
Function and Biomarkers of the Blood-Brain Barrier in a Neonatal Germinal Matrix Haemorrhage Model

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Germinal Matrix Haemorrhage (GMH)
2.3. Functional Blood-Brain Barrier Assessment, Visible Tracers
2.4. Visualisation of Tracers
2.5. Quantification of Leaking Blood Vessels and Hematoma Size
2.6. Functional Blood-Brain Barrier Assessment by Sucrose Extravasation
2.7. Fluorescent Immunohistochemistry and 3D Imaging
2.8. Levels of Tight-Junction Proteins in CSF and Plasma after GMH
2.9. Statistics and Graph
3. Results
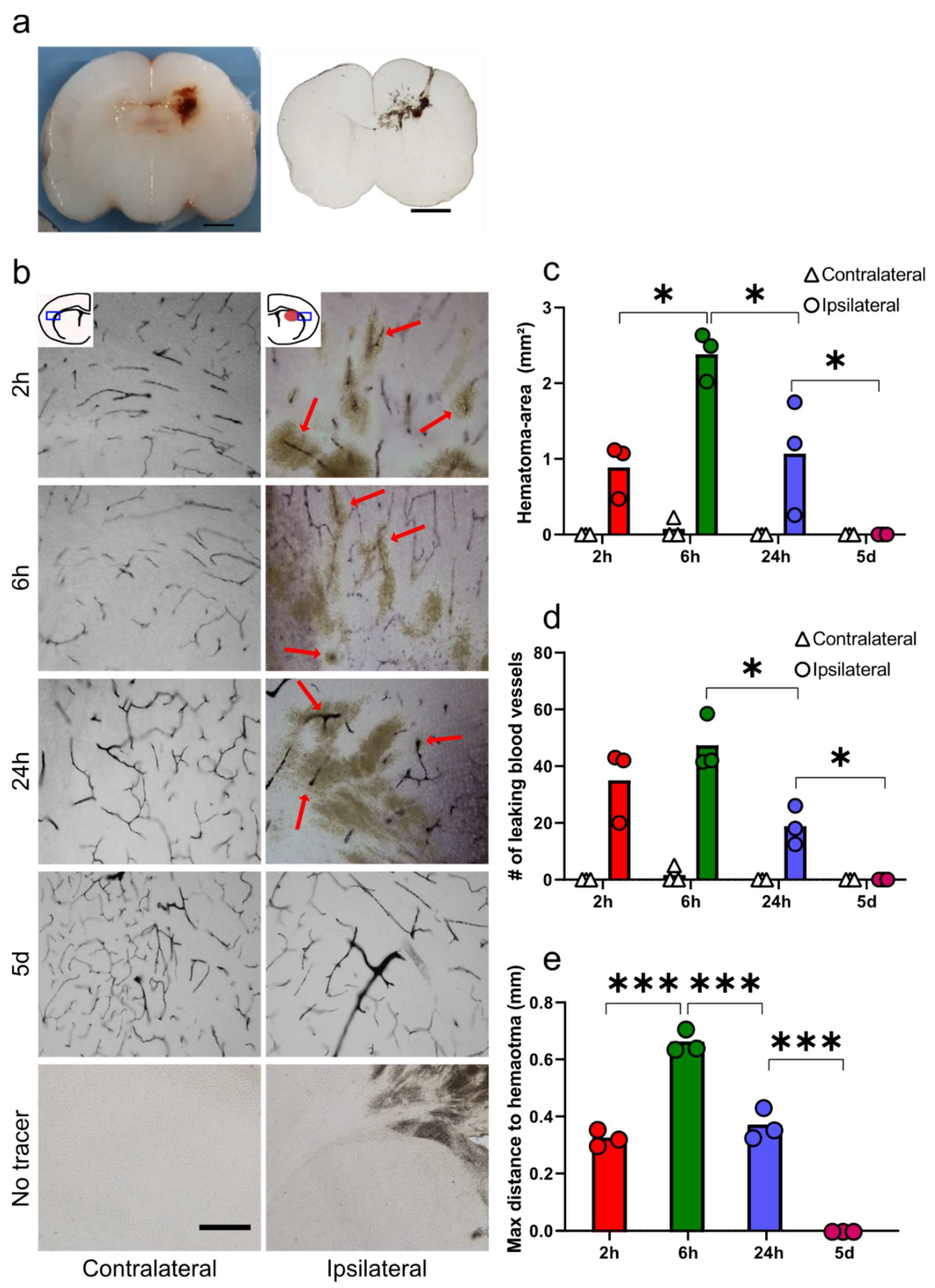
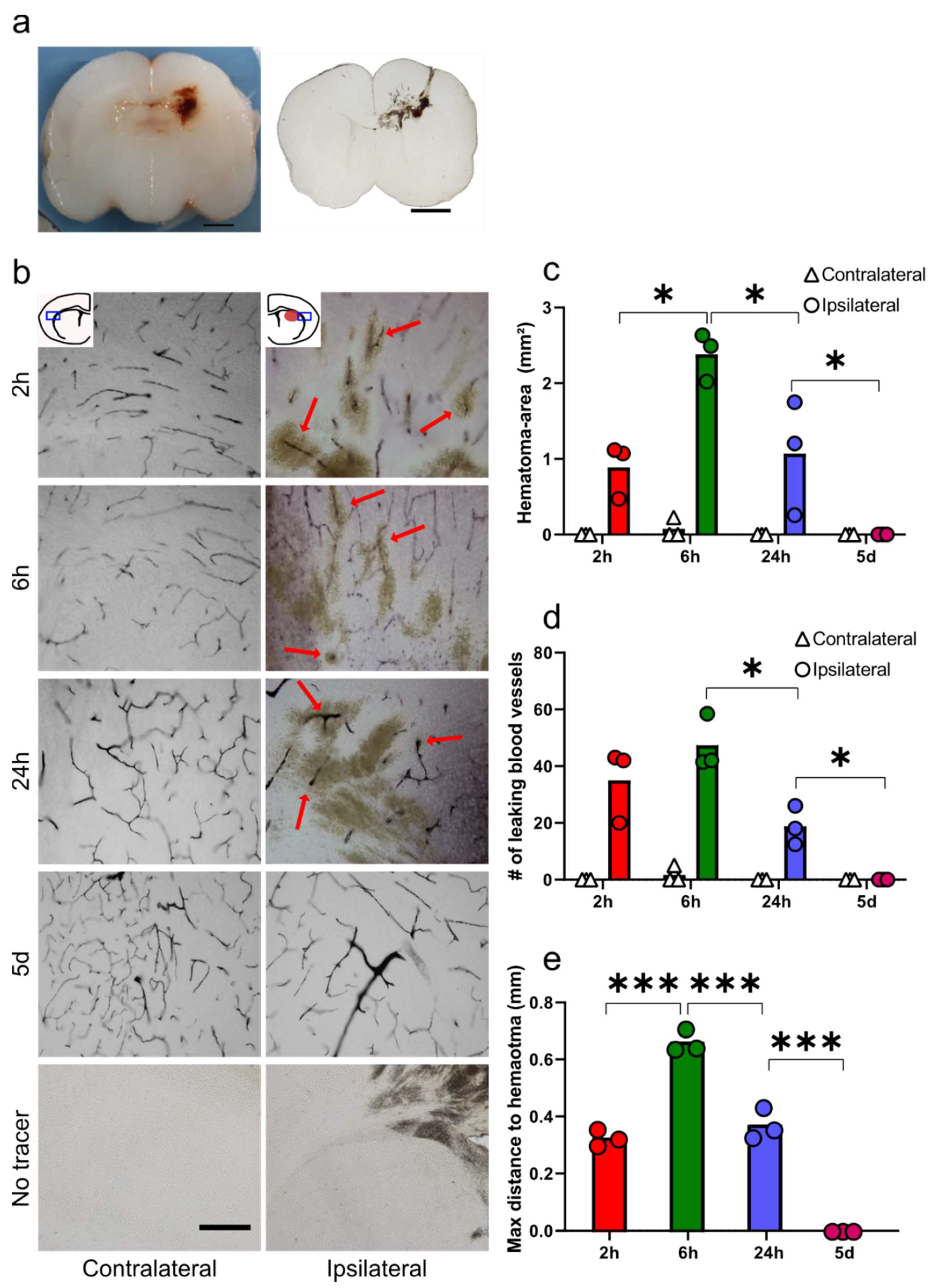
3.1. Functional Blood-Brain Barrier Assessment Using Molecular Tracers
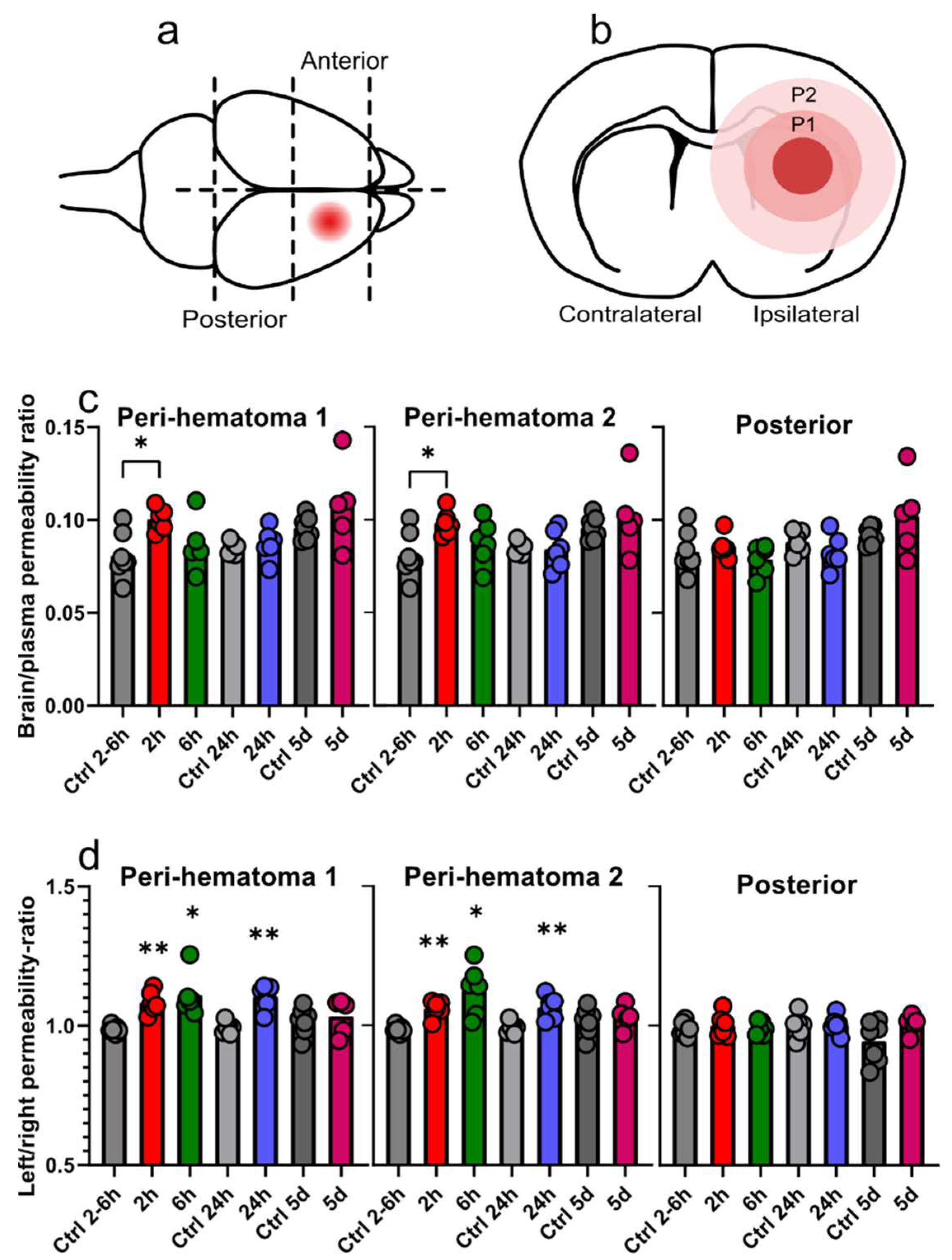
3.2. Regional Sucrose Permeability Across Blood-Brain Barrier after GMH
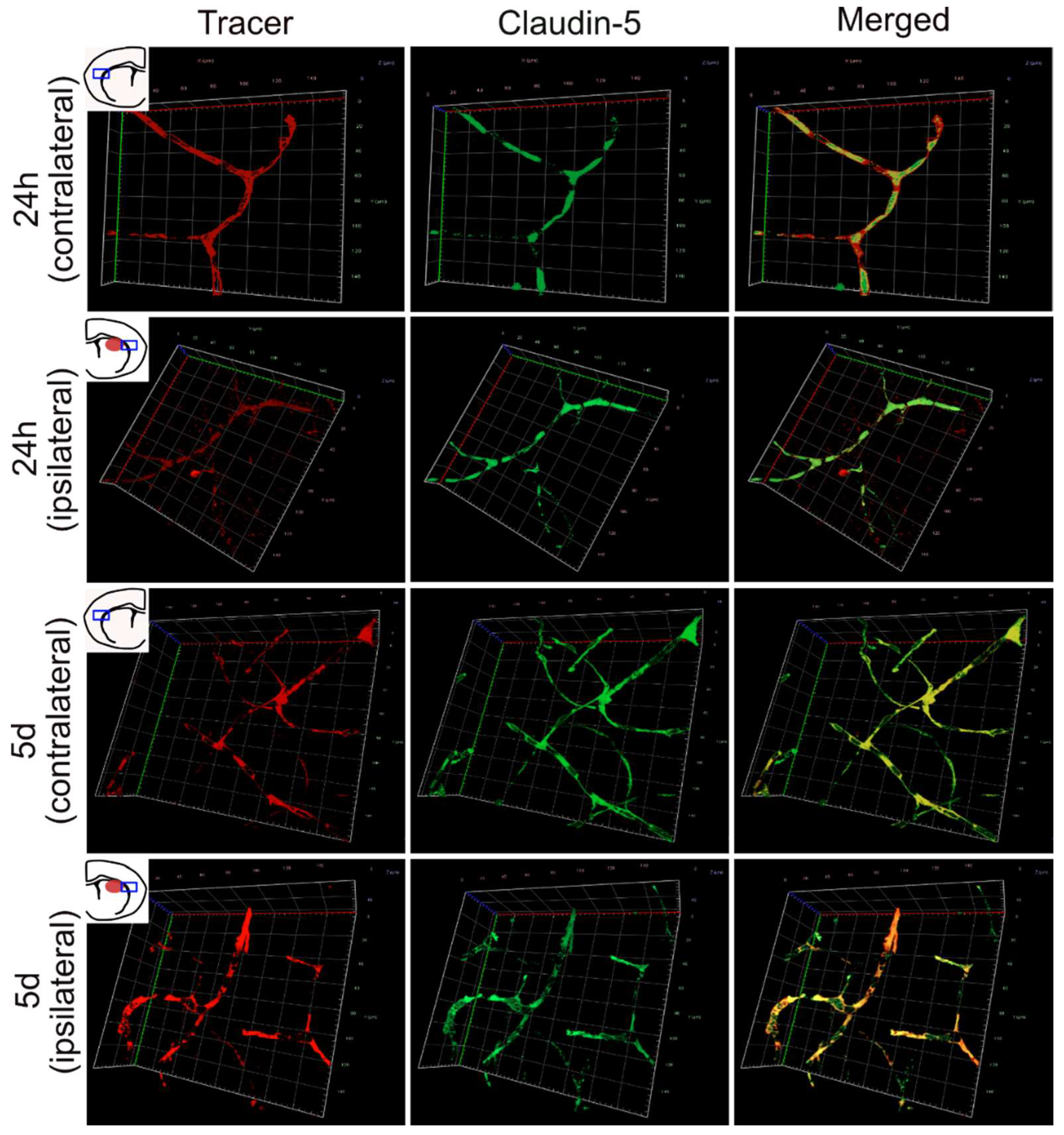
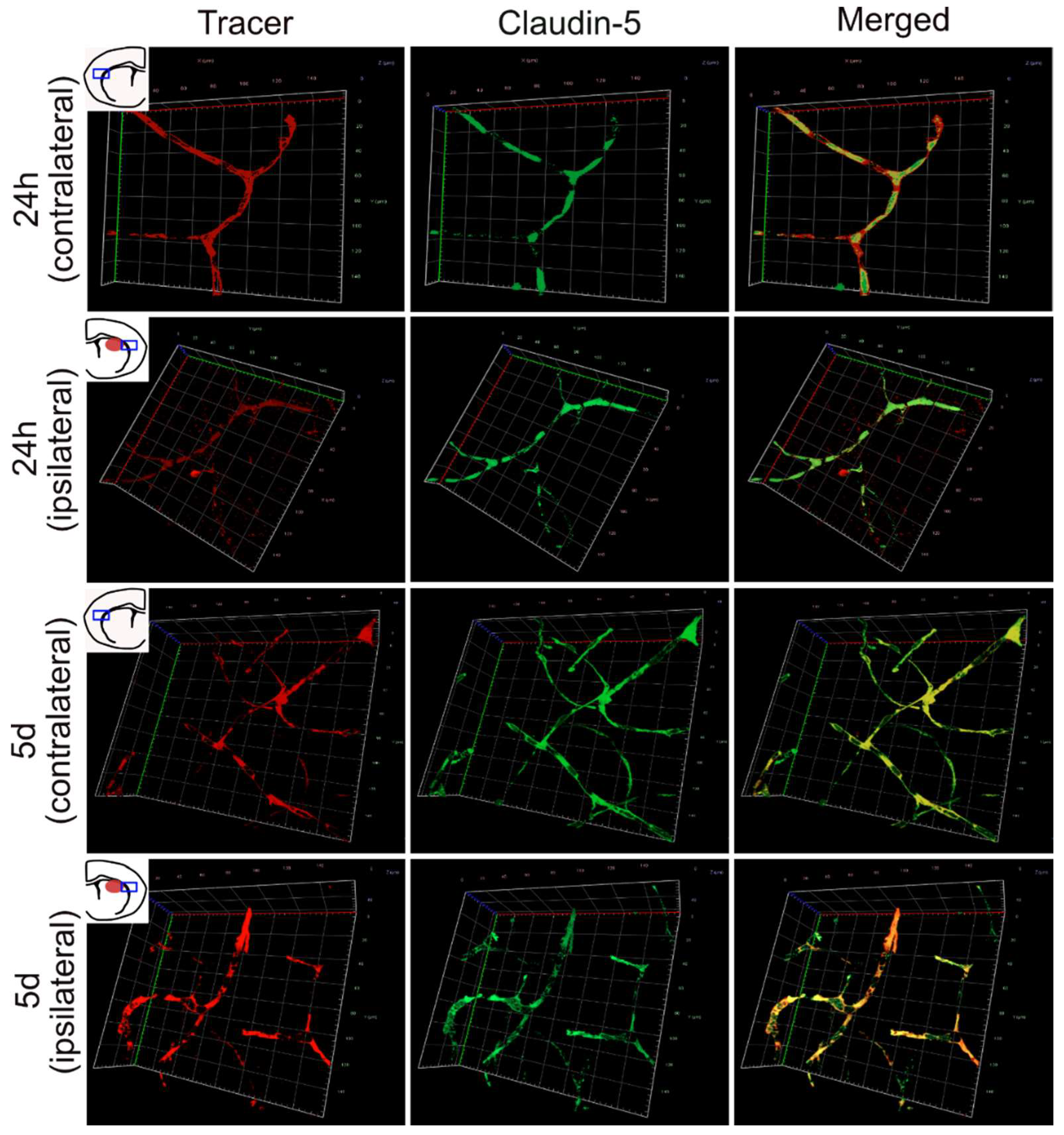
3.3. 3D Imaging of Leaking and Uninjured Brain Blood Vessels
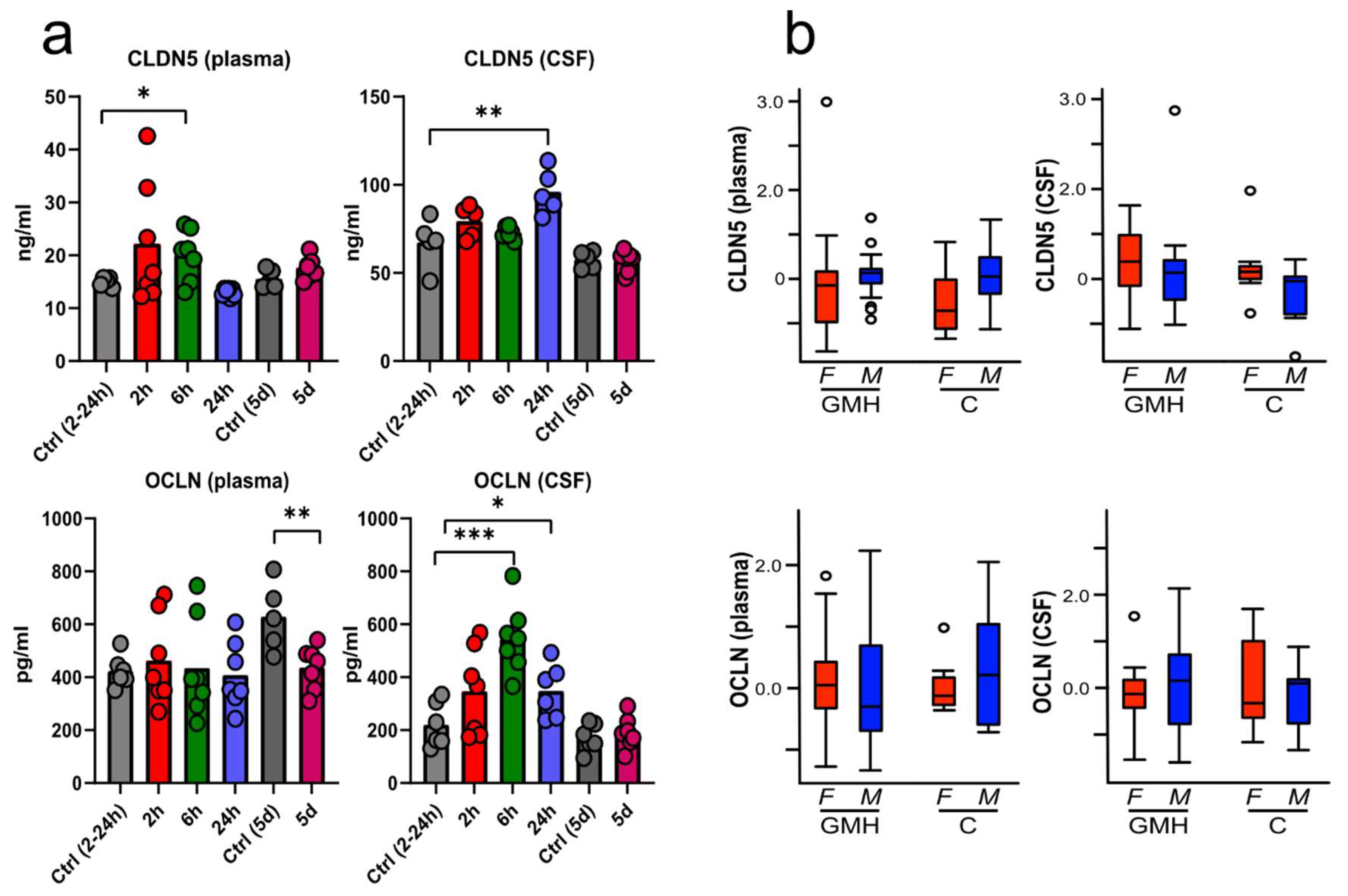
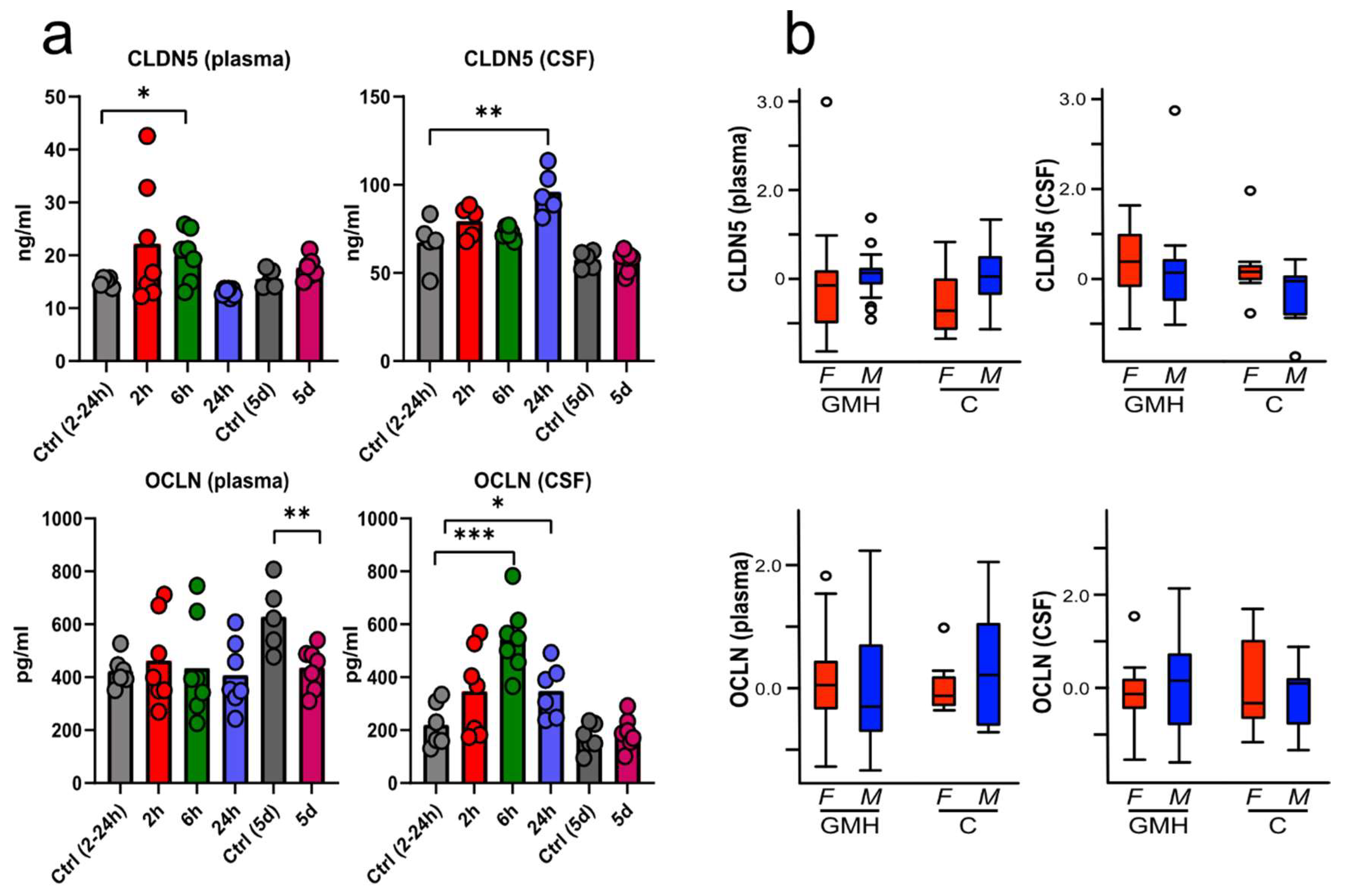
3.4. BBB Proteins Are Increased in Plasma and CSF after GMH
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Özek, E.; Kersin, S.G. Intraventricular hemorrhage in preterm babies. Turk Pediatri Ars. 2020, 55, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Back, S.A.; Miller, S.P. 60–Brain Injury in the Preterm Infant. In Avery’s Diseases of the Newborn, 10th ed.; Gleason, C.A., Juul, S.E., Eds.; Elsevier: Philadelphia, PA, USA, 2018; pp. 879–896.e876. [Google Scholar]
- El-Khoury, N.; Braun, A.; Hu, F.; Pandey, M.; Nedergaard, M.; Lagamma, E.F.; Ballabh, P. Astrocyte End-Feet in Germinal Matrix, Cerebral Cortex, and White Matter in Developing Infants. Pediatr. Res. 2006, 59, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Braun, A.; Xu, H.; Hu, F.; Kocherlakota, P.; Siegel, D.; Chander, P.; Ungvari, Z.; Csiszar, A.; Nedergaard, M.; Ballabh, P. Paucity of Pericytes in Germinal Matrix Vasculature of Premature Infants. J. Neurosci. 2007, 27, 12012–12024. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, M.M.; Stone, B.S.; Shepard, J.A.; Czosnyka, M.; Easley, R.B.; Brady, K.M. Relationship between cerebrovascular dysautoregulation and arterial blood pressure in the premature infant. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2011, 31, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Perlman, J.M.; Goodman, S.; Kreusser, K.L.; Volpe, J.J. Reduction in intraventricular hemorrhage by elimination of fluctuating cerebral blood-flow velocity in preterm infants with respiratory distress syndrome. N. Engl. J. Med. 1985, 312, 1353–1357. [Google Scholar] [CrossRef]
- Atienza-Navarro, I.; Alves-Martinez, P.; Lubian-Lopez, S.; Garcia-Alloza, M. Germinal Matrix-Intraventricular Hemorrhage of the Preterm Newborn and Preclinical Models: Inflammatory Considerations. Int. J. Mol. Sci. 2020, 21, 8343. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, R.S.; Banks, J.T.; Soleau, S.; Smyth, M.D.; Wellons, J.C.; Blount, J.P.; Grabb, P.A.; Oakes, W.J. Complications of ventriculosubgaleal shunts in infants and children. Child’s Nerv. Syst. 2005, 21, 48–51. [Google Scholar] [CrossRef]
- Ek, C.J.; Habgood, M.D.; Dziegielewska, K.M.; Saunders, N.R. Structural characteristics and barrier properties of the choroid plexuses in developing brain of the opossum (Monodelphis Domestica). J. Comp. Neurol. 2003, 460, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Ek, C.J.; Dziegielewska, K.M.; Stolp, H.; Saunders, N.R. Functional effectiveness of the blood-brain barrier to small water-soluble molecules in developing and adult opossum (Monodelphis domestica). J. Comp. Neurol. 2006, 496, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballabh, P. Intraventricular hemorrhage in premature infants: Mechanism of disease. Pediatr. Res. 2010, 67, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballabh, P.; Hu, F.; Kumarasiri, M.; Braun, A.; Nedergaard, M. Development of tight junction molecules in blood vessels of germinal matrix, cerebral cortex, and white matter. Pediatr. Res. 2005, 58, 791–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinnai, M.; Koning, G.; Singh-Mallah, G.; Jonsdotter, A.; Leverin, A.-L.; Svedin, P.; Nair, S.; Takeda, S.; Wang, X.; Mallard, C.; et al. A Model of Germinal Matrix Hemorrhage in Preterm Rat Pups. Front Cell Neurosci 2020, 14, 535320. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallard, C.; Vexler, Z.S. Modeling Ischemia in the Immature Brain: How Translational Are Animal Models? Stroke 2015, 46, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Habgood, M.D.; Bye, N.; Dziegielewska, K.M.; Ek, C.J.; Lane, M.A.; Potter, A.; Morganti-Kossmann, C.; Saunders, N.R. Changes in blood-brain barrier permeability to large and small molecules following traumatic brain injury in mice. Eur. J. Neurosci. 2007, 25, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.A.; Mallard, C.; Ek, C.J. Circulating tight-junction proteins are potential biomarkers for blood–brain barrier function in a model of neonatal hypoxic/ischemic brain injury. Fluids Barriers CNS 2021, 18, 7. [Google Scholar] [CrossRef]
- Mottahedin, A.; Smith, P.L.; Hagberg, H.; Ek, C.J.; Mallard, C. TLR2-mediated leukocyte trafficking to the developing brain. J. Leukoc. Biol. 2017, 101, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Lekic, T.; Flores, J.; Klebe, D.; Doycheva, D.; Rolland, W.B.; Tang, J.; Zhang, J.H. Intranasal IGF-1 Reduced Rat Pup Germinal Matrix Hemorrhage. Acta Neurochir. Suppl. 2016, 121, 209–212. [Google Scholar] [CrossRef]
- Castro Dias, M.; Coisne, C.; Lazarevic, I.; Baden, P.; Hata, M.; Iwamoto, N.; Francisco, D.M.F.; Vanlandewijck, M.; He, L.; Baier, F.A.; et al. Claudin-3-deficient C57BL/6J mice display intact brain barriers. Sci. Rep. 2019, 9, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, W.B.; Krafft, P.R.; Lekic, T.; Klebe, D.; LeGrand, J.; Weldon, A.J.; Xu, L.; Zhang, J.H. Fingolimod confers neuroprotection through activation of Rac1 after experimental germinal matrix hemorrhage in rat pups. J. Neurochem. 2017, 140, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Liu, F.; Chen, Z.; Hua, Y.; Keep, R.F.; Xi, G. Hydrocephalus after Intraventricular Hemorrhage: The Role of Thrombin. J. Cereb. Blood Flow Metab. 2013, 34, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gram, M.; Sveinsdottir, S.; Ruscher, K.; Hansson, S.R.; Cinthio, M.; Åkerström, B.; Ley, D. Hemoglobin induces inflammation after preterm intraventricular hemorrhage by methemoglobin formation. J. Neuroinflamm. 2013, 10, 867. [Google Scholar] [CrossRef] [Green Version]
- Agyemang, A.A.; Sveinsdóttir, K.; Vallius, S.; Sveinsdóttir, S.; Bruschettini, M.; Romantsik, O.; Hellström, A.; Smith, L.E.H.; Ohlsson, L.; Holmqvist, B.; et al. Cerebellar Exposure to Cell-Free Hemoglobin Following Preterm Intraventricular Hemorrhage: Causal in Cerebellar Damage? Transl. Stroke Res. 2017, 8, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Ley, D.; Romantsik, O.; Vallius, S.; Sveinsdóttir, K.; Sveinsdóttir, S.; Agyemang, A.A.; Baumgarten, M.; Mörgelin, M.; Lutay, N.; Bruschettini, M.; et al. High Presence of Extracellular Hemoglobin in the Periventricular White Matter Following Preterm Intraventricular Hemorrhage. Front. Physiol. 2016, 7, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Chen, Q.; Guo, J.; Yang, L.; Tao, Y.; Li, L.; Miao, H.; Feng, H.; Chen, Z.; Zhu, G. Minocycline Attenuates Neonatal Germinal-Matrix-Hemorrhage-Induced Neuroinflamm. and Brain Edema by Activating Cannabinoid Receptor 2. Mol. Neurobiol. 2016, 53, 1935–1948. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, J.; Li, P.; Ding, Y.; Tang, J.; Chen, G.; Zhang, J.H. NT-4 attenuates neuroinflammation via TrkB/PI3K/FoxO1 pathway after germinal matrix hemorrhage in neonatal rats. J. Neuroinflamm. 2020, 17, 158. [Google Scholar] [CrossRef]
- Ek, C.J.; D’Angelo, B.; Baburamani, A.A.; Lehner, C.; Leverin, A.L.; Smith, P.L.; Nilsson, H.; Svedin, P.; Hagberg, H.; Mallard, C. Brain barrier properties and cerebral blood flow in neonatal mice exposed to cerebral hypoxia-ischemia. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2015, 35, 818–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.-Y.; Betz, A.L.; Chenevert, T.L.; Brunberg, J.A.; Hoff, J.T. Experimental intracerebral hemorrhage: Relationship between brain edema, blood flow, and blood-brain barrier permeability in rats. J. Neurosurg. 1994, 81, 93. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Yu, K.; Weatherwax, T.; Zheng, H.; Liu, W.; Liu, K.J. Blood Occludin Level as a Potential Biomarker for Early Blood Brain Barrier Damage Following Ischemic Stroke. Sci. Rep. 2017, 7, 40331. [Google Scholar] [CrossRef]
- Kazmierski, R.; Michalak, S.; Wencel-Warot, A.; Nowinski, W.L. Serum tight-junction proteins predict hemorrhagic transformation in ischemic stroke patients. Neurology 2012, 79, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; He, P.; Li, Y.; Fan, Z.; Si, M.; Xie, Q.; Chang, X.; Huang, D. The Role of Circulating Tight Junction Proteins in Evaluating Blood Brain Barrier Disruption following Intracranial Hemorrhage. Dis. Markers 2015, 2015, 860120. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.A.; Fitch, R.H. Sex differences in mechanisms and outcome of neonatal hypoxia-ischemia in rodent models: Implications for sex-specific neuroprotection in clinical neonatal practice. Neurol. Res. Int. 2012, 2012, 867531. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.A.; Threlkeld, S.W.; Fitch, R.H. Early testosterone modulated sex differences in behavioral outcome following neonatal hypoxia ischemia in rats. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2011, 29, 381–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, M.A.; Aly, H. Male Gender Is Associated With Intraventricular Hemorrhage. Pediatrics 2010, 125, e333–e339. [Google Scholar] [CrossRef]
- Kent, A.L.; Wright, I.M.R.; Abdel-Latif, M.E. Mortality and Adverse Neurologic Outcomes Are Greater in Preterm Male Infants. Pediatrics 2012, 129, 124–131. [Google Scholar] [CrossRef]
- Tioseco, J.A.; Aly, H.; Essers, J.; Patel, K.; El-Mohandes, A.A.E. Male sex and intraventricular hemorrhage. Pediatr. Crit. Care Med. 2006, 7, 40–44. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersson, E.A.; Rocha-Ferreira, E.; Hagberg, H.; Mallard, C.; Ek, C.J. Function and Biomarkers of the Blood-Brain Barrier in a Neonatal Germinal Matrix Haemorrhage Model. Cells 2021, 10, 1677. https://doi.org/10.3390/cells10071677
Andersson EA, Rocha-Ferreira E, Hagberg H, Mallard C, Ek CJ. Function and Biomarkers of the Blood-Brain Barrier in a Neonatal Germinal Matrix Haemorrhage Model. Cells. 2021; 10(7):1677. https://doi.org/10.3390/cells10071677
Chicago/Turabian StyleAndersson, Erik Axel, Eridan Rocha-Ferreira, Henrik Hagberg, Carina Mallard, and Carl Joakim Ek. 2021. "Function and Biomarkers of the Blood-Brain Barrier in a Neonatal Germinal Matrix Haemorrhage Model" Cells 10, no. 7: 1677. https://doi.org/10.3390/cells10071677
APA StyleAndersson, E. A., Rocha-Ferreira, E., Hagberg, H., Mallard, C., & Ek, C. J. (2021). Function and Biomarkers of the Blood-Brain Barrier in a Neonatal Germinal Matrix Haemorrhage Model. Cells, 10(7), 1677. https://doi.org/10.3390/cells10071677