Genetically Determined Physical Activity and Its Association with Circulating Blood Cells

 , , and
, , and
Abstract
1. Introduction
2. Material and Methods
2.1. UK Biobank Participants
2.2. Genotyping and Imputation
2.3. Single Nucleotide Polymorphisms
2.4. Statistical Analyses
3. Results
3.1. Population Characteristics
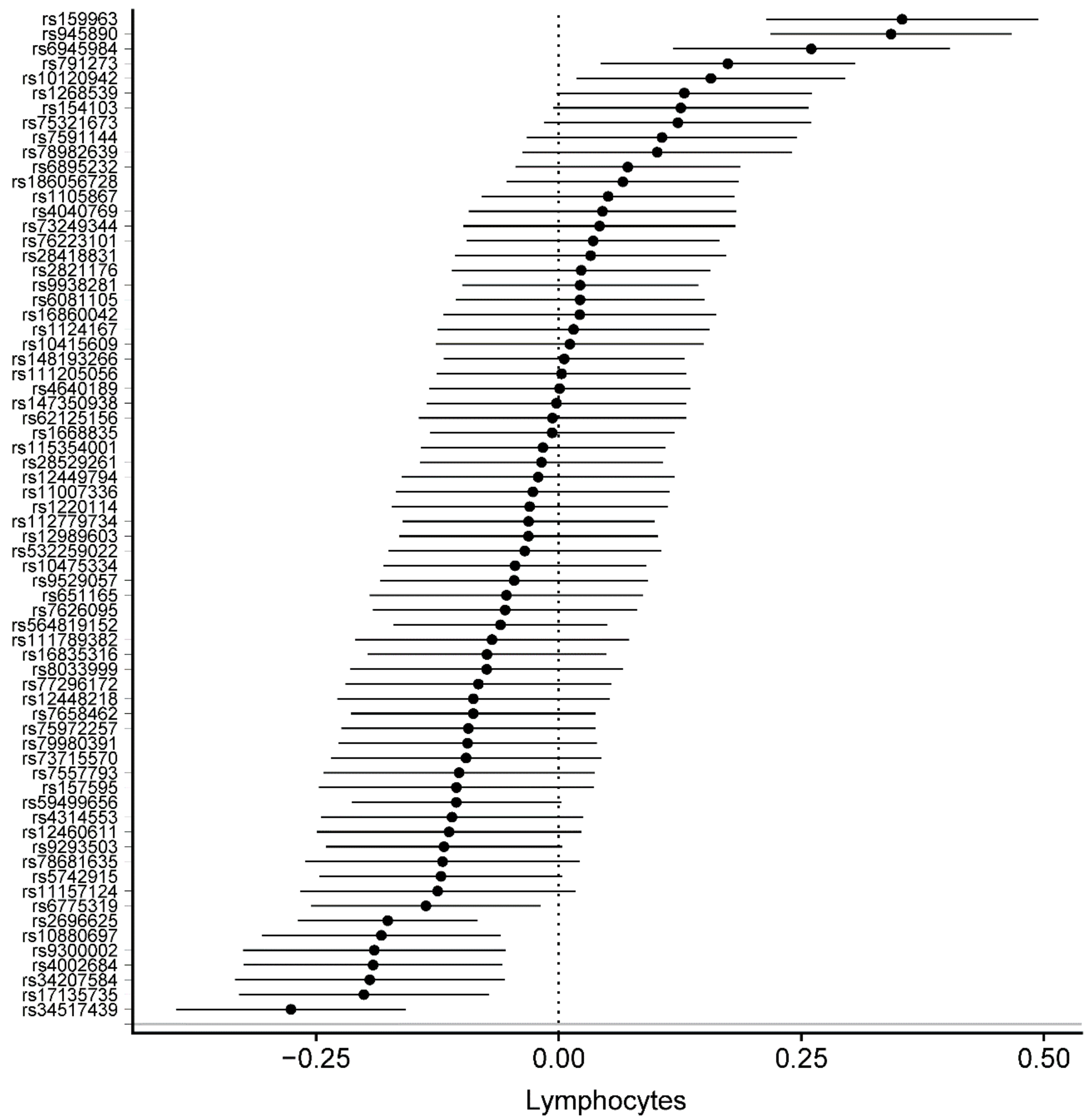
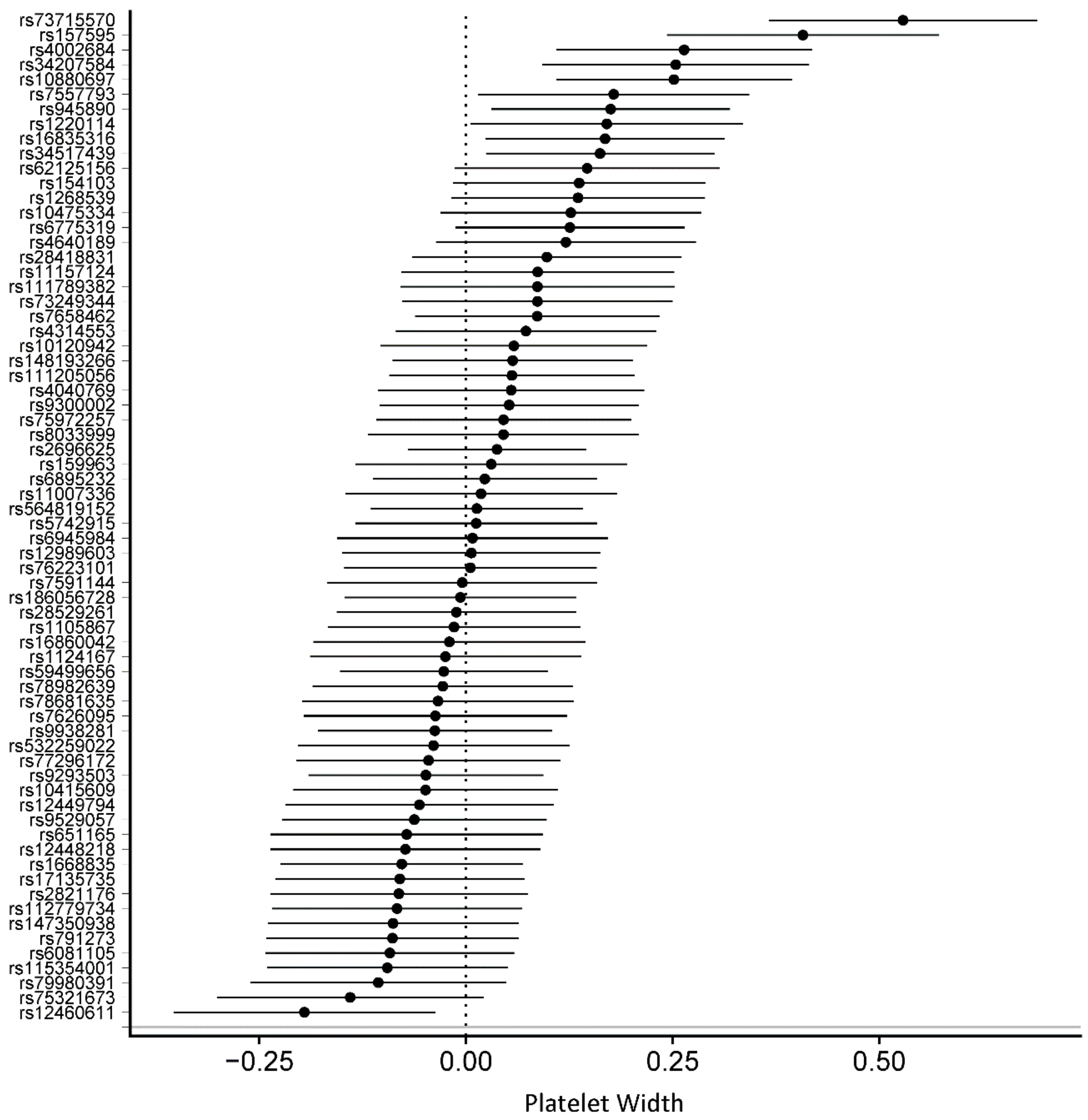
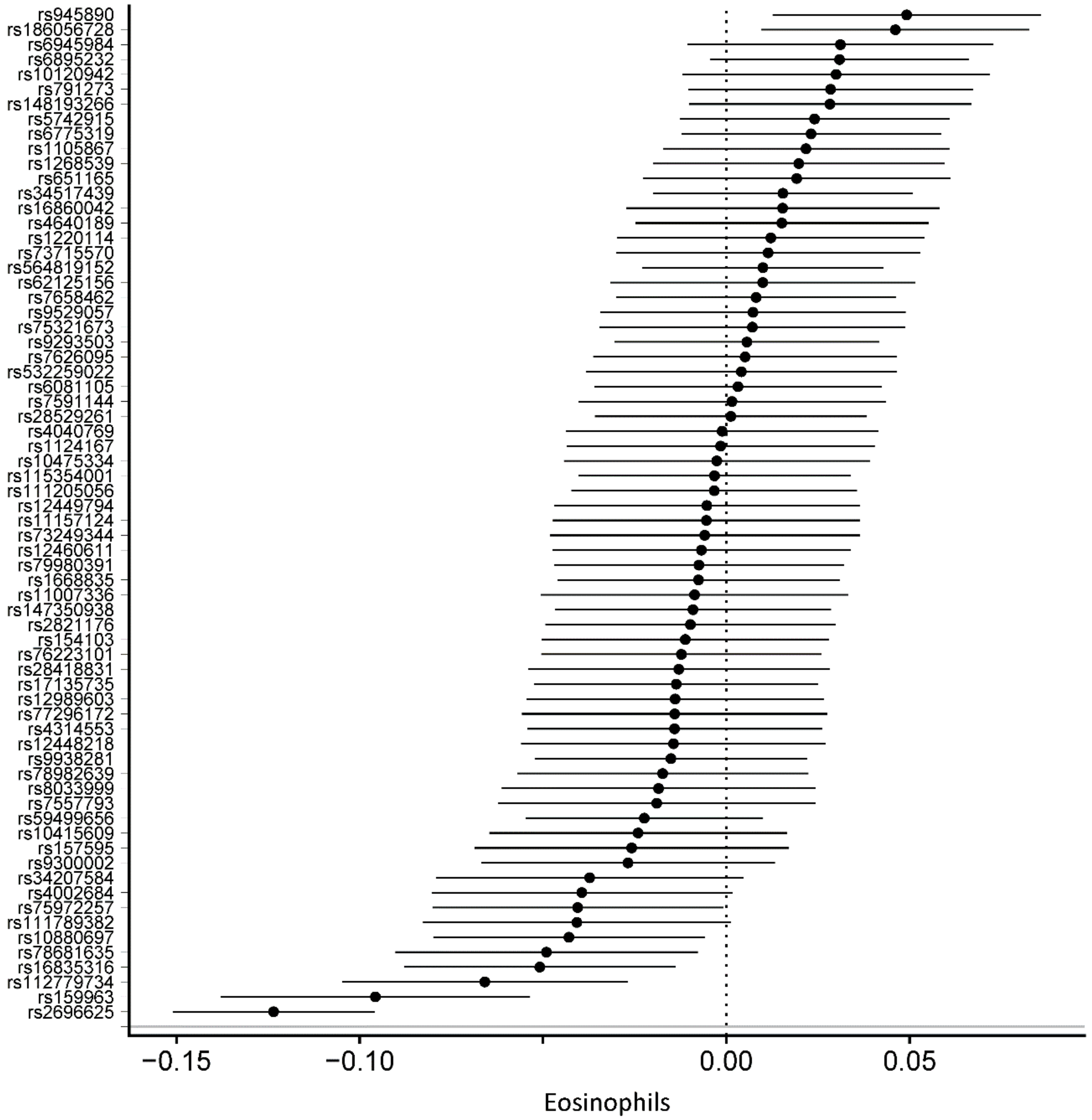
3.2. Genetically Determined Physical Activity and Circulating Blood Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lachman, S.; Boekholdt, S.M.; Luben, R.N.; Sharp, S.J.; Brage, S.; Khaw, K.-T.; Peters, R.J.; Wareham, N.J. Impact of physical activity on the risk of cardiovascular disease in middle-aged and older adults: EPIC Norfolk prospective population study. Eur. J. Prev. Cardiol. 2018, 25, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.; Mcnamara, E.; Tainio, M.; Hé, T.; De Sá, R.; Smith, A.D.; Sharp, S.J.; Edwards, P.; Woodcock, J.; Brage, S.; et al. Sedentary behaviour and risk of all-cause, cardiovascular and cancer mortality, and incident type 2 diabetes: A systematic review and dose response meta-analysis. Eur. J. Epidemiol. 2018, 33, 811–829. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.M.; Shiroma, E.J.; Lobelo, F.; Puska, P.; Blair, S.N.; Katzmarzyk, P.T.; Alkandari, J.R.; Andersen, L.B.; Bauman, A.E.; Brownson, R.C.; et al. Effect of physical inactivity on major non-communicable diseases worldwide: An analysis of burden of disease and life expectancy. Lancet 2012, 380, 219–229. [Google Scholar] [CrossRef]
- WHO Physical Activity and Adults. Available online: https://www.who.int/dietphysicalactivity/factsheet_adults/en/ (accessed on 1 July 2019).
- European Union. Special Eurobarometer 472. “Sport and Physical Activity”; European Union: Brussels, Belgium, 2018. [Google Scholar]
- Althoff, T.; Sosič, R.; Hicks, J.L.; King, A.C.; Delp, S.L.; Leskovec, J. Large-scale physical activity data reveal worldwide activity inequality. Nature 2017, 547, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Kolbe-Alexander, T.; Nguyen, B.; Katzmarzyk, P.T.; Pratt, M.; Lawson, K.D. The economic burden of physical inactivity: A systematic review and critical appraisal. Br. J. Sports Med. 2017, 51, 1392–1409. [Google Scholar] [CrossRef] [PubMed]
- Willis, E.A.; Shearer, J.J.; Matthews, C.E.; Hofmann, J.N. Association of physical activity and sedentary time with blood cell counts: National Health and Nutrition Survey 2003–2006. PLoS ONE 2018, 13, e0204277. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.N.; Deuster, P.A. Biological mechanisms underlying the role of physical fitness in health and resilience. Interface Focus 2014, 4, 20140040. [Google Scholar] [CrossRef] [PubMed]
- Davey Smith, G.; Ebrahim, S. ‘Mendelian randomization’: Can genetic epidemiology contribute to understanding environmental determinants of disease? Int. J. Epidemiol. 2003, 32, 1–22. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Doherty, A.; Smith-Byrne, K.; Ferreira, T.; Holmes, M.V.; Holmes, C.; Pulit, S.L.; Lindgren, C.M. GWAS identifies 14 loci for device-measured physical activity and sleep duration. Nat. Commun. 2018, 9, 5257. [Google Scholar] [CrossRef]
- UK Biobank. Coordinating Centre UK Biobank: Protocol for a Large-Scale Prospective Epidemiological Resource; UKBB-PROT-09-06 (Main Phase); UK Biobank Coordinating Centre: Stockport, UK, 2007; Volume 6, pp. 1–112. [Google Scholar]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; Connell, J.; et al. Genome-wide genetic data on ~500,000 UK Biobank participants. bioRxiv 2017, 166298. [Google Scholar]
- UK Biobank. Genotyping and Quality Control of UK Biobank, a Large-Scale, Extensively Phenotyped Prospective Resource; UK Biobank: Stockport, UK, 2018. [Google Scholar]
- Marchini, J. UK Biobank Phasing and Imputation Documentation; UK Biobank: Stockport, UK, 2018. [Google Scholar]
- Abdullah Said, M.; Eppinga, R.N.; Lipsic, E.; Verweij, N.; van der Harst, P. Relationship of arterial stiffness index and pulse pressure with cardiovascular disease and mortality. J. Am. Heart Assoc. 2018, 7, e007621. [Google Scholar]
- Bowden, J.; Del Greco, M.F.; Minelli, C.; Davey Smith, G.; Sheehan, N.; Thompson, J. A framework for the investigation of pleiotropy in two-sample summary data Mendelian randomization. Stat. Med. 2017, 36, 1783–1802. [Google Scholar] [CrossRef] [PubMed]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.P.; Turner, J.E. Debunking the Myth of Exercise-Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health across the Lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Effects of stress on immune function: The good, the bad, and the beautiful. Immunol. Res. 2014, 58, 193–210. [Google Scholar] [CrossRef]
- Libby, P.; Theroux, P. Pathophysiology of coronary artery disease. Circulation 2005, 111, 3481–3488. [Google Scholar] [CrossRef]
- Rana, J.S.; Boekholdt, S.M.; Ridker, P.M.; Jukema, J.W.; Luben, R.; Bingham, S.A.; Day, N.E.; Wareham, N.J.; Kastelein, J.J.P.; Khaw, K.-T. Differential leucocyte count and the risk of future coronary artery disease in healthy men and women: The EPIC-Norfolk Prospective Population Study. J. Intern. Med. 2007, 262, 678–689. [Google Scholar] [CrossRef]
- McKune, A.J.; Smith, L.L.; Semple, S.J.; Wadee, A.A. Non-allergic activation of eosinophils after strenuous endurance exercise. S. Afr. J. Sport. Med. 2004, 16, 12–16. [Google Scholar] [CrossRef]
- Gao, S.; Deng, Y.; Wu, J.; Zhang, L.; Deng, F.; Zhou, J.; Yuan, Z.; Wang, L. Eosinophils count in peripheral circulation is associated with coronary artery disease. Atherosclerosis 2019, 286, 128–134. [Google Scholar] [CrossRef]
- Sakai, T.; Inoue, S.; Matsuyama, T.-A.; Takei, M.; Ota, H.; Katagiri, T.; Koboyashi, Y. Eosinophils may be involved in thrombus growth in acute coronary syndrome. Int. Heart J. 2009, 50, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wang, X.; Shen, L.; Yao, K.; Ge, L.; Ma, J.; Zhang, F.; Qian, J.; Ge, J. Association of eosinophil-to-monocyte ratio with 1-month and long-term all-cause mortality in patients with ST-elevation myocardial infarction undergoing primary percutaneous coronary intervention. J. Thorac. Dis. 2018, 10, 5449–5458. [Google Scholar] [CrossRef] [PubMed]
- Loprinzi, P.D.; Hall, M.E. Physical activity and dietary behavior with red blood cell distribution width. Physiol. Behav. 2015, 149, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Hammam, N.; Ezeugwu, V.E.; Manns, P.J.; Pritchard-Wiart, L. Relationships between sedentary behaviour, physical activity levels and red blood cell distribution width in children and adolescents. Heal. Promot. Perspect. 2018, 8, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Loprinzi, P.D.; Ford, E.S. The Association between Objectively Measured Sedentary Behavior and Red Blood Cell Distribution Width in a National Sample of US Adults. Am. J. Epidemiol. 2015, 181, 357–359. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef]
- Keyes, K.M.; Westreich, D. UK Biobank, big data, and the consequences of non-representativeness. Lancet 2019, 393, 1297. [Google Scholar] [CrossRef]
- Kichaev, G.; Bhatia, G.; Loh, P.-R.; Gazal, S.; Burch, K.; Freund, M.K.; Schoech, A.; Pasaniuc, B.; Price, A.L. Leveraging Polygenic Functional Enrichment to Improve GWAS Power. Am. J. Hum. Genet. 2019, 104, 65–75. [Google Scholar] [CrossRef]
- Ferrari, R.; Wang, Y.; Vandrovcova, J.; Guelfi, S.; Witeolar, A.; Karch, C.M.; Schork, A.J.; Fan, C.C.; Brewer, J.B.; International FTD-Genomics Consortium (IFGC); et al. Genetic architecture of sporadic frontotemporal dementia and overlap with Alzheimer’s and Parkinson’s diseases. J. Neurol. Neurosurg. Psychiatry 2017, 88, 152–164. [Google Scholar] [CrossRef]
- Hubel, C.; Gaspar, H.A.; Coleman, J.R.I.; Finucane, H.; Purves, K.L.; Hanscombe, K.B.; Prokopenko, I.; Graff, M.; Ngwa, J.S.; Workalemahu, T.; et al. Genomics of body fat percentage may contribute to sex bias in anorexia nervosa. Am. J. Med. Genet. B. Neuropsychiatr. Genet. 2019, 180, 428–438. [Google Scholar] [CrossRef]
- Akiyama, M.; Okada, Y.; Kanai, M.; Takahashi, A.; Momozawa, Y.; Ikeda, M.; Iwata, N.; Ikegawa, S.; Hirata, M.; Matsuda, K.; et al. Genome-wide association study identifies 112 new loci for body mass index in the Japanese population. Nat. Genet. 2017, 49, 1458–1467. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, Y.; Du, C.; Zhao, W.; Zhang, H.; Yang, Y.; Sun, A.; Song, X.; Feng, Z. Pattern Recognition Receptor-Mediated Chronic Inflammation in the Development and Progression of Obesity-Related Metabolic Diseases. Mediators Inflamm. 2019, 2019, 5271295. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. (%) |
|---|---|
| Total, No | 222,645 |
| Age, mean (SD), years | 56 (8) |
| Sex, male (%) | 105,970 (47.6%) |
| Body Mass Index, mean (SD), kg/m2 | 27.0 (4.4) |
| Smoking behavior, No (%) | |
| Never or <100 cigarettes | 125,777 (58.1%) |
| Ex-smokers | 66,895 (30.9%) |
| Current | 23,849 (11.0%) |
| Hypertension, No (%) | 60,900 (27.4%) |
| Hyperlipidemia, No (%) | 40,099 (18.0%) |
| Diabetes Mellitus type 2, No (%) | 6,984 (3.1%) |
| PA phenotypes | |
| Moderate PA, median (IQR), h/week | 4.9 (1.5–11.3) |
| Vigorous PA, median (IQR), h/week | 0.8 (0.0–2.7) |
| Blood cell counts | |
| Leukocytes, median (IQR), 109/L | 6.57 (5.60–7.70) |
| Erythrocytes, median (IQR), 1012/L | 4.54 (4.29–4.82) |
| Neutrophils, median (IQR), 109/L | 3.95 (3.20–4.82) |
| Lymphocytes, median (IQR), 109/L | 0.92 (0.60–1.20) |
| Monocytes, median (IQR), 109/L | 0.44 (0.36–0.55) |
| Eosinophils, median (IQR), 109/L | 0.13 (0.10–0.21) |
| Basophils, median (IQR), 109/L | 0.02 (0.00–0.04) |
| Reticulocytes, median (IQR), 1012/L | 0.06 (0.04–0.07) |
| Thrombocytes, median (IQR), 109/L | 246.8 (212.9–285.0) |
| N snps | Inverse Variance Weighted (Fixed Effects) | Inverse Variance Weighted (Multiplicative Random Effects) | MR Egger Fixed Effects | Weighted Median | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beta | SE | p-Value | Beta | SE | p-Value | Beta | SE | p-Value | Beta | SE | p-Value | ||
| Leukocyte count (10^9 cells/L) | 68 | –0.0406 | 0.0307 | 1.86 × 10−1 | –0.0406 | 0.0465 | 3.83 × 10−1 | 0.0106 | 0.1455 | 9.42 × 10−1 | 0.0024 | 0.0487 | 9.61 × 10−1 |
| Erythrocyte count (10^12 cells/L) | 68 | 0.0001 | 0.0063 | 9.85 × 10−1 | 0.0001 | 0.0137 | 9.93 × 10−1 | 0.0811 | 0.0417 | 5.64 × 10−2 | −0.0126 | 0.0098 | 1.99 × 10−1 |
| Erythrocyte distribution width (%) | 68 | 0.0078 | 0.0166 | 6.40 × 10−1 | 0.0078 | 0.0281 | 7.82 × 10−1 | 0.0562 | 0.0878 | 5.25 × 10−1 | −0.0047 | 0.0251 | 8.53 × 10−1 |
| Neutrophil count (10^9 cells/L) | 68 | 0.0055 | 0.0243 | 8.20 × 10−1 | 0.0055 | 0.0357 | 8.77 × 10−1 | 0.0768 | 0.1118 | 4.95 × 10−1 | −0.0099 | 0.0369 | 7.88 × 10−1 |
| Neutrophils (%) | 68 | 0.4236 | 0.1502 | 4.80 × 10−3 | 0.4236 | 0.2288 | 6.41 × 10−2 | 0.8990 | 0.7164 | 2.14 × 10−1 | 0.0012 | 0.2359 | 9.96 × 10−1 |
| Lymphocyte count (10^9 cells/L) | 68 | –0.0259 | 0.0081 | 1.40 × 10−3 | –0.0259 | 0.0144 | 7.21 × 10−2 | –0.0317 | 0.0454 | 4.87 × 10−1 | −0.0305 | 0.0134 | 2.24 × 10-2 |
| Lymphocyte (%) | 68 | –0.3606 | 0.1294 | 5.30 × 10−3 | –0.3606 | 0.2016 | 7.37 × 10−2 | –0.8313 | 0.6300 | 1.92 × 10−1 | 0.0836 | 0.1995 | 6.75 × 10−1 |
| Monocyte count (10^9 cells/L) | 68 | –0.0010 | 0.0035 | 7.66 × 10−1 | –0.0010 | 0.0045 | 8.21 × 10−1 | 0.0057 | 0.0140 | 6.86 × 10−1 | −0.0013 | 0.0052 | 8.09 × 10−1 |
| Monocytes (%) | 68 | 0.0158 | 0.0462 | 7.33 × 10−1 | 0.0158 | 0.0613 | 7.97 × 10−1 | 0.0549 | 0.1919 | 7.76 × 10−1 | 0.0116 | 0.0739 | 8.75 × 10−1 |
| Eosinophil count (10^9 cells/L) | 68 | –0.0078 | 0.0024 | 1.40 × 10−3 | –0.0078 | 0.0039 | 4.90 × 10−2 | –0.0071 | 0.0123 | 5.67 × 10−1 | −0.0036 | 0.0036 | 3.23 × 10−1 |
| Eosinophils (%) | 68 | –0.0687 | 0.0340 | 4.36 × 10−2 | –0.0687 | 0.0619 | 2.67 × 10−1 | –0.1215 | 0.1946 | 5.35 × 10−1 | 0.0021 | 0.0534 | 9.69 × 10−1 |
| Basophil count (10^9 cells/L) | 68 | –0.0010 | 0.0009 | 2.89 × 10−1 | –0.0010 | 0.0009 | 2.71 × 10−1 | –0.0006 | 0.0028 | 8.32 × 10−1 | −0.0027 | 0.0013 | 4.54 × 10−2 |
| Basophils (%) | 68 | –0.0141 | 0.0110 | 1.99 × 10−1 | –0.0141 | 0.0110 | 1.99 × 10−1 | –0.0131 | 0.0332 | 6.94 × 10−1 | −0.0262 | 0.0161 | 1.04 × 10−1 |
| Reticulocyte count (10^12 cells/L) | 68 | –0.0256 | 0.0111 | 2.04 × 10−2 | –0.0256 | 0.0188 | 1.73 × 10−1 | 0.0260 | 0.0586 | 6.59 × 10−1 | −0.0455 | 0.0173 | 8.40 × 10−3 |
| Reticulocytes (%) | 68 | –0.0242 | 0.0108 | 2.50 × 10−2 | –0.0242 | 0.0165 | 1.41 × 10−1 | –0.0001 | 0.0517 | 9.99 × 10−1 | −0.0479 | 0.0161 | 3.00 × 10−3 |
| Reticulocyte volume (femtolitres) | 68 | –0.3151 | 0.1397 | 2.41 × 10−2 | –0.3151 | 0.2491 | 2.06 × 10−1 | –0.9755 | 0.7778 | 2.14 × 10−1 | 0.0022 | 0.2097 | 9.92 × 10−1 |
| Immature reticuloyctes fraction | 68 | –0.0027 | 0.0011 | 1.42 × 10-2 | –0.0027 | 0.0016 | 1.01 × 10−1 | 0.0026 | 0.0051 | 6.15 × 10−1 | −0.0055 | 0.0017 | 1.30 × 10−3 |
| Platelet count (10^9 cells/L) | 68 | –2.5671 | 1.0319 | 1.29 × 10−2 | –2.5671 | 1.8209 | 1.59 × 10−1 | 2.7242 | 5.6655 | 6.32 × 10−1 | 1.4266 | 1.6611 | 3.90 × 10−1 |
| Platelet volume (femtolitres) | 68 | –0.0001 | 0.0199 | 9.96 × 10−1 | –0.0001 | 0.0352 | 9.98 × 10−1 | –0.1221 | 0.1092 | 2.68 × 10−1 | 0.0083 | 0.0334 | 8.04 × 10−1 |
| Platelet packed cell volume (%) | 68 | –0.0025 | 0.0008 | 2.50 × 10−3 | –0.0025 | 0.0014 | 6.65 × 10−2 | –0.0003 | 0.0043 | 9.41 × 10−1 | 0.0000 | 0.0013 | 9.76 × 10−1 |
| Platelet distribution width (%) | 68 | 0.0369 | 0.0094 | 1.00 × 10−4 | 0.0369 | 0.0146 | 1.14 × 10−2 | –0.0499 | 0.0443 | 2.64 × 10−1 | 0.0106 | 0.0144 | 4.64 × 10−1 |
| Hemoglobin (g/dL) | 68 | 0.0009 | 0.0177 | 9.60 × 10−1 | 0.0009 | 0.0315 | 9.78 × 10−1 | 0.1597 | 0.0974 | 1.06 × 10−1 | −0.0216 | 0.0274 | 4.30 × 10−1 |
| Hematocrit (%) | 68 | –0.0500 | 0.0521 | 3.37 × 10−1 | –0.0500 | 0.0948 | 5.98 × 10−1 | 0.5543 | 0.2887 | 5.92 × 10−2 | −0.0150 | 0.0816 | 8.54 × 10−1 |
| Mean corpuscular volume (femtoliters) | 68 | –0.1090 | 0.0783 | 1.64 × 10−1 | –0.1090 | 0.1126 | 3.33 × 10−1 | –0.3913 | 0.3527 | 2.71 × 10−1 | 0.0492 | 0.1203 | 6.83 × 10−1 |
| Mean corpuscular hemoglobin (picograms) | 68 | –0.0078 | 0.0321 | 8.08 × 10−1 | –0.0078 | 0.0447 | 8.61 × 10−1 | –0.2389 | 0.1367 | 8.52 × 10−2 | 0.0500 | 0.0488 | 3.05 × 10−1 |
| Mean corpuscular hemoglobin concentration (grams/dL) | 68 | 0.0402 | 0.0187 | 3.19 × 10−2 | 0.0402 | 0.0237 | 8.90 × 10−2 | –0.0970 | 0.0719 | 1.82 × 10−1 | 0.0304 | 0.0284 | 2.84 × 10−1 |
| Mean sphered cells volume (femtoliters) | 68 | –0.0564 | 0.0955 | 5.55 × 10−1 | –0.0564 | 0.1836 | 7.59 × 10−1 | –0.5939 | 0.5733 | 3.04 × 10−1 | 0.2760 | 0.1451 | 5.72 × 10−2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prins, F.M.; Said, M.A.; van de Vegte, Y.J.; Verweij, N.; Groot, H.E.; van der Harst, P. Genetically Determined Physical Activity and Its Association with Circulating Blood Cells. Genes 2019, 10, 908. https://doi.org/10.3390/genes10110908
Prins FM, Said MA, van de Vegte YJ, Verweij N, Groot HE, van der Harst P. Genetically Determined Physical Activity and Its Association with Circulating Blood Cells. Genes. 2019; 10(11):908. https://doi.org/10.3390/genes10110908
Chicago/Turabian StylePrins, Femke M., M. Abdullah Said, Yordi J. van de Vegte, Niek Verweij, Hilde E. Groot, and Pim van der Harst. 2019. "Genetically Determined Physical Activity and Its Association with Circulating Blood Cells" Genes 10, no. 11: 908. https://doi.org/10.3390/genes10110908
APA StylePrins, F. M., Said, M. A., van de Vegte, Y. J., Verweij, N., Groot, H. E., & van der Harst, P. (2019). Genetically Determined Physical Activity and Its Association with Circulating Blood Cells. Genes, 10(11), 908. https://doi.org/10.3390/genes10110908