
Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes


Abstract
1. Introduction
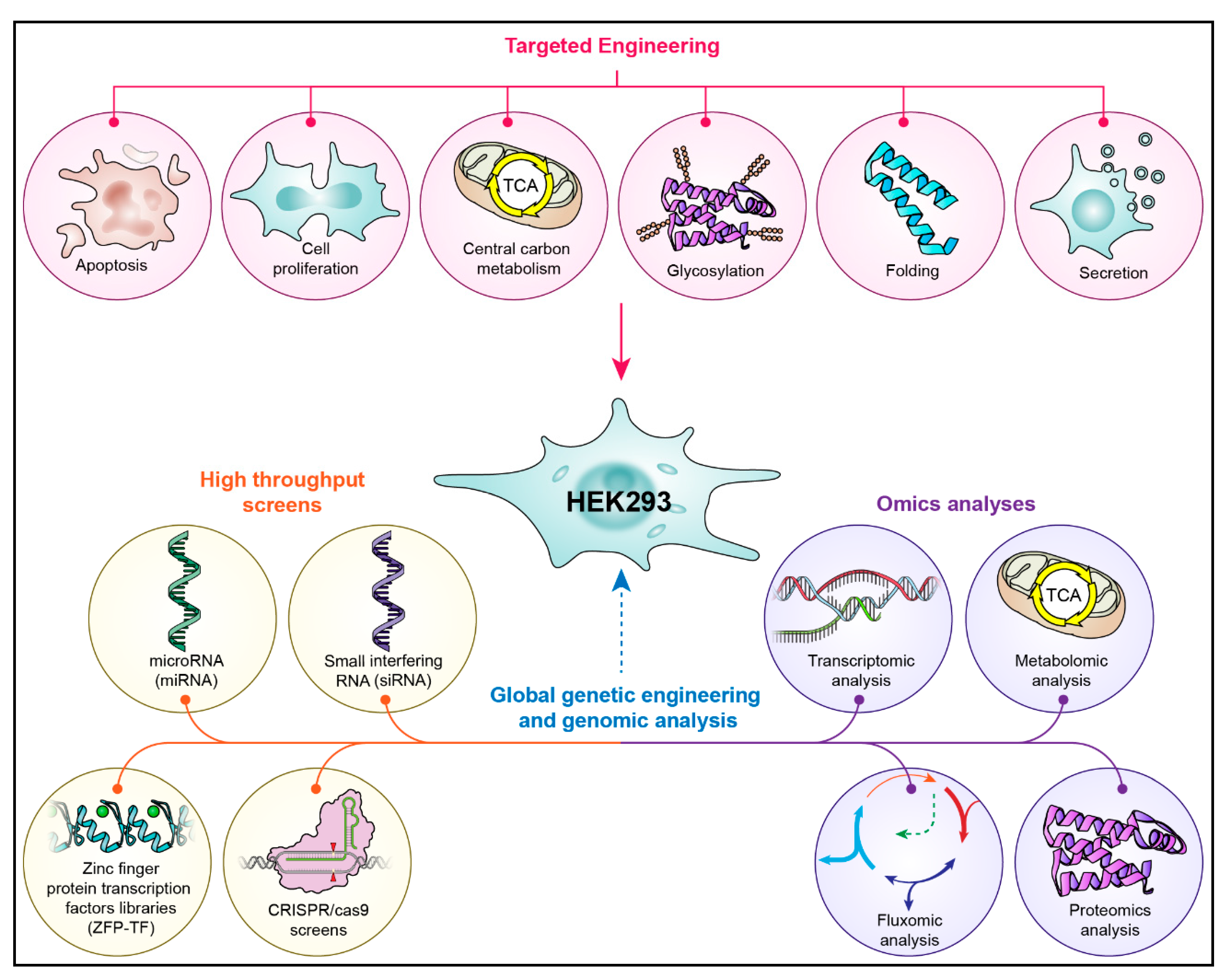
2. Strain/Genetic Engineering for Enhanced Recombinant Protein Expression in the HEK293 Cell Line
2.1. Targeted Genetic Engineering Approaches
2.1.1. Genes Involved in Cell Proliferation and Apoptosis
2.1.2. Genes Associated with Central Carbon Metabolism
2.1.3. Genes Involved in Protein Maturation and Processing
2.2. Global Genetic Engineering and Genomic Analysis Approaches
2.2.1. High Throughput RNA Interference (RNAi) Screens
2.2.2. Genome-Wide CRISPR/Cas9 Screens
2.2.3. Randomized Zinc Finger Protein Transcription factors (ZFP-TF) Libraries
2.2.4. Omics Analysis
3. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Graham, F.L.; Russell, W.C.; Smiley, J.; Nairn, R. Characteristics of a Human Cell Line Transformed by DNA from Human Adenovirus Type. J. Gen. Virol. 1977, 36, 59–72. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Boone, M.; Meuris, L.; Lemmens, I.; Van Roy, N.; Soete, A.; Reumers, J.; Moisse, M.; Plaisance, S.; Drmanac, R.T.; et al. Genome dynamics of the human embryonic kidney 293 lineage in response to cell biology manipulations. Nat. Commun. 2014, 5, 4767. [Google Scholar] [CrossRef]
- Kim, K.-S.; Kim, M.S.; Moon, J.H.; Jeong, M.S.; Kim, J.; Lee, G.M.; Myung, P.-K.; Hong, H.J. Enhancement of recombinant antibody production in HEK 293E cells by WPRE. Biotechnol. Bioprocess. Eng. 2009, 14, 633–638. [Google Scholar] [CrossRef]
- Jäger, V.; Büssow, K.; Wagner, A.; Weber, S.; Hust, M.; Frenzel, A.; Schirrmann, T. High level transient production of recombinant antibodies and antibody fusion proteins in HEK293 cells. BMC Biotechnol. 2013, 13, 52. [Google Scholar] [CrossRef]
- Geisse, S.; Fux, C. Recombinant Protein Production by Transient Gene Transfer into Mammalian Cells. In Methods in Enzymology; Burgess, R.R., Deutscher, M.P., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 223–238. [Google Scholar]
- Casademunt, E.; Martinelle, K.; Jernberg, M.; Winge, S.; Tiemeyer, M.; Biesert, L.; Knaub, S.; Walter, O.; Schröder, C. The first recombinant human coagulation factor VIII of human origin: Human cell line and manufacturing characteristics. Eur. J. Haematol. 2012, 89, 165–176. [Google Scholar] [CrossRef]
- Stepanenko, A.; Dmitrenko, V. HEK293 in cell biology and cancer research: Phenotype, karyotype, tumorigenicity, and stress-induced genome-phenotype evolution. Gene 2015, 569, 182–190. [Google Scholar] [CrossRef]
- Bloom, F.R.; Price, P.; Lao, G.; Xia, J.L.; Crowe, J.H.; Battista, J.R.; Helm, R.F.; Slaughter, S.; Potts, M. Engineering mammalian cells for solid-state sensor applications. Biosens. Bioelectron. 2001, 16, 603–608. [Google Scholar] [CrossRef]
- Yuan, J.; Xu, W.W.; Jiang, S.; Yu, H.; Poon, H.F. The Scattered Twelve Tribes of HEK293. Biomed. Pharmacol. J. 2018, 11, 621–623. [Google Scholar] [CrossRef]
- Malm, M.; Saghaleyni, R.; Lundqvist, M.; Giudici, M.; Chotteau, V.; Field, R.; Varley, P.G.; Hatton, D.; Grassi, L.; Svensson, T.; et al. Evolution from adherent to suspension: Systems biology of HEK293 cell line development. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Swiech, K.; Picanco-Castro, V.; Covas, D. Human cells: New platform for recombinant therapeutic protein production. Protein Expr. Purif. 2012, 84, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Smart, T.G. HEK293 cell line: A vehicle for the expression of recombinant proteins. J. Pharmacol. Toxicol. Methods 2005, 51, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Dumont, A.J.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R.R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2016, 36, 1110–1122. [Google Scholar] [CrossRef]
- Thompson, C.M.; Petiot, E.; Mullick, A.; Aucoin, M.G.; Henry, O.; Kamen, A.A. Critical assessment of influenza VLP production in Sf9 and HEK293 expression systems. BMC Biotechnol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Le Ru, A.; Jacob, D.; Transfiguracion, J.; Ansorge, S.; Henry, O.; Kamen, A.A. Scalable production of influenza virus in HEK-293 cells for efficient vaccine manufacturing. Vaccine 2010, 28, 3661–3671. [Google Scholar] [CrossRef]
- Grieger, J.C.; Soltys, S.M.; Samulski, R.J. Production of Recombinant Adeno-associated Virus Vectors Using Suspension HEK293 Cells and Continuous Harvest of Vector from the Culture Media for GMP FIX and FLT1 Clinical Vector. Mol. Ther. 2016, 24, 287–297. [Google Scholar] [CrossRef]
- Chahal, P.S.; Schulze, E.; Tran, R.; Montes, J.; Kamen, A.A. Production of adeno-associated virus (AAV) serotypes by transient transfection of HEK293 cell suspension cultures for gene delivery. J. Virol. Methods 2014, 196, 163–173. [Google Scholar] [CrossRef]
- Ansorge, S.; Lanthier, S.; Transfiguracion, J.; Durocher, Y.; Henry, O.; Kamen, A. Development of a scalable process for high-yield lentiviral vector production by transient transfection of HEK293 suspension cultures. J. Gene Med. 2009, 11, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.; Pak, J.E.; Pedersen, B.; Bang, L.J.; Zhang, L.B.; Ngaw, S.M.; Green, R.G.; Sharma, V.; Stroud, R.M. Efficient expression screening of human membrane proteins in transiently transfected Human Embryonic Kidney 293S cells. Methods 2011, 55, 273–280. [Google Scholar] [CrossRef]
- Andréll, J.; Tate, C. Overexpression of membrane proteins in mammalian cells for structural studies. Mol. Membr. Biol. 2012, 30, 52–63. [Google Scholar] [CrossRef]
- Ayuso, E.; Mingozzi, F.; Montane, J.; Leon, X.; Anguela, X.; Haurigot, V.A.; Edmonson, A.S.; Africa, L.; Zhou, S.; High, K.; et al. High AAV vector purity results in serotype- and tissue-independent enhancement of transduction efficiency. Gene Ther. 2009, 17, 503–510. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Wagner, K.; Robinson, N.; Sabatino, D.; Margaritis, P.; Xiao, W.; Herzog, R.W. Optimized Production of High-Titer Recombinant Adeno-Associated Virus in Roller Bottles. BioTechniques 2003, 34, 184–189. [Google Scholar] [CrossRef]
- Dalton, A.C.; Barton, W.A. Over-expression of secreted proteins from mammalian cell lines. Protein Sci. 2014, 23, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Zolotukhin, S.; Potter, M.; Zolotukhin, I.; Sakai, Y.; Loiler, S.; Fraites, T.J.; Chiodo, A.V.; Phillipsberg, T.; Muzyczka, N.; Hauswirth, W.W.; et al. Production and purification of serotype 1, 2, and 5 recombinant adeno-associated viral vectors. Methods 2002, 28, 158–167. [Google Scholar] [CrossRef]
- Potter, M.; Lins, B.; Mietzsch, M.; Heilbronn, R.; Van Vliet, K.; Chipman, P.; Agbandje-McKenna, M.; Cleaver, B.D.; Clément, N.; Byrne, B.J.; et al. A simplified purification protocol for recombinant adeno-associated virus vectors. Mol. Ther. Methods Clin. Dev. 2014, 1, 14034. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.B.; Drury, E.J.; Morton, C.L.; Spence, Y.; Lockey, T.D.; Nathwani, A.C.; Davidoff, A.M.; Meagher, M.M. Distribution of AAV8 particles in cell lysates and culture media changes with time and is dependent on the recombinant vector. Mol. Ther. Methods Clin. Dev. 2016, 3, 16015. [Google Scholar] [CrossRef]
- Emmerling, V.V.; Pegel, A.; Milian, E.G.; Venereo-Sanchez, A.; Kunz, M.; Wegele, J.; Kamen, A.A.; Kochanek, S.; Hoerer, M. Rational plasmid design and bioprocess optimization to enhance recombinant adeno-associated virus (AAV) productivity in mammalian cells. Biotechnol. J. 2016, 11, 290–297. [Google Scholar] [CrossRef]
- Strobel, B.; Zuckschwerdt, K.; Zimmermann, G.; Mayer, C.; Eytner, R.; Rechtsteiner, P.; Kreuz, S.; Lamla, T. Standardized, Scalable, and Timely Flexible Adeno-Associated Virus Vector Production Using Frozen High-Density HEK-293 Cell Stocks and CELLdiscs. Hum. Gene Ther. Methods 2019, 30, 23–33. [Google Scholar] [CrossRef]
- Powers, A.; Piras, B.; Clark, R.K.; Lockey, T.D.; Meagher, M.M. Development and Optimization of AAV hFIX Particles by Transient Transfection in an iCELLis®Fixed-Bed Bioreactor. Hum. Gene Ther. Methods 2016, 27, 112–121. [Google Scholar] [CrossRef]
- Ghaderi, D.; Taylor, E.R.; Padler-Karavani, V.; Diaz, S.; Varki, A. Implications of the presence of N-glycolylneuraminic acid in recombinant therapeutic glycoproteins. Nat. Biotechnol. 2010, 28, 863–867. [Google Scholar] [CrossRef]
- Fliedl, L.; Grillari, J.; Grillari-Voglauer, R. Human cell lines for the production of recombinant proteins: On the horizon. N. Biotechnol. 2015, 32, 673–679. [Google Scholar] [CrossRef]
- Böhm, E.; Seyfried, B.K.; Dockal, M.; Graninger, M.; Hasslacher, M.; Neurath, M.; Konetschny, C.; Matthiessen, P.F.; Mitterer, A.; Scheiflinger, F. Differences in N-glycosylation of recombinant human coagulation factor VII derived from BHK, CHO, and HEK293 cells. BMC Biotechnol. 2015, 15, 1–15. [Google Scholar] [CrossRef]
- Kronman, C.; Chitlaru, T.; Elhanany, E.; Velan, B.; Shafferman, A. hierarchy of post-translational modifications involved in the circulatory longevity of glycoproteins. Demonstration of concerted contributions of glycan sialylation and subunit assembly to the pharmacokinetic behavior of bovine acetylcholinesterase. J. Biol. Chem. 2000, 275, 29488–29502. [Google Scholar] [CrossRef]
- Berkner, K. Expression of recombinant vitamin K-dependent proteins in mammalian cells: Factors IX and VII. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1993; Volume 222, pp. 450–477. [Google Scholar]
- Kannicht, C.; Ramström, M.; Kohla, G.; Tiemeyer, M.; Casademunt, E.; Walter, O.; Sandberg, H. Characterisation of the post-translational modifications of a novel, human cell line-derived recombinant human factor VIII. Thromb. Res. 2013, 131, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.T.; Toby, G.; Lu, Q.; Liu, T.; Kulman, J.D.; Low, S.C.; Bitonti, A.J.; Pierce, G.F. Biochemical and functional characterization of a recombinant monomeric factor VIII–Fc fusion protein. J. Thromb. Haemost. 2013, 11, 132–141. [Google Scholar] [CrossRef] [PubMed]
- McCue, J.; Osborne, D.; Dumont, J.; Peters, R.; Mei, B.; Pierce, G.F.; Kobayashi, K.; Euwart, D. Validation of the manufacturing process used to produce long-acting recombinant factor IX Fc fusion protein. Haemophilia 2014, 20, e327–e335. [Google Scholar] [CrossRef] [PubMed]
- McCue, J.; Kshirsagar, R.; Selvitelli, K.; Lu, Q.; Zhang, M.; Mei, B.; Peters, R.; Pierce, G.F.; Dumont, J.; Raso, S.; et al. Manufacturing process used to produce long-acting recombinant factor VIII Fc fusion protein. Biologicals 2015, 43, 213–219. [Google Scholar] [CrossRef]
- Goh, J.B.; Ng, S.K. Impact of host cell line choice on glycan profile. Crit. Rev. Biotechnol. 2018, 38, 851–867. [Google Scholar] [CrossRef]
- Havenga, M.; Holterman, L.; Melis, I.; Smits, S.; Kaspers, J.; Heemskerk, E.; van der Vlugt, R.; Koldijk, M.; Schouten, G.; Hateboer, G.; et al. Serum-free transient protein production system based on adenoviral vector and PER.C6 technology: High yield and preserved bioactivity. Biotechnol. Bioeng. 2008, 100, 273–283. [Google Scholar] [CrossRef]
- Wong, H.-L.; Wang, M.-X.; Cheung, P.-T.; Yao, K.-M.; Chan, B.P. A 3D collagen microsphere culture system for GDNF-secreting HEK293 cells with enhanced protein productivity. Biomaterials 2007, 28, 5369–5380. [Google Scholar] [CrossRef]
- Chin, C.L.; Goh, J.B.; Srinivasan, H.; Liu, K.I.; Gowher, A.; Shanmugam, R.; Lim, H.L.; Choo, M.; Tang, W.Q.; Tan, A.H.-M.; et al. A human expression system based on HEK293 for the stable production of recombinant erythropoietin. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Mensah, E.O.; Guo, X.-Y.; Gao, X.-D.; Fujita, M. Establishment of DHFR-deficient HEK293 cells for high yield of therapeutic glycoproteins. J. Biosci. Bioeng. 2019, 128, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Baek, M.; Lee, G.M. Comprehensive characterization of dihydrofolate reductase-mediated gene amplification for the establishment of recombinant human embryonic kidney 293 cells producing monoclonal antibodies. Biotechnol. J. 2021, 16, e2000351. [Google Scholar] [CrossRef]
- Yu, D.Y.; Lee, S.Y.; Lee, G.M. Glutamine synthetase gene knockout-human embryonic kidney 293E cells for stable production of monoclonal antibodies. Biotechnol. Bioeng. 2018, 115, 1367–1372. [Google Scholar] [CrossRef]
- Liste-Calleja, L.; Lecina, M.; Cairó, J.J. HEK293 cell culture media study: Increasing cell density for different bioprocess applications. BMC Proc. 2013, 7, P51. [Google Scholar] [CrossRef]
- Ooi, A.; Wong, A.; Esau, L.; Lemtiri-Chlieh, F.; Gehring, C. A Guide to Transient Expression of Membrane Proteins in HEK-293 Cells for Functional Characterization. Front. Physiol. 2016, 7, 300. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.E.; Staley, E.M.; Mayginnes, J.P.; Pintel, D.J.; Tullis, G.E. Transfection of mammalian cells using linear polyethylenimine is a simple and effective means of producing recombinant adeno-associated virus vectors. J. Virol. Methods 2006, 138, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Román, R.; Miret, J.; Scalia, F.; Casablancas, A.; Lecina, M.; Cairó, J.J. Enhancing heterologous protein expression and secretion in HEK293 cells by means of combination of CMV promoter and IFNα2 signal peptide. J. Biotechnol. 2016, 239, 57–60. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Huang, Z.; Wen, W.; Wu, A.; Wang, C.; Niu, L. Enhancing Protein Expression in HEK-293 Cells by Lowering Culture Temperature. PLoS ONE 2015, 10, e0123562. [Google Scholar] [CrossRef]
- Cervera, L.; Fuenmayor, J.; González-Domínguez, I.; Gutiérrez-Granados, S.; Segura, M.M.; Gòdia, F. Selection and optimization of transfection enhancer additives for increased virus-like particle production in HEK293 suspension cell cultures. Appl. Microbiol. Biotechnol. 2015, 99, 9935–9949. [Google Scholar] [CrossRef]
- Loignon, M.; Perret, S.; Kelly, J.; Boulais, D.; Cass, B.; Bisson, L.; Afkhamizarreh, F.; Durocher, Y. Stable high volumetric production of glycosylated human recombinant IFNalpha2b in HEK293 cells. BMC Biotechnol. 2008, 8, 65. [Google Scholar] [CrossRef]
- Spidel, J.L.; Vaessen, B.; Chan, Y.Y.; Grasso, L.; Kline, J.B. Rapid high-throughput cloning and stable expression of antibodies in HEK293 cells. J. Immunol. Methods 2016, 439, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, A.; Hust, M.; Schirrmann, T. Expression of Recombinant Antibodies. Front. Immunol. 2013, 4, 217. [Google Scholar] [CrossRef]
- Patil, A.A.; Bhor, S.A.; Rhee, W.J. Cell death in culture: Molecular mechanisms, detections, and inhibition strategies. J. Ind. Eng. Chem. 2020, 91, 37–53. [Google Scholar] [CrossRef]
- Jaluria, P.; Betenbaugh, M.; Konstantopoulos, K.; Shiloach, J. Enhancement of cell proliferation in various mammalian cell lines by gene insertion of a cyclin-dependent kinase homolog. BMC Biotechnol. 2007, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Roobol, A.; Smith, M.E.; Carden, M.J.; Hershey, J.W.; Willis, A.E.; Smales, C.M. Engineered transient and stable overexpression of translation factors eIF3i and eIF3c in CHOK1 and HEK293 cells gives enhanced cell growth associated with increased c-Myc expression and increased recombinant protein synthesis. Metab. Eng. 2020, 59, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Backliwal, G.; Hildinger, M.; Chenuet, S.; Wulhfard, S.; De Jesus, M.; Wurm, F.M. Rational vector design and multi-pathway modulation of HEK 293E cells yield recombinant antibody titers exceeding 1 g/l by transient transfection under serum-free conditions. Nucleic Acids Res. 2008, 36, e96. [Google Scholar] [CrossRef]
- Werner, N.S.; Weber, W.; Fussenegger, M.; Geisse, S. A gas-inducible expression system in HEK.EBNA cells applied to controlled proliferation studies by expression of P27kip1. Biotechnol. Bioeng. 2007, 96, 1155–1166. [Google Scholar] [CrossRef]
- Arena, A.T.; Chou, B.; Harms, P.D.; Wong, A.W. An anti-apoptotic HEK293 cell line provides a robust and high titer platform for transient protein expression in bioreactors. mAbs 2019, 11, 977–986. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Shan, L.; Zhao, J.; Mao, Q.; Xia, H. Generation of apoptosis-resistant HEK293 cells with CRISPR/Cas mediated quadruple gene knockout for improved protein and virus production. Biotechnol. Bioeng. 2017, 114, 2539–2549. [Google Scholar] [CrossRef]
- Sauerwald, T.M.; Betenbaugh, M.J.; Oyler, G.A. Inhibiting apoptosis in mammalian cell culture using the caspase inhibitor XIAP and deletion mutants. Biotechnol. Bioeng. 2002, 77, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.S.; Al-Rubeai, M. The effect of Bcl-2, YAMA, and XIAP over-expression on apoptosis and adenovirus production in HEK293 cell line. Biotechnol. Bioeng. 2009, 104, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Sauerwald, T.M.; Oyler, G.A.; Betenbaugh, M.J. Study of caspase inhibitors for limiting death in mammalian cell culture. Biotechnol. Bioeng. 2002, 81, 329–340. [Google Scholar] [CrossRef]
- Abbasi-Malati, Z.; Amiri, F.; Mohammadipour, M.; Roudkenar, M.H. HEK293 Cells Overexpressing Nuclear Factor E2-Related Factor-2 Improve Expression of Recombinant Coagulation Factor VII. Mol. Biotechnol. 2019, 61, 317–324. [Google Scholar] [CrossRef]
- Formas-Oliveira, A.S.; Basílio, J.S.; Rodrigues, A.F.; Coroadinha, A.S. Overexpression of ER Protein Processing and Apoptosis Regulator Genes in Human Embryonic Kidney 293 Cells Improves Gene Therapy Vectors Production. Biotechnol. J. 2020, 15, e1900562. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, L.; Jenkins, H.A.; Butler, M. Glucose and glutamine metabolism of a murine B-lymphocyte hybridoma grown in batch culture. Appl. Biochem. Biotechnol. 1993, 43, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Glacken, M.W.; Adema, E.; Sinskey, A.J. Mathematical descriptions of hybridoma culture kinetics: I. Initial metabolic rates. Biotechnol. Bioeng. 1988, 32, 491–506. [Google Scholar] [CrossRef]
- Lanks, K.W.; Li, P.-W. End products of glucose and glutamine metabolism by cultured cell lines. J. Cell. Physiol. 1988, 135, 151–155. [Google Scholar] [CrossRef]
- Petch, D.; Butler, M. Profile of energy metabolism in a murine hybridoma: Glucose and glutamine utilization. J. Cell. Physiol. 1994, 161, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Goergen, J.L.; Marc, A.; Engasser, J.M. Influence of lactate and ammonia on the death rate of hybridoma. In Animal Cell Technology; Spier, R.E., Griffiths, J.B., Berthold, W., Eds.; Butterworth-Heinemann: Oxford, UK, 1994; pp. 161–163. [Google Scholar]
- Ozturk, S.S.; Riley, M.R.; Palsson, B.O. Effects of ammonia and lactate on hybridoma growth, metabolism, and antibody production. Biotechnol. Bioeng. 1992, 39, 418–431. [Google Scholar] [CrossRef]
- Newland, M.; Greenfield, P.F.; Reid, S. Hybridoma growth limitations: The roles of energy metabolism and ammonia production. Cytotechnology 1990, 3, 215–229. [Google Scholar] [CrossRef]
- Gawlitzek, M.; Valley, U.; Wagner, R. Ammonium ion and glucosamine dependent increases of oligosaccharide complexity in recombinant glycoproteins secreted from cultivated BHK-21 cells. Biotechnol. Bioeng. 1998, 57, 518–528. [Google Scholar] [CrossRef]
- Goochee, C.F.; Gramer, M.J.; Andersen, D.C.; Bahr, J.B.; Rasmussen, J.R. The Oligosaccharides of Glycoproteins: Bioprocess Factors Affecting Oligosaccharide Structure and their Effect on Glycoprotein Properties. Bio/Technology 1991, 9, 1347–1355. [Google Scholar] [CrossRef]
- Grammatikos, S.I.; Valley, U.; Nimtz, M.; Conradt, H.S.; Wagner, R. Intracellular UDP-N-Acetylhexosamine Pool Affects N-Glycan Complexity: A Mechanism of Ammonium Action on Protein Glycosylation. Biotechnol. Prog. 1998, 14, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Elias, C.B.; Carpentier, E.; Durocher, Y.; Bisson, L.; Wagner, R.; Kamen, A. Improving Glucose and Glutamine Metabolism of Human HEK 293 and Trichoplusia ni Insect Cells Engineered To Express a Cytosolic Pyruvate Carboxylase Enzyme. Biotechnol. Prog. 2003, 19, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Karengera, E.; Robotham, A.; Kelly, J.; Durocher, Y.; De Crescenzo, G.; Henry, O. Altering the central carbon metabolism of HEK293 cells: Impact on recombinant glycoprotein quality. J. Biotechnol. 2017, 242, 73–82. [Google Scholar] [CrossRef]
- Vallée, C.; Durocher, Y.; Henry, O. Exploiting the metabolism of PYC expressing HEK293 cells in fed-batch cultures. J. Biotechnol. 2014, 169, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Henry, O.; Durocher, Y. Enhanced glycoprotein production in HEK-293 cells expressing pyruvate carboxylase. Metab. Eng. 2011, 13, 499–507. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rodrigues, A.F.; Guerreiro, M.; Formas-Oliveira, A.S.; Fernandes, P.; Blechert, A.-K.; Genzel, Y.; Alves, P.; Hu, W.S.; Coroadinha, A.S. Increased titer and reduced lactate accumulation in recombinant retrovirus production through the down-regulation of HIF1 and PDK. Biotechnol. Bioeng. 2016, 113, 150–162. [Google Scholar] [CrossRef]
- Aricescu, A.R.; Owens, R.J. Expression of recombinant glycoproteins in mammalian cells: Towards an integrative approach to structural biology. Curr. Opin. Struct. Biol. 2013, 23, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Yamane-Ohnuki, N.; Satoh, M. Production of therapeutic antibodies with controlled fucosylation. mAbs 2009, 1, 230–236. [Google Scholar] [CrossRef]
- Lepenies, B.; Seeberger, P.H. Simply better glycoproteins: Reducing the glycan heterogeneity of recombinant proteins may improve the efficacy of biopharmaceuticals. Nat. Biotechnol. 2014, 32, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Kozak, S.; Bloch, Y.; De Munck, S.; Mikula, A.; Bento, I.; Savvides, S.N.; Meijers, R. Homogeneously N-glycosylated proteins derived from the GlycoDelete HEK293 cell line enable diffraction-quality crystallogenesis. Acta Crystallogr. Sect. D Struct. Biol. 2020, 76, 1244–1255. [Google Scholar] [CrossRef]
- Ren, W.-W.; Jin, Z.-C.; Dong, W.; Kitajima, T.; Gao, X.-D.; Fujita, M. Glycoengineering of HEK293 cells to produce high-mannose-type N-glycan structures. J. Biochem. 2019, 166, 245–258. [Google Scholar] [CrossRef]
- Reeves, P.J.; Callewaert, N.; Contreras, R.; Khorana, H.G. Structure and function in rhodopsin: High-level expression of rhodopsin with restricted and homogeneous N-glycosylation by a tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proc. Natl. Acad. Sci. USA 2002, 99, 13419–13424. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.-X.; Ren, W.-W.; Li, Y.; Yang, G.; Gao, X.-D.; Fujita, M. Cell engineering for the production of hybrid-type N-glycans in HEK293 cells. J. Biochem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.-C.; Kitajima, T.; Dong, W.; Huang, Y.-F.; Ren, W.-W.; Guan, F.; Chiba, Y.; Gao, X.-D.; Fujita, M. Genetic disruption of multiple α1,2-mannosidases generates mammalian cells producing recombinant proteins with high-mannose–type N-glycans. J. Biol. Chem. 2018, 293, 5572–5584. [Google Scholar] [CrossRef]
- Meuris, L.; Santens, F.; Elson, G.; Festjens, N.; Boone, M.; Dos Santos, A.; Devos, S.; Rousseau, F.; Plets, E.; Houthuys, E.; et al. GlycoDelete engineering of mammalian cells simplifies N-glycosylation of recombinant proteins. Nat. Biotechnol. 2014, 32, 485–489. [Google Scholar] [CrossRef]
- Zhang, P.; Tan, D.L.; Heng, D.; Wang, T.; Mariati; Yang, Y.; Song, Z. A functional analysis of N-glycosylation-related genes on sialylation of recombinant erythropoietin in six commonly used mammalian cell lines. Metab. Eng. 2010, 12, 526–536. [Google Scholar] [CrossRef]
- Raymond, C.; Robotham, A.; Kelly, J.; Lattová, E.; Perreault, H.; Durocher, Y. Production of Highly Sialylated Monoclonal Antibodies. Glycosylation 2012, 397–418. [Google Scholar]
- Nakatsukasa, K.; Brodsky, J.L. The Recognition and Retrotranslocation of Misfolded Proteins from the Endoplasmic Reticulum. Traffic 2008, 9, 861–870. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Jiang, J.-H.; Li, S.-T. Functional expression of human α7 nicotinic acetylcholine receptor in human embryonic kidney 293 cells. Mol. Med. Rep. 2016, 14, 2257–2263. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-M.; Jin, D.-Y.; Camire, R.M.; Stafford, D.W. Vitamin K epoxide reductase significantly improves carboxylation in a cell line overexpressing factor X. Blood 2005, 106, 3811–3815. [Google Scholar] [CrossRef]
- Wajih, N.; Owen, J.; Wallin, R. Enhanced functional recombinant factor VII production by HEK 293 cells stably transfected with VKORC1 where the gamma-carboxylase inhibitor calumenin is stably suppressed by shRNA transfection. Thromb. Res. 2008, 122, 405–410. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barnes, L.M.; Dickson, A.J. Mammalian cell factories for efficient and stable protein expression. Curr. Opin. Biotechnol. 2006, 17, 381–386. [Google Scholar] [CrossRef]
- Becker, E.; Florin, L.; Pfizenmaier, K.; Kaufmann, H. An XBP-1 dependent bottle-neck in production of IgG subtype antibodies in chemically defined serum-free Chinese hamster ovary (CHO) fed-batch processes. J. Biotechnol. 2008, 135, 217–223. [Google Scholar] [CrossRef]
- Borth, N.; Mattanovich, D.; Kunert, R.; Katinger, H. Effect of Increased Expression of Protein Disulfide Isomerase and Heavy Chain Binding Protein on Antibody Secretion in a Recombinant CHO Cell Line. Biotechnol. Prog. 2008, 21, 106–111. [Google Scholar] [CrossRef]
- Davis, R.; Schooley, K.; Rasmussen, B.; Thomas, J.; Reddy, P. Effect of PDI Overexpression on Recombinant Protein Secretion in CHO Cells. Biotechnol. Prog. 2000, 16, 736–743. [Google Scholar] [CrossRef]
- Ku, S.C.; Ng, D.T.; Yap, M.G.; Chao, S.-H. Effects of overexpression of X-box binding protein 1 on recombinant protein production in Chinese hamster ovary and NS0 myeloma cells. Biotechnol. Bioeng. 2008, 99, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Smales, C.; Dinnis, D.; Stansfield, S.; Alete, D.; Sage, E.; Birch, J.; Racher, A.; Marshall, C.; James, D. Comparative proteomic analysis of GS-NS0 murine myeloma cell lines with varying recombinant monoclonal antibody production rate. Biotechnol. Bioeng. 2004, 88, 474–488. [Google Scholar] [CrossRef]
- Smith, J.D.; Tang, B.C.; Robinson, A.S. Protein disulfide isomerase, but not binding protein, overexpression enhances secretion of a non-disulfide-bonded protein in yeast. Biotechnol. Bioeng. 2004, 85, 340–350. [Google Scholar] [CrossRef]
- Tigges, M.; Fussenegger, M. Xbp1-based engineering of secretory capacity enhances the productivity of Chinese hamster ovary cells. Metab. Eng. 2006, 8, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Lang, T.; Südhof, T.C. Membrane Fusion. Cell 2003, 112, 519–533. [Google Scholar] [CrossRef]
- Wickner, W.; Schekman, R. Membrane fusion. Nat. Struct. Mol. Biol. 2008, 15, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.-W.; Guetg, C.; Tigges, M.; Fussenegger, M. The vesicle-trafficking protein munc18b increases the secretory capacity of mammalian cells. Metab. Eng. 2010, 12, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Schiller, L.T.; Diaz, N.L.; Ferreira, R.R.; Böker, K.O.; Gruber, J. Enhanced Production of Exosome-Associated AAV by Overexpression of the Tetraspanin CD. Mol. Ther. Methods Clin. Dev. 2018, 9, 278–287. [Google Scholar] [CrossRef]
- Peng, R.-W.; Fussenegger, M. Molecular engineering of exocytic vesicle traffic enhances the productivity of Chinese hamster ovary cells. Biotechnol. Bioeng. 2009, 102, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Chen, Y.-C.; Betenbaugh, M.J.; Martin, S.E.; Shiloach, J. MiRNA mimic screen for improved expression of functional neurotensin receptor from HEK 293 cells. Biotechnol. Bioeng. 2015, 112, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Inwood, S.; Betenbaugh, M.J.; Lal, M.; Shiloach, J. Genome-Wide High-Throughput RNAi Screening for Identification of Genes Involved in Protein Production. Methods Mol. Biol. 2018, 1850, 209–219. [Google Scholar] [CrossRef]
- Inwood, S.; Abaandou, L.; Betenbaugh, M.; Shiloach, J. Improved protein expression in HEK293 cells by over-expressing miR-22 and knocking-out its target gene, HIPK. N. Biotechnol. 2020, 54, 28–33. [Google Scholar] [CrossRef]
- Lwa, T.R.; Tan, C.H.; Lew, Q.J.; Chu, K.L.; Tan, J.; Lee, Y.Y.; Chao, S.-H. Identification of cellular genes critical to recombinant protein production using a Gaussia luciferase-based siRNA screening system. J. Biotechnol. 2010, 146, 160–168. [Google Scholar] [CrossRef]
- Xiao, S.; Chen, Y.C.; Buehler, E.; Mandal, S.; Mandal, A.; Betenbaugh, M.; Park, M.H.; Martin, S.; Shiloach, J. Genome-scale RNA interference screen identifies antizyme 1 (OAZ1) as a target for improvement of recombinant protein production in mammalian cells. Biotechnol. Bioeng. 2016, 113, 2403–2415. [Google Scholar] [CrossRef] [PubMed]
- Abaandou, L.; Shiloach, J. Knocking out Ornithine Decarboxylase Antizyme 1 (OAZ1) Improves Recombinant Protein Expression in the HEK293 Cell Line. Med. Sci. 2018, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Abaandou, L.; Sharma, A.K.; Shiloach, J. Knockout of the caspase 8-associated protein 2 gene improves recombinant protein expression in HEK293 cells through up-regulation of the cyclin-dependent kinase inhibitor 2A gene. Biotechnol. Bioeng. 2021, 118, 186–198. [Google Scholar] [CrossRef]
- Dietmair, S.; Hodson, M.P.; Quek, L.-E.; Timmins, N.E.; Gray, P.; Nielsen, L.K. A Multi-Omics Analysis of Recombinant Protein Production in Hek293 Cells. PLoS ONE 2012, 7, e43394. [Google Scholar] [CrossRef]
- Rodrigues, A.F.; Formas-Oliveira, A.; Bandeira, V.; Alves, P.; Hu, W.; Coroadinha, A. Metabolic pathways recruited in the production of a recombinant enveloped virus: Mining targets for process and cell engineering. Metab. Eng. 2013, 20, 131–145. [Google Scholar] [CrossRef]
- Lavado-García, J.; Jorge, I.; Cervera, L.; Vázquez, J.; Gòdia, F. Multiplexed Quantitative Proteomic Analysis of HEK293 Provides Insights into Molecular Changes Associated with the Cell Density Effect, Transient Transfection, and Virus-Like Particle Production. J. Proteome Res. 2020, 19, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Lavado-García, J.; Díaz-Maneh, A.; Canal-Paulí, N.; Pérez-Rubio, P.; Gòdia, F.; Cervera, L. Metabolic engineering of HEK293 cells to improve transient transfection and cell budding of HIV-1 virus-like particles. Biotechnol. Bioeng. 2021, 118, 1630–1644. [Google Scholar] [CrossRef]
- Inwood, S.; Buehler, E.; Betenbaugh, M.; Lal, M.; Shiloach, J. Identifying HIPK1 as Target of miR-22-3p Enhancing Recombinant Protein Production From HEK 293 Cell by Using Microarray and HTP siRNA Screen. Biotechnol. J. 2018, 13. [Google Scholar] [CrossRef]
- Shalem, O.; Sanjana, N.E.; Hartenian, E.; Shi, X.; Scott, D.A.; Mikkelsen, T.S.; Heckl, D.; Ebert, B.L.; Root, D.E.; Doench, J.G.; et al. Genome-Scale CRISPR-Cas9 Knockout Screening in Human Cells. Science 2014, 343, 84–87. [Google Scholar] [CrossRef]
- Popa, S.; Villeneuve, J.; Stewart, S.; Perez Garcia, E.; Petrunkina Harrison, A.; Moreau, K. Genome-wide CRISPR screening identifies new regulators of glycoprotein secretion. Wellcome Open Res. 2019, 4, 119. [Google Scholar] [CrossRef]
- Tian, S.; Muneeruddin, K.; Choi, M.Y.; Tao, L.; Bhuiyan, R.H.; Ohmi, Y.; Furukawa, K.; Furukawa, K.; Boland, S.; Shaffer, S.A.; et al. Genome-wide CRISPR screens for Shiga toxins and ricin reveal Golgi proteins critical for glycosylation. PLoS Biol. 2018, 16, e2006951. [Google Scholar] [CrossRef]
- Park, K.-S.; Lee, D.-K.; Lee, H.; Lee, Y.; Jang, Y.-S.; Kim, Y.H.; Yang, H.-Y.; Lee, S.-I.; Seol, W.; Kim, J.-S. Phenotypic alteration of eukaryotic cells using randomized libraries of artificial transcription factors. Nat. Biotechnol. 2003, 21, 1208–1214. [Google Scholar] [CrossRef]
- Park, K.-S.; Seol, W.; Yang, H.-Y.; Lee, S.-I.; Kim, S.K.; Kwon, R.J.; Kim, E.-J.; Roh, Y.-H.; Seong, B.L.; Kim, J.-S. Identification and Use of Zinc Finger Transcription Factors That Increase Production of Recombinant Proteins in Yeast and Mammalian Cells. Biotechnol. Prog. 2008, 21, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, G.M. Functional expression of human pyruvate carboxylase for reduced lactic acid formation of Chinese hamster ovary cells (DG44). Appl. Microbiol. Biotechnol. 2007, 76, 659–665. [Google Scholar] [CrossRef]
- Vazquez-Lombardi, R.; Nevoltris, D.; Luthra, A.; Schofield, P.; Zimmermann, C.; Christ, D. Transient expression of human antibodies in mammalian cells. Nat. Protoc. 2018, 13, 99–117. [Google Scholar] [CrossRef]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G. Viral vector platforms within the gene therapy landscape. Signal. Transduct. Target. Ther. 2021, 6, 1–24. [Google Scholar] [CrossRef]
- Pulix, M.; Lukashchuk, V.; Smith, D.C.; Dickson, A.J. Molecular characterization of HEK293 cells as emerging versatile cell factories. Curr. Opin. Biotechnol. 2021, 71, 18–24. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| HEK 293 Variant | Derivation | Commercially Available? | Desired Characteristic(s) | Year of Derivation |
|---|---|---|---|---|
| HEK293 | Transformation of Human embryonic kidney cells with sheared fragments of adenovirus type 5 (Ad5) DNA, selected for immortalization. | Yes | Parental HEK293 cell line | 1973 |
| HEK293S | Adapted for suspension growth | Yes | Grows in suspension in modified minimal Eagle’s medium | 1984 |
| HEK293T | Stable transfection of the HEK 293 cell line with a plasmid encoding a temperature-sensitive mutant of the SV40 large T antigen | Yes | Amplification of vectors containing the SV40 ori, considerably increasing the protein expression levels during transient transfection. | Before 1985 |
| HEK293FT | Fast-growing variant of HEK293T. Expresses the SV40 large T antigen from the pCMVSPORT6TAg.neo plasmid. | Designed for lentiviral production. | ||
| HEK293F | Cloned from the HEK293 cell line and adapted to commercial medium | Yes | Fast growth and high transfectivity. Growth in chemically defined medium | 2014 |
| HEK293H | Cloned from HEK293 to select a clone with good adherence during plaque assays. Later adapted to growth in serum-free medium (SFM) (GIBCO 293H) | Yes | Fast growth in SFM, good adherence during plaque assays, superior transfection efficiencies and a high level of protein expression | 1993 |
| HEK293E(EBNA) | Expresses the EBNA-1 protein for episomal replication of oriP harboring plasmids | Yes | Amplification of vectors containing oriP, considerably increasing the protein expression levels during transient transfection. | |
| HEK2936E | This cell line is transfected with EBNA1t, a truncated version of EBV EBNA1 lacking the Gly-Gly-Ala repeats region | Has an enhanced ability to produce recombinant protein compared to HEK293-EBNA | ||
| HEK293FTM | Derived from 293 cells by stable transfection of an FRT-site containing plasmid and of a TetR expression plasmid. | Used for fast and easy generation of a stably transfected cell pool by co-transfecting a Flp-InTM expression vector containing a gene of interest and a Flp recombinase expression vector. | Before 2001 | |
| HEK293SG | Ricin toxin-resistant clone derived from HEK293S by ethylmethanesulfonate (EMS). | Yes | Lacks N-acetylglucosaminyltransferase I activity (encoded by the MGAT1 gene) and accordingly predominantly modifies glycoproteins with the Man5GlcNAc2 N-glycan. HEK293SG is used to produce homogenously N-glycosylated proteins | 2001-2002 |
| HEK293SGGD (Glycodelete) | Derives from 293SG through expression of a Golgi targeted form of endoT, an endoglycosidase from the fungus Trichoderma reesei | HEK293SGGD is mainly used to produce proteins for glycosylation studies and structural analysis | 2010 | |
| HEK293A | Subclone of the HEK293 cells with a relatively flat morphology | Facilitates the initial production, amplification and titering of replication-incompetent adenovirus | ||
| HEK293MSR | Genetically engineered from HEK293 to express the human macrophage scavenger receptor | Strongly adheres to standard tissue culture plates for dependable results |
| Gene | Function | Modification | Outcome | Recombinant Protein(s) | Reference |
|---|---|---|---|---|---|
| Cyclin-dependent kinase like 3 (CDKL3) | Promotes cell cycle G1-S transition in mammalian cells | Overexpression | 19% improvement in growth rate, 20% improvement in r. protein production, 8% greater maximum viable cell density | Secreted recombinant adipocyte complement-related protein of 30kDa | [56] |
| Cytochrome c oxidase subunit 15 (COX15) | Counteracts apoptosis due to its involvement in the Synthesis of heme | 13% improvement in growth rate and 11% improvement in r. protein production | |||
| Eukaryotic initiation factor 3 subunit i (EIF3I) | Subunit of the eIF3 targets and initiates translation of a subset of mRNAs involved in cell proliferation, including cell cycling, differentiation, and apoptosis. | Overexpression | Faster growth and increased c-Myc expression. | Renilla and Firefly luciferase | [57] |
| Cyclin-dependent kinase inhibitor 1B (CDKN1B) | Binds to and prevents the activation of cyclin E-CDK2 or cyclin D-CDK4 complexes, and thus controls the cell cycle progression at G1 | Inducible over-expression | 5.9-fold increase in recombinant protein expression | Recombinant secreted alkaline phosphatase (SEAP) | [59] |
| Cyclin-dependent kinase inhibitor 1A (CDKN1A) | Binds to and inhibits the activity of cyclin-cyclin-dependent kinase2 or cyclin-dependent kinase4 complexes, and thus functions as a regulator of cell cycle progression at G1 | Overexpression (Coupled with rational vector design and valporic acid treatment) | Approximately 27-fold increase in recombinant protein production, from 40 mg/L to 1.1 g/L in transient system | IgG | [58] |
| Cyclin-dependent Kinase inhibitor 2C (CDKN2C) | Interacts with CDK4 or CDK6, and prevent the activation of the CDK kinases, thus functioning as a cell growth regulator that controls cell cycle G1 progression | ||||
| Acidic fibroblast growth factor (FGF1) | Involved in a variety of biological processes, including embryonic development, cell growth, morphogenesis, tissue repair, tumor growth and invasion |
| Gene | Function | Modification | Outcome | Recombinant Protein(s) | Reference |
|---|---|---|---|---|---|
| X-linked inhibitor of apoptosis (XIAP) | Inhibits the apoptosis executioners caspase3, caspase 7, and caspase 9 | Overexpression | 8-fold higher number of viable cells., extension of stationary phase by 48 h | N/A | [62] [63] |
| B-cell lymphoma 2 (BCL2) | Inhibits the pro-apoptotic proteins Bak and Bax | Overexpression Over-expression | Extension of stationary phase by 48 h 53% increase in volumetric production | N/A Lentiviral vectors | [63] [66] |
| Caspase 3 (CASP3) , Caspase 6 (CASP6) , Caspase 7 (CASP7) and Allograft inflammatory factor 1 (AIF1) | Executioners of apoptosis in the caspase dependent pathway | Quadruple Knockout | Higher expression levels of r. proteins and higher packaging efficiency of recombinant viral particles | Bax, TRAIL, Luciferase Lentivirus | [61] |
| Bcl-2-associated X protein (BAX) and BCL2 Antagonist/Killer 1 (BAK) | Activation of caspase proteins | Double knockout | Resistance to apoptosis and sheer stress, 40% improvement in recombinant protein titers | Human IgG1 antibody | [60] |
| CrmA (viral) | Apoptosis inhibitor | Overexpression | Resistance to numerous apoptotic insults | N/A | [64] |
| Nuclear factor erythroid 2-related factor 2 (NRF2) | Transcription factor that upregulates antioxidant response elements (AREs)-mediated expression of antioxidant enzyme and cytoprotective proteins. | Overexpression | Higher growth rate, more resistant to oxidative stress. 1.7-fold improvement in r. protein expression | Recombinant coagulation factor VII | [65] |
| Gene | Function | Modification | Outcome | Recombinant Protein(s) | Reference |
|---|---|---|---|---|---|
| Pyruvate Carboxylase (PC) | Catalyzes the conversion of pyruvate to oxaloacetate | Overexpression | Increased maximal cell density, viability, and glucose utilization. | Adenovirus | [77] |
| Decreased lactate and ammonia production. | Recombinant Interferon α2b | [78] | |||
| Improved glucose metabolism. | [79] | ||||
| Improved total volumetric recombinant protein production | [80] | ||||
| Pyruvate dehydrogenase kinase (PDK) | Inactivates the pyruvate dehydrogenase enzyme complex, increasing the conversion of pyruvate to lactate in the cytosol. | Knockdown | 30-fold increase in specific productivity of infectious viral particles and a 4-fold decrease in lactate production | Retroviral vectors | [81] |
| Hypoxia inducible factor 1 (HIF1A) | Activates an over-expression cascade of glycolytic enzymes and glycolysis-related genes, including pyruvate dehydrogenase kinase (PDK) and lactate dehydrogenase (LDH) | Knockdown |
| Gene | Function | Modification | Outcome | Recombinant Protein(s) | Reference |
|---|---|---|---|---|---|
| N-acetylglucosaminyltransferase I (GnTI) protein encoded by the MGAT1 gene | Processing of high-mannose to hybrid and complex N-glycans | Knockout | Inability to synthesize complex glycans. Only the Man5GlcNAc2 N-Glycan present | Rhodopsin | [87] |
| Endo-β-N-acetylglucosaminidase from Hypocrea jecorina (endoT8) fused to the Golgi targeting domain of the human β-galactoside-α-2,6-sialyltransferase 1 (in GnTI-/- cells) | Catalyzes the hydrolysis of mannose modifications to produce free oligosaccharides. Catalyzes the transfer of sialic acid from CMP-sialic acid to galactose-containing substrates | Overexpression | Improved homogeneity in glycan expression | Granulocyte macrophage colony-stimulating factor (GM-CSF), monoclonal anti-CD20 antibody GA101 | [90] |
| Golgi mannosidases MAN1A1 | Catalyzes the removal of 3 distinct mannose residues from peptide-bound Man (9)-GlcNAc (2) oligosaccharides. | Triple knockout | Production of simple Man9GlcNAc2 and Man8GlcNAc2 structures of high-Man–type glycans; limited complex-type N-glycans at relatively low abundance | Lysosomal enzymes, α-galactosidase-A (GLA) and lysosomal acid lipase (LIPA) | [89] |
| MAN1A2 | Progressively trim alpha-1,2-linked mannose residues from Man(9)GlcNAc(2) to produce Man(5)GlcNAc(2). | ||||
| ER mannosidaseMAN1B1 | Plays an important role in the disposal of misfolded glycoproteins | ||||
| MAN1C1 and MGAT1 (quadruple and quintuple knockouts based on the triple knockout by Jin and colleagues [89] | Trim alpha-1,2-linked mannose residues from Man(9)GlcNAc(2) to produce first Man(8)GlcNAc(2) then Man(6)GlcNAc and a small amount of Man(5)GlcNAc. Essential for the conversion of high-mannose to hybrid and complex N-glycans | Quadruple and quintuple knockouts | Elimination of all the hybrid-type and complex-type N-glycans with only the high-mannose-type N-glycans present | Lysosomal acid lipase (LIPA) and immunoglobulin G1 (IgG1) | [86] |
| Sialyl transferases, ST6GAL1, ST3GAL3 and ST3GAL4 ST6GAL1 | Add sialic acid to the terminal portions of the N- or O-linked sugar chains of glycoproteins. | Overexpression Overexpression | Enhanced sialylation Increased α2,6 sialylation | EPO Trastuzumab F243A mutant antibody | [91] [92] |
| Golgi mannosidases MAN2A1 and MAN2A2 | Control conversion of high mannose to complex N-glycans | Double Knockout | Hybrid-type N-glycans only | Lysosomal acid lipase (LIPA) and Fc of immunoglobulin G1 (IgG1) | [88] |
| Fucosyltransferase 8 (FUT8) | Catalyzes the addition of fucose in alpha 1-6 linkage to the first GlcNAc residue, next to the peptide chains in N-glycans. | Triple knockout with MAN2A1 and MAN2A2 | Hybrid-type N-glycans only without core fucosylation | Lysosomal acid lipase (LIPA) and Fc of immunoglobulin G1 (IgG1) | [88] |
| Gene | Function | Modification | Outcome | Recombinant Protein | Reference |
|---|---|---|---|---|---|
| Human resistance to inhibitors of cholinesterase 3 (RIC3) | Molecular chaperone of nicotinic acetylcholine receptors | Overexpression | Expression of high levels of functional receptor protein | α7 nicotinic acetyl cholinesterase receptor | [95] |
| X-box binding protein (XBP1) | Induces the unfolded protein response by upregulating target genes encoding ER chaperones and ER-associated degradation (ERAD) components to enhance the capacity of productive folding and degradation mechanism, respectively. | Overexpression | Improvement in volumetric and cell specific gamma retroviral productivity | Gamma retroviral vectors | [66] |
| Protein Disulfide Isomerase Family A Member 2 (PDIA2) | Folding of nascent proteins in the endoplasmic reticulum by forming disulfide bonds through its thiol isomerase, oxidase and reductase activity | ||||
| Heat shock protein family A (Hsp70) member 5 (HSPA5) | Plays a key role in protein folding and quality control in the endoplasmic reticulum lumen. Involved in the correct folding of proteins and degradation of misfolded proteins. | ||||
| Vitamin K Epoxide Reductase Complex Subunit 1 (VKORC1) | Carboxylation of glutamic acid residues in some blood-clotting proteins, including factor VII, factor IX, and factor X | Overexpression | Improved γ-carboxylation | rhFVII | [97] |
| Calumenin (CALU) | Inhibitor of γ-carboxylase | shRNA inhibition | Up to 7.5-fold increase in production yield of active enzyme | rhFX | [96] |
| Gene | Function | Modification | Outcome | Recombinant Protein | Reference |
|---|---|---|---|---|---|
| SNARE receptor coding genes, SNAP-23 and VAMP8 | Mediate/catalyze the fusion of vesicles to target membranes during exocytosis | Overexpression | Improved recombinant protein expression | SEAP, SAMY | [110] |
| SM protein coding genes MUNC18 and sly1 | Regulate SNARE complex assembly and then cooperate with SNAREs to stimulate membrane fusion. | ||||
| Tetraspanin CD9 | Play a role in many cellular processes including exosome biogenesis. | Overexpression | Up to 26% improvement in virus production | Exosome-Associated AAV | [109] |
| Type of Screen/Analysis | Genes/Genomic Elements/ Pathways Identified | Recombinant Protein(s) | Ref. | Genes/Genomic Elements Identified or Confirmed in Follow-up Study(ies) | Modification | Outcome | Ref. |
|---|---|---|---|---|---|---|---|
| miRNA | hsa-miR-22-5p, hsa-miR-18a-5p, hsa-miR-22-3p, hsa-miR-429 and hsa miR-2110 | Neurotensin receptor, cytoplasmic luciferase and secreted glypican −3 hFc-fusion protein (GPC3) | [111] | HIPK1 | Knockdown | 3.2-fold higher Luciferase and 2.3-fold higher GPC3 expression | [112] |
| miRNA-22-3b | Stable overexpression | 2.4-fold higher luciferase activity | [113] | ||||
| HIPK1 | Knockout | 4.7-fold higher luciferase expression | |||||
| siRNA | CCAAT/enhancer binding protein gamma (CEBPG) | Recombinant EPO, interferon γ and monoclonal antibody | [114] | ||||
| siRNA | INTS1, HNRHPC, CASP8AP2, OAZ1 and PPP2R1A | Luciferase, GPC3, neurotensin receptor type I and serotonin transporter | [115] | OAZ1 | Knockout | 5-fold higher specific luciferase and 2.5-fold higher specific SEAP production | [116] |
| CASP8AP2 | Knockout | 7-fold increase in specific luciferase and 2.5-fold increase in specific SEAP expression | [117] | ||||
| Multi-omics analysis (transcriptomic, fluxomic, metabolomic) | FABP5, RETNLB, pyruvate carboxylase (PC) X-box binding protein (XBP1), HSPA5 and HERPUD1 | A heavy chain variable region fused to the Fc region of a human IgG (dAb-Fc) | [118] | ||||
| Transcriptomic analysis | Amino acid metabolism, carbohydrate metabolism, polyamine metabolism, glutathione metabolism, nucleotide metabolism, the pentose phosphate pathway and lipid metabolism | A murine leukemia virus-based vector. | [119] | ||||
| Proteomic analysis | NEDD8, NEDD4L, ESCRT1, lipid biosynthesis, glycosphingolipids metabolism, calcium regulation, oxidant detoxification, xenobiotic metabolism, peptidase activity, DNA detoxification, endocytosis and late endosomal-related pathways | HIV-gag VLP | [120] | NEDD8 | Overexpression | 1.5-fold increase in VLP production. | [121] |
| NEDD4L | Overexpression | 3.3- fold increase in VLP production. | |||||
| UGCG | Overexpression | 2.9- fold increase in VLP production. | |||||
| CIT | Overexpression | 2.4-fold increase in VLP production. | |||||
| CNP | ShRNA knockdown | 2.7-fold increase in VLP production |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abaandou, L.; Quan, D.; Shiloach, J. Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes. Cells 2021, 10, 1667. https://doi.org/10.3390/cells10071667
Abaandou L, Quan D, Shiloach J. Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes. Cells. 2021; 10(7):1667. https://doi.org/10.3390/cells10071667
Chicago/Turabian StyleAbaandou, Laura, David Quan, and Joseph Shiloach. 2021. "Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes" Cells 10, no. 7: 1667. https://doi.org/10.3390/cells10071667
APA StyleAbaandou, L., Quan, D., & Shiloach, J. (2021). Affecting HEK293 Cell Growth and Production Performance by Modifying the Expression of Specific Genes. Cells, 10(7), 1667. https://doi.org/10.3390/cells10071667