Improved Antitumor Efficacy of Combined Vaccine Based on the Induced HUVECs and DC-CT26 Against Colorectal Carcinoma


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Cell Lines
2.2. DCs Generation from Mouse Bone Marrow
2.3. Preparation of the CT26 Cell Culture Supernatant
2.4. Preparation of the HUVEC Vaccine
2.5. Preparation of the Induced HUVEC Vaccine
2.6. Preparation of the DC-CT26 Vaccine
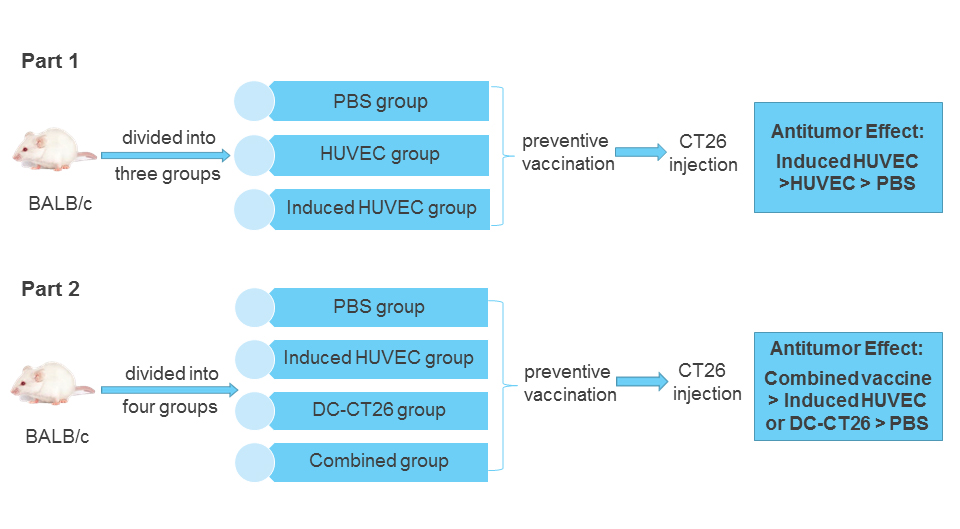
2.7. Vaccination Protocols in Tumor Models
Tumor Weight in the Experimental Group) / Average Tumor Weight in the Control
Group × 100%
2.8. Wound-Healing Assay
2.9. Transwell Assay
2.10. Immunohistochemistry and H&E Staining
2.11. Detection of the Anti-Induced HUVEC Membrane Protein Antibody by ELISA
2.12. Hemoglobin Assay
2.13. Western Blot
2.14. Flow Cytometry
2.15. Measurement of Cytokines
2.16. Cytotoxic T-Lymphocyte (CTL) Killing Assay
Release − Target Spontaneous Release)/(Target Maximal Release − Target
Spontaneous Release) × 100 %
2.17. Statistical Data Analysis
3. Results
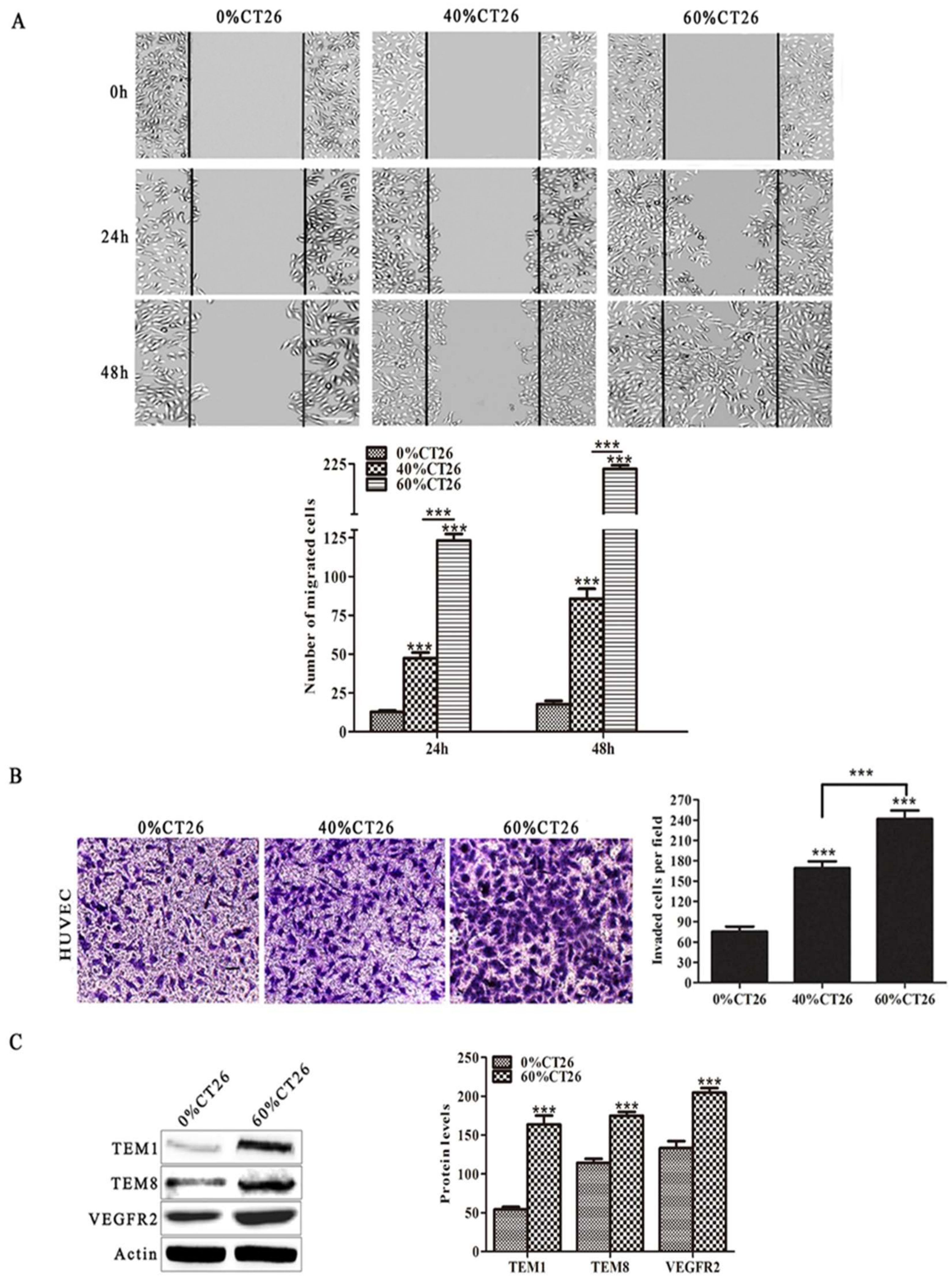
3.1. HUVECs Induced by 60% CT26 Cell Supernatant Had Characteristics Similar to Tumor Vascular Endothelial Cells
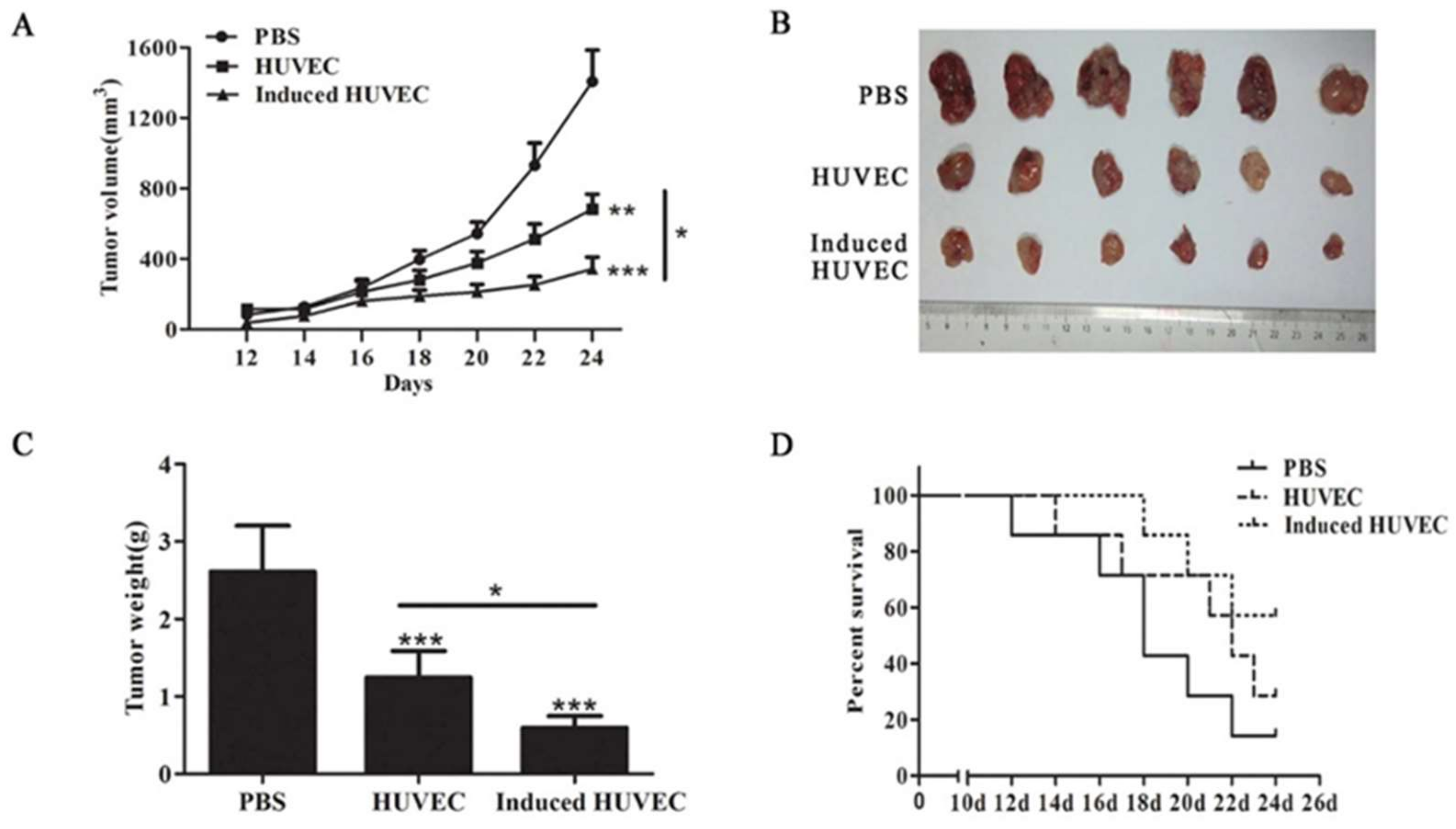
3.2. HUVEC Vaccine Induced by TCM Elicited a Better Antitumor Effect Than the HUVEC Vaccine
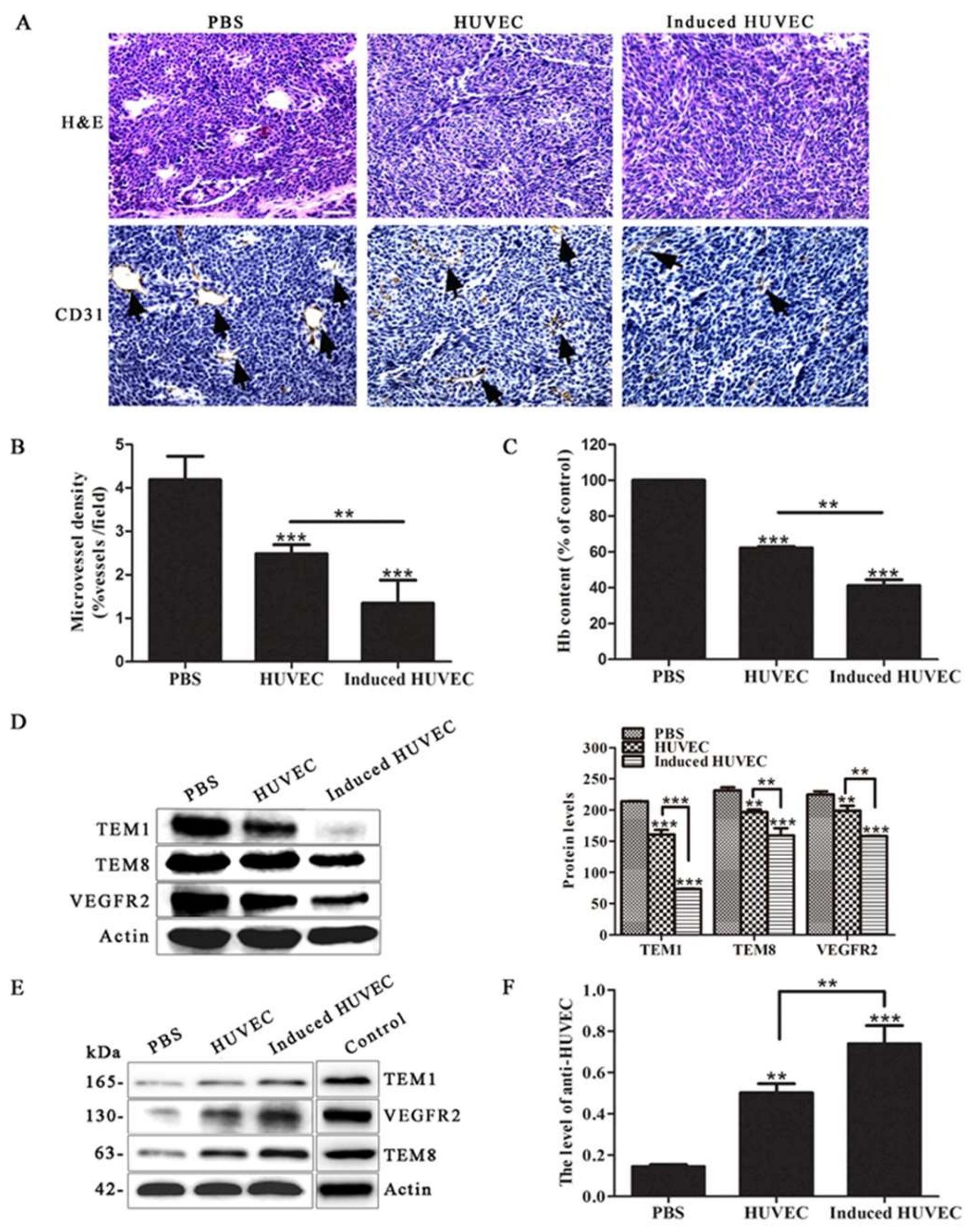
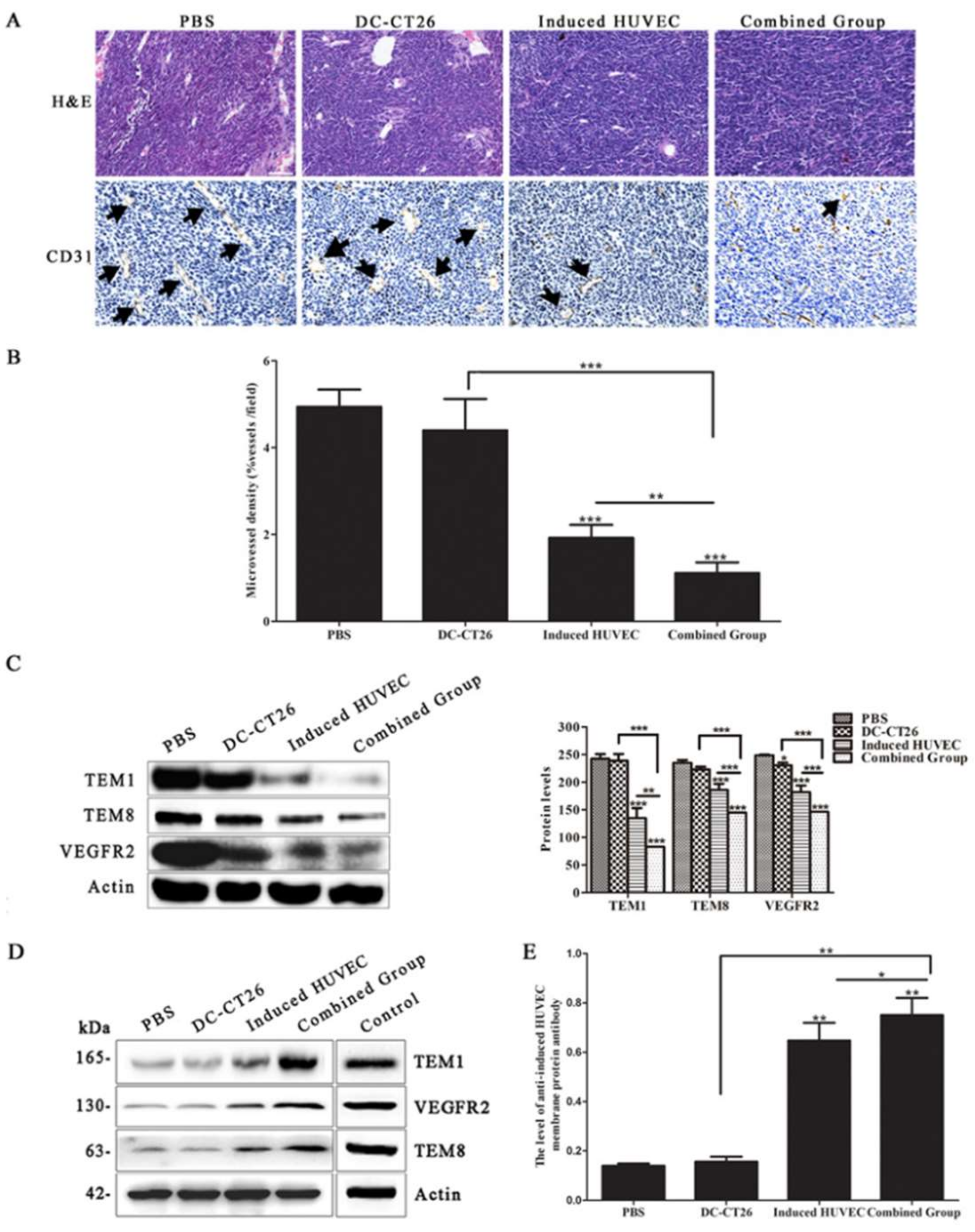
3.3. HUVEC Vaccine Induced by TCM Inhibited Tumor Angiogenesis
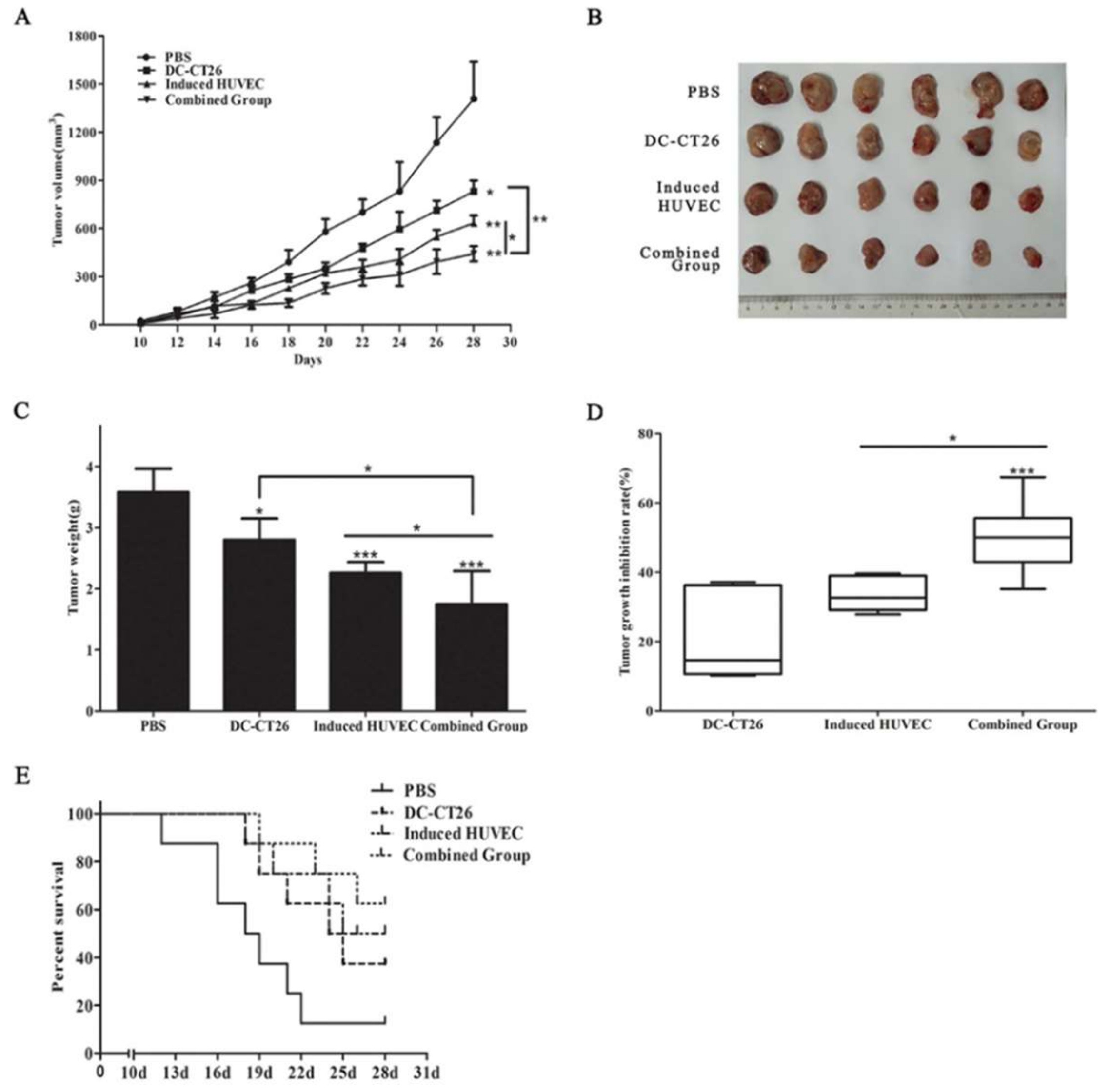
3.4. Combined Vaccine of DC-CT26 with Induced HUVECs Enhanced the Antitumor Effect
3.5. Combined Vaccine of DC-CT26 and Induced HUVECs Exerted a Better Antitumor Angiogenesis Effect Than DC-CT26 or Induced HUVECs Alone
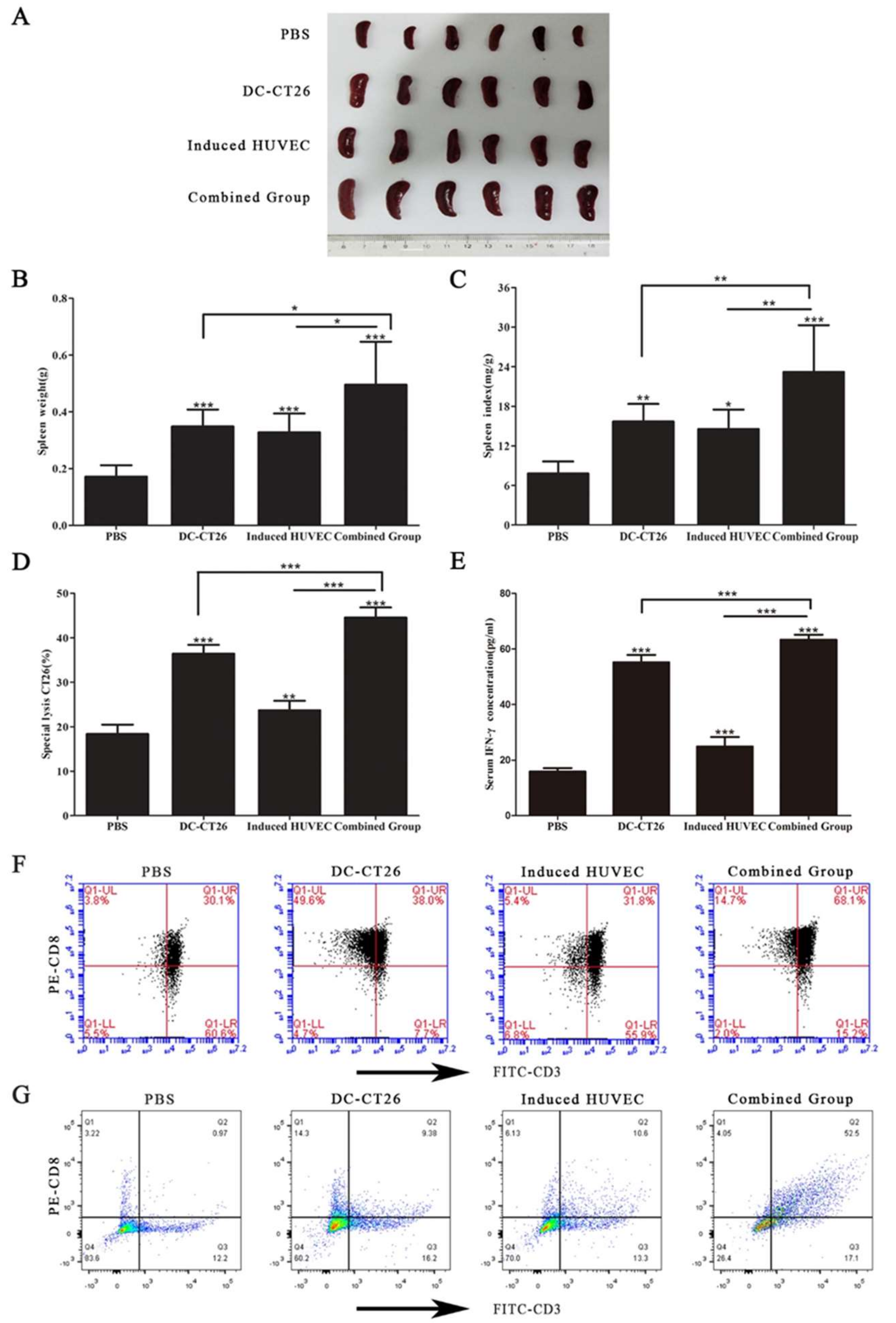
3.6. Combined Vaccine of DC-CT26 with Induced HUVECs Increased the Immune Function of the Spleen
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CRC | Colorectal carcinoma |
| CTLs | Cytotoxic T lymphocytes |
| DCs | Dendritic cells |
| DC-CT26 | Induced HUVECs and dendritic cell-loading CT26 antigen |
| FCM | Flow cytometry |
| H&E | Hematoxylin-eosin |
| HUVECs | Human umbilical vein endothelial cells |
| IFN-γ | Interferon-γ |
| LSD | Least standard difference |
| MVD | Microvessel density |
| TCM | Tumor conditioned medium |
| TEM | Tumor endothelial marker |
| VECs | Vascular endothelial cells |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Li, Y.; Yang, F.L.; Sang, L.X.; Zhu, J.F.; Han, X.; Shan, F.P.; Li, S.J.; Zhai, J.B.; Wang, D.N.; Lu, C.L.; et al. Enhanced therapeutic effects against murine colon carcinoma induced by a Colon 26/Ag85A-CD226 tumor cell vaccine. Oncol. Rep. 2015, 34, 1795–1804. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ebos, J.M.; Lee, C.R.; Cruz-Munoz, W.; Bjarnason, G.A.; Christensen, J.G.; Kerbel, R.S. Accelerated metastasis after short-term treatment with a potent inhibitor of tumor angiogenesis. Cancer Cell 2009, 15, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Farsaci, B.; Donahue, R.N.; Coplin, M.A.; Grenga, I.; Lepone, L.M.; Molinolo, A.A.; Hodge, J.W. Immune Consequences of Decreasing Tumor Vasculature with Antiangiogenic Tyrosine Kinase Inhibitors in Combination with Therapeutic Vaccines. Cancer Immunol. Res. 2014, 2, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Morera, Y.; Sanchez, J.; Bequet-Romero, M.; Selman-Housein, K.H.; de la Torre, A.; Hernandez-Bernal, F.; Martin, Y.; Garabito, A.; Pinero, J.; Bermudez, C.; et al. Specific humoral and cellular immune responses in cancer patients undergoing chronic immunization with a VEGF-based therapeutic vaccine. Vaccine 2017, 35, 3582–3590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lu, J.; Zhou, L.; Zhao, J.; Dong, Z. Efficacy for lung metastasis induced by the allogeneic bEnd3 vaccine in mice. Hum. Vaccin. Immunother. 2018, 14, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Q.; Wang, Q.R.; Zhao, X.; Yang, L.; Tian, L.; Lu, Y.; Kang, B.; Lu, C.J.; Huang, M.J.; Lou, Y.Y.; et al. Immunotherapy of tumors with xenogeneic endothelial cells as a vaccine. Nat. Med. 2000, 6, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Si, C.; Li, D.; Lu, M.; Zhong, W.; Xie, Z.; Guo, L.; Zhang, S.; Xu, M. Assessment of in vivo anti-tumor activity of human umbilical vein endothelial cell vaccines prepared by various antigen forms. Eur. J. Pharm. Sci. 2018, 114, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Zhang, W.; Zhang, W.; Wu, S.; Bi, F.; Su, J.J.; Tan, X.Y.; Liu, J.N.; Zhang, J. Vaccination with viable human umbilical vein endothelial cells prevents metastatic tumors by attack on tumor vasculature with both cellular and humoral immunity. Clin. Cancer Res. 2006, 12, 5834–5840. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, J.; Yang, Y.; Jin, G.; Zhang, X.; Wang, D.; Xie, C.; Liu, K.; Chen, X.; Li, X.; et al. Human umbilical vein endothelial cell vaccine suppresses the angiogenesis of esophageal squamous cell carcinoma in a humanized mouse model. Oncol. Rep. 2018, 40, 3006–3014. [Google Scholar] [CrossRef]
- Mu, X.Y.; Fang, C.J.; Zhou, J.; Xi, Y.F.; Zhang, L.; Wei, Y.Q.; Yi, T.; Wu, Y.; Zhao, X. Fusion with human lung cancer cells elongates the life span of human umbilical endothelial cells and enhances the anti-tumor immunity. J. Cancer Res. Clin. 2016, 142, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.L.; Zhou, L.; Zhang, Y.L.; Xie, Z.P.; Zhang, J.; Guo, L.; Wang, C.H.; Yang, X.P. A Fixed Human Umbilical Vein Endothelial Cell Vaccine With 2 Tandem Repeats of Microbial HSP70 Peptide Epitope 407–426 As Adjuvant for Therapy of Hepatoma in Mice. J. Immunother. 2015, 38, 276–284. [Google Scholar] [CrossRef]
- Tanaka, M.; Tsuno, N.H.; Fujii, T.; Todo, T.; Saito, N.; Takahashi, K. Human umbilical vein endothelial cell vaccine therapy in patients with recurrent glioblastoma. Cancer Sci. 2013, 104, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Alitalo, K. Clinical applications of angiogenic growth factors and their inhibitors. Nat. Med. 1999, 5, 1359–1364. [Google Scholar] [CrossRef]
- Balan, S.; Finnigan, J.; Bhardwaj, N. Dendritic Cell Strategies for Eliciting Mutation-Derived Tumor Antigen Responses in Patients. Cancer J. 2017, 23, 131–137. [Google Scholar] [CrossRef]
- Zhou, J.; Xi, Y.F.; Mu, X.Y.; Zhao, R.C.; Chen, H.D.; Zhang, L.; Wu, Y.; Li, Q. Antitumor immunity induced by VE-cadherin modified DC vaccine. Oncotarget 2017, 8, 67369–67379. [Google Scholar] [CrossRef]
- Shi, G.N.; Zhang, C.N.; Xu, R.; Niu, J.F.; Song, H.J.; Zhang, X.Y.; Wang, W.W.; Wang, Y.M.; Li, C.; Wei, X.Q.; et al. Enhanced antitumor immunity by targeting dendritic cells with tumor cell lysate-loaded chitosan nanoparticles vaccine. Biomaterials 2017, 113, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liang, S.J.; Zhang, C.L.; Liu, Y.L.; Yang, M.; Zhang, J.P.; Zhi, X.; Pan, F.; Cui, D.X. Allogenic dendritic cell and tumor cell fused vaccine for targeted imaging and enhanced immunotherapeutic efficacy of gastric cancer. Biomaterials 2015, 54, 177–187. [Google Scholar] [CrossRef]
- Garu, A.; Moku, G.; Gulla, S.K.; Chaudhuri, A. Genetic Immunization With In Vivo Dendritic Cell-targeting Liposomal DNA Vaccine Carrier Induces Long-lasting Antitumor Immune Response. Mol. Ther. 2016, 24, 385–397. [Google Scholar] [CrossRef]
- Gordy, J.T.; Luo, K.; Francica, B.; Drake, C.; Markham, R.B. Anti-IL-10-mediated Enhancement of Antitumor Efficacy of a Dendritic Cell-targeting MIP3alpha-gp100 Vaccine in the B16F10 Mouse Melanoma Model Is Dependent on Type I Interferons. J. Immunother. 2018, 41, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Rosalia, R.A.; Quakkelaar, E.D.; Redeker, A.; Khan, S.; Camps, M.; Drijfhout, J.W.; Silva, A.L.; Jiskoot, W.; van Hall, T.; van Veelen, P.A.; et al. Dendritic cells process synthetic long peptides better than whole protein, improving antigen presentation and T-cell activation. Eur. J. Immunol. 2013, 43, 2554–2565. [Google Scholar] [CrossRef]
- Meka, R.R.; Mukherjee, S.; Patra, C.R.; Chaudhuri, A. Shikimoyl-ligand decorated gold nanoparticles for use in ex vivo engineered dendritic cell based DNA vaccination. Nanoscale 2019, 11, 7931–7943. [Google Scholar] [CrossRef] [PubMed]
- Kenter, G.G.; Welters, M.J.P.; Valentijn, A.R.P.M.; Lowik, M.J.G.; Berends-van der Meer, D.M.A.; Vloon, A.P.G.; Essahsah, F.; Fathers, L.M.; Offringa, R.; Drijfhout, J.W.; et al. Vaccination against HPV-16 Oncoproteins for Vulvar Intraepithelial Neoplasia. N. Engl. J. Med. 2009, 361, 1838–1847. [Google Scholar] [CrossRef]
- Sabbatini, P.; Tsuji, T.; Ferran, L.; Ritter, E.; Sedrak, C.; Tuballes, K.; Jungbluth, A.A.; Ritter, G.; Aghajanian, C.; Bell-McGuinn, K.; et al. Phase I Trial of Overlapping Long Peptides from a Tumor Self-Antigen and Poly-ICLC Shows Rapid Induction of Integrated Immune Response in Ovarian Cancer Patients. Clin. Cancer Res. 2012, 18, 6497–6508. [Google Scholar] [CrossRef]
- Zeestraten, E.C.M.; Speetjens, F.M.; Welters, M.J.P.; Saadatmand, S.; Stynenbosch, L.F.M.; Jongen, R.; Kapiteijn, E.; Gelderblom, H.; Nijman, H.W.; Valentijn, A.R.P.M.; et al. Addition of interferon-alpha to the p53-SLP (R) vaccine results in increased production of interferon-gamma in vaccinated colorectal cancer patients: A phase I/II clinical trial. Int. J. Cancer 2013, 132, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Speetjens, F.M.; Kuppen, P.K.; Welters, M.J.P.; Essahsah, F.; van den Brink, A.M.E.G.V.; Lantrua, M.G.K.; Valentijn, A.R.P.M.; Oostendorp, J.; Fathers, L.M.; Nijman, H.W.; et al. Induction of p53-Specific Immunity by a p53 Synthetic Long Peptide Vaccine in Patients Treated for Metastatic Colorectal Cancer. Clin. Cancer Res. 2009, 15, 1086–1095. [Google Scholar] [CrossRef]
- Lu, J.; Zhao, J.; Liu, K.; Zhao, J.; Yang, H.; Huang, Y.; Qin, Z.; Bai, R.; Li, P.; Ma, J.; et al. MAPK/ERK1/2 signaling mediates endothelial-like differentiation of immature DCs in the microenvironment of esophageal squamous cell carcinoma. Cell Mol. Life Sci. 2010, 67, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Suri, R.M.; Niimi, M.; Ogilvie, A.L.; Kukutsch, N.A.; Rossner, S.; Schuler, G.; Austyn, J.M. Immature dendritic cells generated with low doses of GM-CSF in the absence of IL-4 are maturation resistant and prolong allograft survival in vivo. Eur. J. Immunol. 2000, 30, 1813–1822. [Google Scholar] [CrossRef]
- Yang, Y.; Lu, J.; Liu, H.F.; Jin, G.G.; Bai, R.H.; Li, X.; Wang, D.Y.; Zhao, J.M.; Huang, Y.T.; Liu, K.D.; et al. Dendritic cells loading autologous tumor lysate promote tumor angiogenesis. Tumor Biol. 2016, 37, 15687–15695. [Google Scholar] [CrossRef]
- Okaji, Y.; Tsuno, N.H.; Kitayama, J.; Saito, S.; Takahashi, T.; Kawai, K.; Yazawa, K.; Asakage, M.; Hori, N.; Watanabe, T.; et al. Vaccination with autologous endothelium inhibits angiogenesis and metastasis of colon cancer through autoimmunity. Cancer Sci. 2004, 95, 85–90. [Google Scholar] [CrossRef]
- Xu, M.L.; Xing, Y.; Zhou, L.; Yang, X.; Yao, W.J.; Xiao, W.; Ge, C.Y.; Ma, Y.J.; Yang, J.; Wu, J.; et al. Improved efficacy of therapeutic vaccination with viable human umbilical vein endothelial cells against murine melanoma by introduction of OK432 as adjuvant. Tumor Biol. 2013, 34, 1399–1408. [Google Scholar] [CrossRef]
- Nanda, A.; St Croix, B. Tumor endothelial markers: New targets for cancer therapy. Curr. Opin. Oncol. 2004, 16, 44–49. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Cyster, J.G. How T Cells Earn the Follicular Rite of Passage. Immunity 2011, 35, 671–680. [Google Scholar] [CrossRef]
- Mu, X.Y.; Sang, Y.X.; Fang, C.J.; Shao, B.; Yang, L.; Yao, K.; Zhao, X.T.; Gou, J.H.; Wei, Y.Q.; Yi, T.; et al. Immunotherapy of tumors with human telomerase reverse transcriptase immortalized human umbilical vein endothelial cells. Int. J. Oncol. 2015, 47, 1901–1911. [Google Scholar] [CrossRef]
- Scappaticci, F.A.; Contreras, A.; Boswell, C.A.; Lewis, J.S.; Nolan, G. Polyclonal antibodies to xenogeneic endothelial cells induce apoptosis and block support of tumor growth in mice. Vaccine 2003, 21, 2667–2677. [Google Scholar] [CrossRef]
- Jin, G.; Yang, Y.; Liu, K.; Zhao, J.; Chen, X.; Liu, H.; Bai, R.; Li, X.; Jiang, Y.; Zhang, X.; et al. Combination curcumin and (-)-epigallocatechin-3-gallate inhibits colorectal carcinoma microenvironment-induced angiogenesis by JAK/STAT3/IL-8 pathway. Oncogenesis 2017, 6. [Google Scholar] [CrossRef]
- Wagner, S.C.; Ichim, T.E.; Ma, H.; Szymanski, J.; Perez, J.A.; Lopez, J.; Bogin, V.; Patel, A.N.; Marincola, F.M.; Kesari, S. Cancer anti-angiogenesis vaccines: Is the tumor vasculature antigenically unique? J. Transl. Med. 2015, 13, 340. [Google Scholar] [CrossRef]
- Hida, K.; Ohga, N.; Akiyama, K.; Maishi, N.; Hida, Y. Heterogeneity of tumor endothelial cells. Cancer Sci. 2013, 104, 1391–1395. [Google Scholar] [CrossRef]
- Carson-Walter, E.B.; Watkins, D.N.; Nanda, A.; Vogelstein, B.; Kinzler, K.W.; Croix, B.S. Cell surface tumor endothelial markers are conserved in mice and humans. Cancer Res. 2001, 61, 6649–6655. [Google Scholar]
- Jyothsna, M.; Rammanohar, M.; Kumar, K. Histomorphometric Analysis of Angiogenesis using CD31 Immunomarker and Mast Cell Density in Oral Premalignant and Malignant Lesions: A Pilot Study. J. Clin. Diagn. Res. 2017, 11, 37–40. [Google Scholar] [CrossRef]
- Miyata, Y.; Sagara, Y.; Watanabe, S.; Asai, A.; Matsuo, T.; Ohba, K.; Hayashi, T.; Sakai, H. CD105 is a more appropriate marker for evaluating angiogenesis in urothelial cancer of the upper urinary tract than CD31 or CD34. Virchows Arch. 2013, 463, 673–679. [Google Scholar] [CrossRef]
- Sharma, B.; Singh, N.; Gupta, N.; Lal, P.; Pande, S.; Chauhan, S. Diagnostic Modalities of Precancerous and Cancerous Cervical Lesions with Special Emphasis on CD31 Angiogenesis Factor as a Marker. Patholog. Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Li, C.S.; Chacko, A.M.; Hu, J.; Hasegawa, K.; Swails, J.; Grasso, L.; El-Deiry, W.S.; Nicolaides, N.; Muzykantov, V.R.; Divgi, C.R.; et al. Antibody-based tumor vascular theranostics targeting endosialin/TEM1 in a new mouse tumor vascular model. Cancer Biol. Ther. 2014, 15, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Hilton, M.B.; Seaman, S.; Haines, D.C.; Stevenson, S.; Lemotte, P.K.; Tschantz, W.R.; Zhang, X.Y.M.; Saha, S.; Fleming, T.; et al. TEM8/ANTXR1 Blockade Inhibits Pathological Angiogenesis and Potentiates Tumoricidal Responses against Multiple Cancer Types. Cancer Cell 2012, 21, 212–226. [Google Scholar] [CrossRef]
- Zafarnia, S.; Bzyl-Ibach, J.; Spivak, I.; Li, Y.P.; Koletnik, S.; Doleschel, D.; Rix, A.; Pochon, S.; Tardy, I.; Koyadan, S.; et al. Nilotinib Enhances Tumor Angiogenesis and Counteracts VEGFR2 Blockade in an Orthotopic Breast Cancer Xenograft Model with Desmoplastic Response. Neoplasia 2017, 19, 896–907. [Google Scholar] [CrossRef]
- Chiang, C.L.; Benencia, F.; Coukos, G. Whole tumor antigen vaccines. Semin. Immunol. 2010, 22, 132–143. [Google Scholar] [CrossRef]
- Wallin, R.P.; Sundquist, V.S.; Brakenhielm, E.; Cao, Y.; Ljunggren, H.G.; Grandien, A. Angiostatic effects of NK cell-derived IFN-gamma counteracted by tumour cell Bcl-xL expression. Scand. J. Immunol. 2014, 79, 90–97. [Google Scholar] [CrossRef]
- Allen, H.; Shraga-Heled, N.; Blumenfeld, M.; Dego-Ashto, T.; Fuchs-Telem, D.; Gilert, A.; Aberman, Z.; Ofir, R. Human Placental-Derived Adherent Stromal Cells Co-Induced with TNF-alpha and IFN-gamma Inhibit Triple-Negative Breast Cancer in Nude Mouse Xenograft Models. Sci. Rep. 2018, 8, 670. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Banik, N.L.; Ray, S.K. Combination of hTERT knockdown and IFN-gamma treatment inhibited angiogenesis and tumor progression in glioblastoma. Clin. Cancer Res. 2009, 15, 7186–7195. [Google Scholar] [CrossRef]
- Gong, Z.; Xu, H.; Su, Y.; Wu, W.; Hao, L.; Han, C. Establishment of a Novel Bladder Cancer Xenograft Model in Humanized Immunodeficient Mice. Cell Physiol. Biochem. 2015, 37, 1355–1368. [Google Scholar] [CrossRef]
- Zhu, L.; Cao, J.; Chen, G.; Xu, Y.; Lu, J.; Fang, F.; Chen, K. Anti-tumor and immunomodulatory activities of an exopolysaccharide from Rhizopus nigricans on CT26 tumor-bearing mice. Int. Immunopharmacol. 2016, 36, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Yi, R.; Li, G.; Sun, P.; Qian, Y.; Zhao, X. Immunomodulatory Effect of Tremella Polysaccharides against Cyclophosphamide-Induced Immunosuppression in Mice. Molecules 2018, 23, 239. [Google Scholar] [CrossRef]
- Nazemalhosseini-Mojarad, E.; Mohammadpour, S.; Torshizi Esafahani, A.; Gharib, E.; Larki, P.; Moradi, A.; Amin Porhoseingholi, M.; Asadzade Aghdaei, H.; Kuppen, P.J.K.; Zali, M.R. Intratumoral infiltrating lymphocytes correlate with improved survival in colorectal cancer patients: Independent of oncogenetic features. J. Cell Physiol. 2019, 234, 4768–4777. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, S.L.; Zhang, H.R.; Gao, L.; Gao, X.; Liu, P.J.; Yi, Z.Y.; Li, N.; Xu, Z.Q. Combination radiotherapy and cantharidin inhibits lung cancer growth through altering tumor infiltrating lymphocytes. Future Oncol. 2017, 13, 1173–1180. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Xie, C.; Wang, D.; Yang, Y.; Liu, H.; Liu, K.; Zhao, J.; Chen, X.; Zhang, X.; Yang, W.; et al. Improved Antitumor Efficacy of Combined Vaccine Based on the Induced HUVECs and DC-CT26 Against Colorectal Carcinoma. Cells 2019, 8, 494. https://doi.org/10.3390/cells8050494
Zhang Q, Xie C, Wang D, Yang Y, Liu H, Liu K, Zhao J, Chen X, Zhang X, Yang W, et al. Improved Antitumor Efficacy of Combined Vaccine Based on the Induced HUVECs and DC-CT26 Against Colorectal Carcinoma. Cells. 2019; 8(5):494. https://doi.org/10.3390/cells8050494
Chicago/Turabian StyleZhang, Qiushuang, Chao Xie, Dongyu Wang, Yi Yang, Hangfan Liu, Kangdong Liu, Jimin Zhao, Xinhuan Chen, Xiaoyan Zhang, Wanjing Yang, and et al. 2019. "Improved Antitumor Efficacy of Combined Vaccine Based on the Induced HUVECs and DC-CT26 Against Colorectal Carcinoma" Cells 8, no. 5: 494. https://doi.org/10.3390/cells8050494
APA StyleZhang, Q., Xie, C., Wang, D., Yang, Y., Liu, H., Liu, K., Zhao, J., Chen, X., Zhang, X., Yang, W., Li, X., Tian, F., Dong, Z., & Lu, J. (2019). Improved Antitumor Efficacy of Combined Vaccine Based on the Induced HUVECs and DC-CT26 Against Colorectal Carcinoma. Cells, 8(5), 494. https://doi.org/10.3390/cells8050494