
Morpho-Physiological and Transcriptome Changes in Tomato Anthers of Different Developmental Stages under Drought Stress



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Phenotypic and Fruit Yield Characterization
2.3. Assessment of Gametophyte Fertility
 × DS
× DS  ); and (4) stigmas of flowers on DS plants cross-pollinated with pollen grains from WW plants (DS × WW ). The emasculation of female flowers was performed 1 d to anthesis and hand-pollinated the following evening using anthers from flowers that opened on the same day as a pollen source.
); and (4) stigmas of flowers on DS plants cross-pollinated with pollen grains from WW plants (DS × WW ). The emasculation of female flowers was performed 1 d to anthesis and hand-pollinated the following evening using anthers from flowers that opened on the same day as a pollen source.2.4. Histo-Cytological Studies
2.5. Transcriptome Analysis
2.5.1. Samples for RNA-Sequencing
2.5.2. Preparation of RNA-Seq Library and Sequencing
2.5.3. Analysis of RNA-Seq Data
2.6. Validation of RNA-Seq Data
2.7. Extraction and Quantification of Sucrose, Fructose and Glucose
2.8. Phytohormones Extraction and Quantification
2.9. Data Analysis
3. Results
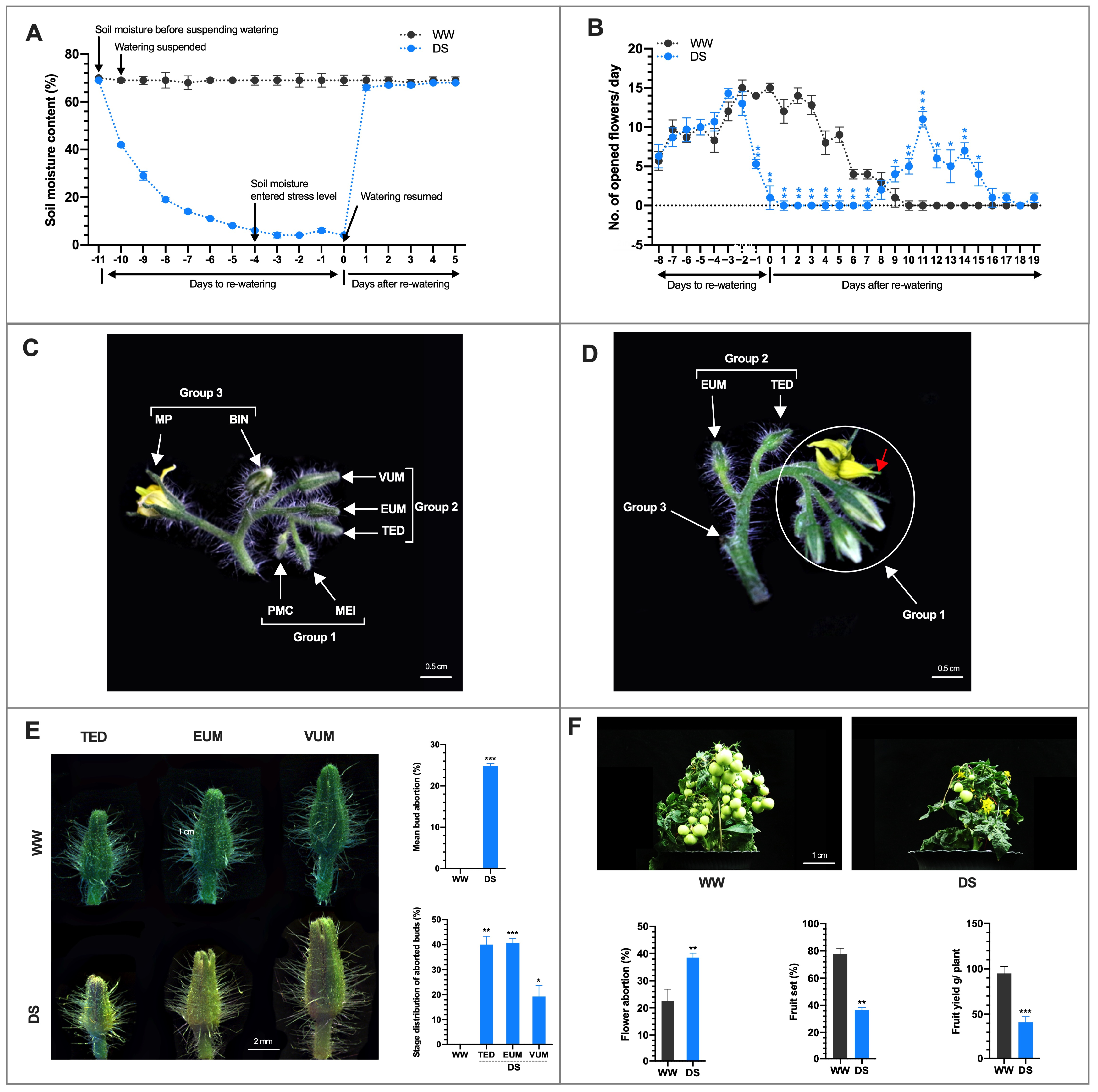
3.1. Drought Stress Affects Flower Development and Fruit Yield
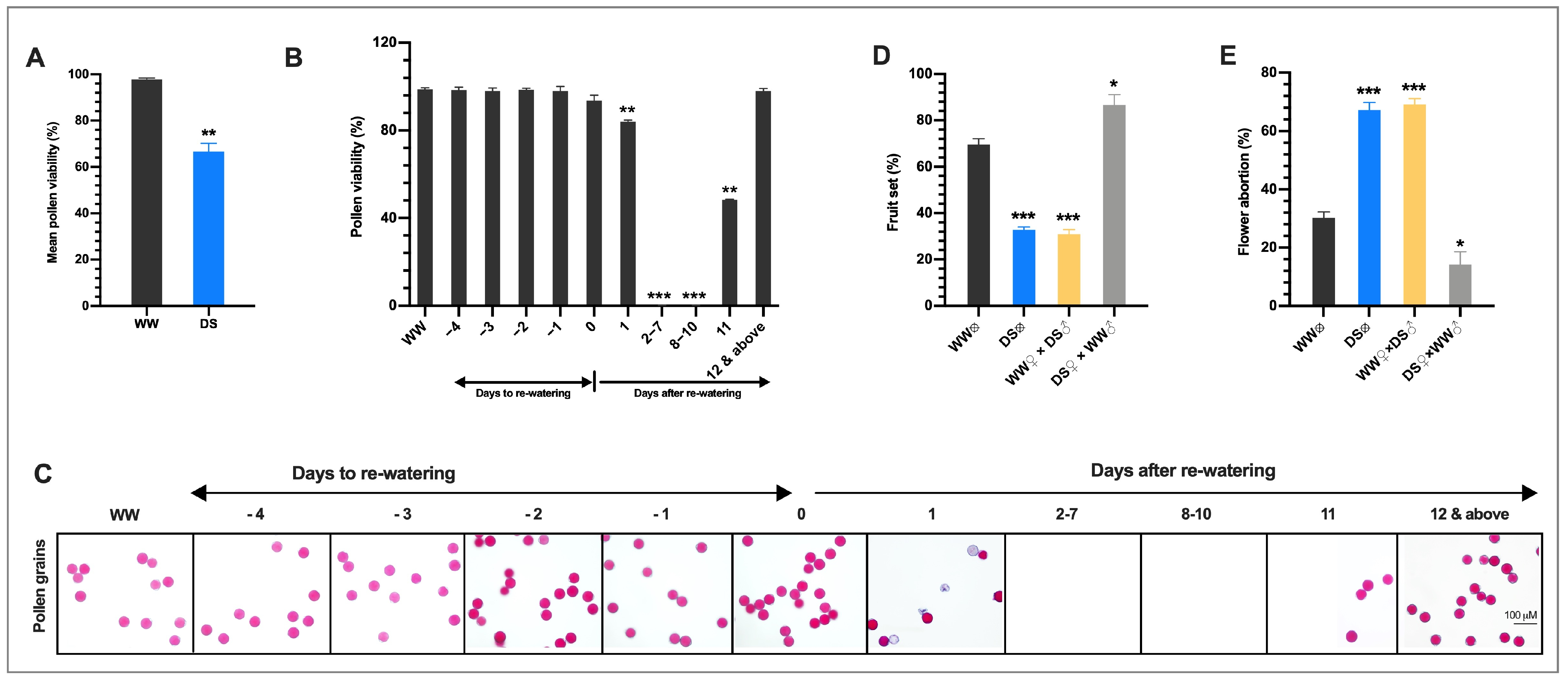
3.2. Drought Stress Reduces the Fertility of the Male Gametophyte
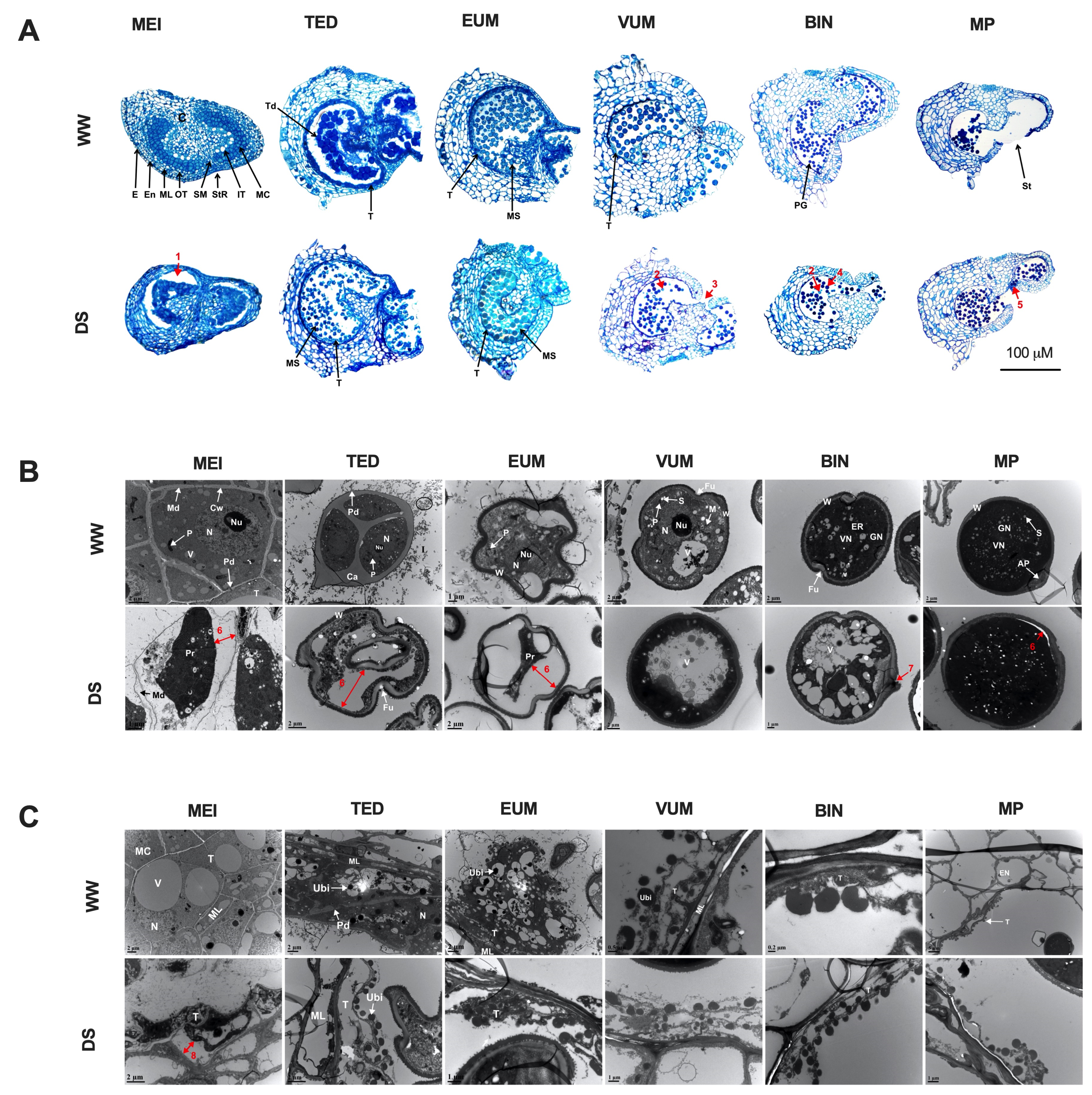
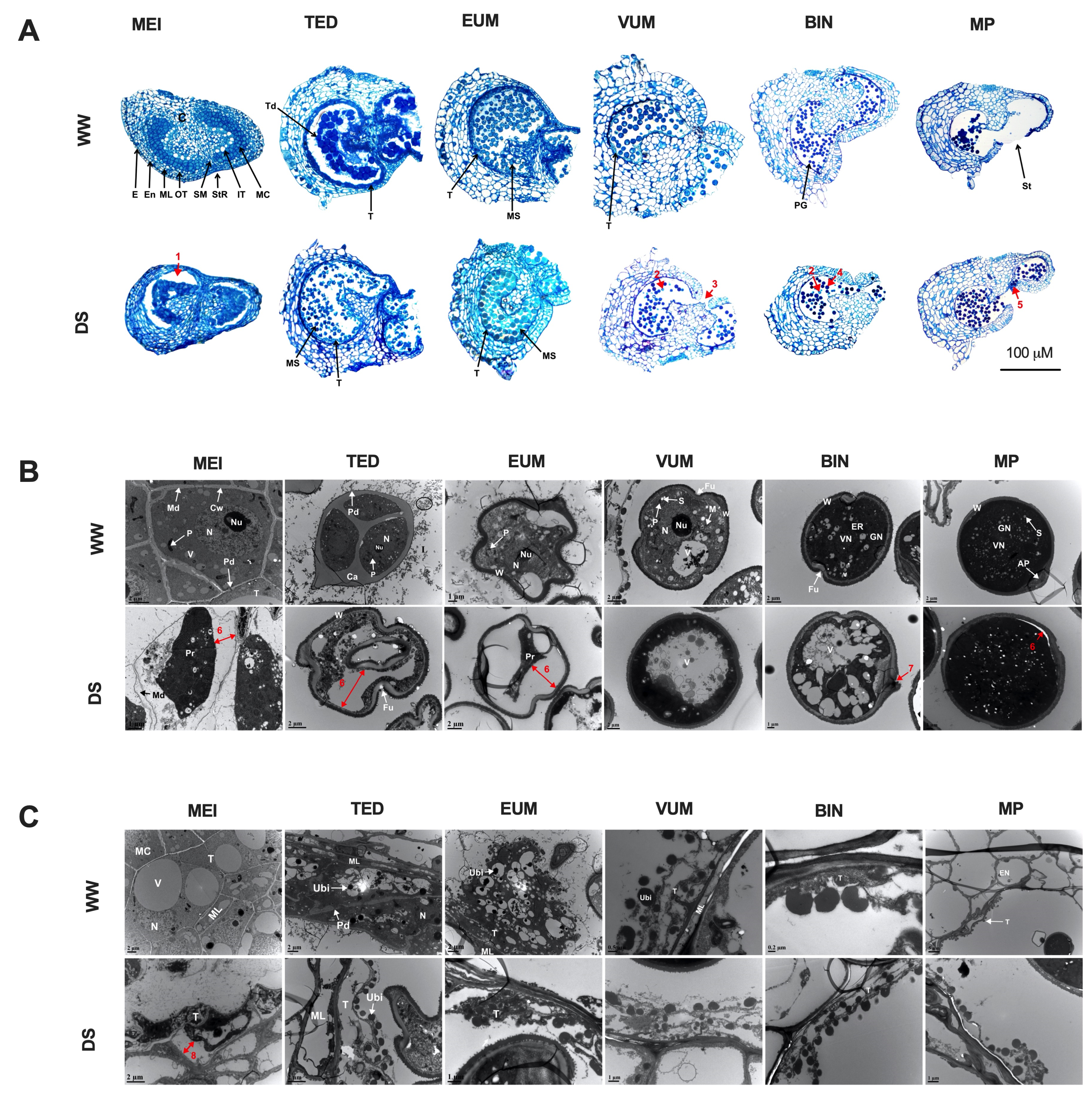
× DS ); and (4) stigmas of flowers on DS plants cross-pollinated with pollen grains from WW plants (DS × WW ). When the stigmas of flowers on WW plants were pollinated with pollen of flowers from DS plants (WW × DS ), fruit set was 55.5% lower than in WW plants, which was consistent with a higher flower abortion rate (Figure 2D), indicating that DS severely affected the male fertility. In contrast, when the stigmas of flowers on DS plants were pollinated with sound pollen of flowers from WW plants (DS × WW ), fruit set and flower abortion were 85.7% and 14.3% which were moderately higher and lower respectively than those in WW plants (Figure 2D,E), indicating that the female fertility was not significantly affected. These results indicated that drought stress-induced irregularities in the male organ is the main cause of floral bud and open flower abortions and the ensuing low fruit set/yield of drought-stressed tomato plants.3.3. Histo-Cytological Analysis of Anther Development under Drought Stress
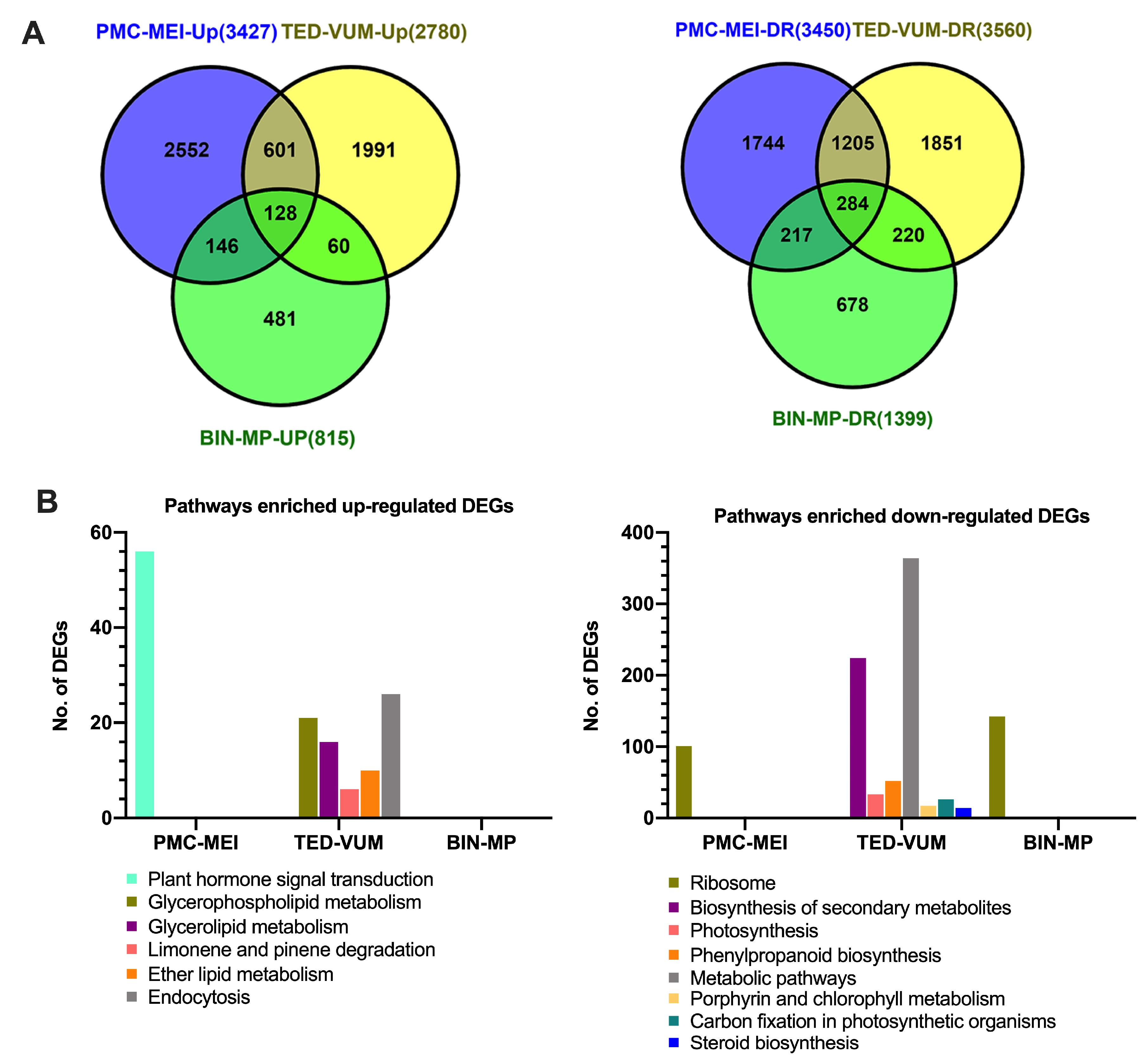
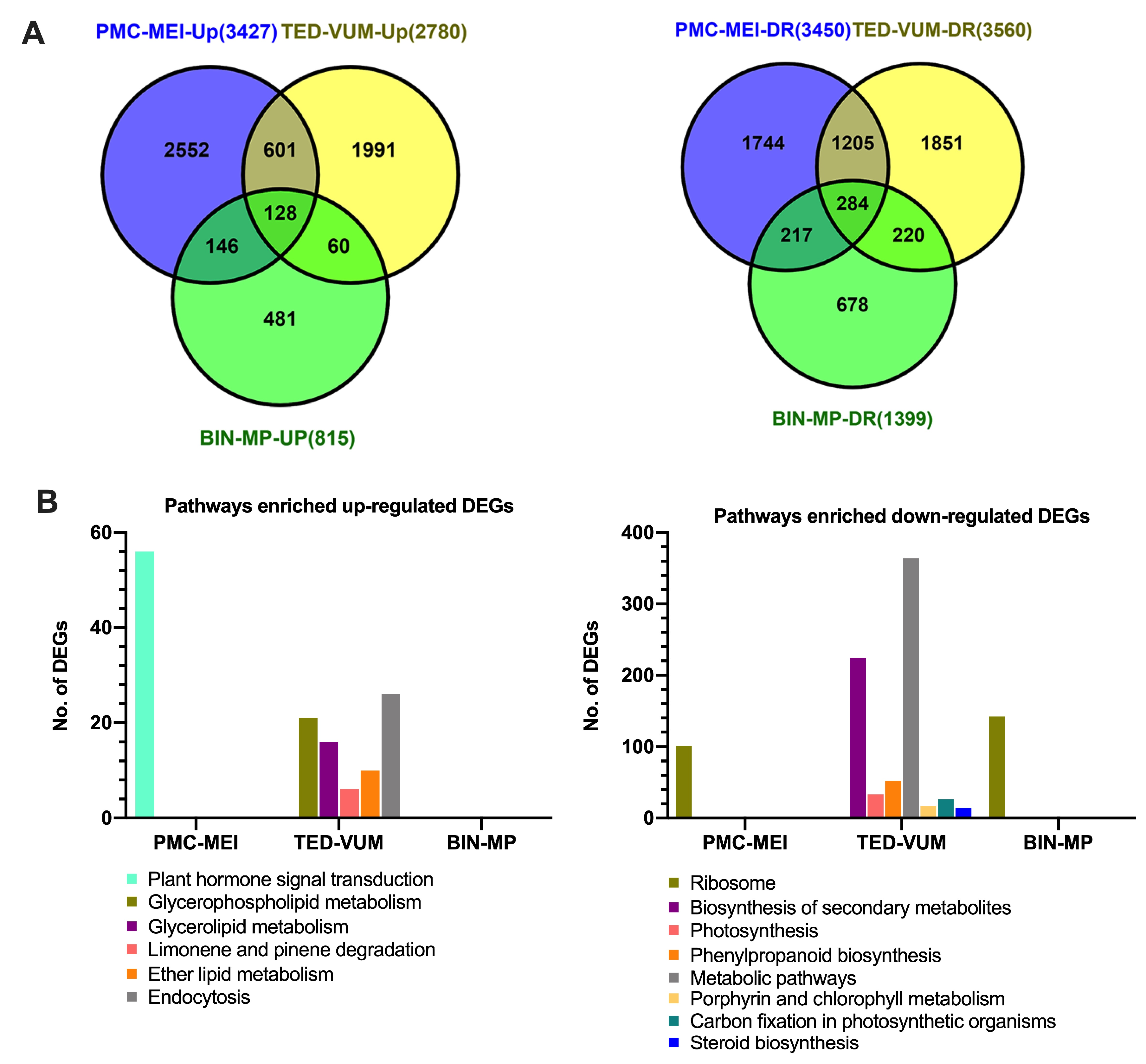
3.4. Transcriptome Changes in Tomato Anthers under Drought Stress
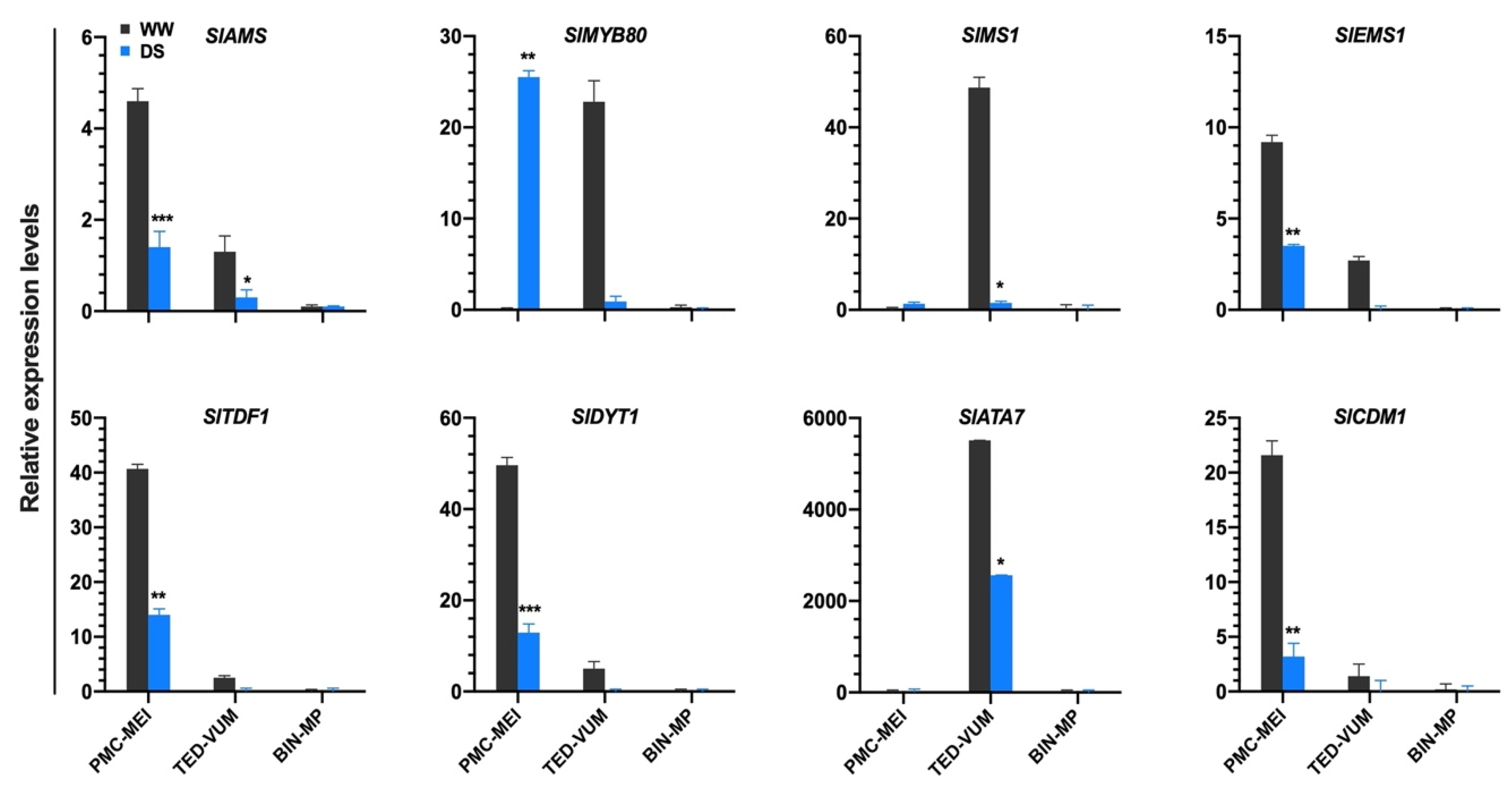
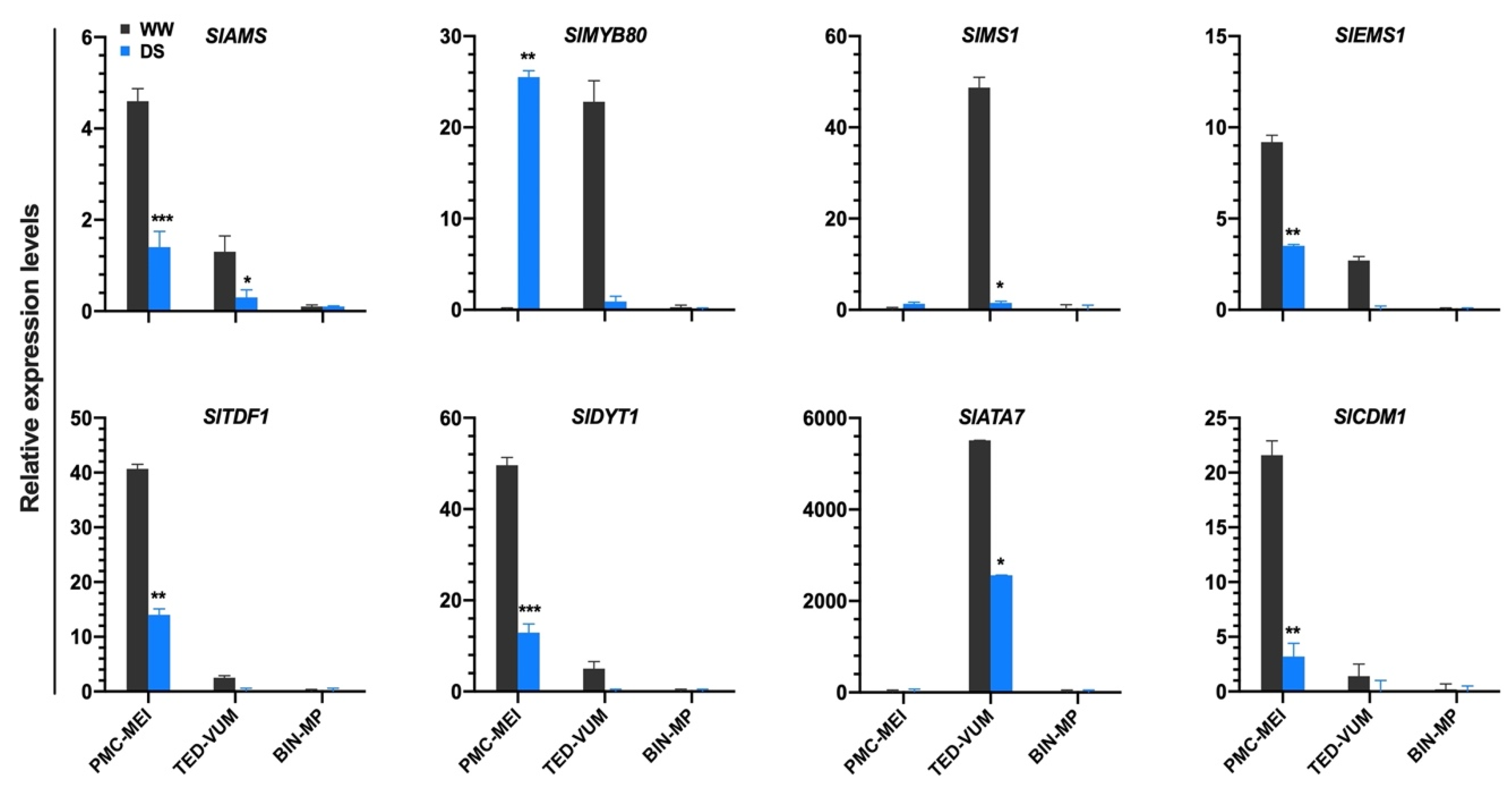
3.5. Impact of Drought Stress on Tapetum-Specific Expressed Genes
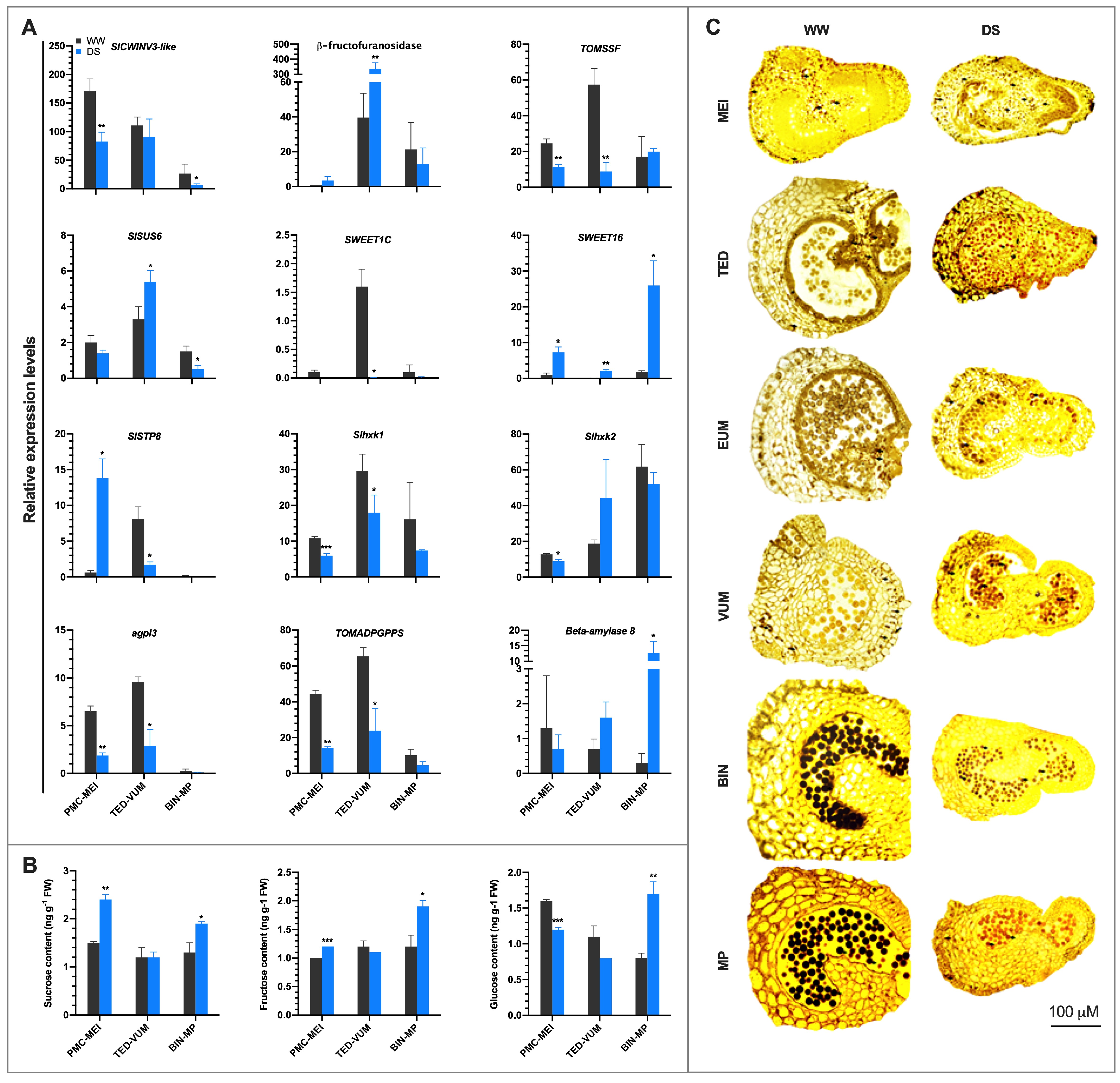
3.6. Drought Stress Disturbs Sucrose and Starch Metabolism and Soluble Sugar Transport
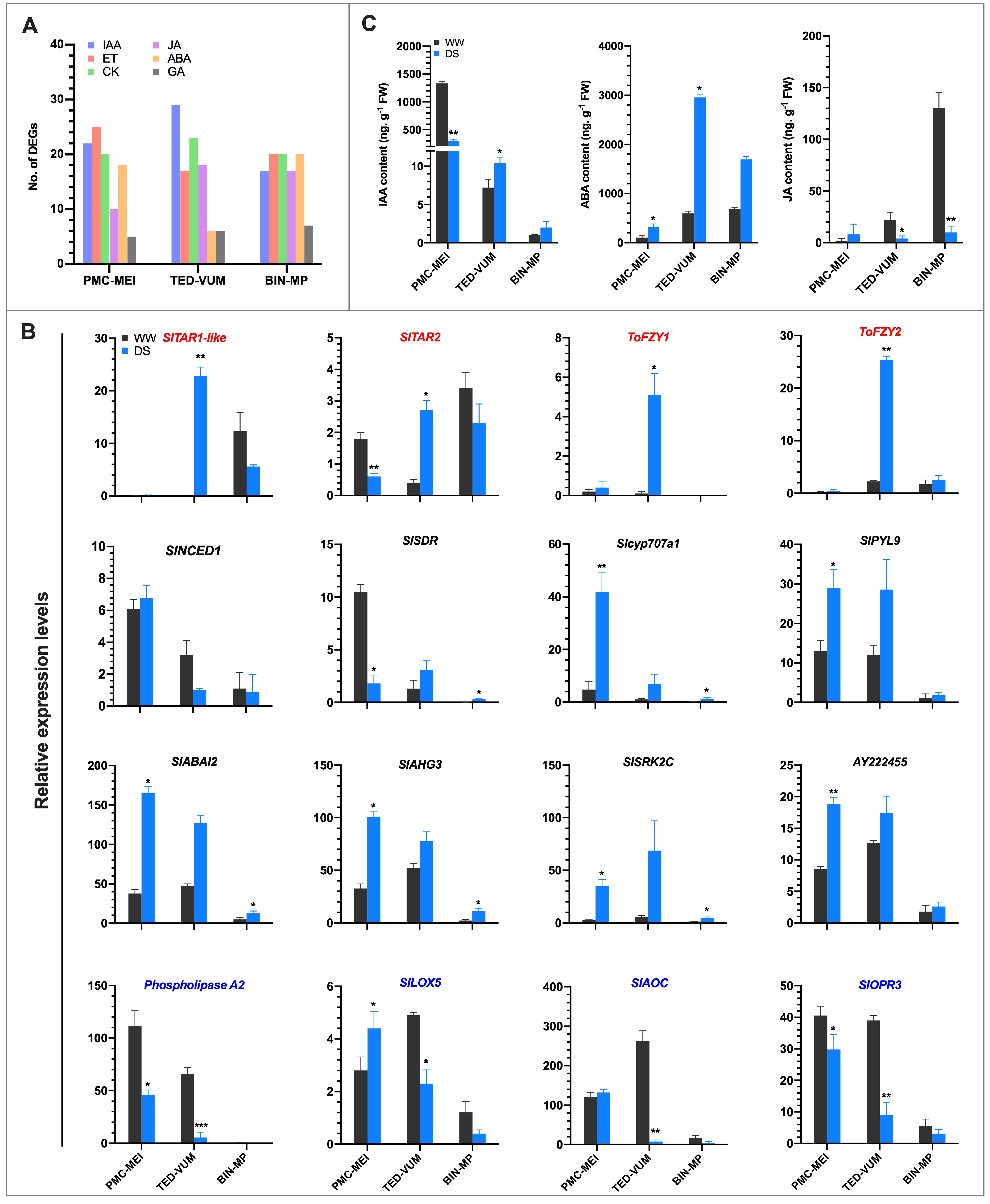
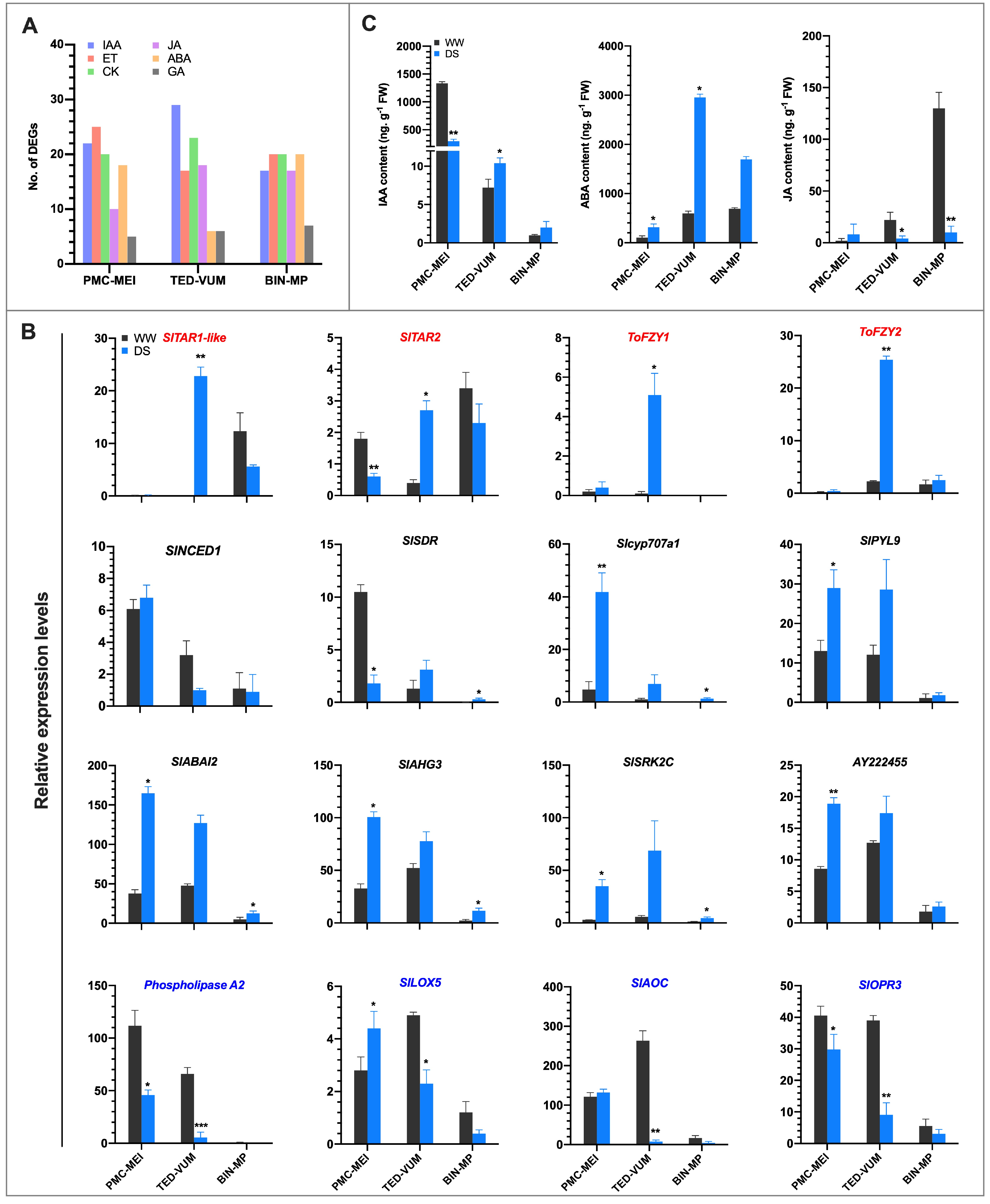
3.7. Effect of Drought on Phytohormones Metabolism and Signaling
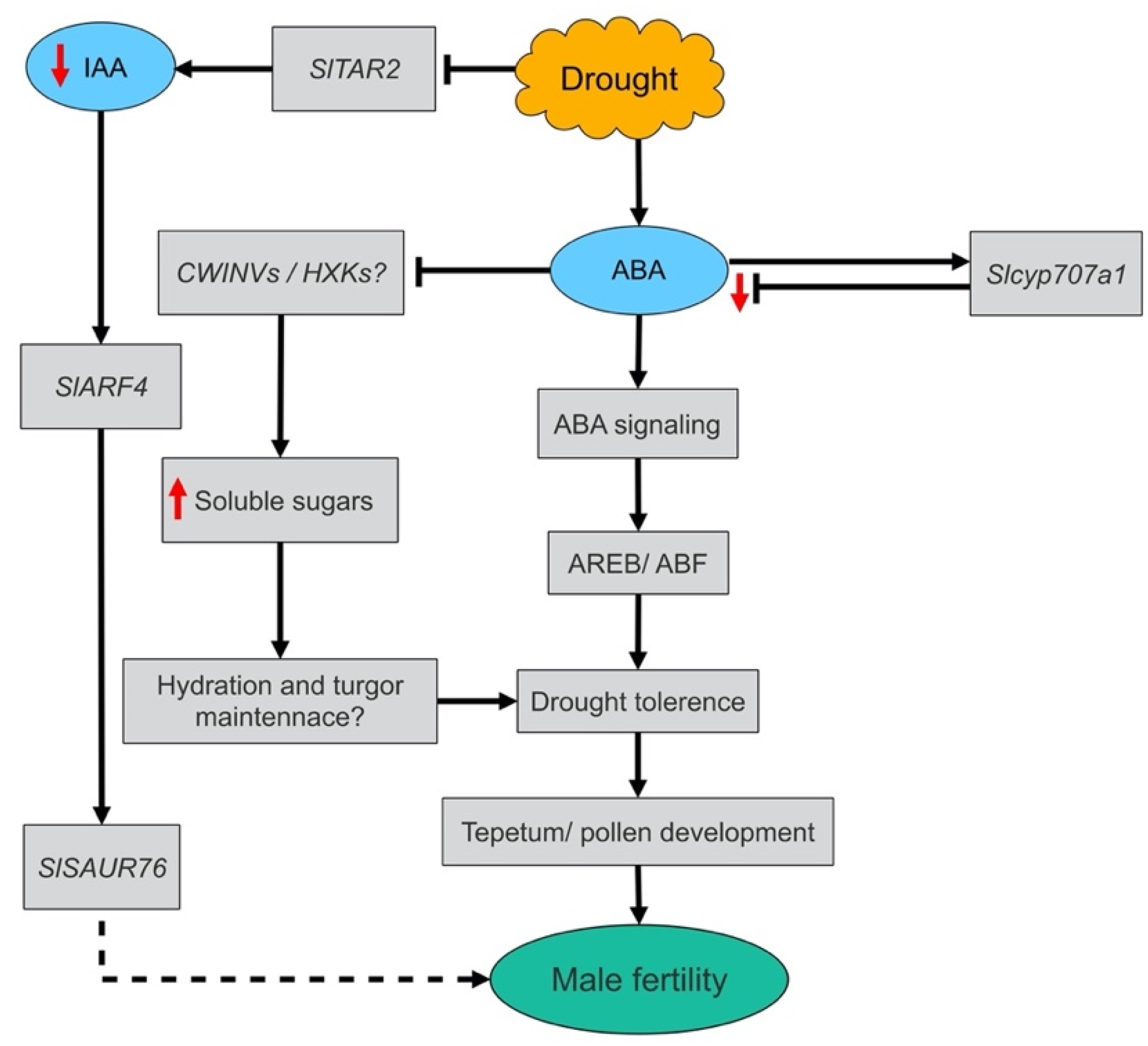
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Zheng, X.; Liu, H.; Able, J.A.; Yang, H.; Zhao, H.; Zhang, M.; Qiao, Y.; Wang, Y.; Liu, M. Effects of drought stress on pollen sterility, grain yield, abscisic acid and protective enzymes in two winter wheat cultivars. Front. Plant. Sci. 2017, 8, 1008. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Serraj, R. Involvement of peduncle elongation, anther dehiscence and spikelet sterility in upland rice response to reproductive-stage drought stress. Environ. Exp. Bot. 2012, 75, 120–127. [Google Scholar] [CrossRef]
- Saini, H.S.; Aspinall, D. Effect of Water Deficit on Sporogenesis in Wheat (Triticum aestivum L.). Ann. Bot. 1981, 48, 623–633. [Google Scholar] [CrossRef]
- Bingham, J. Varietal response in wheat to water supply in the field, and male sterility caused by a period of drought in a glasshouse experiment. Ann. Appl. Biol. 1966, 57, 365–377. [Google Scholar] [CrossRef]
- Seng, K.H. The Effects of Drought, Waterlogging and Heat Stress on Tomatoes (Solanum lycopersicon L.); Lincoln University: Philadelphia, PA, USA, 2014. [Google Scholar]
- Izhar, S.; Frankel, R. Mechanism of male sterility in Petunia: The relationship between pH, callase activity in the anthers, and the breakdown of the microsporogenesis. Theor. Appl. Genet. 1971, 41, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Parish, R.W.; Li, S.F. Death of a tapetum: A programme of developmental altruism. Plant. Sci. 2010, 178, 73–89. [Google Scholar] [CrossRef]
- Sawhney, V.K.; Bhadula, S.K. Microsporogenesis in the normal and male-sterile stamenIess-2 mutant of tomato (Lycopersicon esculentum). Can. J. Bot. 1988, 66, 2013–2021. [Google Scholar] [CrossRef]
- Graybosch, R.A.; Palmer, R.G. Male sterility in soybean (Glycine max). II. Phenotypic expression of the ms4 mutant. Am. J. Bot. 1985, 72, 1751–1764. [Google Scholar] [CrossRef]
- Cecchetti, V.; Altamura, M.M.; Brunetti, P.; Petrocelli, V.; Falasca, G.; Ljung, K.; Costantino, P.; Cardarelli, M. Auxin controls A rabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis. Plant. J. 2013, 74, 411–422. [Google Scholar] [CrossRef]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant. Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Saini, H.S.; Westgate, M.E. Reproductive development in grain crops during drought. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1999; Volume 68, pp. 59–96. [Google Scholar]
- Fu, G.-F.; Jian, S.; Xiong, J.; Li, Y.-R.; Chen, H.-Z.; Le, M.-K.; Tao, L.-X. Changes of oxidative stress and soluble sugar in anthers involve in rice pollen abortion under drought stress. Agric. Sci. China 2011, 10, 1016–1025. [Google Scholar] [CrossRef]
- Oliver, S.N.; Van Dongen, J.T.; Alfred, S.C.; Mamun, E.A.; Zhao, X.; Saini, H.S.; Fernandes, S.F.; Blanchard, C.L.; Sutton, B.G.; Geigenberger, P. Cold-induced repression of the rice anther-specific cell wall invertase gene OSINV4 is correlated with sucrose accumulation and pollen sterility. PlantCell Environ. 2005, 28, 1534–1551. [Google Scholar] [CrossRef]
- Koonjul, P.K.; Minhas, J.S.; Nunes, C.; Sheoran, I.S.; Saini, H.S. Selective transcriptional down-regulation of anther invertases precedes the failure of pollen development in water-stressed wheat. J. Exp. Bot. 2005, 56, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yang, H.; Wei, Z.; Ma, H.; Ge, X. Rice male development under drought stress: Phenotypic changes and stage-dependent transcriptomic reprogramming. Mol. Plant. 2013, 6, 1630–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalonde, S.; Morse, D.; Saini, H.S. Expression of a wheat ADP-glucose pyrophosphorylase gene during development of normal and water-stress-affected anthers. Plant. Mol. Biol. 1997, 34, 445–453. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crop. Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Ji, X.; Dong, B.; Shiran, B.; Talbot, M.J.; Edlington, J.E.; Hughes, T.; White, R.G.; Gubler, F.; Dolferus, R. Control of abscisic acid catabolism and abscisic acid homeostasis is important for reproductive stage stress tolerance in cereals. Plant. Physiol. 2011, 156, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.N.; Dennis, E.S.; Dolferus, R. ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice. Plant. Cell Physiol. 2007, 48, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Sharma, L.; Dalal, M.; Verma, R.K.; Kumar, S.V.; Yadav, S.K.; Pushkar, S.; Kushwaha, S.R.; Bhowmik, A.; Chinnusamy, V. Auxin protects spikelet fertility and grain yield under drought and heat stresses in rice. Environ. Exp. Bot. 2018, 150, 9–24. [Google Scholar] [CrossRef]
- De Ollas Valverde, C.J.; Arbona, V.; Gómez Cadenas, A. Jasmonoyl isoleucine accumulation is needed for abscisic acid build-up in roots of Arabidopsis under water stress conditions. PlantCell Environ. 2015, 38, 2157–2170. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liu, H.; Xiong, L. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant. Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Yang, D.; Zhao, X.; Jiao, C.; Yan, Y.; Lamin-Samu, A.T.; Wang, Q.; Xu, X.; Fei, Z.; Lu, G. Tomato stigma exsertion induced by high temperature is associated with the jasmonate signalling pathway. PlantCell Environ. 2019, 42, 1205–1221. [Google Scholar] [CrossRef]
- Alexander, M. Differential staining of aborted and non-aborted pollen. Stain Tech. 1967, 2, 117–137. [Google Scholar]
- Chen, L.; Yang, D.; Zhang, Y.; Wu, L.; Zhang, Y.; Ye, L.; Pan, C.; He, Y.; Huang, L.; Ruan, Y.L.; et al. Evidence for a specific and critical role of mitogen-activated protein kinase 20 in uni-to-binucleate transition of microgametogenesis in tomato. New Phytol. 2018, 219, 176–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, S.; Joung, J.-G.; Zheng, Y.; Chen, Y.-r.; Liu, B.; Shao, Y.; Xiang, J.Z.; Fei, Z.; Giovannoni, J.J. High-throughput illumina strand-specific RNA sequencing library preparation. Cold Spring Harbor Protoc. 2011, 2011, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.G. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fu, J.; Chu, J.; Sun, X.; Wang, J.; Yan, C. Simple, rapid, and simultaneous assay of multiple carboxyl containing phytohormones in wounded tomatoes by UPLC-MS/MS Using Single SPE Purification and Isotope Dilution. Anal. Sci. 2012, 28, 1081–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Welti, R.; Wang, X. Simultaneous quantification of major phytohormones and related compounds in crude plant extracts by liquid chromatography-electrospray tandem mass spectrometry. Phytochemistry 2008, 69, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Brukhin, V.; Hernould, M.; Gonzalez, N.; Chevalier, C.; Mouras, A. Flower development schedule in tomato Lycopersicon esculentum cv. sweet cherry. Sex. Plant. Reprod. 2003, 15, 311–320. [Google Scholar] [CrossRef]
- Lu, P.; Chai, M.; Yang, J.; Ning, G.; Wang, G.; Ma, H. The Arabidopsis Callose Defective Microspore1 gene is required for male fertility through regulating callose metabolism during microsporogenesis. Plant. Physiol. 2014, 164, 1893–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.-N.; Zhu, J.; Yu, Y.; Teng, X.-D.; Lou, Y.; Xu, X.-F.; Liu, J.-L.; Yang, Z.-N. DYT1 directly regulates the expression of TDF1 for tapetum development and pollen wall formation in Arabidopsis. Plant. J. 2014, 80, 1005–1013. [Google Scholar] [CrossRef]
- Phan, H.A.; Phan, H.A.; Li, S.F.; Li, S.F.; Parish, R.W.; Parish, R.W. MYB80, a regulator of tapetal and pollen development, is functionally conserved in crops. Plant. Mol. Biol. 2012, 78, 171–183. [Google Scholar] [CrossRef]
- Nie, X.-L.; Zhu, J.-W.; Geng, A.-Q.; Xiao, X.-G. Cloning and functional analysis in transgenic tobacco of a tapetum-specific promoter from Arabidopsis. Afr. J. Biotechnol. 2010, 9, 6826–6834. [Google Scholar]
- Feng, X.; Dickinson, H.G. Tapetal cell fate, lineage and proliferation in the Arabidopsis anther. Development 2010, 137, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Nagata, N.; Yoshiba, Y.; Ohme-Takagi, M.; Ma, H.; Shinozaki, K. Arabidopsis Male STERILITY1 encodes a PHD-type transcription factor and regulates pollen and tapetum development. Plant. Cell 2007, 19, 3549–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, A.M.; Kröber, S.; Unte, U.S.; Huijser, P.; Dekker, K.; Saedler, H. The Arabidopsis Aborted Microspores (AMS) gene encodes a MYC class transcription factor. Plant. J. 2003, 33, 413–423. [Google Scholar] [CrossRef]
- Xu, Y.; Iacuone, S.; Li, S.F.; Parish, R.W. MYB80 homologues in Arabidopsis, cotton and Brassica: Regulation and functional conservation in tapetal and pollen development. Bmc Plant. Biol. 2014, 14, 278. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yang, C.; Yuan, Z.; Zhang, D.; Gondwe, M.Y.; Ding, Z.; Liang, W.; Zhang, D.; Wilson, Z.A. The Aborted Microspores regulatory network is required for postmeiotic male reproductive development in Arabidopsis thaliana. Plant. Cell 2010, 22, 91–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Chen, H.; Li, H.; Gao, J.F.; Jiang, H.; Wang, C.; Guan, Y.F.; Yang, Z.N. Defective in Tapetal development and function 1 is essential for anther development and tapetal function for microspore maturation in Arabidopsis. Plant. J. 2008, 55, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Vizcay-Barrena, G.; Conner, K.; Wilson, Z.A. Male Sterility1 is required for tapetal development and pollen wall biosynthesis. Plant. Cell 2007, 19, 3530–3548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granot, D.; David-Schwartz, R.; Kelly, G. Hexose kinases and their role in sugar-sensing and plant development. Front. Plant. Sci. 2013, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Bai, L.; Wang, W.; Shi, H.; Ramón Botella, J.; Zhan, Q.; Liu, K.; Yang, H.Q.; Song, C.P. COP1 promotes ABA-induced stomatal closure by modulating the abundance of ABI/HAB and AHG3 phosphatases. New Phytol. 2021, 229, 2035–2049. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.L.; Benning, G.; Grill, E. ABI2, a second protein phosphatase 2C involved in abscisic acid signal transduction in Arabidopsis. Febs Lett. 1998, 421, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.-C.; Hwang, S.-G.; Endo, A.; Okamoto, M.; Koshiba, T.; Cheng, W.-H. Ectopic expression of Abscisic Acid 2/Glucose Insensitive 1 in Arabidopsis promotes seed dormancy and stress tolerance. Plant. Physiol. 2007, 143, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, M.; Brownfield, L.; Twell, D. Male gametophyte development: A molecular perspective. J. Exp. Bot. 2009, 60, 1465–1478. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, G.N.; Sutton, B.G. Water deficit reduced fertility of young microspores resulting in a decline of viable mature pollen and grain set in rice. J. Agron. Crop. Sci. 2009, 195, 11–18. [Google Scholar] [CrossRef]
- Su, Z.; Ma, X.; Guo, H.; Sukiran, N.L.; Guo, B.; Assmann, S.M.; Ma, H. Flower development under drought stress: Morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis. Plant. Cell 2013, 25, 3785–3807. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Turner, N.C.; Yan, G.; Li, F.; Kadambot, H.M.S. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Saini, H.S.; Sedgley, M.; Aspinall, D. Developmental anatomy in wheat of male-sterility induced by heat-stress, water deficit or abscisic-acid. Aust. J. Plant. Physiol. 1984, 11, 243–253. [Google Scholar] [CrossRef]
- Kiran, A.; Kumar, S.; Nayyar, H.; Sharma, K.D. Low temperature-induced aberrations in male and female reproductive organ development cause flower abortion in chickpea. PlantCell Environ. 2019, 42, 2075–2089. [Google Scholar] [CrossRef]
- Kawanabe, T.; Ariizumi, T.; Kawai-Yamada, M.; Uchimiya, H.; Toriyama, K. Abolition of the tapetum suicide program ruins microsporogenesis. Plant. Cell Physiol. 2006, 47, 784–787. [Google Scholar] [CrossRef] [Green Version]
- Oda, S.; Kaneko, F.; Yano, K.; Fujioka, T.; Masuko, H.; Park, J.-I.; Kikuchi, S.; Hamada, K.; Endo, M.; Nagano, K.; et al. Morphological and gene expression analysis under cool temperature conditions in rice anther development. Genes Genet. Syst. 2010, 85, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.-L.; Jin, Y.; Yang, Y.-J.; Li, G.-J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant. 2010, 3, 942–955. [Google Scholar] [CrossRef]
- Hu, W.; Huang, Y.; Loka, D.A.; Bai, H.; Liu, Y.; Wang, S.; Zhou, Z. Drought-induced disturbance of carbohydrate metabolism in anthers and male abortion of two Gossypium hirsutum cultivars differing in drought tolerance. Plant. Cell Rep. 2019, 39, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.N.; Hailstones, D.L.; Wilkes, M.; Sutton, B.G. Drought stress: Role of carbohydrate metabolism in drought-induced male sterility in rice anthers. J. Agron. Crop. Sci. 2010, 196, 346–357. [Google Scholar] [CrossRef]
- Suwabe, K.; Suzuki, G.; Takahashi, H.; Shiono, K.; Endo, M.; Yano, K.; Fujita, M.; Masuko, H.; Saito, H.; Fujioka, T. Separated transcriptomes of male gametophyte and tapetum in rice: Validity of a laser microdissection (LM) microarray. Plant. Cell Physiol. 2008, 49, 1407–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, L.; Li, Y.; Hu, Q.; Zhu, L.; Gao, W.; Wu, Y.; Ding, Y.; Liu, S.; Yang, X.; Zhang, X. Sugar and auxin signaling pathways respond to high-temperature stress during anther development as revealed by transcript profiling analysis in cotton. Plant. Physiol. 2014, 164, 1293–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, T.; Oshino, T.; Miura, S.; Tomabechi, M.; Tsunaga, Y.; Higashitani, N.; Miyazawa, Y.; Takahashi, H.; Watanabe, M.; Higashitani, A. Auxins reverse plant male sterility caused by high temperatures. Proc. Natl. Acad. Sci. USA 2010, 107, 8569–8574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Kai, W.; Liang, B.; Wang, J.; Jiang, L.; Du, Y.; Sun, Y.; Leng, P. The functional analysis of SlNCED1 in tomato pollen development. Cell. Mol. Life Sci. 2018, 75, 3457–3472. [Google Scholar] [CrossRef]
- Liu, F.; Andersen, M.N.; Jensen, C.R. Loss of pod set caused by drought stress is associated with water status and ABA content of reproductive structures in soybean. Funct. Plant. Biol. 2003, 30, 271–280. [Google Scholar] [CrossRef]
- Setter, T.L.; Flannigan, B.A.; Melkonian, J. Loss of kernel set due to water deficit and shade in maize: Carbohydrate supplies, abscisic acid, and cytokinins. Crop. Sci. 2001, 41, 1530–1540. [Google Scholar] [CrossRef]
 × DS ) and stigma of flowers on DS plants pollinated with pollen grains from WW plants (DS ×WW ). Values presented are means ±SD (n = 150 for WW ⊗ and DS ⊗ experiments; n = 125 for WW × DS and DS × WW experiments. * p < 0.05; ** p < 0.01; *** p < 0.001 (t-test).
× DS ) and stigma of flowers on DS plants pollinated with pollen grains from WW plants (DS ×WW ). Values presented are means ±SD (n = 150 for WW ⊗ and DS ⊗ experiments; n = 125 for WW × DS and DS × WW experiments. * p < 0.05; ** p < 0.01; *** p < 0.001 (t-test).
× DS ) and stigma of flowers on DS plants pollinated with pollen grains from WW plants (DS ×WW ). Values presented are means ±SD (n = 150 for WW ⊗ and DS ⊗ experiments; n = 125 for WW × DS and DS × WW experiments. * p < 0.05; ** p < 0.01; *** p < 0.001 (t-test).
× DS ) and stigma of flowers on DS plants pollinated with pollen grains from WW plants (DS ×WW ). Values presented are means ±SD (n = 150 for WW ⊗ and DS ⊗ experiments; n = 125 for WW × DS and DS × WW experiments. * p < 0.05; ** p < 0.01; *** p < 0.001 (t-test).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamin-Samu, A.T.; Farghal, M.; Ali, M.; Lu, G. Morpho-Physiological and Transcriptome Changes in Tomato Anthers of Different Developmental Stages under Drought Stress. Cells 2021, 10, 1809. https://doi.org/10.3390/cells10071809
Lamin-Samu AT, Farghal M, Ali M, Lu G. Morpho-Physiological and Transcriptome Changes in Tomato Anthers of Different Developmental Stages under Drought Stress. Cells. 2021; 10(7):1809. https://doi.org/10.3390/cells10071809
Chicago/Turabian StyleLamin-Samu, Anthony Tumbeh, Mohamed Farghal, Muhammad Ali, and Gang Lu. 2021. "Morpho-Physiological and Transcriptome Changes in Tomato Anthers of Different Developmental Stages under Drought Stress" Cells 10, no. 7: 1809. https://doi.org/10.3390/cells10071809
APA StyleLamin-Samu, A. T., Farghal, M., Ali, M., & Lu, G. (2021). Morpho-Physiological and Transcriptome Changes in Tomato Anthers of Different Developmental Stages under Drought Stress. Cells, 10(7), 1809. https://doi.org/10.3390/cells10071809