Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies

 ,
, 
Abstract
1. Introduction
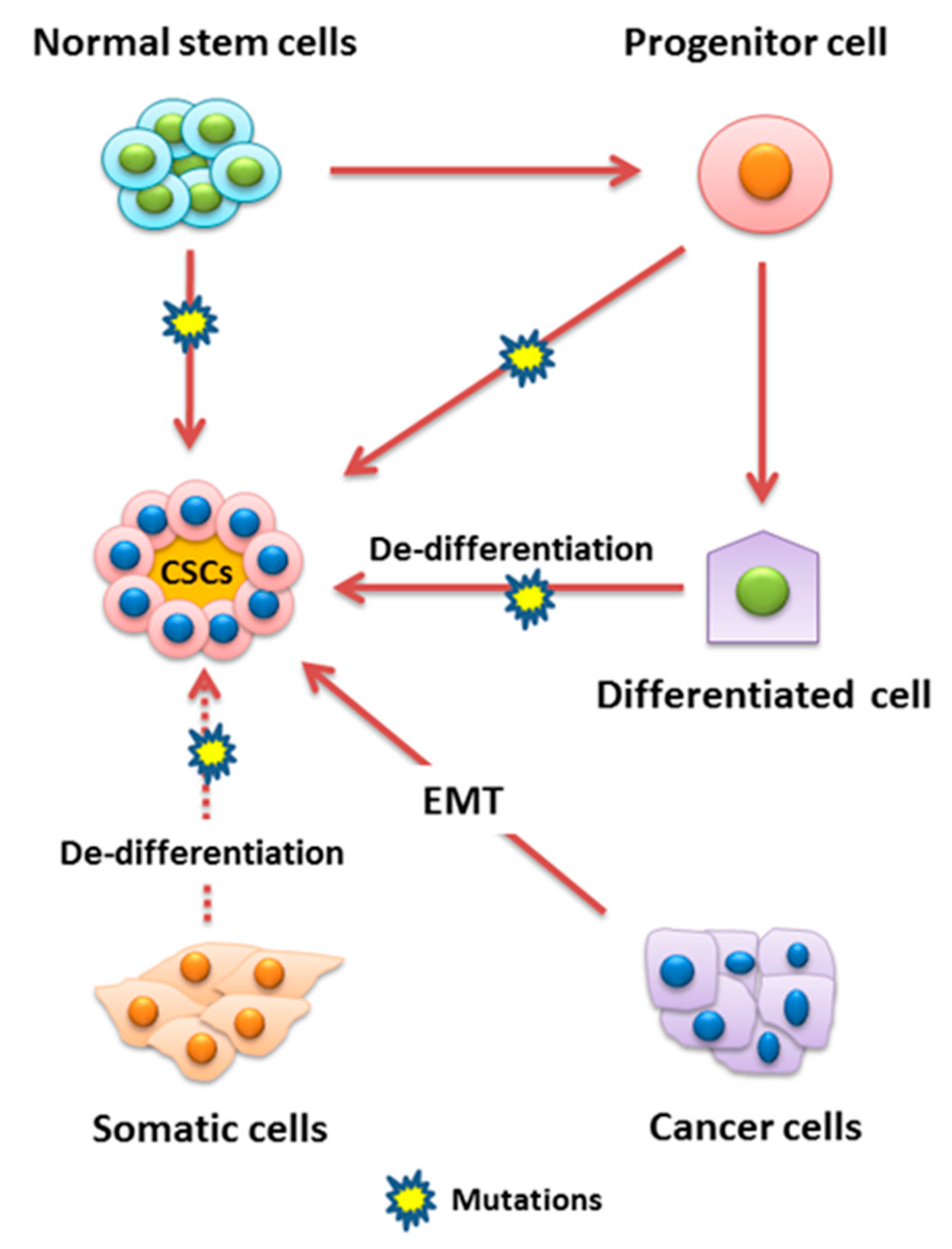
2. Normal and Cancer Stem Cells
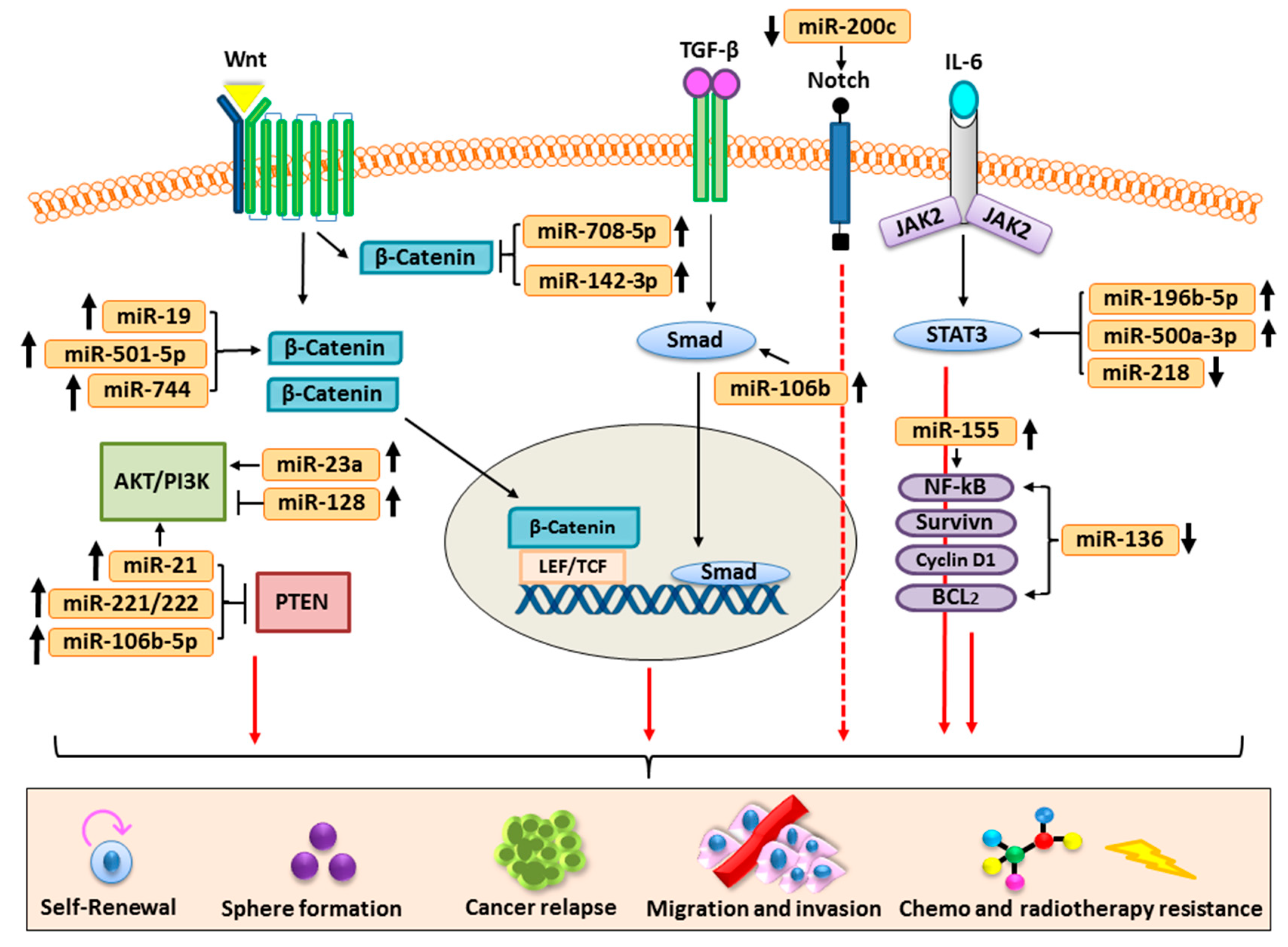
3. Cancer Stem Cells and Signaling Pathways
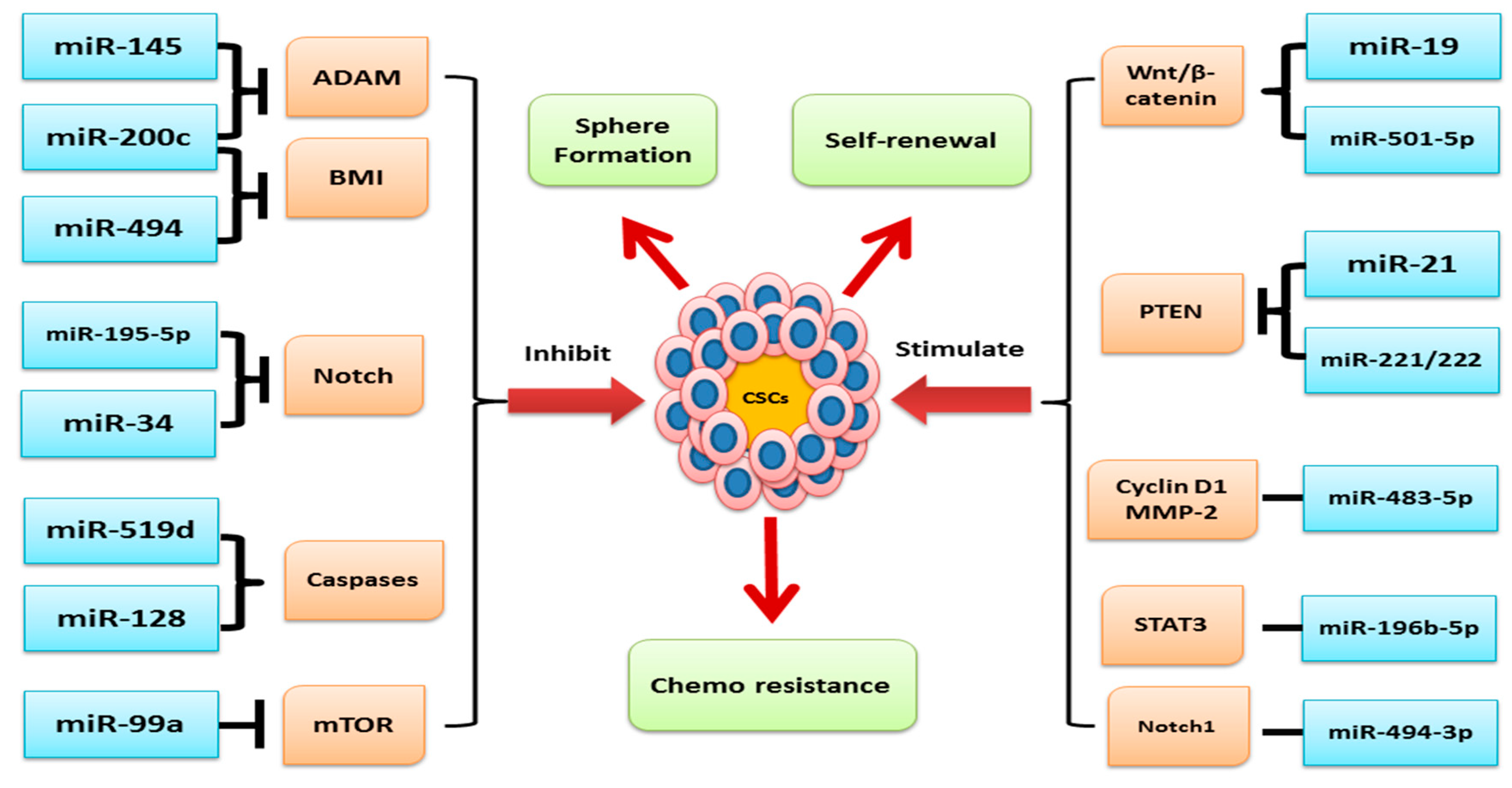
4. Role of miRNA and Oncogenic Signaling Pathways in Human Cancer Stem Cells
4.1. Colorectal Cancer
4.2. Lung Cancer
4.3. Breast Cancer
4.4. Gastric Cancer
4.5. Prostate Cancer
4.6. Pancreatic Cancer
4.7. Hepatocellular Carcinoma (HCC)
4.8. Head and Neck
4.9. Other Human Malignancies
5. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| microRNAs | miRNAs |
| Cancer stem cells | CSCs |
| Prostate cancer | PCa |
| colorectal cancer | CRC |
| untranslated region | UTR |
| 5-fluorouracil | FU |
| tumor microenvironment | TME |
| epithelial to mesenchymal transition | EMT |
| transforming growth factor β | TGFβ |
| head and neck cancer | HNSC |
| aldehyde dehydrogenase1 | ALDH1 |
| Janus-activated kinase/signal transducer and activator of transcription | JAK/STAT |
| phosphatidylinositol 3-kinase/phosphatase and tensin homolog | PI3K/PTEN |
| nuclear factor-kB | NF- kB |
| triple negative breast cancer | TNBC |
| gastric cancer | GC |
References
- Nassar, D.; Blanpain, C. Cancer stem cells: Basic concepts and therapeutic implications. Annu. Rev. Pathol. 2016, 11, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Long, B.; Han, W.; Yuan, S.; Wang, K. Micrornas: Important regulators of stem cells. Stem Cell Res. Ther. 2017, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Pestell, T.G.; Lisanti, M.P.; Pestell, R.G. Cancer stem cells. Int. J. Biochem. Cell Boil. 2012, 44, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Asadzadeh, Z.; Mansoori, B.; Mohammadi, A.; Aghajani, M.; Haji-Asgarzadeh, K.; Safarzadeh, E.; Mokhtarzadeh, A.; Duijf, P.H.G.; Baradaran, B. Micrornas in cancer stem cells: Biology, pathways, and therapeutic opportunities. J. Cell Physiol. 2019, 234, 10002–10017. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.W.; Earle, C.; Shiina, M. Activation of matrix hyaluronan-mediated cd44 signaling, epigenetic regulation and chemoresistance in head and neck cancer stem cells. Int. J. Mol. Sci. 2017, 18, 1849. [Google Scholar] [CrossRef] [PubMed]
- Vlashi, E.; Pajonk, F. Cancer stem cells, cancer cell plasticity and radiation therapy. Semin. Cancer Boil. 2015, 31, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Zhao, S.; Zhang, Y.; Zhao, Z.; Ye, R.; Wen, J.; Li, J. Emerging role of micrornas in cancer and cancer stem cells. J. Cell. Biochem. 2014, 115, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Papaccio, F.; Paino, F.; Regad, T.; Papaccio, G.; Desiderio, V.; Tirino, V. Concise review: Cancer cells, cancer stem cells, and mesenchymal stem cells: Influence in cancer development. Stem Cells Transl. Med. 2017, 6, 2115–2125. [Google Scholar] [CrossRef]
- Chakraborty, C.; Chin, K.Y.; Das, S. Mirna-regulated cancer stem cells: Understanding the property and the role of mirna in carcinogenesis. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 13039–13048. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. The migration ability of stem cells can explain the existence of cancer of unknown primary site. Rethinking metastasis. Oncoscience 2015, 2, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Stem cell division theory of cancer. Cell Cycle 2015, 14, 2547–2548. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Krishnamurthy, S.; Nor, J.E. Head and neck cancer stem cells. J. Dent. Res. 2012, 91, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Kusoglu, A.; Biray Avci, C. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Moharil, R.B.; Dive, A.; Khandekar, S.; Bodhade, A. Cancer stem cells: An insight. J. Oral Maxillofac. Pathol. 2017, 21, 463. [Google Scholar] [CrossRef] [PubMed]
- Matsui, W.H. Cancer stem cell signaling pathways. Medicine 2016, 95, S8–S19. [Google Scholar] [CrossRef] [PubMed]
- Garg, M. Emerging role of micrornas in cancer stem cells: Implications in cancer therapy. World J. Stem Cells 2015, 7, 1078–1089. [Google Scholar] [CrossRef]
- Koury, J.; Zhong, L.; Hao, J. Targeting signaling pathways in cancer stem cells for cancer treatment. Stem Cells Int. 2017, 2017, 2925869. [Google Scholar] [CrossRef]
- Borah, A.; Raveendran, S.; Rochani, A.; Maekawa, T.; Kumar, D.S. Targeting self-renewal pathways in cancer stem cells: Clinical implications for cancer therapy. Oncogenesis 2015, 4, e177. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/beta-catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.K.; Shao, C.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/beta-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, X.; Zhang, H.; Wang, Z.; Nan, G.; Li, Y.; Zhang, F.; Mohammed, M.K.; Haydon, R.C.; Luu, H.H.; et al. The evolving roles of canonical wnt signaling in stem cells and tumorigenesis: Implications in targeted cancer therapies. Lab. Investig. J. Tech. Methods Pathol. 2016, 96, 116–136. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, C.R.; Szczepny, A.; Watkins, D.N.; Cain, J.E. Hedgehog signaling in the maintenance of cancer stem cells. Cancers 2015, 7, 1554–1585. [Google Scholar] [CrossRef]
- Fernandez-Valdivia, R.; Takeuchi, H.; Samarghandi, A.; Lopez, M.; Leonardi, J.; Haltiwanger, R.S.; Jafar-Nejad, H. Regulation of mammalian notch signaling and embryonic development by the protein o-glucosyltransferase rumi. Development 2011, 138, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Schumacher, N.; Maier, M.; Sendtner, M.; Gessler, M. The notch target genes hey1 and hey2 are required for embryonic vascular development. Genes Dev. 2004, 18, 901–911. [Google Scholar] [CrossRef]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef]
- Xiao, W.; Gao, Z.; Duan, Y.; Yuan, W.; Ke, Y. Notch signaling plays a crucial role in cancer stem-like cells maintaining stemness and mediating chemotaxis in renal cell carcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 41. [Google Scholar] [CrossRef]
- Galoczova, M.; Coates, P.; Vojtesek, B. Stat3, stem cells, cancer stem cells and p63. Cell. Mol. Biol. Lett. 2018, 23, 12. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. Nf-kappab signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Cildir, G.; Low, K.C.; Tergaonkar, V. Noncanonical nf-kappab signaling in health and disease. Trends Mol. Med. 2016, 22, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Shen, S.; Verma, I.M. Nf-kappab, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Smahi, A.; Courtois, G.; Rabia, S.H.; Doffinger, R.; Bodemer, C.; Munnich, A.; Casanova, J.L.; Israel, A. The nf-kappab signalling pathway in human diseases: From incontinentia pigmenti to ectodermal dysplasias and immune-deficiency syndromes. Hum. Mol. Genet. 2002, 11, 2371–2375. [Google Scholar] [CrossRef]
- Rinkenbaugh, A.L.; Baldwin, A.S. The nf-kappab pathway and cancer stem cells. Cells 2016, 5, 16. [Google Scholar] [CrossRef]
- Iorio, M.V.; Croce, C.M. Microrna dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microrna function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Reddy, K.B. Microrna (mirna) in cancer. Cancer Cell Int. 2015, 15, 38. [Google Scholar] [CrossRef]
- Takahashi, R.U.; Miyazaki, H.; Ochiya, T. The role of micrornas in the regulation of cancer stem cells. Front. Genet. 2014, 4, 295. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. Micrornas: Small rnas with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Baek, D.; Villen, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of micrornas on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bushati, N.; Cohen, S.M. Microrna functions. Annu. Rev. Cell Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef] [PubMed]
- Yair, E.; Gath, I. High resolution pole-zero analysis of parkinsonian speech. IEEE Trans. Biomed. Eng. 1991, 38, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. Micrornas in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.S.; Malakar, A.K.; Chakraborty, S. Interplay between mirnas and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Croce, C.M. Microrna signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Ors-Kumoglu, G.; Gulce-Iz, S.; Biray-Avci, C. Therapeutic micrornas in human cancer. Cytotechnology 2019, 71, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Vannini, I.; Fanini, F.; Fabbri, M. Emerging roles of micrornas in cancer. Curr. Opin. Genet. Dev. 2018, 48, 128–133. [Google Scholar] [CrossRef]
- Prokopi, M.; Kousparou, C.A.; Epenetos, A.A. The secret role of micrornas in cancer stem cell development and potential therapy: A notch-pathway approach. Front. Oncol. 2014, 4, 389. [Google Scholar] [CrossRef]
- Lou, W.; Liu, J.; Gao, Y.; Zhong, G.; Ding, B.; Xu, L.; Fan, W. Microrna regulation of liver cancer stem cells. Am. J. Cancer Res. 2018, 8, 1126–1141. [Google Scholar]
- Mahabadi, J.A.; Sabzalipoor, H.; Nikzad, H.; Seyedhosseini, E.; Enderami, S.E.; Gheibi Hayat, S.M.; Sahebkar, A. The role of micrornas in embryonic stem cell and induced pluripotent stem cell differentiation in male germ cells. J. Cell. Physiol. 2018, 234, 12278–12289. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Okada, F.; Ochiya, T. Mirna therapy targeting cancer stem cells: A new paradigm for cancer treatment and prevention of tumor recurrence. Ther. Deliv. 2015, 6, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Medvid, R.; Melton, C.; Jaenisch, R.; Blelloch, R. Dgcr8 is essential for microrna biogenesis and silencing of embryonic stem cell self-renewal. Nat. Genet. 2007, 39, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Leichter, A.L.; Sullivan, M.J.; Eccles, M.R.; Chatterjee, A. Microrna expression patterns and signalling pathways in the development and progression of childhood solid tumours. Mol. Cancer 2017, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Micrornas, signaling pathways and diseases. Ann. Transl. Med. 2015, 3, 329. [Google Scholar]
- Mamoori, A.; Gopalan, V.; Smith, R.A.; Lam, A.K. Modulatory roles of micrornas in the regulation of different signalling pathways in large bowel cancer stem cells. Biol. Cell 2016, 108, 51–64. [Google Scholar] [CrossRef]
- Ju, J. Implications of mirnas in colorectal cancer chemoresistance. Int. Drug Discov. 2011, 2011, 2063. [Google Scholar]
- Shi, L.; Xi, J.; Xu, X.; Peng, B.; Zhang, B. Mir-148a suppressed cell invasion and migration via targeting wnt10b and modulating beta-catenin signaling in cisplatin-resistant colorectal cancer cells. Biomed. Pharmacother. 2019, 109, 902–909. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, D.; Kuai, J.; Cheng, M.; Fang, X.; Li, G. Upregulation of mir-199a/b contributes to cisplatin resistance via wnt/beta-catenin-abcg2 signaling pathway in aldha1(+) colorectal cancer stem cells. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2017, 39. [Google Scholar] [CrossRef]
- Ren, D.; Lin, B.; Zhang, X.; Peng, Y.; Ye, Z.; Ma, Y.; Liang, Y.; Cao, L.; Li, X.; Li, R.; et al. Maintenance of cancer stemness by mir-196b-5p contributes to chemoresistance of colorectal cancer cells via activating stat3 signaling pathway. Oncotarget 2017, 8, 49807–49823. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, M.; Mazza, T.; Fusilli, C.; Loiacono, L.; Poeta, M.L.; Sanchez, M.; Massi, E.; Lamorte, G.; Diodoro, M.G.; Pescarmona, E.; et al. Ephb2 stem-related and epha2 progression-related mirna-based networks in progressive stages of crc evolution: Clinical significance and potential mirna drivers. Mol. Cancer 2018, 17, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, J.; Zhao, J.; Bai, J. Knockdown of mir-27a sensitizes colorectal cancer stem cells to trail by promoting the formation of apaf-1-caspase-9 complex. Oncotarget 2017, 8, 45213–45223. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Yu, P.; Li, B.; Guo, Y.H.; Liang, Z.R.; Zheng, L.L.; Yang, J.H.; Xu, H.; Liu, S.; Zheng, L.S.; et al. Mir-372 and mir-373 enhance the stemness of colorectal cancer cells by repressing differentiation signaling pathways. Mol. Oncol. 2018, 12, 1949–1964. [Google Scholar] [CrossRef]
- Xu, P.; Wang, J.; Sun, B.; Xiao, Z. Comprehensive analysis of mirnas expression profiles revealed potential key mirna/mrnas regulating colorectal cancer stem cell self-renewal. Gene 2018, 656, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Hisamori, S.; Oshima, N.; Sato, F.; Shimono, Y.; Sakai, Y. Mir-137 regulates the tumorigenicity of colon cancer stem cells through the inhibition of dclk1. Mol. Cancer Res. 2016, 14, 354–362. [Google Scholar] [CrossRef]
- Wellner, U.; Schubert, J.; Burk, U.C.; Schmalhofer, O.; Zhu, F.; Sonntag, A.; Waldvogel, B.; Vannier, C.; Darling, D.; zur Hausen, A.; et al. The emt-activator zeb1 promotes tumorigenicity by repressing stemness-inhibiting micrornas. Nat. Cell Boil. 2009, 11, 1487–1495. [Google Scholar] [CrossRef]
- Jin, Y.; Wang, M.; Hu, H.; Huang, Q.; Chen, Y.; Wang, G. Overcoming stemness and chemoresistance in colorectal cancer through mir-195-5p-modulated inhibition of notch signaling. Int. J. Biol. Macromol. 2018, 117, 445–453. [Google Scholar] [CrossRef]
- Kwon, T.; Chandimali, N.; Huynh, D.L.; Zhang, J.J.; Kim, N.; Bak, Y.; Yoon, D.Y.; Yu, D.Y.; Lee, J.C.; Gera, M.; et al. Brm270 inhibits cancer stem cell maintenance via microrna regulation in chemoresistant a549 lung adenocarcinoma cells. Cell Death Dis. 2018, 9, 244. [Google Scholar] [CrossRef]
- Yang, Y.; Ding, L.; Hu, Q.; Xia, J.; Sun, J.; Wang, X.; Xiong, H.; Gurbani, D.; Li, L.; Liu, Y.; et al. Microrna-218 functions as a tumor suppressor in lung cancer by targeting il-6/stat3 and negatively correlates with poor prognosis. Mol. Cancer 2017, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Feng, X.; Zhou, W.; Wu, Y.; Yang, Y. Mir-128 reverses the gefitinib resistance of the lung cancer stem cells by inhibiting the c-met/pi3k/akt pathway. Oncotarget 2016, 7, 73188–73199. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cheng, Y.; Li, Y.; Jin, Z.; Pan, Y.; Liu, G.; Fu, S.; Zhang, Y.; Feng, K.; Feng, Y. Microrna-128 plays a critical role in human non-small cell lung cancer tumourigenesis, angiogenesis and lymphangiogenesis by directly targeting vascular endothelial growth factor-c. Eur. J. Cancer 2014, 50, 2336–2350. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Meng, Q.; Qiao, W.; Ma, R.; Ju, W.; Hu, J.; Lu, H.; Cui, J.; Jin, Z.; Zhao, Y.; et al. Mir-181b/notch2 overcome chemoresistance by regulating cancer stem cell-like properties in nsclc. Stem Cell Res. Ther. 2018, 9, 327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, T.; Han, L.; Qin, P.; Wu, Z.; Xu, B.; Gao, Q.; Song, Y. Tgfbeta1-induced down-regulation of microrna-138 contributes to epithelial-mesenchymal transition in primary lung cancer cells. Biochem. Biophys. Res. Commun. 2018, 496, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Xie, Y.K.; Wu, Y.; Li, L.L.; Liu, Y.; Miao, X.B.; Liu, Q.Z.; Yao, K.T.; Xiao, G.H. Sulforaphane inhibits cancer stem-like cell properties and cisplatin resistance through mir-214-mediated downregulation of c-myc in non-small cell lung cancer. Oncotarget 2017, 8, 12067–12080. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, X.; Chen, T.; Luo, Z.; Hu, X. Downregulation of dnmt3a by mir-708-5p inhibits lung cancer stem cell-like phenotypes through repressing wnt/beta-catenin signaling. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 1748–1760. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, L.; Ling, C.; Heng, W. Hur facilitates cancer stemness of lung cancer cells via regulating mir-873/cdk3 and mir-125a-3p/cdk3 axis. Biotechnol. Lett. 2018, 40, 623–631. [Google Scholar] [CrossRef]
- Han, Z.; Zhou, X.; Li, S.; Qin, Y.; Chen, Y.; Liu, H. Inhibition of mir-23a increases the sensitivity of lung cancer stem cells to erlotinib through pten/pi3k/akt pathway. Oncol. Rep. 2017, 38, 3064–3070. [Google Scholar] [CrossRef]
- Faversani, A.; Amatori, S.; Augello, C.; Colombo, F.; Porretti, L.; Fanelli, M.; Ferrero, S.; Palleschi, A.; Pelicci, P.G.; Belloni, E.; et al. Mir-494-3p is a novel tumor driver of lung carcinogenesis. Oncotarget 2017, 8, 7231–7247. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, S.; Chen, Y.; Li, X.; Jiang, Y.; Yang, X.; Li, Y.; Wang, X.; Meng, Y.; Zhu, M.; et al. Mir-19 targeting of gsk3beta mediates sulforaphane suppression of lung cancer stem cells. J. Nutr. Biochem. 2017, 44, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbacher, D.; Klec, C.; Pasculli, B.; Cerk, S.; Rinner, B.; Karbiener, M.; Ivan, C.; Barbano, R.; Ling, H.; Wulf-Goldenberg, A.; et al. Mir-1287-5p inhibits triple negative breast cancer growth by interaction with phosphoinositide 3-kinase cb, thereby sensitizing cells for pi3kinase inhibitors. Breast Cancer Res. 2019, 21, 20. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Huang, Y.; Lou, C.; He, Y.; Zhang, Y.; Zhang, Q. Fstl1 enhances chemoresistance and maintains stemness in breast cancer cells via integrin beta3/wnt signaling under mir-137 regulation. Cancer Boil. Ther. 2019, 20, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Luo, N.; Hu, Y.; Li, X.; Zhang, K. Mir-137 suppresses triple-negative breast cancer stemness and tumorigenesis by perturbing bcl11a-dnmt1 interaction. Cell. Physiol. Biochem. 2018, 47, 2147–2158. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, P.; Climent, M.; Panebianco, F.; Tordonato, C.; Santoro, A.; Marzi, M.J.; Pelicci, P.G.; Ventura, A.; Nicassio, F. Dual role for mir-34a in the control of early progenitor proliferation and commitment in the mammary gland and in breast cancer. Oncogene 2019, 38, 360–374. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, B.; Eades, G.; Zhou, Q. Roles of microrna-140 in stem cell-associated early stage breast cancer. World J. Stem Cells 2014, 6, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Mayoral-Varo, V.; Calcabrini, A.; Sanchez-Bailon, M.P.; Martin-Perez, J. Mir205 inhibits stem cell renewal in sum159pt breast cancer cells. PLoS ONE 2017, 12, e0188637. [Google Scholar] [CrossRef]
- Lv, C.; Li, F.; Li, X.; Tian, Y.; Zhang, Y.; Sheng, X.; Song, Y.; Meng, Q.; Yuan, S.; Luan, L.; et al. Mir-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing wnt signaling antagonists. Nat. Commun. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Li, B.; Lu, Y.; Yu, L.; Han, X.; Wang, H.; Mao, J.; Shen, J.; Wang, B.; Tang, J.; Li, C.; et al. Mir-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting pten and sustained akt/nf-kappab/cox-2 activation. Chem. Biol. Interact. 2017, 277, 33–42. [Google Scholar] [CrossRef]
- Xie, Q.; Wang, S.; Zhao, Y.; Zhang, Z.; Qin, C.; Yang, X. Mir-519d impedes cisplatin-resistance in breast cancer stem cells by down-regulating the expression of mcl-1. Oncotarget 2017, 8, 22003–22013. [Google Scholar] [CrossRef]
- Peng, X.; Kang, Q.; Wan, R.; Wang, Z. Mir-26a/hoxc9 dysregulation promotes metastasis and stem cell-like phenotype of gastric cancer. Cell. Physiol. Biochem. 2018, 49, 1659–1676. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Xu, J.; Guan, X.; Zhou, B.; Wei, W.; Deng, R.; Li, D.; Xu, X.; Zhu, H. In vitro and in vivo effects of mirna-19b/20a/92a on gastric cancer stem cells and the related mechanism. Int. J. Med. Sci. 2018, 15, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Ma, L.; Zhu, J. Mir4835p promotes growth, invasion and selfrenewal of gastric cancer stem cells by wnt/betacatenin signaling. Mol. Med. Rep. 2016, 14, 3421–3428. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Shin, H.S.; Lee, Y.S.; Lee, Y.C. Mir-106b modulates cancer stem cell characteristics through tgf-beta/smad signaling in cd44-positive gastric cancer cells. Lab. Investig. J. Tech. Methods Pathol. 2014, 94, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Guo, X.; Zhang, D.; Fan, Y.; Qin, L.; Dong, S.; Zhang, L. Upregulated mir-132 in lgr5(+) gastric cancer stem cell-like cells contributes to cisplatin-resistance via sirt1/creb/abcg2 signaling pathway. Mol. Carcinog. 2017, 56, 2022–2034. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Ren, B.; Yang, X.; Liu, J.; Zhang, Z. Upregulation of mir-501-5p activates the wnt/beta-catenin signaling pathway and enhances stem cell-like phenotype in gastric cancer. J. Exp. Clin. Cancer Res. 2016, 35, 177. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Dai, Y.; Liu, Q.; Ning, S.; Liu, J.; Shen, Z.; Zhu, D.; Jiang, F.; Zhang, J.; et al. Sulforaphane improves chemotherapy efficacy by targeting cancer stem cell-like properties via the miR-124/IL-6R/STAT3 axis. Sci. Rep. 2016, 6, 36796. [Google Scholar] [CrossRef]
- Song, X.L.; Huang, B.; Zhou, B.W.; Wang, C.; Liao, Z.W.; Yu, Y.; Zhao, S.C. Mir-1301-3p promotes prostate cancer stem cell expansion by targeting sfrp1 and gsk3beta. Biomed. Pharmacother. 2018, 99, 369–374. [Google Scholar] [CrossRef]
- Dallavalle, C.; Albino, D.; Civenni, G.; Merulla, J.; Ostano, P.; Mello-Grand, M.; Rossi, S.; Losa, M.; D’Ambrosio, G.; Sessa, F.; et al. Microrna-424 impairs ubiquitination to activate stat3 and promote prostate tumor progression. J. Clin. Investig. 2016, 126, 4585–4602. [Google Scholar] [CrossRef]
- Guo, Y.; Cui, J.; Ji, Z.; Cheng, C.; Zhang, K.; Zhang, C.; Chu, M.; Zhao, Q.; Yu, Z.; Zhang, Y.; et al. Mir-302/367/lats2/yap pathway is essential for prostate tumor-propagating cells and promotes the development of castration resistance. Oncogene 2017, 36, 6336–6347. [Google Scholar] [CrossRef]
- Jeong, J.H.; Park, S.J.; Dickinson, S.I.; Luo, J.L. A constitutive intrinsic inflammatory signaling circuit composed of mir-196b, meis2, ppp3cc, and p65 drives prostate cancer castration resistance. Mol. Cell 2017, 65, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Ngalame, N.N.; Makia, N.L.; Waalkes, M.P.; Tokar, E.J. Mitigation of arsenic-induced acquired cancer phenotype in prostate cancer stem cells by mir-143 restoration. Toxicol. Appl. Pharmacol. 2016, 312, 11–18. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chang, Y.L.; Zhou, P.J.; Wei, L.; Li, W.; Ji, Z.; Fang, Y.X.; Gao, W.Q. Microrna-7 inhibits the stemness of prostate cancer stem-like cells and tumorigenesis by repressing klf4/pi3k/akt/p21 pathway. Oncotarget 2015, 6, 24017–24031. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, S.; Stebbing, J.; Frampton, A.E.; Zagorac, S.; Krell, J.; de Giorgio, A.; Trabulo, S.M.; Nguyen, V.T.M.; Magnani, L.; Feng, H.; et al. Tgf-beta induces mir-100 and mir-125b but blocks let-7a through lin28b controlling pdac progression. Nat. Commun. 2018, 9, 1845. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Lin, C.M.; Huang, Y.J.; Wei, L.; Ting, L.L.; Kuo, C.C.; Hsu, C.; Chiou, J.F.; Wu, A.T.H.; Lee, W.H. Garcinol downregulates notch1 signaling via modulating mir-200c and suppresses oncogenic properties of panc-1 cancer stem-like cells. Biotechnol. Appl. Biochem. 2017, 64, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Du, J.; Liu, Z.; Zhang, D.; Yao, X.; Yang, Y. Mir-6875-3p promotes the proliferation, invasion and metastasis of hepatocellular carcinoma via btg2/fak/akt pathway. J. Exp. Clin. Cancer Res. 2019, 38, 7. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.M.; Bian, X.Y.; Qin, C.D.; Wu, W.Z. Mir-106b-5p promotes stem cell-like properties of hepatocellular carcinoma cells by targeting pten via pi3k/akt pathway. Onco Targets Ther. 2018, 11, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, Q.; Wang, D.; Luo, F.; Liu, X.; Xue, J.; Yang, P.; Xu, H.; Lu, J.; Zhang, A.; et al. Microrna-191, regulated by hif-2alpha, is involved in emt and acquisition of a stem cell-like phenotype in arsenite-transformed human liver epithelial cells. Toxicol. In Vitro 2018, 48, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Yu, M.; Xie, X.; Huang, G.; Peng, Y.; Ren, D.; Lin, M.; Liu, B.; Liu, M.; Wang, W.; et al. Mir-217 targeting dkk1 promotes cancer stem cell properties via activation of the wnt signaling pathway in hepatocellular carcinoma. Oncol. Rep. 2017, 38, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Long, J.; Liu, B.; Xu, M.; Wang, W.; Xie, X.; Wang, X.; Kuang, M. Mir-500a-3p promotes cancer stem cells properties via stat3 pathway in human hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 99. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Huang, J.; Ma, L.; Shan, J.; Shen, J.; Yang, Z.; Liu, L.; Luo, Y.; Yao, C.; Qian, C. Microrna-122 confers sorafenib resistance to hepatocellular carcinoma cells by targeting igf-1r to regulate ras/raf/erk signaling pathways. Cancer Lett. 2016, 371, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, J.; Shao, M.; Cui, B.; Peng, F.; Li, J.; Ran, Y.; Jin, D.; Kong, J.; Chang, J.; et al. Downregulation of mir-218-5p promotes invasion of oral squamous cell carcinoma cells via activation of cd44-rock signaling. Biomed. Pharmacother. 2018, 106, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, L.Y.; Earle, C.; Wong, G.; Spevak, C.C.; Krueger, K. Stem cell marker (nanog) and stat-3 signaling promote microrna-21 expression and chemoresistance in hyaluronan/cd44-activated head and neck squamous cell carcinoma cells. Oncogene 2012, 31, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.C.; Tsai, L.L.; Wang, M.L.; Yu, C.H.; Lo, W.L.; Chang, Y.C.; Chiou, G.Y.; Chou, M.Y.; Chiou, S.H. Mir145 targets the sox9/adam17 axis to inhibit tumor-initiating cells and il-6-mediated paracrine effects in head and neck cancer. Cancer Res. 2013, 73, 3425–3440. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.L.; Yu, C.C.; Chiou, G.Y.; Chen, Y.W.; Huang, P.I.; Chien, C.S.; Tseng, L.M.; Chu, P.Y.; Lu, K.H.; Chang, K.W.; et al. Microrna-200c attenuates tumour growth and metastasis of presumptive head and neck squamous cell carcinoma stem cells. J. Pathol. 2011, 223, 482–495. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Banerjee, A.; Cui, T.; Han, C.; Cai, S.; Liu, L.; Wu, D.; Cui, R.; Li, Z.; Zhang, X.; et al. Inhibition of mir-328-3p impairs cancer stem cell function and prevents metastasis in ovarian cancer. Cancer Res. 2019, 79, 2314–2326. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Dong, Z.; Chen, Y.; Yang, L.; Lai, D. Enrichment of ovarian cancer stem-like cells is associated with epithelial to mesenchymal transition through an mirna-activated akt pathway. Cell Prolif. 2013, 46, 436–446. [Google Scholar] [CrossRef]
- Tung, S.L.; Huang, W.C.; Hsu, F.C.; Yang, Z.P.; Jang, T.H.; Chang, J.W.; Chuang, C.M.; Lai, C.R.; Wang, L.H. Mirna-34c-5p inhibits amphiregulin-induced ovarian cancer stemness and drug resistance via downregulation of the areg-egfr-erk pathway. Oncogenesis 2017, 6, e326. [Google Scholar] [CrossRef]
- Chen, M.W.; Yang, S.T.; Chien, M.H.; Hua, K.T.; Wu, C.J.; Hsiao, S.M.; Lin, H.; Hsiao, M.; Su, J.L.; Wei, L.H. The stat3-mirna-92-wnt signaling pathway regulates spheroid formation and malignant progression in ovarian cancer. Cancer Res. 2017, 77, 1955–1967. [Google Scholar] [CrossRef]
- Wu, G.; Liu, A.; Zhu, J.; Lei, F.; Wu, S.; Zhang, X.; Ye, L.; Cao, L.; He, S. Mir-1207 overexpression promotes cancer stem cell-like traits in ovarian cancer by activating the wnt/beta-catenin signaling pathway. Oncotarget 2015, 6, 28882–28894. [Google Scholar]
- Liu, T.; Qin, W.; Hou, L.; Huang, Y. Microrna-17 promotes normal ovarian cancer cells to cancer stem cells development via suppression of the lkb1-p53-p21/waf1 pathway. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 1881–1893. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Kang, H.; Kim, T.H.; Kim, G.; Heo, J.H.; Kwon, A.Y.; Kim, S.; Jung, S.G.; An, H.J. Microrna-136 inhibits cancer stem cell activity and enhances the anti-tumor effect of paclitaxel against chemoresistant ovarian cancer cells by targeting notch3. Cancer Lett. 2017, 386, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Haghpanah, V.; Fallah, P.; Tavakoli, R.; Naderi, M.; Samimi, H.; Soleimani, M.; Larijani, B. Antisense-mir-21 enhances differentiation/apoptosis and reduces cancer stemness state on anaplastic thyroid cancer. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Chen, Y.; Gong, Y.; Dong, T.; Zhang, B.; Gao, W. Mir-148a inhibits self-renewal of thyroid cancer stem cells via repressing ino80 expression. Oncol. Rep. 2016, 36, 3387–3396. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huang, S.; Tan, X.; Huang, Z.; Chen, Z.; Lin, P.; Fu, S.W. Microrna biomarkers in colorectal cancer liver metastasis. J. Cancer 2018, 9, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Desantis, C.; Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 104–117. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, M.; Poeta, M.L.; Signori, E.; Fazio, V.M. Current understanding and clinical utility of mirnas regulation of colon cancer stem cells. Semin. Cancer Boil. 2018, 53, 232–247. [Google Scholar] [CrossRef]
- Chhabra, R.; Saini, N. Micrornas in cancer stem cells: Current status and future directions. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 8395–8405. [Google Scholar] [CrossRef]
- Hirsch, F.R.; Scagliotti, G.V.; Mulshine, J.L.; Kwon, R.; Curran, W.J., Jr.; Wu, Y.L.; Paz-Ares, L. Lung cancer: Current therapies and new targeted treatments. Lancet 2017, 389, 299–311. [Google Scholar] [CrossRef]
- Castro, D.; Moreira, M.; Gouveia, A.M.; Pozza, D.H.; De Mello, R.A. Micrornas in lung cancer. Oncotarget 2017, 8, 81679–81685. [Google Scholar] [CrossRef]
- Fan, T.; Wang, W.; Zhang, B.; Xu, Y.; Chen, L.; Pan, S.; Hu, H.; Geng, Q. Regulatory mechanisms of micrornas in lung cancer stem cells. SpringerPlus 2016, 5, 1762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Q.; Wang, S. Micrornas: A new key in lung cancer. Cancer Chemother. Pharmacol. 2014, 74, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.C.; Chin, T.M.; Yang, H.; Nga, M.E.; Lunny, D.P.; Lim, E.K.; Sun, L.L.; Pang, Y.H.; Leow, Y.N.; Malusay, S.R.; et al. Tumour-initiating cell-specific mir-1246 and mir-1290 expression converge to promote non-small cell lung cancer progression. Nat. Commun. 2016, 7, 11702. [Google Scholar] [CrossRef] [PubMed]
- Vahidian, F.; Mohammadi, H.; Ali-Hasanzadeh, M.; Derakhshani, A.; Mostaan, M.; Hemmatzadeh, M.; Baradaran, B. Micrornas and breast cancer stem cells: Potential role in breast cancer therapy. J. Cell. Physiol. 2019, 234, 3294–3306. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Shi, W.; Tang, T.; Wang, Y.; Yin, X.; Chen, Y.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. Mir-29a contributes to breast cancer cells epithelial-mesenchymal transition, migration, and invasion via down-regulating histone h4k20 trimethylation through directly targeting suv420h2. Cell Death Dis. 2019, 10, 176. [Google Scholar] [CrossRef]
- Huang, Q.D.; Zheng, S.R.; Cai, Y.J.; Chen, D.L.; Shen, Y.Y.; Lin, C.Q.; Hu, X.Q.; Wang, X.H.; Shi, H.; Guo, G.L. Imp3 promotes tnbc stem cell property through mirna-34a regulation. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2688–2696. [Google Scholar]
- Lin, X.; Chen, W.; Wei, F.; Zhou, B.P.; Hung, M.C.; Xie, X. Nanoparticle delivery of mir-34a eradicates long-term-cultured breast cancer stem cells via targeting c22orf28 directly. Theranostics 2017, 7, 4805–4824. [Google Scholar] [CrossRef]
- Lin, C.; Gao, B.; Yan, X.; Lei, Z.; Chen, K.; Li, Y.; Zeng, Q.; Chen, Z.; Li, H. Microrna 628 suppresses migration and invasion of breast cancer stem cells through targeting sos1. Onco Targets Ther. 2018, 11, 5419–5428. [Google Scholar] [CrossRef]
- Tang, T.; Yang, Z.; Zhu, Q.; Wu, Y.; Sun, K.; Alahdal, M.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. Up-regulation of mir-210 induced by a hypoxic microenvironment promotes breast cancer stem cells metastasis, proliferation, and self-renewal by targeting e-cadherin. FASEB J. 2018, 32, 6965–6981. [Google Scholar] [CrossRef]
- Cheng, C.W.; Yu, J.C.; Hsieh, Y.H.; Liao, W.L.; Shieh, J.C.; Yao, C.C.; Lee, H.J.; Chen, P.M.; Wu, P.E.; Shen, C.Y. Increased cellular levels of microrna-9 and microrna-221 correlate with cancer stemness and predict poor outcome in human breast cancer. Cell. Physiol. Biochem. 2018, 48, 2205–2218. [Google Scholar] [CrossRef]
- Troschel, F.M.; Bohly, N.; Borrmann, K.; Braun, T.; Schwickert, A.; Kiesel, L.; Eich, H.T.; Gotte, M.; Greve, B. Mir-142-3p attenuates breast cancer stem cell characteristics and decreases radioresistance in vitro. Tumour Boil. J. Int. Soc. Oncodev. Biol. Med. 2018, 40. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.Q.; Fu, F.; Han, J. Mir-27a modulates radiosensitivity of triple-negative breast cancer (tnbc) cells by targeting cdc27. Med. Sci. Monit. 2015, 21, 1297–1303. [Google Scholar] [PubMed]
- Wu, D.; Zhang, J.; Lu, Y.; Bo, S.; Li, L.; Wang, L.; Zhang, Q.; Mao, J. Mir-140-5p inhibits the proliferation and enhances the efficacy of doxorubicin to breast cancer stem cells by targeting wnt1. Cancer Gene Ther. 2019, 26, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Li, Y.; Fan, X.; Ma, J.; Li, J.; Lu, G.; Zhang, Y.; Huang, Y.; Li, W.; Huang, X.; et al. Mir-4319 suppress the malignancy of triple-negative breast cancer by regulating self-renewal and tumorigenesis of stem cells. Cell. Physiol. Biochem. 2018, 48, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Zhang, J.; Li, J.; Shao, J.; Fang, L. Mir-130a-3p inhibits migration and invasion by regulating rab5b in human breast cancer stem cell-like cells. Biochem. Biophys. Res. Commun. 2018, 501, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, C.; Xiao, G.; Meng, J.; Wang, J.; Tang, S.C.; Qin, S.; Du, N.; Li, G.; Ren, H.; et al. Breast cancer stem-like cells are sensitized to tamoxifen induction of self-renewal inhibition with enforced let-7c dependent on wnt blocking. Int. J. Mol. Med. 2018, 41, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Yu, Y.; Zhu, M.; Jing, W.; Yu, M.; Chai, H.; Liang, C.; Tu, J. Inhibition of mir-155, a therapeutic target for breast cancer, prevented in cancer stem cell formation. Cancer Biomark. 2018, 21, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Bott, A.; Erdem, N.; Lerrer, S.; Hotz-Wagenblatt, A.; Breunig, C.; Abnaof, K.; Worner, A.; Wilhelm, H.; Munstermann, E.; Ben-Baruch, A.; et al. Mirna-1246 induces pro-inflammatory responses in mesenchymal stem/stromal cells by regulating pka and pp2a. Oncotarget 2017, 8, 43897–43914. [Google Scholar] [CrossRef]
- El Helou, R.; Pinna, G.; Cabaud, O.; Wicinski, J.; Bhajun, R.; Guyon, L.; Rioualen, C.; Finetti, P.; Gros, A.; Mari, B.; et al. Mir-600 acts as a bimodal switch that regulates breast cancer stem cell fate through wnt signaling. Cell Rep. 2017, 18, 2256–2268. [Google Scholar] [CrossRef]
- Fu, H.; Fu, L.; Xie, C.; Zuo, W.S.; Liu, Y.S.; Zheng, M.Z.; Yu, J.M. Mir-375 inhibits cancer stem cell phenotype and tamoxifen resistance by degrading hoxb3 in human er-positive breast cancer. Oncol. Rep. 2017, 37, 1093–1099. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Du, P.; Zhao, J.; Hu, C.; Qin, Y.; Huang, G. Gastric cancer stem cells: Mechanisms and therapeutic approaches. Yonsei Med. J. 2018, 59, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Pinheiro, H.; Figueiredo, J.; Seruca, R.; Carneiro, F. Familial gastric cancer: Genetic susceptibility, pathology, and implications for management. Lancet Oncol. 2015, 16, e60–e70. [Google Scholar] [CrossRef]
- Brungs, D.; Aghmesheh, M.; Vine, K.L.; Becker, T.M.; Carolan, M.G.; Ranson, M. Gastric cancer stem cells: Evidence, potential markers, and clinical implications. J. Gastroenterol. 2016, 51, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Han, T.S.; Voon, D.C.; Oshima, H.; Nakayama, M.; Echizen, K.; Sakai, E.; Yong, Z.W.E.; Murakami, K.; Yu, L.; Minamoto, T.; et al. Interleukin 1 up-regulates microrna 135b to promote inflammation-associated gastric carcinogenesis in mice. Gastroenterology 2019, 156, 1140–1155. [Google Scholar] [CrossRef]
- Gong, Y.C.; Ren, G.L.; Liu, B.; Li, F.; Zhao, H.P.; Chen, J.B.; Li, Y.P.; Yu, H.H. Mir206 inhibits cancer initiating cells by targeting ehf in gastric cancer. Oncol. Rep. 2017, 38, 1688–1694. [Google Scholar] [CrossRef]
- Wu, Q.; Yang, Z.; Wang, F.; Hu, S.; Yang, L.; Shi, Y.; Fan, D. Mir-19b/20a/92a regulates the self-renewal and proliferation of gastric cancer stem cells. J. Cell Sci. 2013, 126, 4220–4229. [Google Scholar] [CrossRef]
- Taitt, H.E. Global trends and prostate cancer: A review of incidence, detection, and mortality as influenced by race, ethnicity, and geographic location. Am. J. Mens Heal. 2018, 12, 1807–1823. [Google Scholar] [CrossRef]
- Leao, R.; Domingos, C.; Figueiredo, A.; Hamilton, R.; Tabori, U.; Castelo-Branco, P. Cancer stem cells in prostate cancer: Implications for targeted therapy. Urol. Int. 2017, 99, 125–136. [Google Scholar] [CrossRef]
- Guan, B.; Mu, L.; Zhang, L.; Wang, K.; Tian, J.; Xu, S.; Wang, X.; He, D.; Du, Y. Microrna-218 inhibits the migration, epithelial-mesenchymal transition and cancer stem cell properties of prostate cancer cells. Oncol. Lett. 2018, 16, 1821–1826. [Google Scholar] [CrossRef]
- Liu, T.; Chi, H.; Chen, J.; Chen, C.; Huang, Y.; Xi, H.; Xue, J.; Si, Y. Curcumin suppresses proliferation and in vitro invasion of human prostate cancer stem cells by cerna effect of mir-145 and lncrna-ror. Gene 2017, 631, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, R.; Zhang, D.; Deng, Q.; Liu, B.; Chao, H.P.; Rycaj, K.; Takata, Y.; Lin, K.; Lu, Y.; et al. Microrna-141 suppresses prostate cancer stem cells and metastasis by targeting a cohort of pro-metastasis genes. Nat. Commun. 2017, 8, 14270. [Google Scholar] [CrossRef] [PubMed]
- Bucay, N.; Sekhon, K.; Yang, T.; Majid, S.; Shahryari, V.; Hsieh, C.; Mitsui, Y.; Deng, G.; Tabatabai, Z.L.; Yamamura, S.; et al. Microrna-383 located in frequently deleted chromosomal locus 8p22 regulates cd44 in prostate cancer. Oncogene 2017, 36, 2667–2679. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, C.; Zhang, D.; Liu, B.; Chen, X.; Rycaj, K.; Jeter, C.; Calhoun-Davis, T.; Li, Y.; Yang, T.; et al. Mir-199a-3p targets stemness-related and mitogenic signaling pathways to suppress the expansion and tumorigenic capabilities of prostate cancer stem cells. Oncotarget 2016, 7, 56628–56642. [Google Scholar] [CrossRef] [PubMed]
- Albino, D.; Civenni, G.; Dallavalle, C.; Roos, M.; Jahns, H.; Curti, L.; Rossi, S.; Pinton, S.; D’Ambrosio, G.; Sessa, F.; et al. Activation of the lin28/let-7 axis by loss of ese3/ehf promotes a tumorigenic and stem-like phenotype in prostate cancer. Cancer Res. 2016, 76, 3629–3643. [Google Scholar] [CrossRef]
- Fan, X.; Chen, X.; Deng, W.; Zhong, G.; Cai, Q.; Lin, T. Up-regulated microrna-143 in cancer stem cells differentiation promotes prostate cancer cells metastasis by modulating fndc3b expression. BMC Cancer 2013, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, I.S.; Chang, K.C.; Tsai, Y.T.; Ke, J.Y.; Lu, P.J.; Lee, K.H.; Yeh, S.D.; Hong, T.M.; Chen, Y.L. Microrna-320 suppresses the stem cell-like characteristics of prostate cancer cells by downregulating the wnt/beta-catenin signaling pathway. Carcinogenesis 2013, 34, 530–538. [Google Scholar] [CrossRef]
- Liu, C.; Kelnar, K.; Liu, B.; Chen, X.; Calhoun-Davis, T.; Li, H.; Patrawala, L.; Yan, H.; Jeter, C.; Honorio, S.; et al. The microrna mir-34a inhibits prostate cancer stem cells and metastasis by directly repressing cd44. Nat. Med. 2011, 17, 211–215. [Google Scholar] [CrossRef]
- Ercan, G.; Karlitepe, A.; Ozpolat, B. Pancreatic cancer stem cells and therapeutic approaches. Anticancer. Res. 2017, 37, 2761–2775. [Google Scholar]
- Moriya, C.; Taniguchi, H.; Miyata, K.; Nishiyama, N.; Kataoka, K.; Imai, K. Inhibition of prdm14 expression in pancreatic cancer suppresses cancer stem-like properties and liver metastasis in mice. Carcinogenesis 2017, 38, 638–648. [Google Scholar] [CrossRef]
- Bimonte, S.; Barbieri, A.; Leongito, M.; Palma, G.; Del Vecchio, V.; Falco, M.; Palaia, R.; Albino, V.; Piccirillo, M.; Amore, A.; et al. The role of mirnas in the regulation of pancreatic cancer stem cells. Stem Cells Int. 2016, 2016, 8352684. [Google Scholar] [CrossRef] [PubMed]
- Tsukasa, K.; Ding, Q.; Miyazaki, Y.; Matsubara, S.; Natsugoe, S.; Takao, S. Mir-30 family promotes migratory and invasive abilities in cd133(+) pancreatic cancer stem-like cells. Hum. Cell 2016, 29, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, Y.; Gou, S.; Xiong, J.; Wu, H.; Wang, C.; Yan, H.; Liu, T. Mir-744 increases tumorigenicity of pancreatic cancer by activating wnt/beta-catenin pathway. Oncotarget 2015, 6, 37557–37569. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Huang, T.; Ding, Y.C.; Yu, W.; Wang, Q.; Meng, B.; Luo, S.X. Microrna-200c overexpression inhibits chemoresistance, invasion and colony formation of human pancreatic cancer stem cells. Int. J. Clin. Exp. Pathol. 2015, 8, 6533–6539. [Google Scholar] [PubMed]
- Laursen, L. A preventable cancer. Nature 2014, 516, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhu, Y. Targeting liver cancer stem cells for the treatment of hepatocellular carcinoma. Ther. Adv. Gastroenterol. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, X.; Wang, Z.; Cheng, Q.; Ji, G.; Yang, H.; Wan, L.; Ge, C.; Zeng, Q.; Huang, H.; et al. Microrna4865p functions as a tumor suppressor of proliferation and cancer stemlike cell properties by targeting sirt1 in liver cancer. Oncol. Rep. 2019, 41, 1938–1948. [Google Scholar]
- Gu, Y.; Wei, X.; Sun, Y.; Gao, H.; Zheng, X.; Wong, L.L.; Jin, L.; Liu, N.; Hernandez, B.; Peplowska, K.; et al. Mir-192-5p silencing by genetic aberrations is a key event in hepatocellular carcinomas with cancer stem cell features. Cancer Res. 2019, 79, 941–953. [Google Scholar] [CrossRef]
- Chen, C.; Luo, F.; Yang, Q.; Wang, D.; Yang, P.; Xue, J.; Dai, X.; Liu, X.; Xu, H.; Lu, J.; et al. Nf-kappab-regulated mir-155, via repression of qki, contributes to the acquisition of csc-like phenotype during the neoplastic transformation of hepatic cells induced by arsenite. Mol. Carcinog. 2018, 57, 483–493. [Google Scholar] [CrossRef]
- Lu, A.Q.; Lv, B.; Qiu, F.; Wang, X.Y.; Cao, X.H. Upregulation of mir-137 reverses sorafenib resistance and cancer-initiating cell phenotypes by degrading ant2 in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 2071–2078. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, P.; Shuai, L.; Chen, K.; Li, Z.; Zhang, Y.; Jiang, Y.; Li, X. Mir-589-5p inhibits map3k8 and suppresses cd90(+) cancer stem cells in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 176. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Ng, K.Y.; Tong, M.; Lau, E.Y.; Lee, T.K.; Chan, K.W.; Yuan, Y.F.; Cheung, T.T.; Cheung, S.T.; Wang, X.Q.; et al. Octamer 4/microrna-1246 signaling axis drives wnt/beta-catenin activation in liver cancer stem cells. Hepatology 2016, 64, 2062–2076. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, D.L.; Dong, L.L.; Wen, D.; Shi, D.M.; Zhou, J.; Fan, J.; Wu, W.Z. Mir-612 suppresses stem cell-like property of hepatocellular carcinoma cells by modulating sp1/nanog signaling. Cell Death Dis. 2016, 7, e2377. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Liu, J.; Yang, Z.; Wu, L.; Xie, H.; Jiang, C.; Lin, B.; Chen, T.; Xing, C.; Liu, Z.; et al. Microrna-452 promotes stem-like cells of hepatocellular carcinoma by inhibiting sox7 involving wnt/beta-catenin signaling pathway. Oncotarget 2016, 7, 28000–28012. [Google Scholar] [CrossRef] [PubMed]
- Courthod, G.; Franco, P.; Palermo, L.; Pisconti, S.; Numico, G. The role of microrna in head and neck cancer: Current knowledge and perspectives. Molecules 2014, 19, 5704–5716. [Google Scholar] [CrossRef] [PubMed]
- Nagadia, R.; Pandit, P.; Coman, W.B.; Cooper-White, J.; Punyadeera, C. Mirnas in head and neck cancer revisited. Cell. Oncol. 2013, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.F.; Lin, S.C.; Chang, K.W. Microrna aberrances in head and neck cancer: Pathogenetic and clinical significance. Curr. Opin. Otolaryngol. Head Neck Surg. 2013, 21, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Chen, P.Y.; Ho, D.C.; Tsai, L.L.; Hsieh, P.L.; Lu, M.Y.; Yu, C.C.; Yu, C.H. Mir-145 mediates the anti-cancer stemness effect of photodynamic therapy with 5-aminolevulinic acid (ala) in oral cancer cells. J. Formos. Med. Assoc. 2018, 117, 738–742. [Google Scholar] [CrossRef]
- Peng, C.Y.; Wang, T.Y.; Lee, S.S.; Hsieh, P.L.; Liao, Y.W.; Tsai, L.L.; Fang, C.Y.; Yu, C.C.; Hsieh, C.S. Let-7c restores radiosensitivity and chemosensitivity and impairs stemness in oral cancer cells through inhibiting interleukin-8. J. Oral. Pathol. Med. 2018, 47, 590–597. [Google Scholar] [CrossRef]
- Chang, Y.C.; Jan, C.I.; Peng, C.Y.; Lai, Y.C.; Hu, F.W.; Yu, C.C. Activation of microrna-494-targeting bmi1 and adam10 by silibinin ablates cancer stemness and predicts favourable prognostic value in head and neck squamous cell carcinomas. Oncotarget 2015, 6, 24002–24016. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, Y.; Wang, Y.; Zhang, Y.; Luo, Q.; Man, X.; Wei, F.; Yu, X. Downregulation of foxo3 and trim31 by mir-551b in side population promotes cell proliferation, invasion, and drug resistance of ovarian cancer. Med. Oncol. 2016, 33, 126. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.A.; Fridman, M.V.; Kushlinskii, N.E. Molecular mechanisms of ovarian carcinoma metastasis: Key genes and regulatory micrornas. Biochemistry 2017, 82, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Qian, Z.; Ma, X.; Lin, X.; You, Y.; Li, Y.; Chen, T.; Jiang, H. Mir-628-5p decreases the tumorigenicity of epithelial ovarian cancer cells by targeting at fgfr2. Biochem. Biophys. Res. Commun. 2018, 495, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Gao, C.; Wang, K.; Cui, L.; Li, W.; Zhao, X.; Liu, F.; Wu, M.; Deng, G.; Ding, W.; et al. Effect of microrna-210 on prognosis and response to chemotherapeutic drugs in pediatric acute lymphoblastic leukemia. Cancer Sci. 2014, 1056, 43–472. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Sun, S.; Zhang, X.Q.; Zhang, P.D.; Ho, A.S.; Kiang, K.M.; Fung, C.F.; Lui, W.M.; Leung, G.K. Microrna-210 and endoplasmic reticulum chaperones in the regulation of chemoresistance in glioblastoma. J. Cancer 2015, 6, 227–232. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cancer Type | miRNA | Signaling Pathways/Targeting Gene | References |
|---|---|---|---|
| Colorectal | miR-21 | PTEN, AKT, Ras | [58] |
| MiRNA-215 | DTL | [59] | |
| miR-148a | WNT, β-catenin | [60] | |
| miR-199a/b | Gsk3β, Wnt/β-catenin-ABCG2 | [61] | |
| miR-196b-5p | STAT3 | [62] | |
| miRs-31 | EphB2 and EphA2 | [63] | |
| miR-27a | Apaf-1/caspase-9 | [64] | |
| miR-372/373 | Nanog, Hedgehog, NFκB, MAPK/Erk, VDR Jak-STAT, TGF-beta, PI3K-Akt, MAPK. | [65,66] | |
| miR-137 | DCLK1 | [67] | |
| miR-146a | β-catenin | [68] | |
| miR-195-5p | STAT3, BIRC5, BCL2, BCL-XL, SOX2, CD133, RBPJ, Notch2 | [62,69] | |
| Lung | miR-128 | AKT/ERK, p38, c-met/PI3K/AKT, VEGF/PI3K/AKT, IL-6-JAK-STAT3 | [70,71,72,73] |
| miR-181b | Notch2 | [74] | |
| miR-138 | TGFβ | [75] | |
| miR-5100 | Rab6 | [76] | |
| miR-214 | c-MYC | [76] | |
| miR-708-5p | Wnt/β-catenin | [77] | |
| miR-873 miR-125a-3p | Oct4, Nanog and ALDH | [78] | |
| miR-23a | PTEN/PI3K/Akt | [79] | |
| miR-494-3p | NOTCH1-PI3K | [80] | |
| miR-19a/19b | Wnt/β-catenin | [81] | |
| Breast | miR-1287-5p | PI3Kinase | [82] |
| miR-137 | β3/Wnt, BCL11A | [83,84] | |
| miR-34a | Wnt/beta-catenin | [85] | |
| miRNA-140-5p | Wnt/β-catenin | [86] | |
| miR205 | STAT3 | [87] | |
| miR-31 | Prlr/Stat5, TGFβ and Wnt/β-catenin | [88] | |
| miR-221/222 | ALDH1, PTEN, p65, pp65, p-AKT, COX-2 | [89] | |
| miR-519d | Bcl-2, MCL-1 | [90] | |
| Gastric | MiR-26a | HOXC9 | [91] |
| miRNA-19b/20a/92a | β-catenin | [92] | |
| miR-483-5p | Wnt/β-catenin | [93] | |
| miR-106b | TGF-β/Smad | [94] | |
| miR-132 | SIRT1/CREB/ABCG2 | [95] | |
| miR-501-5p | DKK1, NKD1, GSK3β, IL-6R/STAT3 | [96,97] | |
| Prostate | miR-218 | OCT4, SOX2, NANOG, CD44, KLF4, c-MYC, Wnt, | [98] |
| miR-424 | STAT3 | [99] | |
| miR-302/367 | NANOG, SOX2, OCT4, KLF4, BMI-1, LATS2/YAP | [100] | |
| miR-199a-3p | c-MYC, cyclin D1 (CCND1), EGFR | [101] | |
| miR-143 | MMP-9, MMP-2 | [102] | |
| miR-7 | KLF4/PI3K/Akt/p21 | [103] | |
| Pancreatic | miR-100 miR-125b | TGF-β | [104] |
| miR-200c | Notch1 | [105] | |
| Liver | miR-6875-3p | BTG2/FAK/Akt | [106] |
| miR-106b-5p | PI3K/Akt | [107] | |
| miR-191 | HIF-2α | [108] | |
| miR-217 | Wnt | [109] | |
| miR-500a-3p | SOCS2, SOCS4, PTPN11, STAT3 | [110] | |
| miR-122 | RAS/RAF/ERK | [111] | |
| Head and Neck | miR-218-5p | CD44-ROCK | [112] |
| Mir-21 | Nanog-Stat-3 | [113] | |
| miR-145 | ADAM17, SOX9 | [114] | |
| miR-200c | ALDH1, Nanog, Oct4, SOX2. | [115] | |
| Ovarian | miR-328 | ERK | [116] |
| miR-20a miR-200c | PI3K/AKT | [117] | |
| miRNA-34c-5p | EGFR-ERK | [118] | |
| miR-92a | Wnt | [119] | |
| miR-1207 | Wnt/β-catenin | [120] | |
| miR-17 | LKB1-p53-p21/WAF1 | [121] | |
| miR-136 | NOTCH3, NF-kB, Cyclin D1, Survivin, BCL-XL, BCL2 | [122] | |
| Thyroid | miR-21 | ABCG2, Oct4 | [123] |
| miR-148a | ATC-CSCs | [124] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.Q.; Ahmed, E.I.; Elareer, N.R.; Junejo, K.; Steinhoff, M.; Uddin, S. Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells 2019, 8, 840. https://doi.org/10.3390/cells8080840
Khan AQ, Ahmed EI, Elareer NR, Junejo K, Steinhoff M, Uddin S. Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells. 2019; 8(8):840. https://doi.org/10.3390/cells8080840
Chicago/Turabian StyleKhan, Abdul Q., Eiman I. Ahmed, Noor R. Elareer, Kulsoom Junejo, Martin Steinhoff, and Shahab Uddin. 2019. "Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies" Cells 8, no. 8: 840. https://doi.org/10.3390/cells8080840
APA StyleKhan, A. Q., Ahmed, E. I., Elareer, N. R., Junejo, K., Steinhoff, M., & Uddin, S. (2019). Role of miRNA-Regulated Cancer Stem Cells in the Pathogenesis of Human Malignancies. Cells, 8(8), 840. https://doi.org/10.3390/cells8080840