
Iron Supplement-Enhanced Growth and Development of Hydrangea macrophylla In Vitro under Normal and High pH



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. Iron and pH Treatments
2.3. Measurement of the Growth Characteristics
2.4. Contents of Chlorophyll, Carotenoid, and Total Anthocyanin
2.5. Total Soluble Proteins and Antioxidant Enzyme Activities
2.6. FCR Activity
2.7. Scanning Electron Microscopic (SEM) Analysis of the Stomata
2.8. Determination of Macro- and Micronutrient Contents Using Inductively Coupled Plasma Spectrometer
2.9. Quantitative Real-Time PCR Analysis
2.10. Data Collection and Analysis
3. Results
3.1. Analysis of the Morphological and Growth Parameters
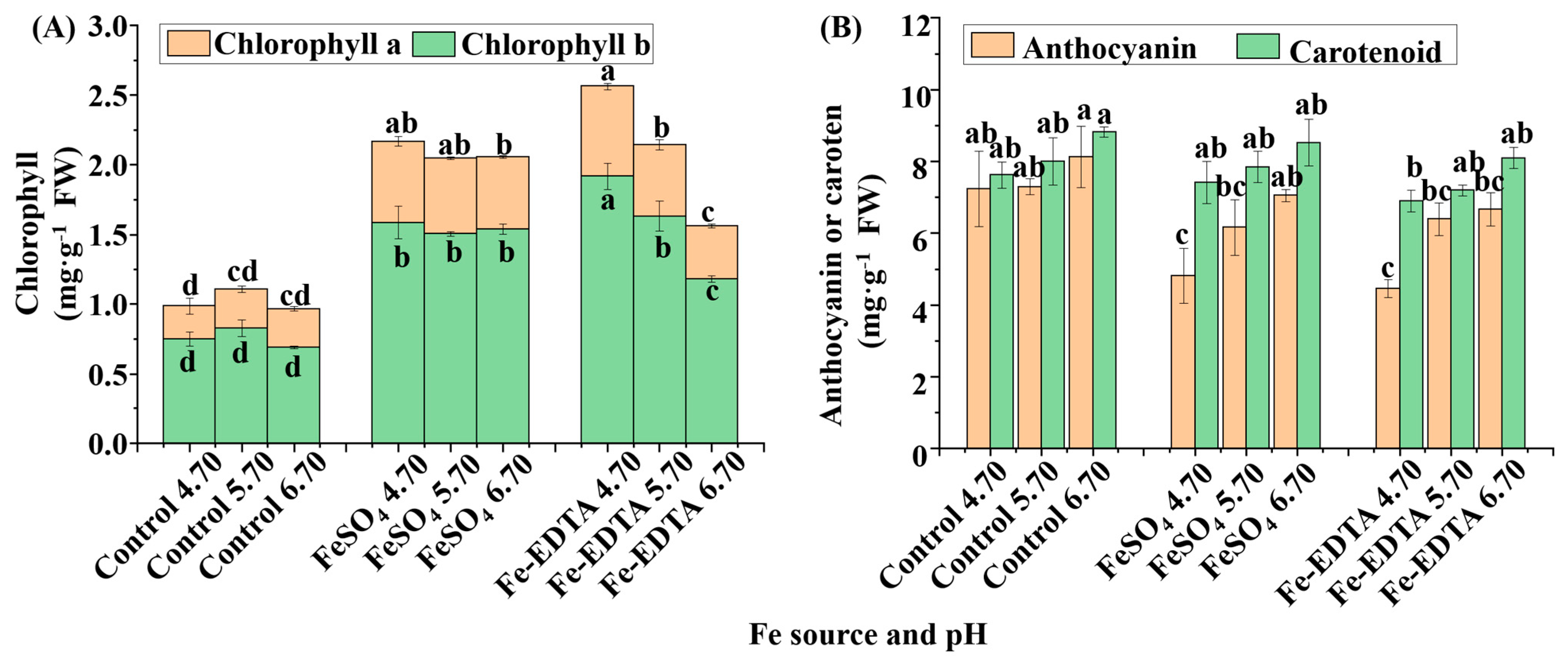
3.2. Leaf Color and Pigment Content Analyses
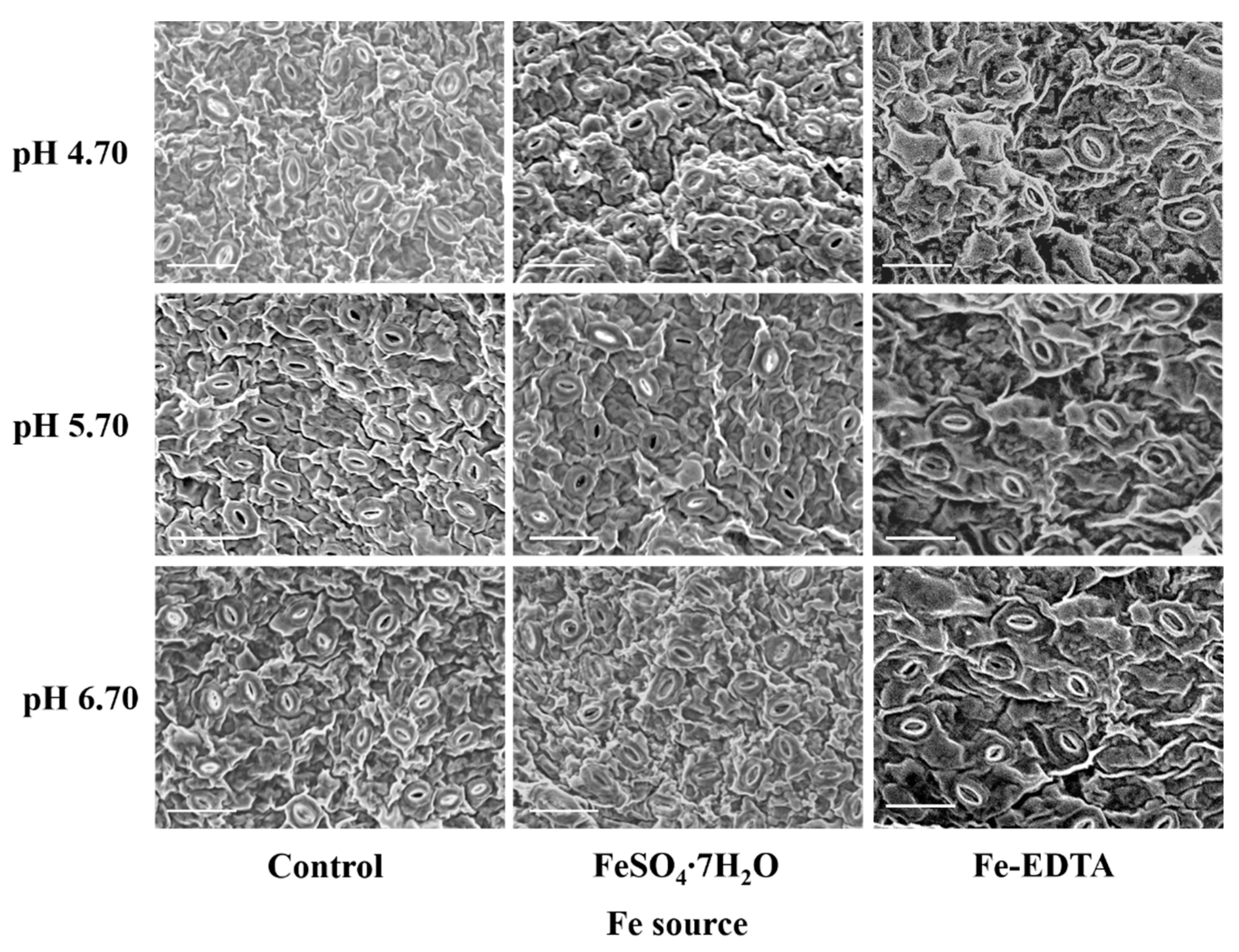
3.3. Analysis of the Stomata and Chlorophyll Fluorescence Parameters
3.4. Soluble Proteins, FCR, and Antioxidant Enzyme Activities
3.5. Micro- and Macronutrient Contents
3.6. Quantitative Real-Time RT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomidis, T.; Tsipouridis, C. Influence of rootstocks, pH, iron supply (in nutrient solutions) and Agrobacterium radiobacter on chlorophyll and iron concentration in leaves of a peach variety. J. Plant Nutr. 2005, 28, 1833–1842. [Google Scholar] [CrossRef]
- Peng, J.; Xue, C.; Dong, X.; Zeng, C.; Wu, Y.; Cao, F. Gene cloning and analysis of the pattern of expression of the transcription factor HymMYB2 related to blue flower formation in hydrangea macrophylla. Euphytica 2021, 217, 115. [Google Scholar] [CrossRef]
- McClintock, E. A monograph of the genus Hydrangea. Proc. Calif. Acad. Sci. 1957, 29, 147–256. [Google Scholar]
- Matsuoka, K.K.; Furuhata, S.I.; Fujisaki, R.I.; Kawakamia, T.; Mogi, S.; Yoshihara, H.; Aokia, N.; Ishii, A.; Shibuya, T. Anti-malarial activity of leaf-extract of Hydrangea macrophylla, a common japanese plant. Acta Med. Okayama 2000, 54, 227–232. [Google Scholar]
- Head, K.A. Natural approaches to prevention and treatment of infections of the lower urinary tract. Altern. Med. Rev. 2008, 13, 227–244. [Google Scholar]
- Zhang, H.; Matsuda, H.; Kumahara, A.; Ito, Y.; Nakamura, S.; Yoshikawa, M. New type of anti-diabetic compounds from the processed leaves of Hydrangea macrophylla var. thunbergii (Hydrangeae Dulcis Folium). Bioorg. Med. Chem. Lett. 2007, 17, 4972–4976. [Google Scholar] [CrossRef]
- Zhang, S.; Xin, H.; Li, Y.; Zhang, D.; Shi, J.; Yang, J.; Chen, X. Skimmin, a coumarin from Hydrangea paniculata, slows down the progression of membranous glomerulonephritis by anti-inflammatory effects and inhibiting immune complex deposition. Evid. Based Complementary Altern. Med. 2013, 2013, 819296. [Google Scholar]
- Sebastian, T.K.; Heuser, C.W. In vitro propagation of Hydrangea quercifolia Bartr. Sci. Hortic. 1987, 31, 303–309. [Google Scholar] [CrossRef]
- Ruffoni, B.; Sacco, E.; Savona, M. In vitro propagation of Hydrangea spp. In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Humana Press: Totowa, NJ, USA, 2012; pp. 231–244. [Google Scholar]
- Malyarovskaya, V.; Samarina, L. In vitro morphogenesis of ornamental shrubs Camellia japonica and Hydrangea macrophylla. Plant Tissue Cult. Biotech. 2017, 27, 181–187. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (cu, zn, mn, fe, ni, mo, b, cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Lascelles, J. Adaptation to form bacteriochlorophyll in Rhodopseudomonas spheroides: Changes in activity of enzymes concerned in pyrrole synthesis. Biochem. J. 1959, 72, 508–518. [Google Scholar] [CrossRef]
- Abadía, J. Leaf responses to fe deficiency: A review. J. Plant Nutr. 1992, 15, 1699–1713. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Kissel, M. Different strategies in higher plants in mobilization and uptake of iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Martín-Barranco, A.; Spielmann, J.; Dubeaux, G.; Vert, G.; Zelazny, E. Dynamic control of the high-affinity iron uptake complex in root epidermal cells. Plant Physiol. 2020, 184, 1236–1250. [Google Scholar] [CrossRef]
- Colangelo, E.P.; Guerinot, M.L. The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell 2004, 16, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, L.; Ying, Y.; Wang, J.; Shao, J.F.; Yamaji, N.; Whelan, J.; Ma, J.F.; Shou, H. A transcription factor OsbHLH156 regulates Strategy ii iron acquisition through localising IRO2 to the nucleus in rice. New Phytol. 2020, 225, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef]
- Connorton, J.M.; Balk, J.; Rodríguez-Celma, J. Iron homeostasis in plants—A brief overview. Metallomics 2017, 9, 813–823. [Google Scholar] [CrossRef]
- Von Wiren, N.; Klair, S.; Bansal, S.; Briat, J.F.; Khodr, H.; Shioiri, T.; Leigh, R.A.; Hider, R.C. Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants. Plant Physiol. 1999, 119, 1107–1114. [Google Scholar] [CrossRef]
- Vert, G.A.; Briat, J.F.; Curie, C. Dual regulation of the Arabidopsis high-affinity root iron uptake system by local and long-distance signals. Plant Physiol. 2003, 132, 796–804. [Google Scholar] [CrossRef]
- Rodríguez-Celma, J.; Pan, I.; Li, W.D.; Lan, P.D.; Buckhout, T.J.; Schmidt, W. The transcriptional response of Arabidopsis leaves to Fe deficiency. Front. Plant Sci. 2013, 4, 276. [Google Scholar] [CrossRef]
- Santi, S.; Schmidt, W. Dissecting iron deficiency-induced proton extrusion in Arabidopsis roots. New Phytol. 2009, 183, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Römheld, V. Strategies of plants for acquisition of iron. Plant Soil 1994, 165, 261–274. [Google Scholar] [CrossRef]
- Kalbasl, M.; Manuchehri, N.; Filsoof, F. Local acidification of soil as a means to alleviate iron chlorosis in quince orchards. J. Plant Nutr. 1986, 9, 1001–1007. [Google Scholar] [CrossRef]
- Bertamini, M.; Muthuchelian, K.; Nedunchezhian, N. Iron deficiency induced changes on the donor side of PS II in field grown grapevine (Vitis vinifera L. Cv. Pinot noir) leaves. Plant Sci. 2002, 162, 599–605. [Google Scholar] [CrossRef]
- Platt-Aloia, K.; Thomson, W.; Terry, N. Changes in plastid ultrastructure during iron nutrition-mediated chloroplast development. Protoplasma 1983, 114, 85–92. [Google Scholar] [CrossRef]
- Rombolà, A.D.; Gogorcena, Y.; Larbi, A.; Morales, F.; Baldi, E.; Marangoni, B.; Tagliavini, M.; Abadía, J. Iron deficiency-induced changes in carbon fixation and leaf elemental composition of sugar beet (Beta vulgaris) plants. Plant Soil 2005, 271, 39–45. [Google Scholar] [CrossRef][Green Version]
- Morales, F.; Abadía, A.; Abadía, J. Characterization of the xanthophyll cycle and other photosynthetic pigment changes induced by iron deficiency in sugar beet (Beta vulgaris L.). Plant Physiol. 1990, 94, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Larbi, A.; Abadía, A.; Morales, F.; Abadía, J. Fe resupply to Fe-deficient sugar beet plants leads to rapid changes in the violaxanthin cycle and other photosynthetic characteristics without significant de novo chlorophyll synthesis. Photosynth. Res. 2004, 79, 59–69. [Google Scholar] [CrossRef]
- Xiao, J.; Park, Y.G.; Guo, G.; Jeong, B.R. Effect of iron source and medium pH on growth and development of Sorbus commixta in vitro. Int. J. Mol. Sci. 2021, 22, 133. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Şükran, D.E.R.E.; Güneş, T.; Sivaci, R. Spectrophotometric determination of chlorophyll-A, B and total carotenoid contents of some algae species using different solvents. Turk. J. Bot. 1998, 22, 13–18. [Google Scholar]
- Pirie, A.; Mullins, M.G. Changes in anthocyanin and phenolics content of grapevine leaf and fruit tissues treated with sucrose, nitrate, and abscisic acid. Plant Physiol. 1976, 58, 468–472. [Google Scholar] [CrossRef]
- Soundararajan, P.; Manivannan, A.; Ko, C.H.; Park, J.E.; Jeong, B.R. Evaluation of relative toxicity caused by deicing agents on photosynthesis, redox homeostasis, and the osmoregulatory system in creeper-type plants. Hortic. Environ. Biotechnol. 2019, 60, 175–186. [Google Scholar] [CrossRef]
- Soundararajan, P.; Manivannan, A.; Park, Y.G.; Muneer, S.; Jeong, B.R. Silicon alleviates salt stress by modulating antioxidant enzyme activities in Dianthus caryophyllus ‘tula’. Hortic. Environ. Biotechnol. 2015, 56, 233–239. [Google Scholar] [CrossRef]
- Bienfait, H.F.; van den Briel, W.; Mesland-Mul, N.T. Free space iron pools in roots: Generation and mobilization. Plant Physiol. 1985, 78, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Tanaka, M. Scanning electron microscope observation of leaf surface of Hydrangea macrophylla. Sci. Repts. Yokohama Natl. Univ. 1989, 2, 35–44. [Google Scholar]
- Zhang, H.; Dotson, P. The use of microwave muffle furnace for dry ashing plant tissue samples. Commun. Soil Sci. Plant Anal. 1994, 25, 1321–1327. [Google Scholar] [CrossRef]
- Chen, Q.; Yu, H.; Wang, X.; Xie, X.; Yue, X.; Tang, H. An alternative cetyltrimethylammonium bromide-based protocol for RNA isolation from blackberry (Rubus L.). Genet. Mol. Res. GMR 2012, 11, 1773–1782. [Google Scholar] [CrossRef]
- Hariri, M.R.; Yusuf, M.S.; Nurmamulyosari, L.D.; Kharisma, A.E. Effect of soil ph on the anthocyanin level of hortensia (Hydrangea macrophylla). KnE Life Sci. 2015, 2, 613–616. [Google Scholar] [CrossRef]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- McCauley, A.; Jones, C.; Jacobsen, J. Soil pH and organic matter. Nutr. Manag. Modul. 2009, 8, 1–12. [Google Scholar]
- Gabriela, P.R.; Ramona, P.I. Indicator plants of fertility and soil acidity. Ann. ’Constantin Brancusi’ Univ. Targu-Jiu. Eng. Ser. 2018, 4, 32–36. [Google Scholar]
- Suoning, Y.; Li, Z.; Chun, L.; Suxia, Y.; Junyun, Z.; Gaitian, Z. Influences of temperature and soils acidity on the growth and development of Hydrangea macrophylla ‘bailmer’. J. Agric. Sci. Technol. 2020, 22, 24. [Google Scholar]
- Lee, J.S.; Ahn, J.S.; Gi, G.Y.; Hwang, I.T.; Yoon, B.K. Growth charateristic and alumimum contents by nutrient solution of regulated pH in cut Hydrangea ‘adria’. Trends Agric. Life Sci. 2015, 50, 53–55. [Google Scholar]
- Bailey, D.A.; Nelson, P.V.; Fonteno, W.C. Substrates pH and Water Quality; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Pérez-Labrada, F.; Benavides-Mendoza, A.; Juárez-Maldonado, A.; Solís-Gaona, S.; González-Morales, S. Organic acids combined with Fe-chelate improves ferric nutrition in tomato grown in calcisol soil. J. Soil Sci. Plant Nutr. 2020, 20, 673–683. [Google Scholar] [CrossRef]
- Bauer, P.; Hell, R. Translocation of iron in plant tissues. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Springer: Dordrecht, The Nertherlands, 2006; pp. 279–288. [Google Scholar]
- Chapman, S.K.; White, S.A.; Reid, G.A. Flavocytochrome B2. Adv. Inorg. Chem. 1991, 36, 257–301. [Google Scholar]
- Moog, P.R.; Brüggemann, W. Iron reductase systems on the plant plasma membrane—A review. Plant Soil 1994, 165, 241–260. [Google Scholar] [CrossRef]
- Susin, S.; Abadia, A.; González-Reyes, J.A.; Lucena, J.J.; Abadia, J. The ph requirement for in vivo activity of the iron-deficiency-induced “turbo” ferric chelate reductase (a comparison of the iron-deficiency-induced iron reductase activities of intact plants and isolated plasma membrane fractions in sugar beet). Plant Physiol. 1996, 110, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Oh, M.M.; Park, S.A. Ferric-chelate reductase activity is a limiting factor in iron uptake in spinach and kale roots. Hortic. Environ. Biotechnol. 2016, 57, 462–469. [Google Scholar] [CrossRef]
- Abbaszadeh-Dahaji, P.; Masalehi, F.; Akhgar, A. Improved growth and nutrition of sorghum (Sorghum bicolor) plants in a low-fertility calcareous soil treated with plant growth–promoting rhizobacteria and Fe-EDTA. J. Soil Sci. Plant Nutr. 2020, 20, 31–42. [Google Scholar] [CrossRef]
- Sunda, W.; Huntsman, S. Effect of pH, light, and temperature on Fe–EDTA chelation and Fe hydrolysis in seawater. Mar. Chem. 2003, 84, 35–47. [Google Scholar] [CrossRef]
- Chaney, R.L. Plants can utilize iron form Fe-N, N’-di-(2-hydroxybenzoyl)-ethylenediamine-N, N’-diacetic acid, a ferric chelate with 106 greater formation constant than Fe-EDDHA. J. Plant Nutr. 1988, 11, 1033–1050. [Google Scholar] [CrossRef]
- Fisher, P.R.; Wik, R.M.; Smith, B.R.; Pasian, C.C.; Kmetz-González, M.; Argo, W.R. Correcting iron deficiency in calibrachoa grown in a container medium at high pH. HortTechnology 2003, 13, 308–313. [Google Scholar] [CrossRef]
- Puschenreiter, M.; Gruber, B.; Wenzel, W.W.; Schindlegger, Y.; Hann, S.; Spangl, B.; Schenkeveld, W.D.; Kraemer, S.M.; Oburger, E. Phytosiderophore-induced mobilization and uptake of Cd, Cu, Fe, Ni, Pb and Zn by wheat plants grown on metal-enriched soils. Environ. Exp. Bot. 2017, 138, 67–76. [Google Scholar] [CrossRef]
- Ghasemi-Fasaei, R.; Ronaghi, A. Interaction of iron with copper, zinc, and manganese in wheat as affected by iron and manganese in a calcareous soil. J. Plant Nutr. 2008, 31, 839–848. [Google Scholar] [CrossRef]
- Xue, H.; Sigg, L.; Kari, F.G. Speciation of EDTA in natural waters: Exchange kinetics of Fe-EDTA in river water. Environ. Sci. Technol. 1995, 29, 59–68. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Z.; Gao, Y.; Liu, L.; Yu, R.; Jin, J.; Luo, L.; Hui, X.; Li, F.; Li, M. EDTA alone enhanced soil zinc availability and winter wheat grain Zn concentration on calcareous soil. Environ. Exp. Bot. 2017, 141, 19–27. [Google Scholar] [CrossRef]
- Slamet-Loedin, I.H.; Johnson-Beebout, S.E.; Impa, S.; Tsakirpaloglou, N. Enriching rice with Zn and fe while minimizing Cd risk. Front. Plant Sci. 2015, 6, 121. [Google Scholar] [CrossRef]
- Terry, N.; Low, G. Leaf chlorophyll content and its relation to the intracellular localization of iron. J. Plant Nutr. 1982, 5, 301–310. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Klaus, A.A.; Kartashov, A.V.; Kusnetsov, V.V. Distribution of Cd and other cations between the stroma and thylakoids: A quantitative approach to the search for Cd targets in chloroplasts. Photosynth. Res. 2019, 139, 337–358. [Google Scholar] [CrossRef]
- Peterson, G.C.; Onken, A.B. Relationship between chlorophyll concentration and iron chlorosis in grain sorghum. Crop. Sci. 1992, 32, 964–967. [Google Scholar] [CrossRef]
- Mann, A.; Singh, A.; Oza, S.; Goswami, N.; Mehta, D.; Chaudhari, V. Effect of iron source on iron deficiency induced chlorosis in groundnut. Legume Res. Int. J. 2017, 40, 241–249. [Google Scholar] [CrossRef]
- Nemati Lafmejani, Z.; Jafari, A.A.; Moradi, P.; Ladan Moghadam, A. Impact of foliar application of iron-chelate and iron nano particles on some morpho-physiological traits and rssential oil composition of peppermint (Mentha piperita L.). J. Essent. Oil Bear. Plants 2018, 21, 1374–1384. [Google Scholar] [CrossRef]
- Fernández, V.; Eichert, T.; Del Río, V.; López-Casado, G.; Heredia-Guerrero, J.A.; Abadía, A.; Heredia, A.; Abadía, J. Leaf structural changes associated with iron deficiency chlorosis in field-grown pear and peach: Physiological implications. Plant Soil 2008, 311, 161–172. [Google Scholar] [CrossRef]
- Franks, P.J.; Berry, J.A.; Lombardozzi, D.L.; Bonan, G.B. Stomatal function across temporal and spatial scales: Deep-time trends, land-atmosphere coupling and global models. Plant Physiol. 2017, 174, 583–602. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef] [PubMed]
- Dow, G.J.; Berry, J.A.; Bergmann, D.C. The physiological importance of developmental mechanisms that enforce proper stomatal spacing in a Rabidopsis thaliana. New Phytol. 2014, 201, 1205–1217. [Google Scholar] [CrossRef]
- Yin, Q.; Tian, T.; Kou, M.; Liu, P.; Wang, L.; Hao, Z.; Yue, M. The relationships between photosynthesis and stomatal traits on the loess plateau. Glob. Ecol. Conserv. 2020, 23, e01146. [Google Scholar] [CrossRef]
- Schmidt, S.B.; Eisenhut, M.; Schneider, A. Chloroplast transition metal regulation for efficient photosynthesis. Trends Plant Sci. 2020, 25, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.; Abadía, A.; Belkhodja, R.; Abadía, J. Iron deficiency-induced changes in the photosynthetic pigment composition of field-grown pear (Pyrus communis L) leaves. Plant, Cell Environ. 1994, 17, 1153–1160. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; Abadía, J. Photosynthesis, quenching of chlorophyll fluorescence and thermal energy dissipation in iron-deficient sugar beet leaves. Funct. Plant Biol. 1998, 25, 403–412. [Google Scholar] [CrossRef]
- Stocking, C.R. Iron deficiency and the structure and physiology of maize chloroplasts. Plant Physiol. 1975, 55, 626–631. [Google Scholar] [CrossRef]
- Larbi, A.; Abadía, A.; Abadía, J.; Morales, F. Down co-regulation of light absorption, photochemistry, and carboxylation in Fe-deficient plants growing in different environments. Photosynth. Res. 2006, 89, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Li, B.; Chen, H.; Song, C.; Meng, J.; Xi, Z.; Zhang, Z. Iron supply affects anthocyanin content and related gene expression in berries of Vitis vinifera cv. Cabernet sauvignon. Molecules 2017, 22, 283. [Google Scholar] [CrossRef]
- Khan, A.; Jalil, S.; Cao, H.; Tsago, Y.; Sunusi, M.; Chen, Z.; Shi, C.; Jin, X. The purple leaf (pl6) mutation regulates leaf color by altering the anthocyanin and chlorophyll contents in rice. Plants 2020, 9, 1477. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.O.; Gould, K.S. Anthocyanins in leaves: Light attenuators or antioxidants? Funct. Plant Biol. 2003, 30, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Maruta, T.; Noshi, M.; Tanouchi, A.; Tamoi, M.; Yabuta, Y.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. H2O2-triggered retrograde signaling from chloroplasts to nucleus plays specific role in response to stress. J. Biol. Chem. 2012, 287, 11717–11729. [Google Scholar] [CrossRef]
- Kim, J.-G.; Jung, S. Differential antioxidant mechanisms of rice plants in response to oxyfluorfen and paraquat. Weed Turfgrass Sci. 2013, 2, 254–259. [Google Scholar] [CrossRef]
- Jimenez, A.; Hernandez, J.A.; del Río, L.A.; Sevilla, F. Evidence for the presence of the ascorbate-glutathione cycle in mitochondria and peroxisomes of pea leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [CrossRef]
- Vasconcelos, M.W.; Clemente, T.; Grusak, M.A. Evaluation of constitutive iron reductase (AtFRO2) expression on mineral accumulation and distribution in soybean (Glycine max. L). Front. Plant Sci. 2014, 5, 112. [Google Scholar] [CrossRef]
- Ling, H.Q.; Bauer, P.; Bereczky, Z.; Keller, B.; Ganal, M. The tomato fer gene encoding a bHLH protein controls iron-uptake responses in roots. Proc. Natl. Acad. Sci. USA 2002, 99, 13938–13943. [Google Scholar] [CrossRef] [PubMed]
- Klatte, M.; Schuler, M.; Wirtz, M.; Fink-Straube, C.; Hell, R.; Bauer, P. The analysis of Arabidopsis nicotianamine synthase mutants reveals functions for nicotianamine in seed iron loading and iron deficiency responses. Plant Physiol. 2009, 150, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Pich, A.; Scholz, G. Translocation of copper and other micronutrients in tomato plants (Lycopersicon esculentum Mill.): Nicotianamine-stimulated copper transport in the xylem. J. Exp. Bot. 1996, 47, 41–47. [Google Scholar] [CrossRef]
- Yu, C.; Wang, Y.; Zhang, X.; Han, Z. A preliminary study on physiological and molecular effects of iron deficiency in Fuji/Chistock 1. J. Plant Nutr. 2014, 37, 2170–2178. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Xie, Q.; Akmakjian, G.Z.; Jobe, T.O.; Patel, A.; Stacey, M.G.; Song, L.; Demoin, D.W.; Jurisson, S.S.; Stacey, G.; et al. OPT3 Is a component of the iron-signaling network between leaves and roots and misregulation of OPT3 leads to an over-accumulation of cadmium in seeds. Mol. Plant 2014, 7, 1455–1469. [Google Scholar] [CrossRef]
- Jeong, J.; Merkovich, A.; Clyne, M.; Connolly, E.L. Directing iron transport in dicots: Regulation of iron acquisition and translocation. Curr. Opin. Plant Biol. 2017, 39, 106–113. [Google Scholar] [CrossRef]
- Maas, F.M.; van de Wetering, D.A.; van Beusichem, M.L.; Bienfait, H.F. Characterization of phloem iron and its possible role in the regulation of Fe-efficiency reactions. Plant Physiol. 1988, 87, 167–171. [Google Scholar] [CrossRef]
- García, M.J.; Romera, F.J.; Stacey, M.G.; Stacey, G.; Villar, E.; Alcántara, E.; Pérez-Vicente, R. Shoot to root communication is necessary to control the expression of iron-acquisition genes in Strategy I plants. Planta 2013, 237, 65–75. [Google Scholar] [CrossRef]
- Sági-Kazár, M.; Zelenyánszki, H.; Müller, B.; Cseh, B.; Gyuris, B.; Farkas, S.Z.; Fodor, F.; Tóth, B.; Kovács, B.; Koncz, A.; et al. Supraoptimal iron nutrition of Brassica napus plants suppresses the iron uptake of chloroplasts by down-regulating chloroplast ferric chelate reductase. Front. Plant Sci. 2021, 12, 748. [Google Scholar] [CrossRef]
- Msilini, N.; Zaghdoudi, M.; Govindachary, S.; Lachaâl, M.; Ouerghi, Z.; Carpentier, R. Inhibition of photosynthetic oxygen evolution and electron transfer from the quinone acceptor QA− to QB by iron deficiency. Photosynth. Res. 2011, 107, 247–256. [Google Scholar] [CrossRef]
- Morales, F.; Moise, N.; Quílez, R.; Abadía, A.; Abadía, J.; Moya, I. Iron deficiency interrupts energy transfer from a disconnected part of the antenna to the rest of Photosystem II. Photosynth. Res. 2001, 70, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Yadavalli, V.; Neelam, S.; Rao, A.S.; Reddy, A.R.; Subramanyam, R. Differential degradation of photosystem I subunits under iron deficiency in rice. J. Plant Physiol. 2012, 169, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Haldrup, A.; Naver, H.; Scheller, H.V. The interaction between plastocyanin and photosystem i is inefficient in transgenic Arabidopsis plants lacking the PSI-N subunit of photosystem. Plant J. 1999, 17, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Hippler, M.; Drepper, F.; Haehnel, W.; Rochaix, J.-D. The N-terminal domain of PsaF: Precise recognition site for binding and fast electron transfer from cytochrome c6 and plastocyanin to photosystem I of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1998, 95, 7339–7344. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Li, K.; Cai, X.; Wu, M.; Chen, G. Fe deficiency induced changes in rice (Oryza sativa L.) thylakoids. Environ. Sci. Pollut. Res. 2017, 24, 1380–1388. [Google Scholar] [CrossRef]
- Ma, F.; Zhang, X.; Zhu, X.; Li, T.; Zhan, J.; Chen, H.; He, C.; Wang, Q. Dynamic changes of IsiA-containing complexes during long-term iron deficiency in Synechocystis sp. PCC 6803. Mol. Plant 2017, 10, 143–154. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Name | Abbreviation | Locus ID |
|---|---|---|
| AtFerritin 1 | AtFER1 | AT5G01600 |
| AtFerric reductase oxidase 2 | AtFRO2 | AT1G01580 |
| AtPhotosystem I subunit N | AtPsAN | AT5G64040 |
| AtPhotosystem I subunit F | AtPsAF | AT1G31330 |
| AtOligopeptide transporters 3 | AtOPT3 | AT4G16370 |
| AtNicotianamine synthase 3 | AtNAS4 | AT1G09240.1 |
| AtNicotianamine synthase 4 | AtNAS3 | AT1G09240.2 |
| AtYellow stripe-like 5 | AtYSL5 | AT3G17650 |
| AtYellow stripe-like 7 | AtYSL7 | AT1G65730 |
| Gene Name | Forward Primers (5’ to 3’) | Reverse Primers (5’ to 3’) |
|---|---|---|
| Hm-AtFER1 | ATGCTTTATATGCAATGGAGTTGGC | CTGGTCGAAATGCCAAACTCCG |
| Hm-AtFRO2 | CTTCCTTCCGACACTATGGAGC | CAGAAGAAGAAAGCCTCTGGTG |
| Hm-AtPsAN | TCATTGACGACTACCTGGAAAAGAG | CCAGAAAACATTGGAACCGCAC |
| Hm-AtPsAF | ACTTGATAGTGAACGGAGACCAGCG | AGAAACTCTCTGTAGGCAGCAACGG |
| Hm-AtOPT3 | GCACTTCCAGGCTATCTATTCCC | TCCAGTAACAAACAGGGACGATG |
| Hm-AtNAS4 | AAAATCTGTGGTGAGGCTGAAG | TTGAGATGGGAAGAGGCAAGAAC |
| Hm-AtNAS3 | CGCTTCTCTTCTGGTTTCCTCTG | AAATAGACAAAACCTCGAACCCCTG |
| Hm-AtYSL5 | TATCAACAGCTTTCACACCCCTC | ACGGACATATCATTCCAACACCAAC |
| Hm-AtYSL7 | CATTTGGGTTGGATGATTGGATTTC | TTGAAAGAAACCCCACAAGAAACTG |
| Hmactin | GCCTGCCATGTATGTTGCCATC | CGGAATCCAGCACAATACCAGTTG |
| Fe Source (A) | pH (B) | Leaf | Shoot | Root | Stem Diameter (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | Area (cm2) | Length (cm) | Width (cm) | Length (cm) | Fresh Weight (mg) | Dry Weight (mg) | Number | Length (cm) | Fresh Weight (mg) | Dry Weight (mg) | |||
| Control | 4.70 | 9 b–d z | 10.3 a–c | 2.1 bc | 1.9 a | 3.8 a | 300.4 bc | 57.0 a–c | 3 d | 5.3 b–d | 51.2 cd | 3.0 b–d | 2.4 ab |
| 5.70 | 7 cd | 4.9 bc | 1.9 c | 1.3 a | 4.7 a | 179.7 c | 34.6 c | 3 d | 2.7 e | 21.7 d | 0.9 d | 2.7 ab | |
| 6.70 | 6 d | 3.6 c | 2.1 bc | 1.3 a | 4.4 a | 180.0 c | 42.4 bc | 3 d | 5.1 cd | 46.3 cd | 1.7 cd | 3.0 a | |
| FeSO4 H2O | 4.70 | 14 a | 13.2 ab | 3.2 a | 1.7 a | 5.6 a | 757.2 a | 86.9 a | 8 a | 7.0 a | 117.9 a | 6.9 a | 2.1 ab |
| 5.70 | 12 ab | 9.7 a–c | 2.8 ab | 1.5 a | 4.4 a | 377.6 bc | 56.9 a–c | 6 a–c | 5.7 a–d | 59.5 b–d | 4.1 a–c | 2.5 a | |
| 6.70 | 10 bc | 14.6 a | 2.7 a–c | 2.1 a | 4.6 a | 394.6 bc | 61.9 a–c | 7 ab | 6.7 a–c | 88.9 a–c | 5.2 ab | 1.7 ac | |
| Fe-EDTA | 4.70 | 12 ab | 12.1 a–c | 2.6 a–c | 1.7 a | 4.6 a | 424.1 b | 69.9 ab | 6 a–c | 6.9 ab | 107.0 ab | 6.9 a | 3.0 a |
| 5.70 | 11 ab | 15.0 a | 3.2 a | 1.8 a | 4.7 a | 309.1 bc | 51.4 bc | 5 bc | 5.2 cd | 60.9 b–d | 3.2 bc | 2.1 ab | |
| 6.70 | 9 b–d | 9.1 a–c | 2.6 a–c | 1.7 a | 5.1 a | 490.1 b | 63.3 a–c | 5 bc | 4.8 d | 50.5 cd | 4.4 a–c | 2.8 a | |
| F-test y | A | *** | * | ** | NS | NS | *** | NS | *** | *** | * | ** | ** |
| B | * | NS | NS | NS | NS | NS | * | NS | NS | NS | NS | NS | |
| A × B | NS | NS | NS | NS | NS | ** | ** | * | ** | ** | NS | ** | |
| Fe Source (A) | pH (B) | L* | a | b |
|---|---|---|---|---|
| Control | 4.70 | 44.7 ab z | −15.1 a | 15.7 ab |
| 5.70 | 48.2 a | −16.1 a | 19.9 a | |
| 6.70 | 48.3 a | −11.0 a | 21.9 a | |
| FeSO4 | 4.70 | 41.3 ab | −15.5 a | 6.3 bc |
| 5.70 | 37.7 bc | −18.0 a | 11.5 bc | |
| 6.70 | 37.8 bc | −14.1 a | 10.0 bc | |
| Fe-EDTA | 4.70 | 37.7 bc | −14.6 a | 3.0 c |
| 5.70 | 30.8 c | −11.0 a | 3.6 bc | |
| 6.70 | 37.8 bc | −12.0 a | 8.8 bc | |
| F-test y | A | *** | NS | * |
| B | NS | NS | NS | |
| A × B | *** | NS | NS |
| Correlation Coefficient | a * | b * | Chlorophyll a | Chlorophyll b | Anthocyanin | Carotenoid |
|---|---|---|---|---|---|---|
| L * | −0.159 | 0.646 ** | −0.580 ** | −0.511 ** | 0.010 | 0.137 |
| a * | −0.244 | −0.096 | −0.095 | 0.028 | 0.248 | |
| b * | −0.501 ** | −0.425 * | 0.039 | 0.040 | ||
| Chlorophyll a | 0.964 ** | −0.370 | −0.256 | |||
| Chlorophyll b | −0.411 * | −0.160 | ||||
| Anthocyanin | −0.038 |
| Fe Source (A) | pH (B) | Micronutrient (mg·g−1 DW) | Macronutrient (mg·g−1 DW) | Si (mg·g−1 DW) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fe | Mn | Zn | Cu | P | K | Ca | S | Mg | |||
| Control | 4.70 | 0.13 f z | 0.20 b | 0.06 c | 0.09 c | 9.63 e | 18.61 f | 15.02 e | 26.34 c | 2.84 e | 0.13 f |
| 5.70 | 0.10 f | 0.20 b | 0.04 d | 0.10 b | 10.08 e | 23.27 e | 33.16 d | 21.95 d | 3.23 e | 0.12 f | |
| 6.70 | 0.06 g | 0.47 a | 0.04 d | 0.28 a | 3.17 f | 6.69 g | 4.80 f | 10.03 g | 2.07 f | 0.07 g | |
| FeSO4 | 4.70 | 0.36 c | 0.10 e | 0.06 c | 0.02 fg | 27.88 b | 80.37 b | 161.29 a | 37.96 b | 6.59 c | 0.48 b |
| 5.70 | 0.25 de | 0.19 b | 0.10 b | 0.04 e | 18.21 c | 39.89 d | 94.00 b | 25.25 c | 12.63 a | 0.35 c | |
| 6.70 | 0.23 e | 0.14 d | 0.07 c | 0.07 d | 10.31 e | 21.13 ef | 14.85 e | 12.32 f | 3.45 e | 0.14 f | |
| Fe-EDTA | 4.70 | 1.12 a | 0.07 f | 0.18 a | 0.02 fg | 52.51 a | 134.80 a | 74.21 c | 63.45 a | 6.43 cd | 0.80 a |
| 5.70 | 0.49 b | 0.10 e | 0.09 b | 0.02 fg | 17.53 c | 47.04 c | 33.08 d | 14.12 e | 10.70 b | 0.23 d | |
| 6.70 | 0.26 d | 0.17 c | 0.07 c | 0.03 f | 14.67 d | 37.95 d | 37.71 d | 12.77 f | 5.88 d | 0.19 e | |
| F-test y | A | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| A × B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, J.; Guo, G.; Jeong, B.R. Iron Supplement-Enhanced Growth and Development of Hydrangea macrophylla In Vitro under Normal and High pH. Cells 2021, 10, 3151. https://doi.org/10.3390/cells10113151
Xiao J, Guo G, Jeong BR. Iron Supplement-Enhanced Growth and Development of Hydrangea macrophylla In Vitro under Normal and High pH. Cells. 2021; 10(11):3151. https://doi.org/10.3390/cells10113151
Chicago/Turabian StyleXiao, Jie, Ge Guo, and Byoung Ryong Jeong. 2021. "Iron Supplement-Enhanced Growth and Development of Hydrangea macrophylla In Vitro under Normal and High pH" Cells 10, no. 11: 3151. https://doi.org/10.3390/cells10113151
APA StyleXiao, J., Guo, G., & Jeong, B. R. (2021). Iron Supplement-Enhanced Growth and Development of Hydrangea macrophylla In Vitro under Normal and High pH. Cells, 10(11), 3151. https://doi.org/10.3390/cells10113151