Physiological Adaptation to Simultaneous Chronic Exposure to High-Fat Diet and Dichlorodipheniletylhene (DDE) in Wistar Rat Testis


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Model
2.2. Lipid Peroxidation
2.3. SOD and GPx Activity Assay
2.4. Electrophoresis and Western Blot Analysis
2.5. Immunohistochemistry
2.6. Quantitative Real-Time PCR Analysis
2.7. Histological Analysis
2.8. Serum Testosterone Levels
2.9. Statistical Analysis
3. Results
3.1. High-Fat Diet and DDE Induce Testicular Lipid Peroxidation
3.2. Modulation of Antioxidant Enzymes Activities: SOD and GPx
3.3. High-Fat Diet and DDE Induce Pro-Apoptotic Stimuli
3.4. High-Fat Diet and DDE Induce PCNA Up-Regulation
3.5. Changes of Metallothioneins Expression and Synthesis
3.6. High-Fat Diet and DDE Induce Alterations in Testis Morphology
3.7. Serum Testosterone Levels
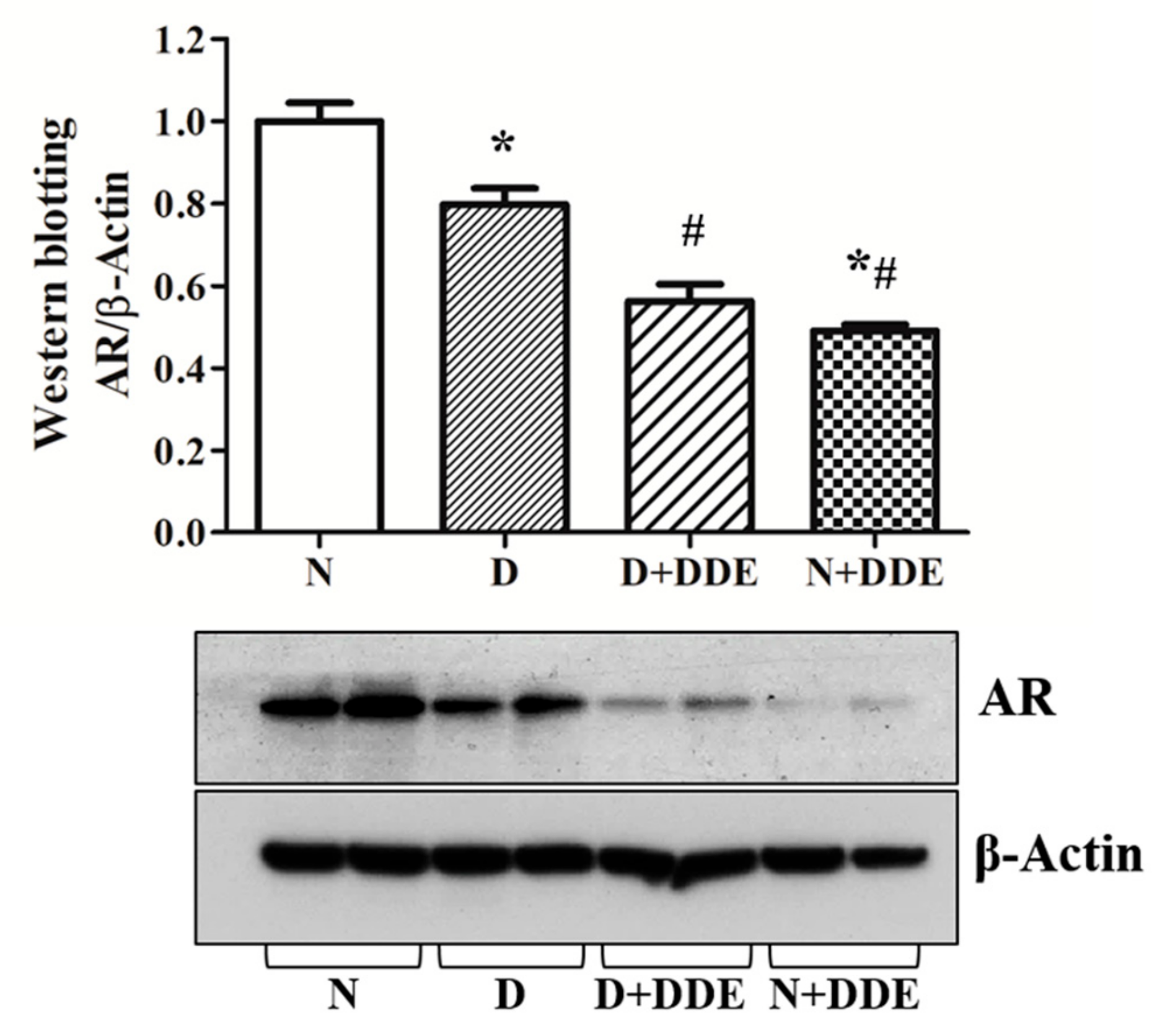
3.8. Androgen Receptor Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chavarro, J.E.; Toth, T.L.; Sadio, S.M.; Hauser, R. Soy food and isoflavone intake in relation to semen quality parameters among men from an infertility clinic. Human Reprod. 2008, 23, 2584–2590. [Google Scholar] [CrossRef]
- Ghanayem, B.I.; Bai, R.; Kissling, G.E.; Travlos, G.; Hoffler, U. Diet-induced obesity in male mice is associated with reduced fertility and potentiation of acrylamide-induced reproductive toxicity. Biol. Reprod. 2010, 82, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Hauser, R. Exposure to Polychlorinated Biphenyls (PCBs) and male reproduction. Syst. Biol. Reprod. Med. 2010, 56, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Bang, D.Y.; Kyung, M.; Kim, M.J.; Jung, B.Y.; Cho, M.C.; Choi, S.M.; Kim, Y.W.; Lim, S.K.; Lim, D.S.; Won, A.J.; et al. Human Risk Assessment of Endocrine-Disrupting Chemicals Derived from Plastic Food Containers. CRFSFS 2012, 11, 453–470. [Google Scholar] [CrossRef]
- Rato, L.; Alves, M.G.; Cavaco, J.E.; Oliveira, P.F. High-energy diets: A threat for male fertility? Obes. Rev. 2014, 15, 996–1007. [Google Scholar] [CrossRef]
- Gabrielsen, J.S.; Tanrikut, C. Chronic exposures and male fertility: The impacts of environment, diet, and drug use on spermatogenesis. Andrology 2016, 4, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Heitmann, B.L.; Blomberg, J.M.; Halldorsson, T.I.; Andersson, A.M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgård, C.; et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2013, 97, 411–418. [Google Scholar] [CrossRef]
- De Vriese, S.R.; Christophe, A.B. Fatty acid remodelling during sperm maturation: Variation of docosahexaenoic acid content. In Male Fertility and Lipid Metabolism; The American Oil Chemists Society: Urbana, IL, USA, 2003; pp. 96–117. [Google Scholar]
- Dagher, S.M.; Talhouk, R.S.; Nasrallah, S.S.; Tannous, R.I.; Mroueh, S.M. Relationship of dietary intake to DDE residues in breast milk of nursing mothers in Beirut. Food Add. Cont. 1999, 16, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.T.; Lee, T.K.M.; Chen, K.; Wong, H.L.; Zheng, J.S.; Giesy, J.P.; Lo, K.K.W.; Yamashita, N.; Lam, P.K.S. Human health risk assessment of organochlorines associated with fish consumption in a coastal city in China. Environ. Pollut. 2005, 136, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Ostby, J.; Monosson, E.; Kelce, W.R.; Gray, L.E. Environmental antiandrogens: Low doses of the fungicide vinclozolin alter sexual differentiation of the male rat. Toxicol. Ind. Health. 1999, 15, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Spira, A.; Multigner, L. Contribution of environmental factors to the risk of male infertility. Human Reprod. 2001, 16, 1768–1776. [Google Scholar] [CrossRef]
- Sikka, S.C. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef] [PubMed]
- De Jager, C.; Farias, P.; Barraza-Villarreal, A.; Avila, M.H.; Ayotte, P.; Dewailly, E.; Dombrowski, C.; Rousseau, F.; Sanchez, V.D.; Bailey, J.L. Reduced seminal parameters associated with environmental DDT exposure and p,p′-DDE concentrations in men in Chiapas, Mexico: A cross-sectional study. J. Androl. 2006, 27, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Schug, T.T.; Janesick, A.; Blumberg, B.; Heindel, J.J. Endocrine disrupting chemicals and disease susceptibility. J. Steroid Biochem. Mol. Biol. 2011, 117, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Streit, B. Bioaccumulation processes in ecosystems. Cell. Mol. Life Sci. 1992, 48, 955–970. [Google Scholar] [CrossRef]
- Jones, K.C.; de Voogt, P. Persistent organic pollutants (POPs): State of the science. Environ. Pollut. 1999, 100, 209–221. [Google Scholar] [CrossRef]
- Quinete, N.; Schettgen, T.; Bertram, J.; Kraus, T. Occurrence and distribution of PCB metabolites in blood and their potential health effects in humans: A review. Environ. Sci. Pollut. Res. Int. 2014, 21, 11951–11972. [Google Scholar] [PubMed]
- Bishopp, F.C. Insect Problems in World War II with Special References to the Insecticide DDT. Am. J. Public Health Nations Health 1945, 35, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Blus, L.J.; Wiemeyer, S.N.; Bunck, C.M. Clarification of effects of DDE on shell thickness, size, mass, and shape of avian eggs. Environ. Pollut. 1997, 95, 67–74. [Google Scholar] [CrossRef]
- Tiemann, U. In vivo and in vitro effects of the organochlorine pesticides DDT, TCPM, methoxychlor, and lindane on the female reproductive tract of mammals: A review. Reprod. Toxicol. 2008, 25, 316–326. [Google Scholar] [CrossRef]
- Norén, K.; Lunden, A.; Pettersson, E.; Bergman, A. Methylsulfonyl metabolites of PCBs and DDE in human milk in Sweden, 1972-1992. Environ. Health Perspect. 1996, 104, 766–772. [Google Scholar] [PubMed]
- Weistrand, C.; Norén, K. Methylsulfonyl metabolites of PCBs and DDE in human tissues. Environ. Health Persp. 1997, 105, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Ropstad, E.; Oskam, I.C.; Lyche, J.L.; Larsen, H.J.; Lie, E.; Haave, M.; Dahl, E.; Wiger, R.; Skaare, J.U. Endocrine disruption induced by organochlorines (OCs): Field studies and experimental models. J. Toxicol. Environ. Health A 2006, 8, 53–76. [Google Scholar] [CrossRef]
- Wang, X.P.; Gong, P.; Yao, T.D.; Jones, K.C. Passive Air Sampling of Organochlorine Pesticides, Polychlorinated Biphenyls, and Polybrominated Diphenyl Ethers Across the Tibetan Plateau. Environ. Sci. Technol. 2010, 44, 2988–2993. [Google Scholar] [CrossRef] [PubMed]
- Rehwagen, R. WHO recommends DDT to control malaria. BMJ 2006, 333, 622. [Google Scholar] [CrossRef]
- Kelce, W.R.; Stone, C.R.; Laws, S.C.; Gray, L.E.; Kemppainen, J.A.; Wilson, E.M. Persistent DDT metabolite p,p’–DDE is a potent androgen receptor antagonist. Nature 1995, 375, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.N.; Tanabe, S.; Tatsukawa, R.; Saito, S.; Miyazaki, N. Reduction in the testosterone levels by PCBs and DDE in Dall’s porpoises of northwestern North Pacific. Mar. Pollut. Bull. 1987, 18, 643–646. [Google Scholar] [CrossRef]
- Daxenberger, A. Pollutants with androgen-disrupting potency. Eur. J. Lipid Sci. Technol. 2002, 104, 124–130. [Google Scholar] [CrossRef]
- Rogan, W.J.; Chen, A. Health risks and benefits of bis(4-chlorophenyl)-1,1,1-trichloroethane (DDT). Lancet 2005, 366, 763–773. [Google Scholar] [CrossRef]
- Laird, B.D.; Goncharov, A.B.; Chan, H.M. Body burden of metals and persistent organic pollutants among Inuit in the Canadian Arctic. Environ. Int. 2013, 59, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.E., Jr.; Kelce, W.R. Latent effects of pesticides and toxic substances on sexual differentiation of rodents. Toxicol. Ind. Health 1996, 12, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.C.; Sun, H.; Chen, J.F.; Bian, Q.; Song, L.; Wang, X.R. Androgen receptor activities of p,p’-DDE, fenvalerate and phoxim detected by androgen receptor reporter gene assay. Toxicol. Lett. 2006, 160, 151–157. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Li, H.W.; Wang, Y.P.; Liu, C.J.; Yang, K.D. p,p′-DDE induces apoptosis and mRNA expression of apoptosis-associated genes in testes of pubertal rats. Environ. Toxicol. 2013, 28, 31–41. [Google Scholar] [CrossRef]
- Song, Y.; Liang, X.; Hu, Y.; Wang, Y.; Yu, H.; Yang, K. p,p′-DDE induces mitochondria-mediated apoptosis of cultured rat Sertoli cells. Toxicology 2008, 253, 53–61. [Google Scholar] [CrossRef]
- Fiorini, C.; Gilleron, J.; Carette, D.; Valette, A.; Tilloy, A.; Chevalier, S.; Segretain, D.; Pointis, G. Accelerated internalization of junctional membrane proteins (connexin 43, N-cadherin and ZO-1) within endocytic vacuoles: An early event of DDT carcinogenicity. Biochim. Biophys. Acta 2008, 1778, 56–67. [Google Scholar] [CrossRef]
- Tebourbi, O.; Driss, M.R.; Sakly, M.; Rhouma, K.B. Metabolism of DDT in different tissues of young rats. J. Environ. Sci. Health B 2006, 41, 167–176. [Google Scholar] [CrossRef]
- Cetkovic-Cvrlje, M.; Olson, M.; Schindler, B.; Gong, H.K. Exposure to DDT metabolite p,p′-DDE increases autoimmune type 1 diabetes incidence in NOD mouse model. J. Immunotoxicol. 2015, 13, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Salgado, B.; Olivero-Verbel, J.; Guerrero-Castilla, A. Direct effect of p,p′- DDT on mice liver. Braz. J. Pharm. Sci. 2016, 52, 287–298. [Google Scholar] [CrossRef]
- Shi, Y.Q.; Wang, Y.P.; Song, Y.; Li, H.W.; Liu, C.J.; Wu, Z.G.; Yang, K.D. p,p-DDE induces testicular apoptosis in prepubertal rats via the Fas/FasL pathway. Toxicol. Lett. 2010, 193, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Longnecker, M.P.; Klebanoff, M.A.; Zhou, H.; Brock, J.W. Association between maternal serum concentration of DDT metabolite DDE and preterm and small-for-gestational-age babies at birth. Lancet 2001, 14, 110–114. [Google Scholar] [CrossRef]
- Auer, T.; Khoschsorur, G.A.; Rabl, H.; Iberer, F.; Petutschnigg, B.; Wasler, A.; Tscheliessnigg, K.H. Detection of lipid peroxidation products by malondialdehyde (MDA-TBA reaction) in organ transplantation. Transplant. Proc. 1995, 27, 2749–2751. [Google Scholar] [PubMed]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. Oxid. Med. Cell. Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C. The enzymatic antioxidant system of human spermatozoa. Adv. Androl. 2014, 2014, 1–15. [Google Scholar]
- Makita, Y.; Matsuura, T.; Ogata, R.; Romero, Y.; Omura, M.; Tanaka, A.; Hirata, M.; Inoue, N. Systemic effects of orally administered p, p’-dde on immature male wistar rats during pubertal period. J. Occup. Health 2003, 45, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Lionetti, L.; Moreno, M.; Lombardi, A.; De Lange, P.; Antonelli, A.; Lanni, A.; Cavaliere, G.; Barletta, A.; Goglia, F. 3,5-diiodo-l-thyronine, by modulating mitochondrial functions, reverses hepatic fat accumulation in rats fed a high-fat diet. J. Hepatol. 2009, 51, 363–370. [Google Scholar] [CrossRef]
- Makita, Y.; Matsuura, T.; Ogata, R.; Omura, M.; Tanaka, A.; Hirata, M.; Inoue, N. Systemic toxicity of p,p′-DDE in aged male Wistar rats following oral administration. Fukuoka Igaku Zasshi. 2003, 94, 59–65. [Google Scholar]
- Laemmli, V.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Scudiero, R.; Cigliano, L.; Verderame, M. Age-related changes of metallothionein 1/2 and metallothionein 3 expression in rat brain. C. R. Biol. 2017, 340, 13–17. [Google Scholar] [CrossRef]
- Spagnuolo, M.S.; Mollica, M.P.; Maresca, B.; Cavaliere, G.; Cefaliello, C.; Trinchese, G.; Scudiero, R.; Crispino, M.; Cigliano, L. High Fat Diet and Inflammation – Modulation of Haptoglobin Level in Rat Brain. Fronti. Cell. Neurosci. 2015, 9, 1–9. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Leblond, C.P.; Clermont, Y. Definition of the stages of the cycle of the seminiferous epithelium in the rat. Ann. N. Y. Acad. Sci. 1952, 55, 548–573. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, V.; Lionetti, L.; Putti, R.; Sica, R.; Scudiero, R. Combined effects of DDE and hyperlipidic diet on metallothionein expression and synthesis in rat tissues. Environ. Toxicol. 2019, 34, 283–293. [Google Scholar] [PubMed]
- Liu, D.; Xu, Y. p53, oxidative stress, and aging. Antioxid. Redox. Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Kuwana, T.; Bouchier-Hayes, L.; Droin, N.M.; Newmeyer, D.D.; Schuler, M.; Green, D.R. Direct Activation of Bax by p53 Mediates Mitochondrial Membrane Permeabilization and Apoptosis. Science 2004, 303, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.H.; Mu, Y.; Wang, B.A.; Li, X.L.; Lu, J.M.; Li, J.Y.; Pan, C.Y.; Yanase, T.; Nawata, H. Saturated free fatty acids palmitic acid and stearic acid induce apoptosis by stimulation of ceramide generation in rat testicular Leydig cells. Biochem. Biophys. Res. Commun. 2003, 303, 1002–1007. [Google Scholar] [CrossRef]
- Siddighi, S.; Patton, W.C.; Jacobson, J.D.; King, A.; Chan, P.J. Correlation of sperm parameters with apoptosis assessed by dual fluorescence dna integrity assay. Arch. Androl. 2009, 50, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Franca, L.R.; Avelar, G.F.; Almeida, F.F. Spermatogenesis and sperm transit through the epi-didymis in mammals with emphasis on pigs. Theriogenology 2005, 63, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Shaha, C.; Tripathi, R.; Mishra, D.P. Male germ cell apoptosis: Regulation and biology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1501–1515. [Google Scholar] [CrossRef]
- Lizama, C.; Alfaro, I.; Reyes, J.G.; Moreno, R.D. Up-regulation of CD95 (Apo-1/Fas) is associated with spermatocyte apoptosis during the first round of spermatogenesis in the rat. Apoptosis 2007, 12, 499–512. [Google Scholar] [CrossRef]
- Vidal, J.D.; Whitney, K.M. Morphologic manifestations of testicular and epididymal toxicity. Spermatogenesis 2014, 4, 1–17. [Google Scholar] [CrossRef]
- Creasy, D.M. Pathogenesis of Male Reproductive Toxicity. Toxicol. Pathol. 2001, 29, 64–76. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, Y.; Xue, K.; Gu, G.; Fan, W.; Xu, Y.; Ding, Z. Diet-induced obesity in male C57BL/6 mice decreases fertility as a consequence of disrupted blood-testis barrier. PLoS ONE 2015, 10, e0120775. [Google Scholar]
- Arrebola, J.P.; Cuellar, M.; Claure, E.; Quevedo, M.; Antelo, S.R.; Mutch, E.; Ramirez, E.; Fernandez, M.F.; Olea, N.; Mercado, L.A. Concentrations of organochlorine pesticides and polychlorinated biphenyls in human serum and adipose tissue from Bolivia. Environ. Res. 2012, 112, 40–47. [Google Scholar] [CrossRef]
- Migliaccio, V.; Scudiero, R.; Sica, R.; Lionetti, L.; Putti, R. Oxidative stress and mitochondrial uncoupling protein 2 expression in hepatic steatosis induced by exposure to xenobiotic DDE and high fat diet in male Wistar rats. PLoS ONE 2019, 14, e0215955. [Google Scholar] [CrossRef]
- Marouani, N.; Hallegue, D.; Sakly, M.; Benkhalifa, M.; Rhouma, K.B.; Tebourbi, O. p,p′-DDT induces testicular oxidative stress-induced apoptosis in adult rats. Reprod. Biol. Endocrinol. 2017, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Liu, J.; Jin, X.; Li, Z.; Zhao, M.; Liu, W. p,p′-Dichlorodiphenyldichloroethylene induces colorectal adenocarcinoma cell proliferation through oxidative stress. PLoS ONE 2014, 9, e112700. [Google Scholar] [CrossRef]
- Penna-Videau, S.; Bustos-Obregon, E.; Cermeno-Vivas, J.R.; Chirino, D. Malathion affects spermatogenic proliferation in mouse. Int. J. Morphol. 2012, 30, 1399–1407. [Google Scholar] [CrossRef]
- Lebda, M.; Gad, S.; Gaafar, H. Effects of lipoic acid on acrylamide induced testicular damage. Mater. Sociomed. 2014, 26, 208–212. [Google Scholar] [CrossRef]
- Türedi, S.; Yuluğ, E.; Alver, A.; Kutlu, O.; Kahraman, C. Effects of resveratrol on doxorubicin induced testicular damage in rats. Experim. Toxicol. Pathol. 2015, 67, 229–235. [Google Scholar] [CrossRef]
- Fiorini, C.; Tilloy-Ellul, A.; Chevalier, S.; Charuel, C.; Pointis, G. Sertoli cell junctional proteins as early targets for different classes of reproductive toxicantsl reprod. Toxicolology 2004, 18, 413–421. [Google Scholar]
- Jandacek, R.J.; Heubi, J.E.; Buckley, D.D.; Khoury, J.C.; Turner, W.E.; Sjödin, A.; Olson, J.R.; Shelton, C.; Helms, K.; Bailey, T.D.; et al. Reduction of the Body Burden of PCBs and DDE by Dietary Intervention in a Randomized Trial. J. Nutr. Biochem. 2014, 25, 483–488. [Google Scholar]
- Pareek, T.K.; Joshi, A.R.; Sanyal, A.; Dighe, R.R. Insights into male germ cell apoptosis due to depletion of gonadotropins caused by GnRH antagonists. Apoptosis 2007, 12, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Danzo, B.J. Environmental xenobiotics may disrupt normal endocrine function by interfering with the binding of physiological ligands to steroid receptors and binding proteins. Environ. Health Perspect. 1997, 105, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Luccio-Camelo, D.C.; Prins, G.S. Disruption of androgen receptor signaling in males by environmental chemicals. J. Steroid Biochem. Mol. Biol. 2011, 127, 74–82. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliaccio, V.; Sica, R.; Scudiero, R.; Simoniello, P.; Putti, R.; Lionetti, L. Physiological Adaptation to Simultaneous Chronic Exposure to High-Fat Diet and Dichlorodipheniletylhene (DDE) in Wistar Rat Testis. Cells 2019, 8, 443. https://doi.org/10.3390/cells8050443
Migliaccio V, Sica R, Scudiero R, Simoniello P, Putti R, Lionetti L. Physiological Adaptation to Simultaneous Chronic Exposure to High-Fat Diet and Dichlorodipheniletylhene (DDE) in Wistar Rat Testis. Cells. 2019; 8(5):443. https://doi.org/10.3390/cells8050443
Chicago/Turabian StyleMigliaccio, Vincenzo, Raffaella Sica, Rosaria Scudiero, Palma Simoniello, Rosalba Putti, and Lillà Lionetti. 2019. "Physiological Adaptation to Simultaneous Chronic Exposure to High-Fat Diet and Dichlorodipheniletylhene (DDE) in Wistar Rat Testis" Cells 8, no. 5: 443. https://doi.org/10.3390/cells8050443
APA StyleMigliaccio, V., Sica, R., Scudiero, R., Simoniello, P., Putti, R., & Lionetti, L. (2019). Physiological Adaptation to Simultaneous Chronic Exposure to High-Fat Diet and Dichlorodipheniletylhene (DDE) in Wistar Rat Testis. Cells, 8(5), 443. https://doi.org/10.3390/cells8050443