
Microgravity Effects on the Matrisome


Abstract
:1. Introduction
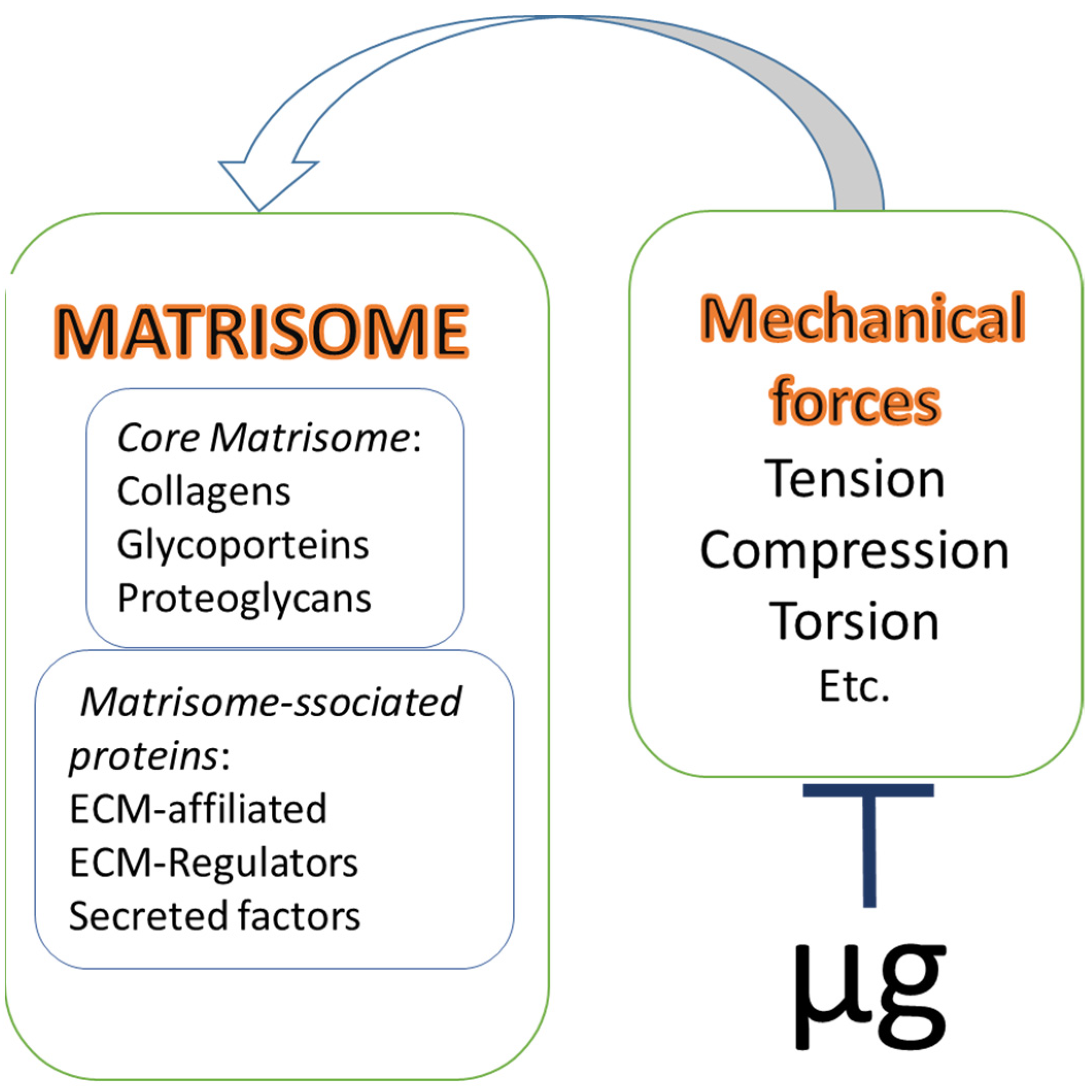
2. Current Concept of the Extracellular Matrix as a Complex of Structural and Regulatory Molecules: Matrisome
3. Matrisome of Connective Tissues
4. Connective Tissue Matrisome and Microgravity
4.1. Space Flight Experiments
4.2. Ground-Based Simulations
5. Proteomic Profile of Human Matrisome-Associated Proteins under Real and Simulated Microgravity
6. Concluding Remarks and Further Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Parra, M.; Jung, J.; Boone, T.D.; Tran, L.; Blaber, E.A.; Brown, M.; Chin, M.; Chinn, T.; Cohen, J.; Doebler, R.; et al. Microgravity validation of a novel system for RNA isolation and multiplex quantitative real time PCR analysis of gene expression on the International Space Station. PLoS ONE 2017, 12, e0183480. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Pietsch, J.; Wehland, M.; Richter, P.; Strauch, S.M.; Lebert, M.; Magnusson, N.E.; Wise, P.; Bauer, J. The impact of microgravity-based proteomics research. Expert Rev. Proteom. 2014, 11, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Buken, C.; Sahana, J.; Corydon, T.J.; Melnik, D.; Bauer, J.; Wehland, M.; Krüger, M.; Balk, S.; Abuagela, N.; Infanger, M.; et al. Morphological and molecular changes in juvenile normal human fibroblasts exposed to simulated microgravity. Sci. Rep. 2019, 9, 11882. [Google Scholar] [CrossRef] [PubMed]
- Argentati, C.; Morena, F.; Tortorella, I.; Bazzucchi, M.; Porcellati, S.; Emiliani, C.; Martino, S. Insight into mechanobiology: How stem cells feel mechanical forces and orchestrate biological functions. Int. J. Mol. Sci. 2019, 20, 5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, I.D.; Humphries, M.J. Integrin structure, activation, and interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingber, D.E. Tensegrity: The architectural basis of cellular mechanotransduction. Ann. Rev. Physiol. 1997, 59, 575–599. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Melnik, D.; Corydon, T.J.; Sahana, J.; Krüger, M.; Wehland, M.; Bauer, T.J.; Liemersdorf, C.; Hemmersbach, R.; et al. Short-term microgravity influences cell adhesion in human breast cancer cells. Int. J. Mol. Sci. 2019, 20, 5730. [Google Scholar] [CrossRef] [Green Version]
- Oganov, V.S. Modern analysis of bone loss mechanisms in microgravity. J. Gravit. Physiol. 2004, 11, 143–146. [Google Scholar]
- Sibonga, J.D.; Evans, H.J.; Sung, H.G.; Spector, E.R.; Lang, T.F.; Oganov, V.S.; Bakulin, A.V.; Shackelford, L.C.; LeBlanc, A.D. Recovery of spaceflight-induced bone loss: Bone mineral density after long-duration missions as fitted with an exponential function. Bone 2007, 41, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced expression of cytoskeletal and extracellular matrix genes in human adult retinal pigment epithelium cells exposed to simulated microgravity. Cell Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Chatziravdeli, V.; Katsaras, G.N.; Lambrou, G.I. Gene expression in osteoblasts and osteoclasts under microgravity conditions: A systematic review. Curr. Genom. 2019, 20, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Bloom, W.; Fawcett, D.W. A Textbook of Histology, 12th ed.; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma 2014, 23, 20–23. [Google Scholar] [CrossRef]
- Vogel, V. Unraveling the mechanobiology of extracellular matrix. Annu. Rev. Physiol. 2018, 80, 353–387. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.R.; Kleinman, H.K.; Terranova, V.P.; Ledbetter, S.; Hassell, J.R. The regulation of basement membrane formation and cell-matrix interactions by defined supramolecular complexes. Ciba Found. Symp. 1984, 108, 197–212. [Google Scholar] [CrossRef]
- Hynes, R.O.; Naba, A. Overview of the matrisome—An inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [Green Version]
- Naba, A.; Clauser, K.R.; Hoersch, S.; Liu, H.; Carr, S.A.; Hynes, R.O. The matrisome: In silico definition and in vivo characterization by proteomics of normal and tumor extracellular matrices. Mol. Cell Proteom. 2012, 11, M111.014647. [Google Scholar] [CrossRef] [Green Version]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- Myllyharju, J.; Kivirikko, K.I. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar] [CrossRef]
- Chung, M.I.; Miao, M.; Stahl, R.J.; Chan, E.; Parkinson, J.; Keeley, F.W. Sequences and domain structures of mammalian, avian, amphibian, and teleost tropoelastins: Clues to the evolutionary history of elastin. Matrix Biol. 2006, 25, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Kadler, E.; Baldock, C.; Bella, J.; Raymond, P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Derm.-Endocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Murphy-Ullrich, J.E.; Iozzo, R.V. Thrombospondins in physiology and disease: New tricks for old dogs. Matrix Biol. 2012, 31, 152–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halper, M.J.; Kjaer, M. Basic components of connective tissues and extracellular matrix: Elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. Adv. Exp. Med. Biol. 2014, 802, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [Green Version]
- Parisi, L.; Toffoli, A.; Ghezzi, B.; Mozzoni, B.; Lumetti, S.; Macaluso, G.M. A glance on the role of fibronectin in controlling cell response at biomaterial interface. Jpn. Dent. Sci. Rev. 2020, 56, 50–55. [Google Scholar] [CrossRef]
- Corsi, X.T.; Chen, X.D.; Boyde, A.; Liang, J.; Mankani, M.; Sommer, B.; Iozzo, R.V.; Eichstetter, I.; Robey, P.G.; Bianco, P.; et al. Phenotypic effects of biglycan deficiency are linked to collagen fibril and mimic Ehlers-Danlos-like changes in bone and other connective abnormalities, are synergized by decorin deficiency, tissues. J. Am. Soc. Bone Miner. 2002, 17, 1180–1189. [Google Scholar] [CrossRef]
- Halasz, K.; Kassner, A.; Morgelin, M.; Heinegård, D. COMP as a catalyst in collagen fi brillogenesis. J. Biol. Chem. 2007, 282, 31166–31173. [Google Scholar] [CrossRef] [Green Version]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mole. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Daley, W.P.; Peters, S.B.; Larsen, M. Extracellular matrix dynamics in development and regenerative medicine. J. Cell Sci. 2008, 121, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta 2014, 1840, 2506–2519. [Google Scholar] [CrossRef]
- Rahman, S.; Patel, Y.; Murray, J.; Patel, K.V.; Sumathipala, R.; Sobel, M.; Wijelath, E.S. Novel hepatocyte growth factor (HGF) binding domains on fibronectin and vitronectin coordinate a distinct and amplified Met-integrin induced signalling pathway in endothelial cells. BMC Cell Biol. 2005, 6, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Martino, M.M.; Brkic, S.; Bovo, E.; Burger, M.; Schaefer, D.J.; Wolff, T.; Gürke, L.; Briquez, P.S.; Larsson, H.M.; Gianni-Barrera, R.; et al. Extracellular matrix and growth factor engineering for controlled angiogenesis in regenerative medicine. Front. Bioeng. Biotechnol. 2015, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, F.; Rifkin, D.B. Extracellular microfibrils: Contextual platforms for TGFbeta and BMP signaling. Curr. Opin. Cell Biol. 2009, 21, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Munger, J.S.; Sheppard, D. Cross talk among TGF-β signaling pathways, integrins, and the extracellular matrix. Cold Spring Harb. Perspect. Biol. 2013, 3, a005017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Lin, X. Shaping morphogen gradients by proteoglycans. Cold Spring Harb. Perspect. Biol. 2009, 1, a002493. [Google Scholar] [CrossRef]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Ramage, L. Integrins and extracellular matrix in mechanotransduction. Cell Health Cytoskelet. 2011, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Raghunathan, R.; Sethi, M.K.; Klein, J.A.; Zaia, J. Proteomics, glycomics, and glycoproteomics of matrisome molecules. Mol. Cell Proteom. 2019, 18, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Taha, I.N.; Clauser, K.R.; Gao, Y.T.; Naba, A. MatrisomeDB: The ECM-protein knowledge database. Nucleic Acids Res. 2020, 48, D1136–D1144. [Google Scholar] [CrossRef] [PubMed]
- Badhwar, A.; Stanimirovic, D.B.; Hamel, E.; Haqqani, A.S. The proteome of mouse cerebral arteries. J. Cereb. Blood Flow Metab. 2014, 34, 1033–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharaz, Y.A.; Canty-Laird, E.G.; Tew, S.R.; Comerford, E.J. Variations in internal structure, composition and protein distribution between intra- and extra-articular knee ligaments and tendons. J. Anat. 2018, 232, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, K.R.; Saleh, A.M.; Sarah, N.; Lipp, S.N.; Ocken, A.R.; Kinzer-Ursem, T.L.; Calve, S. Extracellular matrix protein composition dynamically changes during murine forelimb development. BioRxiv 2020. [Google Scholar] [CrossRef]
- Li, M.; Zhang, A.; Li, J.; Zhou, J.; Zheng, Y.; Zhang, C.; Xia, D.; Mao, H.; Zhao, J. Osteoblast/fibroblast coculture derived bioactive ECM with unique matrisome profile facilitates bone regeneration. Bioact. Mater. 2020, 5, 938–948. [Google Scholar] [CrossRef]
- Frost, H.M. Wolff’s Law and bone’s structural adaptations to mechanical usage: An overview for clinicians. Angle Orthod. 1994, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhu, D.; Gao, J.; Linwei, L.V.; Zhang, X. An adaptation model for trabecular bone at different mechanical levels. Biomed. Eng. Online 2010, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Mammoto, T.; Ingber, D.E. Mechanical control of tissue and organ development. Development 2010, 137, 1407–1420. [Google Scholar] [CrossRef] [Green Version]
- Bogdanowicz, D.R.; Lu, H.H. Designing the stem cell microenvironment for guided connective tissue regeneration. Ann. N. Y. Acad. Sci. 2017, 1410, 3–25. [Google Scholar] [CrossRef]
- Weaver, V.M. Cell and tissue mechanics: The new cell biology frontier. Mol. Biol. Cell. 2017, 28, 1815–1818. [Google Scholar] [CrossRef] [Green Version]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [Green Version]
- Vico, L.; Collet, P.; Guignandon, A.; Lafage-Proust, M.H.; Thomas, T.; Rehaillia, M.; Alexandre, C. Effects of long-term microgravity exposure on cancellous and cortical weight-bearing bones of cosmonauts. Lancet 2000, 355, 1607–1611. [Google Scholar] [CrossRef]
- Neutelings, T.; Nusgens, B.V.; Liu, Y.; Tavella, S.; Ruggiu, A.; Cancedda, R.; Gabriel, M.; Colige, A.; Lambert, C. Skin physiology in microgravity: A 3-month stay aboard ISS induces dermal atrophy and affects cutaneous muscle and hair follicles cycling in mice. NPJ Microgravity 2015, 1, 15002. [Google Scholar] [CrossRef] [Green Version]
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human pathophysiological adaptations to the space environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Chatani, M.; Morimoto, H.; Takeyama, K.; Mantoku, A.; Tanigawa, N.; Kubota, K.; Suzuki, H.; Uchida, S.; Tanigaki, F.; Shirakawa, M.; et al. Acute transcriptional up-regulation specific to osteoblasts/osteoclasts in medaka fish immediately after exposure to microgravity. Sci. Rep. 2016, 6, 39545. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, G.; Nys, G.; Bouillon, R. Microgravity reduces the differentiation of human osteoblastic MG-63 cells. J. Bone Miner. Res. 1997, 12, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D.; Harris, J.; Halloran, B.P.; Morey-Holton, E. Altered skeletal pattern of gene expression in response to spaceflight and hindlimb elevation. Am. J. Physiol. 1994, 267, E822–E827. [Google Scholar] [CrossRef] [PubMed]
- Landis, W.J.; Hodgens, K.J.; Block, D.; Toma, C.D.; Gerstenfeld, L.C. Spaceflight effects on cultured embryonic chick bone cells. J. Bone Miner. Res. 2000, 15, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Kumei, Y.; Shimokawa, H.; Ohya, K.; Katano, H.; Akiyama, H.; Hirano, M.; Morita, S. Small GTPase Ras and Rho expression in rat osteoblasts during spaceflight. Ann. N. Y. Acad. Sci. 2007, 1095, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, L.; Jiang, Y.; Wang, C.; Geng, B.; Wang, Y.; Chen, J.; Liu, F.; Qiu, P.; Zhai, G.; et al. Space microgravity drives transdifferentiation of human bone marrow-derived mesenchymal stem cells from osteogenesis to adipogenesis. FASEB J. 2018, 32, 4444–4458. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Yang, J.; Guo, C.; Shi, D.; Shen, D.; Zheng, Q.; Chen, R.; Xu, Y.; Xi, Y.; Wang, J. Effects of hindlimb unloading on ex vivo growth and osteogenic/adipogenic potentials of bone marrow-derived mesenchymal stem cells in rats. Stem Cells Dev. 2008, 17, 795–804. [Google Scholar] [CrossRef]
- Ishijima, M.; Tsuji, K.; Rittling, S.R.; Yamashita, T.; Kurosawa, H.; Denhardt, D.T.; Nifuji, A.; Ezura, Y.; Noda, M. Osteopontin is required for mechanical stress-dependent signals to bone marrow cells. J. Endocrinol. 2007, 193, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Ferranti, F.; Del Bianco, M.; Pacelli, C. Advantages and limitations of current microgravity platforms for space biology research. Appl. Sci. 2021, 11, 68. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayzafoon, M.; Gathings, W.E.; McDonald, J.M. Modeled microgravity inhibits osteogenic differentiation of human mesenchymal stem cells and increases adipogenesis. Endocrinology 2004, 145, 2421–2432. [Google Scholar] [CrossRef] [Green Version]
- Bucaro, M.A.; Zahm, A.M.; Risbud, M.V.; Ayyaswamy, P.S.; Mukundakrishnan, K.; Steinbeck, M.J.; Shapiro, I.M.; Adams, C.S. The effect of simulated microgravity on osteoblasts is independent of the induction of apoptosis. J. Cell Biochem. 2007, 102, 483–495. [Google Scholar] [CrossRef]
- Ontiveros, C.; McCabe, L.R. Simulated microgravity suppresses osteoblast phenotype, runx2 levels and AP-1 transactivation. J. Cell Biochem. 2003, 88, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; Smith, C.L.; Weigel, N.L. Vector-averaged gravity-induced changes in cell signaling and vitamin D receptor activity in MG-63 cells are reversed by a 1,25-(OH)2D3 analog, EB. Bone 2002, 31, 381–388. [Google Scholar] [CrossRef]
- Mayer-Wagner, S.; Hammerschmid, F.; Redeker, J.I.; Schmitt, B.; Holzapfel, B.M.; Jansson, V.; Betz, O.B.; Müller, P.E. Simulated microgravity affects chondrogenesis and hypertrophy of human mesenchymal stem cells. Int. Orthop. 2014, 38, 2615–2621. [Google Scholar] [CrossRef]
- Rucci, N.; Rufo, A.; Alamanou, M.; Teti, A. Modeled microgravity stimulates osteoclastogenesis and bone resorption by increasing osteoblast RANKL/OPG ratio. J. Cell. Biochem. 2007, 100, 464–473. [Google Scholar] [CrossRef]
- Ratushnyy, A.Y.; Buravkova, L.B. Expression of focal adhesion genes in mesenchymal stem cells under simulated microgravity. Dokl. Biochem. Biophys. 2017, 477, 354–356. [Google Scholar] [CrossRef]
- Zhivodernikov, I.V.; Ratushnyy, A.Y.; Matveeva, D.K.; Buravkova, L.B. Extracellular matrix proteins and transcription of matrix-associated genes in mesenchymal stromal cells during modeling of the effects of microgravity. Bull. Exp. Biol. Med. 2020, 170, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Buravkova, L.B.; Gershovich, P.M.; Gershovich, J.G.; Grigorev, A.I. Mechanisms of gravitational sensitivity of osteogenic precursor cells. Acta Nat. 2010, 2, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Gershovich, P.M.; Gershovich, J.G.; Zhambalova, A.P.; Romanov, Y.A.; Buravkova, L.B. Cytoskeletal proteins and stem cell markers gene expression in human bone marrow mesenchymal stromal cells after different periods of simulated microgravity. Acta Astronaut. 2012, 70, 36–42. [Google Scholar] [CrossRef]
- Pardo, S.J.; Patel, M.J.; Sykes, M.C.; Platt, M.O.; Boyd, N.L.; Sorescu, G.P.; Xu, M.; van Loon, J.J.; Wang, M.D.; Jo, H. Simulated microgravity using the Random Positioning Machine inhibits differentiation and alters gene expression profiles of 2T3 preosteoblasts. Am. J. Physiol. Cell Physiol. 2005, 288, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Soshi, S.; Fujii, K. Effect of hyper- and microgravity on collagen post-translational controls of MC3T3-E1 osteoblasts. J. Bone Miner. Res. 2003, 18, 1695–1705. [Google Scholar] [CrossRef]
- Ebnerasuly, F.; Hajebrahimi, Z.; Tabaie, S.M.; Darbouy, M. Simulated microgravity condition alters the gene expression of some ECM and adhesion molecules in adipose derived stem cells. Int. J. Mol. Cell Med. 2018, 7, 146–157. [Google Scholar] [CrossRef]
- Makihira, S.; Kawahara, Y.; Yuge, L.; Mine, Y.; Nikawa, H. Impact of the microgravity environment in a 3-dimensional clinostat on osteoblast- and osteoclast-like cells. Cell Biol. Int. 2008, 32, 1176–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.Q.; Wang, R.; Ling, S.K.; Wan, Y.M.; Li, Y.H. Simulated microgravity inhibits the proliferation and osteogenesis of rat bone marrow mesenchymal stem cells. Cell Prolif. 2007, 40, 671–684. [Google Scholar] [CrossRef]
- Rea, G.; Cristofaro, F.; Pani, G.; Pascucci, B.; Ghuge, S.A.; Corsetto, P.A.; Imbriani, M.; Visai, L.; Rizzo, A.M. Microgravity-driven remodeling of the proteome reveals insights into molecular mechanisms and signal networks involved in response to the space flight environment. J. Proteom. 2016, 137, 3–18. [Google Scholar] [CrossRef]
- Prasad, B.; Grimm, D.; Strauch, S.M.; Erzinger, G.S.; Corydon, T.J.; Lebert, M.; Magnusson, N.E.; Infanger, M.; Richter, P.; Krüger, M. Influence of microgravity on apoptosis in cells, tissues, and other systems in vivo and in vitro. Int. J. Mol. Sci. 2020, 21, 9373. [Google Scholar] [CrossRef]
- Strauch, S.M.; Grimm, D.; Corydon, T.J.; Krüger, M.; Bauer, J.; Lebert, M.; Wise, P.; Infanger, M.; Richter, P. Current knowledge about the impact of microgravity on the proteome. Exp. Rev. Proteom. 2019, 16, 5–16. [Google Scholar] [CrossRef]
- Kononikhin, A.S.; Starodubtseva, N.L.; Pastushkova, L.K.; Kashirina, D.N.; Fedorchenko, K.Y.; Brhozovsky, A.G.; Popov, I.A.; Larina, I.M.; Nikolaev, E.N. Spaceflight induced changes in the human proteome. Exp. Rev. Proteom. 2017, 14, 15–29. [Google Scholar] [CrossRef]
- Larina, I.; Pastushkova, L.; Kononikhin, A.; Nikolaev, E.; Orlov, O. Piloted space flight and post-genomic technologies. Reach 2019, 16, 10003. [Google Scholar] [CrossRef]
- Taha, I.N.; Naba, A. Exploring the extracellular matrix in health and disease using proteomics. Essays Biochem. 2019, 63, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Randles, M.J.; Humphries, M.J.; Lennon, R. Proteomic definitions of basement membrane composition in health and disease. Matrix Biol. 2017, 57–58, 12–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barallobre-Barreiro, J.; Lynch, M.; Yin, X.; Mayr, M. Systems biology-opportunities and challenges: The application of proteomics to study the cardiovascular extracellular matrix. Cardiovasc. Res. 2016, 112, 626–636. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, M.L.; Jung, M.; Hall, M.E.; DeLeon-Pennell, K.Y. Proteomic analysis of the cardiac extracellular matrix: Clinical research applications. Expert Rev. Proteom. 2018, 15, 105–112. [Google Scholar] [CrossRef]
- Socovich, A.M.; Naba, A. The cancer matrisome: From comprehensive characterization to biomarker discovery. Semin. Cell Dev. Biol. 2019, 89, 157–166. [Google Scholar] [CrossRef]
- Randles, M.; Lennon, R. Applying proteomics to investigate extracellular matrix in health and disease. Curr. Top. Membr. 2015, 76, 171–196. [Google Scholar] [CrossRef]
- Elagamey, E.; Narula, K.; Chakraborty, N.; Chakraborty, S. Extracellular matrix proteome: Isolation of ECM proteins for proteomics studies. Meth. Mol. Biol. 2020, 2057, 155–172. [Google Scholar] [CrossRef]
- Langley, S.R.; Willeit, K.; Didangelos, A.; Matic, L.P.; Skroblin, P.; Barallobre-Barreiro, J.; Lengquist, M.; Rungger, G.; Kapustin, A.; Kedenko, L.; et al. Extracellular matrix proteomics identifies molecular signature of symptomatic carotid plaques. J. Clin. Investig. 2017, 127, 1546–1560. [Google Scholar] [CrossRef] [PubMed]
- Kozlovskaya, I.B. Fundamental and applied problems of immersion research. Aviakosm. Ekol. Med. 2008, 42, 3–7. [Google Scholar]
- Linossier, M.T.; Amirova, L.E.; Thomas, M.; Normand, M.; Bareille, M.; Gauquelin-Koch, G.; Beck, A.; Bonneau, C.; Gharib, C.; Custaud, M.; et al. Effects of short-term dry immersion on bone remodeling markers, insulin and adipokines. PLoS ONE 2017, 12, e0182970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzhozovskiy, A.G.; Kononikhin, A.S.; Pastushkova, L.C.; Kashirina, D.N.; Indeykina, M.I.; Popov, I.A.; Custaud, M.A.; Larina, I.M.; Nikolaev, E.N. The Effects of Spaceflight Factors on the Human Plasma Proteome, Including Both Real Space Missions and Ground-Based Experiments. Int. J. Mol. Sci. 2019, 20, 3194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liphardt, A.M.; Mündermann, A.; Koo, S.; Bäcker, N.; Andriacchi, T.P.; Zange, J.; Mester, J.; Heer, M. Vibration training intervention to maintain cartilage thickness and serum concentrations of cartilage oligometric matrix protein (COMP) during immobilization. Osteoarthr. Cartil. 2009, 17, 1598–1603. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Hu, X.; Zhang, X.; Duan, X.; Peng Yang, P.; Zhao, F.; Ao, Y. Effects of mechanical stress on chondrocyte phenotype and chondrocyte extracellular matrix expression. Sci. Rep. 2016, 6, 37268. [Google Scholar] [CrossRef] [Green Version]
- Kashirina, D.N.; Brzhozovskiy, A.G.; Pastushkova, L.K.; Kononikhin, A.S.; Borchers, C.H.; Nikolaev, E.N.; Larina, I.M. Semiquantitative proteomic research of protein plasma profile of volunteers in 21-day head-down bed rest. Front. Physiol. 2020, 11, 678. [Google Scholar] [CrossRef]
- da Silveira, W.A.; Fazelinia, H.; Rosenthal, S.B.; Laiakis, E.C.; Kim, M.S.; Meydan, C.; Kidane, Y.; Rathi, K.S.; Smith, S.M.; Stear, B. Comprehensive multi-omics analysis reveals mitochondrial stress as a central biological hub for spaceflight impact. Cell 2020, 183, 1185–1201. [Google Scholar] [CrossRef]
- Pakharukova, N.A.; Pastushkova, L.K.; Samarin, G.I.; Pochuev, V.I.; Morukov, B.V.; Larina, I.M. Direct proteomic profiling of blood serum of cosmonauts after long-term space flights. Hum. Physiol. 2014, 40, 713. [Google Scholar] [CrossRef]
- Larina, I.M.; Percy, A.J.; Yang, J.; Borchers, C.H.; Nosovsky, A.M.; Grigoriev, A.I.; Nikolaev, E.N. Protein expression changes caused by spaceflight as measured for 18 Russian cosmonauts. Sci. Rep. 2019, 9, 8570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latridis, J.C.; MacLean, J.J.; Roughley, P.J.; Alini, M. Effects of mechanical loading on intervertebral disc metabolism in vivo. J. Bone Jt. Surg. Am. 2006, 88, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Pastushkova, L.; Kireev, K.S.; Kononikhin, A.S.; Tiys, E.S.; Popov, I.A.; Starodubtseva, N.L.; Dobrokhotov, I.V.; Ivanisenko, V.A.; Larina, I.M.; Kolchanov, N.A.; et al. Detection of renal tissue and urinary tract proteins in the human urine after space flight. PLoS ONE 2013, 8, e71652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanczyk, M.; Layland, S.L.; Schenke-Layland, K. The role of extracellular matrix in biomechanics and its impact on bioengineering of cells and 3D tissues. Matrix Biol. 2020, 85–86, 1–14. [Google Scholar] [CrossRef]
- Po, A.; Giuliani, A.; Masiello, M.G.; Cucina, A.; Catizone, A.; Ricci, G.; Chiacchiarini, M.; Tafani, M.; Ferretti, E.; Bizzarri, M. Phenotypic transitions enacted by simulated microgravity do not alter coherence in gene transcription profile. NPJ Microgravity 2019, 5, 27. [Google Scholar] [CrossRef]
- Monti, N.; Masiello, M.G.; Proietti, S.; Catizone, A.; Ricci, G.; Harrath, A.H.; Alwasel, S.H.; Cucina, A.; Bizzarri, M. Survival pathways are differently affected by microgravity in normal and cancerous breast cells. Int. J. Mol. Sci. 2021, 22, 862. [Google Scholar] [CrossRef]
- Jiang, M.; Qiu, J.; Zhang, L.; Lü, D.; Long, M.; Chen, L.; Luo, X. Changes in tension regulates proliferation and migration of fibroblasts by remodeling expression of ECM proteins. Exp. Ther. Med. 2016, 12, 1542–1550. [Google Scholar] [CrossRef] [Green Version]
- Uddin, S.M.; Qin, Y.X. Enhancement of osteogenic differentiation and proliferation in human mesenchymal stem cells by a modified low intensity ultrasound stimulation under simulated microgravity. PLoS ONE 2013, 8, e73914. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Yasuda, T.; Watanabe-Asaka, T.; Oda, S.; Mantoku, A.; Takeyama, K.; Chatani, M.; Kudo, A.; Uchida, S.; Suzuki, H.; et al. Histological and transcriptomic analysis of adult Japanese Medaka sampled onboard the international space station. PLoS ONE 2015, 10, e138799. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Object | Cell Type | Duration | Matrisome | Exp. Approach | Study | ||
|---|---|---|---|---|---|---|---|
| Core Matrisome | ECM-Associated Molecules | ||||||
| Collagens | Proteoglycans, Glycoproteins | ||||||
| Space flight | |||||||
| Medaka fish | OB, Ocl (ISS) | 1, 5, 8 d | U: COL10A1 | U: OCN | U: MMP9 | DsRed live-imaging, GGA | [58] |
| Mouse | MG-63, osteoinduced (Foton 10) | 9 d | D: COL1A; ND: collagen type I | D: OCN | RT-PCR, WB | [59] | |
| Mouse | 2T3 OB (STS-54) | 6 d | D: OCN | RT-PCR | [60] | ||
| Chicken | OB (STS-59) | 12 d | D: COL1A; ND: collagen type I | D: OCN | RT-PCR, WB | [61] | |
| Rat | OB (STS-65) | 5 d | D: OCN | RT-PCR | [62] | ||
| Human | OB, MSC-derived (SJ-10) | 2 d | D: COL1A1, -1A2, -3A1, -4A1, -5A1, -6A1, -8A1 | D: TNC, COMP | U: MMP1 | RT-PCR | [63] |
| Human | OB, MSC-derived (SJ-10) | 5 d | D: COL1A1 | RT-PCR | [63] | ||
| Hind-limb suspension (HS) | |||||||
| Rat | MSC-BM, tibia | 5 d | D: OCN | RT-PCR | [60] | ||
| Rat | MSC-BM, tibia | 14 d | U: OCN | RT-PCR | [60] | ||
| Rat | MSC-BM, femur | 28 d | D: RUNX2, COLI, ALP, OCN | D: osteogenic potential; expression of osteoblast gene marker mRNAs under osteogenic conditions. | RT-PCR, HS | [64] | |
| Mouse | MSC-BM, femur | 7 d | D: mineralization | HS | [65] | ||
| Rotating-Wall Vessel (RWV) | |||||||
| Human | MSC-BM | 7 d | D: COL2A1, COL10A1 | D: ON | RT-PCR | [68] | |
| Mouse | OB MC3T3-E1 | 1–14 d | ND: COL1A2 | ND: OCN | RT-PCR | [68] | |
| Mouse | OB/OC MLO-A5 | 1–14 d | ND: COL1A3 | ND: OCN, OPN | RT-PCR | [69] | |
| Mouse | MSC C3H10T(1/2) | 1–14 d | ND: OCN, OPN, COL1A4 | ND: OCN, OPN | RT-PCR | [69] | |
| Mouse | OB MC3T3-E1 | 1 d | D: OCN | RT-PCR | [70] | ||
| Human | MG-63, osteoinduced | 3 d | D: COL1 | D: OCN | RT-PCR | [71] | |
| Human | ChB, MSC-derived | 21 d | D: COL1 | D: AGN | RT-PCR | [72] | |
| Mouse | OB | 1d | ND: COL1A2 | ND: OCN, OPN | RT-PCR | [73] | |
| Random Positioning Machine (RPM) | |||||||
| Human | FB | 3 d | U: COL4A5 | U: FN/FN | U: TGF | RT-PCR, WB | [3] |
| Human | MSC-AT | 4 d | U: COL12A1, COL15A1, COL16A1, COL1A1, COL5A1, COL8A1 | U: THBS1, THBS2, THBS3, LAMA, SPARC, TNC, VCAN, VTN; D: CLEC3B | RT-PCR | [74] | |
| Human | MSC-AT | 10 d | D: COL11A1; D: collagenous proteins | D: LAMB3, TNC; U: non-collagenous proteins | RT-PCR, HC | [75] | |
| Human | MSC-BM, osteoinduced | 20 d | U: COL1A1 | D: OMD; ND: OCN | D: ECM mineralization | RT-PCR, HC | [76] |
| Human | MSC-BM | 5 d | D: COL9A1, COL2A1 | RT-PCR | [77] | ||
| Human | MSC-BM, osteoinduced | 10 d | ND: COL1A1 | D: OMD; ND: OCN | RT-PCR, ICC | 77] | |
| Human | MSC-BM | 20 d | ND: COL1A1; ND: collagen tot | RT-PCR, ICC | [77] | ||
| Human | OB | 20 d | U: ECM mineralization | HC | [77] | ||
| Mouse | 2T3 OB | 3 d | D: OMD | RT-PCR | [78] | ||
| 2D, 3D-clinorotation | |||||||
| Mouse | MC3T3-E2 | 3 d | U: PLOD1, PLOD2; U: enzymes activity | RT-PCR, enzyme assay | [79] | ||
| Human | MSC-AT | 7 d | U: COL1 COL3 | D: FBN1 | D: MMP1 | RT-PCR | [80] |
| Mouse | MC3T3-E1 | 7 d | D: COL1A1 | RT-PCR | [81] | ||
| Rat | MSC | 1–4 d | D: cbfa1/RUNX2 | RT-PCR | [82] | ||
| Protein | Uniprot Index | Biological Process (ANDCell) |
|---|---|---|
| Alpha-2-HS-glycoprotein | FETUA_HUMAN | Positive regulation of ECM constituent secretion; Regulation of ECM assembly; erk 1/2 mitogen-activated protein kinase pathway ANG 2; ANG2 expression of ECM proteins; ANG2 erk1/2 pathway; mek/erk pathway; erk pathway |
| Angiotensinogen | ANGT_HUMAN | erk pathway |
| Apolipoprotein A-I | APOA1_HUMAN | erk pathway |
| Apolipoprotein E | APOE_HUMAN | Positive regulation of ECM constituent secretion |
| Carboxypeptidase B2 | CBPB2_HUMAN | erk pathway |
| Cathelicidin antimicrobial peptide | CAMP_HUMAN | ras-erk pathway |
| CD44 antigen | CD44_HUMAN | MMP9 signaling pathway; ras-erk1/2 pathway; mek/erk pathway; erk pathway. |
| Clusterin | CLUS_HUMAN | Inhibition of ECM disassembly; ECM organization; mapk/erk pathway; MMP9 signaling pathway |
| Cystatin-C | CYTC_HUMAN | ECM organization |
| Fibronectin | FINC_HUMAN | ECM organization; Activation of erk pathway; ECM assembly; erk1/2 pathway; Cell–matrix adhesion; erk pathway; Calcium independent cell matrix adhesion; mapk/erk pathway |
| Fibulin-1 | FBLN1_HUMAN | Cell–matrix adhesion |
| Insulin-like growth factor-binding protein 3 | IBP3_HUMAN | erk1/2 pathway |
| Intercellular adhesion molecule 1 | ICAM1_HUMAN | erk1/2 pathway; mek/erk pathway; Cell–matrix adhesion |
| Kininogen-1 | KNG1_HUMAN | Bradykinin in MMP secretion; ECM secretion; mapk/erk pathway |
| Lumican | LUM_HUMAN | ECM assembly |
| Pigment epithelium-derived factor | PEDF_HUMAN | MMP secretion; apoptotic signaling pathway; erk1/2 pathway |
| Transthyretin | TTHY_HUMAN | Apoptotic signaling pathway; erk1/2 pathway |
| Vitronectin | VTNC_HUMAN | ECM organization; Cell–matrix adhesion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buravkova, L.; Larina, I.; Andreeva, E.; Grigoriev, A. Microgravity Effects on the Matrisome. Cells 2021, 10, 2226. https://doi.org/10.3390/cells10092226
Buravkova L, Larina I, Andreeva E, Grigoriev A. Microgravity Effects on the Matrisome. Cells. 2021; 10(9):2226. https://doi.org/10.3390/cells10092226
Chicago/Turabian StyleBuravkova, Ludmila, Irina Larina, Elena Andreeva, and Anatoly Grigoriev. 2021. "Microgravity Effects on the Matrisome" Cells 10, no. 9: 2226. https://doi.org/10.3390/cells10092226
APA StyleBuravkova, L., Larina, I., Andreeva, E., & Grigoriev, A. (2021). Microgravity Effects on the Matrisome. Cells, 10(9), 2226. https://doi.org/10.3390/cells10092226