacho, J.P.M.; Artoni, R.F. New insights on the origin of B chromosomes in Astyanax scabripinnis obtained by chromosome painting and FISH. Genetica 2011, 139, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Vazzoler, A.E.A.M. Biologia da Reprodução de Peixes Teleósteos: Teoria e Prática; EDUEM: Maringá, Brazil, 1996. [Google Scholar]
- Bruford, M.W.; Hanotte, O.; Brookfield, J.F.Y.; Burke, T. Single-locus and multilocus DNA fingerprinting. In Molecular Genetics Analyses of Populations: A Practical Approach; HoelzelL, A.R., Ed.; IRL Press: Oxford, UK, 1992; pp. 225–269. ISBN 0-19-963277-4. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Carvalho, M.L.; Oliveira, C.; Foresti, F. Nuclear DNA content of thirty species of Neotropical fishes. Genet. Mol. Biol. 1998, 21, 47–54. [Google Scholar] [CrossRef]
- Spargo, A.; Ponstingl, H.; Ning, Z. SSAHA2: Sequence Search and Alignment by Hashing Algorithm. Available online: https://www.sanger.ac.uk/science/tools/ssaha2-0 (accessed on 16 March 2018).
- Pfennig, F.; Standke, A.; Gutzeit, H.O. The role of Amh signaling in teleost fish—Multiple functions not restricted to the gonads. Gen. Comp. Endocrinol. 2015, 223, 87–107. [Google Scholar] [CrossRef]
- Poonlaphdecha, S.; Pepey, E.; Canonne, M.; de Verdal, H.; Baroiller, J.F.; D’Cotta, H. Temperature induced-masculinisation in the Nile tilapia causes rapid up-regulation of both dmrt1 and amh expressions. Gen. Comp. Endocrinol. 2013, 193, 234–242. [Google Scholar] [CrossRef]
- Morinaga, C.; Saito, D.; Nakamura, S.; Sasaki, T.; Asakawa, S.; Shimizu, N.; Mitani, H.; Furutani-Seiki, M.; Tanaka, M.; Kondoh, H. The hotei mutation of medaka in the anti-Mullerian hormone receptor causes the dysregulation of germ cell and sexual development. Proc. Natl. Acad. Sci. USA 2007, 104, 9691–9696. [Google Scholar] [CrossRef]
- Li, Y.; Jing, X.A.; Aldrich, J.C.; Clifford, C.; Chen, J.; Akbari, O.S.; Ferree, P.M. Unique sequence organization and small RNA expression of a “selfish” B chromosome. Chromosoma 2017, 126, 753–768. [Google Scholar] [CrossRef]
- Ramos, É.; Cardoso, A.L.; Brown, J.; Marques, D.F.; Fantinatti, B.E.A.; Cabral-de-Mello, D.C.; Oliveira, R.A.; O’Neill, R.J.; Martins, C. The repetitive DNA element BncDNA, enriched in the B chromosome of the cichlid fish Astatotilapia latifasciata, transcribes a potentially noncoding RNA. Chromosoma 2017, 126, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Maistro, E.L.; Foresti, F.; Oliveira, C. New Occurrence of a Macro-B-Chromosome in Astyanax-Scabripinnis-Paranae (Pisces, Characiformes, Characidae). Braz. J. Genet. 1994, 17, 153–156. [Google Scholar]
- Silva, D.M.Z.D.A.; Utsunomia, R.; Ruiz-Ruano, F.J.; Oliveira, C.; Foresti, F. The complete mitochondrial genome sequence of Astyanax paranae (Teleostei: Characiformes). Mitochondrial DNA Part B 2016, 1, 586–587. [Google Scholar] [CrossRef]
- Castro, J.P.; Moura, M.O.; Moreira-Filho, O.; Shibatta, O.A.; Santos, M.H.; Nogaroto, V.; Vicari, M.R.; de Almeida, M.C.; Artoni, R.F. Diversity of the Astyanax scabripinnis species complex (Teleostei: Characidae) in the Atlantic Forest, Brazil: Species limits and evolutionary inferences. Rev. Fish Biol. Fish. 2015, 25, 231–244. [Google Scholar] [CrossRef]
- Castro, J.P.; Moura, M.O.; Moreira-Filho, O.; Shibatta, O.A.; Santos, M.H.; Nogaroto, V.; Vicari, M.R.; de Almeida, M.C.; Artoni, R.F. Evidence of incipient speciation in Astyanax scabripinnis species complex (Teleostei: Characidae). Neotrop. Ichthyol. 2014, 12, 429–438. [Google Scholar] [CrossRef]
- Pandian, T.J. Sexuality in Fishes; Science Publishers Enfield/CRC Press: New Hampshire, UK, 2010; ISBN 9781578086856. [Google Scholar]
- Pandian, T.J. Genetic Sex Differentiation in Fish; Science Publishers Enfield/CRC Press: New Hampshire, UK, 2012; ISBN 9781466517103. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Use |
|---|---|---|
| ame_amh_Fw1 | CTGGGATGTTGAAGACGA | Conventional PCR |
| ame_amh_Rv1 | GAGGAATTAATCAGCTCCAGAA | Conventional PCR |
| ame_fox2_Fw1 | ACGTTCTTGGGCTCAGAGGA | Conventional PCR |
| ame_foxl2a_Rv1 | AGACTTGCCGGGTTGGAAGTG | Conventional PCR |
| amh_rtf1 | CCTCACTGCTCTTCCTGACG | qRT-PCR |
| amh_rtR1 | AAACACCCAACACAGCTTGC | qRT-PCR |
| foxl2a_rtF1 | ACCTGAGCCTTAACGAGTGC | qRT-PCR |
| foxl2a_rtR1 | ATGTCTTCACACGTCGGGTC | qRT-PCR |
| β-actin F_RT | ATCATGAAGTGCGACGTGGA | qRT-PCR |
| β-actin R_RT | TATTTACGCTCAGGTGGGGC | qRT-PCR |
| Gene | ♀ (B+) | ♀ (B−) | ♂ (B+) | ♂ (B−) | Total |
|---|---|---|---|---|---|
| amh | 3 | 3 | 3 | 3 | 12 |
| foxl2a | 3 | 3 | 3 | 3 | 12 |
| Total | 24 |
| Sex | Samples | Number of B | Number of Reads (mi) | Coverage |
|---|---|---|---|---|
| Male | 2 | 0 | 93.7 | 7.5 |
| 1 | 1 | 52.2 | 4.2 | |
| Female | 1 | 0 | 43.7 | 3.5 |
| 2 | 1 | 96 | 7.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, J.P.; Hattori, R.S.; Yoshinaga, T.T.; Silva, D.M.Z.d.A.; Ruiz-Ruano, F.J.; Foresti, F.; Santos, M.H.; Almeida, M.C.d.; Moreira-Filho, O.; Artoni, R.F. Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals. Genes 2019, 10, 909. https://doi.org/10.3390/genes10110909
Castro JP, Hattori RS, Yoshinaga TT, Silva DMZdA, Ruiz-Ruano FJ, Foresti F, Santos MH, Almeida MCd, Moreira-Filho O, Artoni RF. Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals. Genes. 2019; 10(11):909. https://doi.org/10.3390/genes10110909
Chicago/Turabian StyleCastro, Jonathan Pena, Ricardo Shohei Hattori, Túlio Teruo Yoshinaga, Duílio Mazzoni Zerbinato de Andrade Silva, Francisco J. Ruiz-Ruano, Fausto Foresti, Mateus Henrique Santos, Mara Cristina de Almeida, Orlando Moreira-Filho, and Roberto Ferreira Artoni. 2019. "Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals" Genes 10, no. 11: 909. https://doi.org/10.3390/genes10110909
APA StyleCastro, J. P., Hattori, R. S., Yoshinaga, T. T., Silva, D. M. Z. d. A., Ruiz-Ruano, F. J., Foresti, F., Santos, M. H., Almeida, M. C. d., Moreira-Filho, O., & Artoni, R. F. (2019). Differential Expression of Genes Related to Sexual Determination Can Modify the Reproductive Cycle of Astyanax scabripinnis (Characiformes: Characidae) in B Chromosome Carrier Individuals. Genes, 10(11), 909. https://doi.org/10.3390/genes10110909