

Development of Comorbid Depression after Social Fear Conditioning in Mice and Its Effects on Brain Sphingolipid Metabolism

 ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Overview
2.3. Social Fear Conditioning (SFC) Paradigm
2.4. Social Fear Assessment
2.5. Elevated Plus-Maze (EPM) Test
2.6. Forced Swim Test (FST)
2.7. Measurement of Sphingomyelinase and Ceramidase Activities
2.8. Sphingolipid Quantification by Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
2.9. Statistical Analysis
3. Results
3.1. Short-Term Effects of SFC on Social Fear, Non-Social Anxiety-Like Behavior and Depressive-Like Behavior
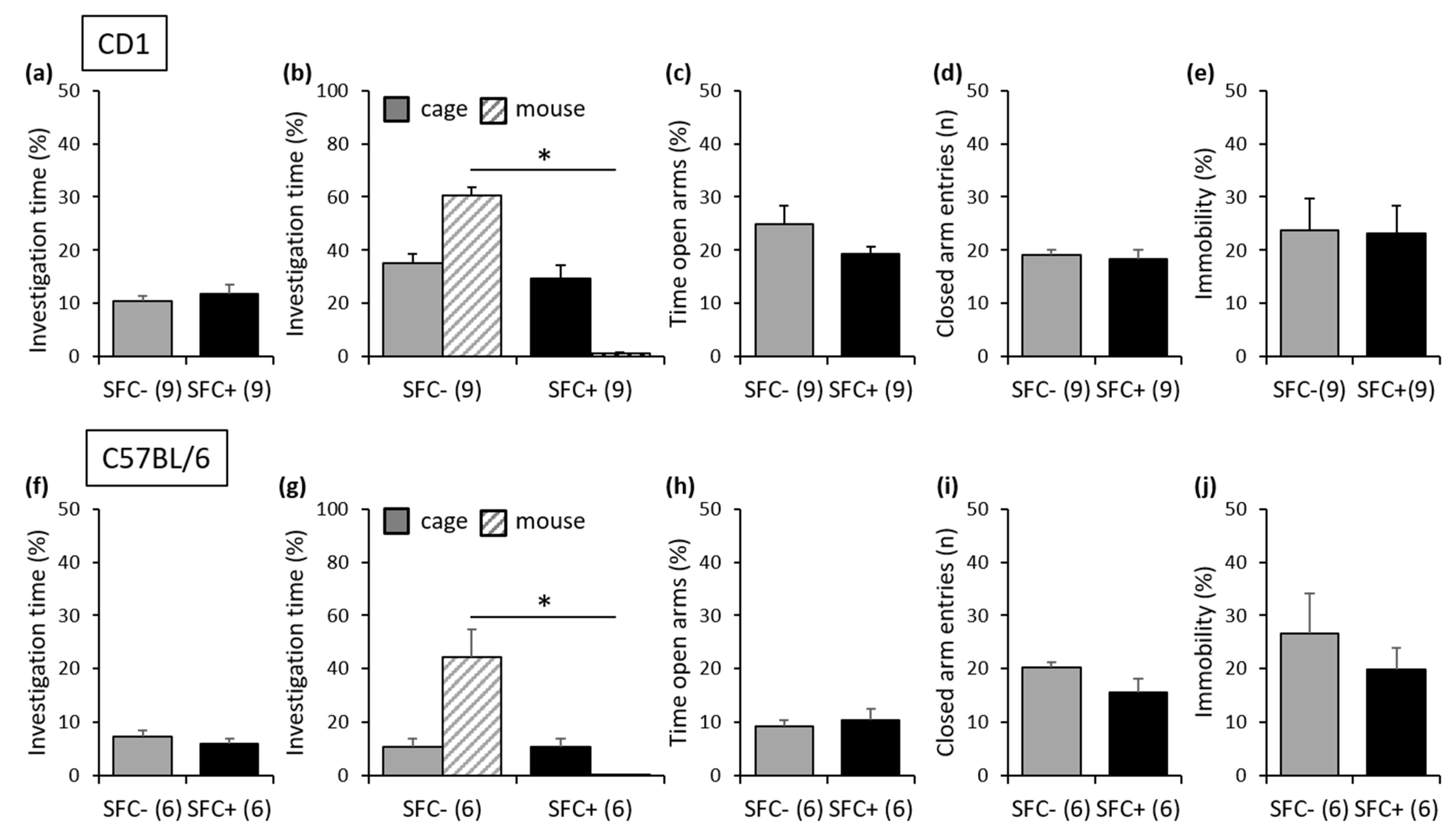
3.1.1. Outbred CD1 Mice
3.1.2. Inbred C57BL/6 Mice
3.2. Long-Term Effects of SFC on Social Fear, Non-Social Anxiety-like Behavior and Depressive-Like Behavior
3.2.1. Outbred CD1 Mice
3.2.2. Inbred C57BL/6 Mice
3.3. Strain Differences in Behavior
3.4. Short-Term Effects of SFC on Brain Sphingolipid Metabolizing Enzymes
3.5. Short-Term Effects of SFC on Brain Sphingolipids
3.6. Long-Term Effects of SFC on Brain Sphingolipid Metabolizing Enzymes
3.7. Long-Term Effects of SFC on Brain Sphingolipids
3.8. Effects of Duration of Single Housing on Brain Sphingolipid Metabolizing Enzymes, Sphingolipid Levels and Sphingolipid Ratios
4. Discussion
4.1. Development of Comorbid Depression after SFC in Male Mice
4.2. Effects of SFC on Brain Sphingolipid Metabolizing Enzymes
4.3. Effects of SFC on Brain Sphingolipids
4.4. Effects of Prolonged Single Housing on Behavior and Brain Sphingolipid Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Berglund, P.; Demler, O.; Jin, R.; Merikangas, K.R.; Walters, E.E. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch. Gen. Psychiatry 2005, 62, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Stangier, U.; Heidenreich, T.; Schermelleh-Engel, K. Safety behaviors and social performance in patients with generalized social phobia. J. Cogn. Psychother. 2006, 20, 17–31. [Google Scholar] [CrossRef]
- Koyuncu, A.; İnce, E.; Ertekin, E.; Tükel, R. Comorbidity in social anxiety disorder: Diagnostic and therapeutic challenges. Drugs Context 2019, 8, 212573. [Google Scholar] [CrossRef] [PubMed]
- Chartier, M.J.; Walker, J.R.; Stein, M.B. Considering comorbidity in social phobia. Soc. Psychiatry Psychiatr. Epidemiol. 2003, 38, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Belzer, K.; Schneier, F.R. Comorbidity of anxiety and depressive disorders: Issues in conceptualization, assessment, and treatment. J. Psychiatr. Pract. 2004, 10, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.; Neumann, I.D. Animal models of social avoidance and social fear. Cell. Tissue Res. 2013, 354, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social fear conditioning: A novel and specific animal model to study social anxiety disorder. Neuropsychopharmacology 2012, 37, 1433–1443. [Google Scholar] [CrossRef]
- Toth, I.; Neumann, I.D.; Slattery, D.A. Social Fear Conditioning as an Animal Model of Social Anxiety Disorder. Curr. Protoc. Neurosci. 2013, 63, 9–42. [Google Scholar] [CrossRef]
- Kornhuber, J.; Huber, S.E.; Zoicas, I. Effects of conditioned social fear on ethanol drinking and vice-versa in male mice. Psychopharmacology 2019, 236, 2059–2067. [Google Scholar] [CrossRef]
- Müller, C.P.; Reichel, M.; Mühle, C.; Rhein, C.; Gulbins, E.; Kornhuber, J. Brain membrane lipids in major depression and anxiety disorders. Biochim. Biophys. Acta 2015, 1851, 1052–1065. [Google Scholar] [CrossRef]
- Bienias, K.; Fiedorowicz, A.; Sadowska, A.; Prokopiuk, S.; Car, H. Regulation of sphingomyelin metabolism. Pharmacol. Rep. 2016, 68, 570–581. [Google Scholar] [CrossRef]
- Milhas, D.; Clarke, C.J.; Hannun, Y.A. Sphingomyelin metabolism at the plasma membrane: Implications for bioactive sphingolipids. FEBS Lett. 2010, 584, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Mühle, C.; BilbaoCanalejas, R.D.; Kornhuber, J. Sphingomyelin synthases in neuropsychiatric health and disease. Neurosignals 2019, 27, 54–76. [Google Scholar] [CrossRef] [PubMed]
- Goñi, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef]
- Lahiri, S.; Futerman, A.H. The metabolism and function of sphingolipids and glycosphingolipids. Cell. Mol. Life Sci. 2007, 64, 2270–2284. [Google Scholar] [CrossRef]
- Okino, N.; He, X.; Gatt, S.; Sandho, K.; Ito, M.; Schuchman, E.H. The reverse activity of human acid ceramidase. J. Biol. Chem. 2003, 278, 29948–29953. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.; Akkaoui, J.; Yamada, C.; Ho, A.; Mao, C.; Movila, A. Elusive roles of the different ceramidases in human health, pathophysiology, and tissue regeneration. Cells 2020, 9, 1379. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, M.; Scandroglio, F.; Prioni, S.; Buccinnà, B.; Loberto, N.; Aureli, M.; Chigorno, V.; Lupino, E.; DeMarco, G.; Lomartire, A.; et al. Deregulated sphingolipid metabolism and membrane organization in neurodegenerative disorders. Mol. Neurobiol. 2010, 41, 314–340. [Google Scholar] [CrossRef]
- Mühle, C.; Reichel, M.; Gulbins, E.; Kornhuber, J. Sphingolipids in psychiatric disorders and pain syndromes. Handb. Exp. Pharmacol. 2013, 216, 431–456. [Google Scholar] [CrossRef]
- Kornhuber, J.; Medlin, A.; Bleich, S.; Jendrossek, V.; Henkel, A.W.; Wiltfang, J.; Gulbins, E. High activity of acid sphingomyelinase in major depression. J. Neural Transm. 2005, 112, 1583–1590. [Google Scholar] [CrossRef]
- Mühle, C.; Wagner, C.J.; Färber, K.; Richter-Schmidinger, T.; Gulbins, E.; Lenz, B.; Kornhuber, J. Secretory acid sphingomyelinase in the serum of medicated patients predicts the prospective course of depression. J. Clin. Med. 2019, 8, 846. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, F.; Edwards, M.J.; Mühle, C.; Carpinteiro, A.; Wilson, G.C.; Wilker, B.; Soddemann, M.; Keitsch, S.; Scherbaum, N.; Müller, B.W.; et al. Ceramide levels in blood plasma correlate with major depressive disorder severity and its neutralization abrogates depressive behavior in mice. J. Biol. Chem. 2022, 298, 102185. [Google Scholar] [CrossRef] [PubMed]
- Gracia-Garcia, P.; Rao, V.; Haughey, N.J.; Bandaru, V.V.; Smith, G.; Rosenberg, P.B.; Lobo, A.; Lyketsos, C.G.; Mielke, M.M. Elevated plasma ceramides in depression. J. Neuropsychiatry Clin. Neurosci. 2011, 23, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Brunkhorst-Kanaan, N.; Klatt-Schreiner, K.; Hackel, J.; Schroter, K.; Trautmann, S.; Hahnefeld, L.; Wicker, S.; Reif, A.; Thomas, D.; Geisslinger, G.; et al. Targeted lipidomics reveal derangement of ceramides in major depression and bipolar disorder. Metabolism 2019, 95, 65–76. [Google Scholar] [CrossRef]
- Dinoff, A.; Saleem, M.; Herrmann, N.; Mielke, M.M.; Oh, P.I.; Venkata, S.L.V.; Haughey, N.J.; Lanctot, K.L. Plasma sphingolipids and depressive symptoms in coronary artery disease. Brain Behav. 2017, 7, e00836. [Google Scholar] [CrossRef]
- Moaddel, R.; Shardell, M.; Khadeer, M.; Lovett, J.; Kadriu, B.; Ravichandran, S.; Morris, P.J.; Yuan, P.; Thomas, C.J.; Gould, T.D.; et al. Plasma metabolomic profiling of a ketamine and placebo crossover trial of major depressive disorder and healthy control subjects. Psychopharmacology 2018, 235, 3017–3030. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Zheng, P.; Zhao, X.; Zhou, C.; Hu, C.; Hou, X.; Wang, H.; Xie, P.; Xu, G. Plasma lipidomics reveals potential lipid markers of major depressive disorder. Anal. Bioanal. Chem. 2016, 408, 6497–6507. [Google Scholar] [CrossRef]
- Gulbins, E.; Palmada, M.; Reichel, M.; Lüth, A.; Böhmer, C.; Amato, D.; Müller, C.P.; Tischbirek, C.H.; Groemer, T.W.; Tabatabai, G.; et al. Acid sphingomyelinase-ceramide system mediates effects of antidepressant drugs. Nature Med. 2013, 19, 934–938. [Google Scholar] [CrossRef]
- Zoicas, I.; Reichel, M.; Gulbins, E.; Kornhuber, J. Role of acid sphingomyelinase in the regulation of social behavior and memory. PLoS ONE. 2016, 11, e0162498. [Google Scholar] [CrossRef]
- Müller, C.P.; Kalinichenko, L.S.; Tiesel, J.; Witt, M.; Stöckl, T.; Sprenger, E.; Fuchser, J.; Beckmann, J.; Praetner, M.; Huber, S.E.; et al. Paradoxical antidepressant effects of alcohol are related to acid sphingomyelinase and its control of sphingolipid homeostasis. Acta Neuropathol. 2017, 133, 463–483. [Google Scholar] [CrossRef]
- Zoicas, I.; Schumacher, F.; Kleuser, B.; Reichel, M.; Gulbins, E.; Fejtova, A.; Kornhuber, J.; Rhein, C. The forebrain-specific overexpression of acid sphingomyelinase induces depressive-like symptoms in mice. Cells 2020, 9, 1244. [Google Scholar] [CrossRef] [PubMed]
- Schmidtner, A.K.; Slattery, D.A.; Gläsner, J.; Hiergeist, A.; Gryksa, K.; Malik, V.A.; Hellmann-Regen, J.; Heuser, I.; Baghai, T.C.; Gessner, A.; et al. Minocycline alters behavior, microglia and the gut microbiome in a trait-anxiety-dependent manner. Transl. Psychiatry 2019, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Zoicas, I.; Mühle, C.; Schmidtner, A.K.; Gulbins, E.; Neumann, I.D.; Kornhuber, J. Anxiety and depression are related to higher activity of sphingolipid metabolizing enzymes in the rat brain. Cells 2020, 9, 1239. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef]
- Oliveira, T.G.; Chan, R.B.; Bravo, F.V.; Miranda, A.; Silva, R.R.; Zhou, B.; Marques, F.; Pinto, V.; Cerqueira, J.J.; Di Paolo, G.; et al. The impact of chronic stress on the rat brain lipidome. Mol. Psychiatry 2016, 21, 80–88. [Google Scholar] [CrossRef]
- Gregus, A.; Wintink, A.J.; Davis, A.C.; Kalynchuk, L.E. Effect of repeated corticosterone injections and restraint stress on anxiety and depression-like behavior in male rats. Behav. Brain Res. 2005, 156, 105–114. [Google Scholar] [CrossRef]
- Miranda, A.M.; Bravo, F.V.; Chan, R.B.; Sousa, N.; Di Paolo, G.; Oliveira, T.G. Differential lipid composition and regulation along the hippocampal longitudinal axis. Transl. Psychiatry 2019, 9, 144. [Google Scholar] [CrossRef]
- Zoicas, I.; Huber, S.E.; Kalinichenko, L.S.; Gulbins, E.; Müller, C.P.; Kornhuber, J. Ceramides affect alcohol consumption and depressive-like and anxiety-like behavior in a brain region- and ceramide species-specific way in male mice. Addict. Biol. 2019, 25, e12847. [Google Scholar] [CrossRef]
- Michalikova, S.; van Rensburg, R.; Chazot, P.L.; Ennaceur, A. Anxiety responses in Balb/c, c57 and CD-1 mice exposed to a novel open space test. Behav. Brain Res. 2010, 207, 402–417. [Google Scholar] [CrossRef]
- Benatti, C.; Alboni, S.; Montanari, C.; Caggia, F.; Tascedda, F.; Brunello, N.; Blom, J.M. Central effects of a local inflammation in three commonly used mouse strains with a different anxious phenotype. Behav. Brain Res. 2011, 224, 23–34. [Google Scholar] [CrossRef]
- Painsipp, E.; Köfer, M.J.; Sinner, F.; Holzer, P. Prolonged depression-like behavior caused by immune challenge: Influence of mouse strain and social environment. PLoS ONE 2011, 6, e20719. [Google Scholar] [CrossRef]
- de Sá-Calçada, D.; Roque, S.; Branco, C.; Monteiro, S.; Cerqueira-Rodrigues, B.; Sousa, N.; Palha, J.A.; Correia-Neves, M. Exploring female mice interstrain differences relevant for models of depression. Front. Behav. Neurosci. 2015, 9, 335. [Google Scholar] [CrossRef] [PubMed]
- Faure, A.; Pittaras, E.; Nosjean, A.; Chabout, J.; Cressant, A.; Granon, S. Social behaviors and acoustic vocalizations in different strains of mice. Behav. Brain Res. 2017, 320, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.S.; Wen, J.H.; Miyares, L.; Lombroso, P.J.; Bordey, A. Outbred CD1 mice are as suitable as inbred C57BL/6J mice in performing social tasks. Neurosci. Lett. 2017, 637, 142–147. [Google Scholar] [CrossRef]
- Zoicas, I.; Slattery, D.A.; Neumann, I.D. Brain oxytocin in social fear conditioning and its extinction: Involvement of the lateral septum. Neuropsychopharmacology 2014, 39, 3027–3035. [Google Scholar] [CrossRef] [PubMed]
- Zoicas, I.; Menon, R.; Neumann, I.D. Neuropeptide S reduces fear and avoidance of con-specifics induced by social fear conditioning and social defeat, respectively. Neuropharmacology 2016, 108, 284–291. [Google Scholar] [CrossRef]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y reduces expression of social fear via simultaneous activation of Y1 and Y2 receptors. J. Psychopharmacol. 2019, 33, 1533–1539. [Google Scholar] [CrossRef]
- Kornhuber, J.; Zoicas, I. Neuropeptide Y as alternative pharmacotherapy for antidepressant-resistant social fear. Int. J. Mol. Sci. 2020, 21, 8220. [Google Scholar] [CrossRef]
- Kornhuber, J.; Zoicas, I. Social fear memory requires two stages of protein synthesis in mice. Int. J. Mol. Sci. 2020, 21, 5537. [Google Scholar] [CrossRef]
- Kornhuber, J.; Zoicas, I. Brain region-dependent effects of neuropeptide Y on conditioned social fear and anxiety-like behavior in male mice. Int. J. Mol. Sci. 2021, 22, 3695. [Google Scholar] [CrossRef]
- Mühle, C.; Kornhuber, J. Assay to measure sphingomyelinase and ceramidase activities efficiently and safely. J. Chromatogr. A 2017, 1481, 137–144. [Google Scholar] [CrossRef]
- Gulbins, A.; Schumacher, F.; Becker, K.A.; Wilker, B.; Soddemann, M.; Boldrin, F.; Müller, C.P.; Edwards, M.J.; Goodman, M.; Caldwell, C.C.; et al. Antidepressants act by inducing autophagy controlled by sphingomyelin-ceramide. Mol. Psychiatry 2018, 23, 2324–2346. [Google Scholar] [CrossRef] [PubMed]
- Naser, E.; Kadow, S.; Schumacher, F.; Mohamed, Z.H.; Kappe, C.; Hessler, G.; Pollmeier, B.; Kleuser, B.; Arenz, C.; Becker, K.A.; et al. Characterization of the small molecule ARC39, a direct and specific inhibitor of acid sphingomyelinase in vitro. J. Lipid Res. 2020, 61, 896–910. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, K.A.; Maren, S. Hippocampal inactivation disrupts contextual retrieval of fear memory after extinction. J. Neurosci. 2001, 21, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Maren, S.; Phan, K.L.; Liberzon, I. The contextual brain: Implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 2013, 14, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Trainor, B.C. Stress responses and the mesolimbic dopamine system: Social contexts and sex differences. Horm. Behav. 2011, 60, 457–469. [Google Scholar] [CrossRef]
- Baldaçara, L.; Borgio, J.G.; Lacerda, A.L.; Jackowski, A.P. Cerebellum and psychiatric disorders. Braz. J. Psychiatry 2008, 30, 281–289. [Google Scholar] [CrossRef]
- Moreno-Rius, J. Is there an “antisocial” cerebellum? Evidence from disorders other than autism characterized by abnormal social behaviours. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 1–8. [Google Scholar] [CrossRef]
- Venkatraman, A.; Edlow, B.L.; Immordino-Yang, M.H. The brainstem in emotion: A review. Front. Neuroanat. 2017, 11, 15. [Google Scholar] [CrossRef]
- Boot, R.G.; Verhoek, M.; Donker-Koopman, W.; Strijland, A.; van Marle, J.; Overkleeft, H.S.; Wennekes, T.; Aerts, J.M. Identification of the non-lysosomal glucosylceramidase as beta-glucosidase 2. J. Biol. Chem. 2007, 282, 1305–1312. [Google Scholar] [CrossRef]
- Thorsell, A.; Slawecki, C.J.; El Khoury, A.; Mathe, A.A.; Ehlers, C.L. The effects of social isolation on neuropeptide Y levels, exploratory and anxiety-related behaviors in rats. Pharmacol. Biochem. Behav. 2006, 83, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Westenbroek, C.; Ter Horst, G.J.; Roos, M.H.; Kuipers, S.D.; Trentani, A.; den Boer, J.A. Gender-specific effects of social housing in rats after chronic mild stress exposure. Progress. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Workman, J.L.; Trainor, B.C.; Finy, M.S.; Nelson, R.J. Effects of neuronal nitric oxide synthase and social housing of anxiety-like behavior. Behav. Brain Res. 2008, 187, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Ruis, M.A.W.; te Brake, J.H.A.; Buwalda, B.; De Boer, S.F.; Meerlo, P.; Korte, S.M.; Blokhuis, H.J.; Koolhaas, J.M. Housing familiar male wildtype rats together reduces the long-term adverse behavioural and physiological effects of social defeat. Psychoneuroendocrinology 1999, 24, 285–300. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoicas, I.; Mühle, C.; Schumacher, F.; Kleuser, B.; Kornhuber, J. Development of Comorbid Depression after Social Fear Conditioning in Mice and Its Effects on Brain Sphingolipid Metabolism. Cells 2023, 12, 1355. https://doi.org/10.3390/cells12101355
Zoicas I, Mühle C, Schumacher F, Kleuser B, Kornhuber J. Development of Comorbid Depression after Social Fear Conditioning in Mice and Its Effects on Brain Sphingolipid Metabolism. Cells. 2023; 12(10):1355. https://doi.org/10.3390/cells12101355
Chicago/Turabian StyleZoicas, Iulia, Christiane Mühle, Fabian Schumacher, Burkhard Kleuser, and Johannes Kornhuber. 2023. "Development of Comorbid Depression after Social Fear Conditioning in Mice and Its Effects on Brain Sphingolipid Metabolism" Cells 12, no. 10: 1355. https://doi.org/10.3390/cells12101355
APA StyleZoicas, I., Mühle, C., Schumacher, F., Kleuser, B., & Kornhuber, J. (2023). Development of Comorbid Depression after Social Fear Conditioning in Mice and Its Effects on Brain Sphingolipid Metabolism. Cells, 12(10), 1355. https://doi.org/10.3390/cells12101355