
Man-Specific Lectins from Plants, Fungi, Algae and Cyanobacteria, as Potential Blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) Coronaviruses: Biomedical Perspectives

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Man-Specific Lectins from Higher Plants, Fungi, Algae, and Cyanobacteria
2.1. Man-Specific Lectins from Higher Plants
- -
- The β-sandwich structure (jelly-roll structural scaffold), present in legume lectins. They are classically divided into two groups of single- and two-chain lectins, according to the cleavage (two-chain) or the absence of cleavage (single-chain) of their protomers. Single-chain, tetravalent lectins consist of the non-covalent association of four identical protomers (α4), each containing a CBS that specifically recognizes Man and its derivatives. In two-chain lectins, a proteolytic cleavage of the protomeric chain results in the separation of a light (α-chain) and a heavy (β-chain), which remain associated with non-covalent bonds. Two-chain, bivalent lectins (α2β2)) result from the non-covalent association of two identical two-chain protomers, each possessing a CBS specific for Man and its derivatives.
- -
- The β-prism I structure (β-barrel structural scaffold), present in the jacalin-related lectin group. Their protomers are organized in three bundles of four antiparallel β-strands arranged into a β-prism structure along a longitudinal axis. Depending on the number of identical protomers associated by non-covalent bonds, Man-specific jacalin-related lectins consist of bivalent (two CBSs) lectins, e.g., Calsepa from hedge bindweed Calystegia sepium [61] (Figure 2C); tetravalent (four CBSs) lectins, e.g., Artocarpin from black mulberry Artocarpus integer [65] (Figure 2D); hexavalent (six CBSs) lectins, e.g., PPL from the African locust bean Parkia platycephala [70] (Figure 2E); or octavalent (eight CBSs) lectins, e.g., Heltuba from the Jerusalem artichoke Helianthus tuberosus [71] (Figure 2F).
- -
- The β-prism II structure (β-trefoil structural scaffold), present in the GNA-related lectin group. The protomer consists of three bundles of four antiparallel β-strands arranged into a flattened β-trefoil structure around a central pseudoaxis. A CBS occurs in each of the bundles of β-strands. Except for gastrodianin, which is composed of a single protomer (Figure 2G), lectins belonging to the GNA-related group result from the non-covalent association of two protomers, e.g., the hexavalent NPA from daffodil (Narcissus pseudonarcissus) bulbs [80] (Figure 2H) or four protomers, e.g., the dodecavalent GNA from snowdrop (Galanthus nivalis) bulbs [94] (Figure 2I).
- -
- Two covalently linked β-trefoil-domains form the B-chain of the lectin from Iris hollandica, a type II ribosome-inactivating protein (RIP-II) (Figure 2J). Each domain contains a CBS which can accommodate Man and Gal/GalNAc. This Man-binding property confers an unusual Man-specificity to the Iris RIP-II, which readily differs from the Gal/GalNAc-specificity of other RIP-IIs [97].
- -
- Nictaba, the jasmonic acid-induced tobacco (Nicotiana tabacum) lectin displays specificity for chitin and GlcNAc oligomers but it can also bind to high mannose glycans with a lower affinity, as shown from glycan array experiments. The molecular structure of Nictaba is still unknown but modeling experiments suggest a β-sandwich structure for this tobacco lectin [98] (Figure 2K).
2.2. Man-Specific Lectins from Lower Plants and Fungi
- -
- The β-sandwich structure (jelly-roll structural scaffold) typical for legume lectins, also occurs in the Man-specific lectin PeCL from Penicillium chrysogenum, and in floculins Flo1p from the yeast Saccharomyces pasteurianum and Flo5p from Saccharomyces cerevisiae.
- -
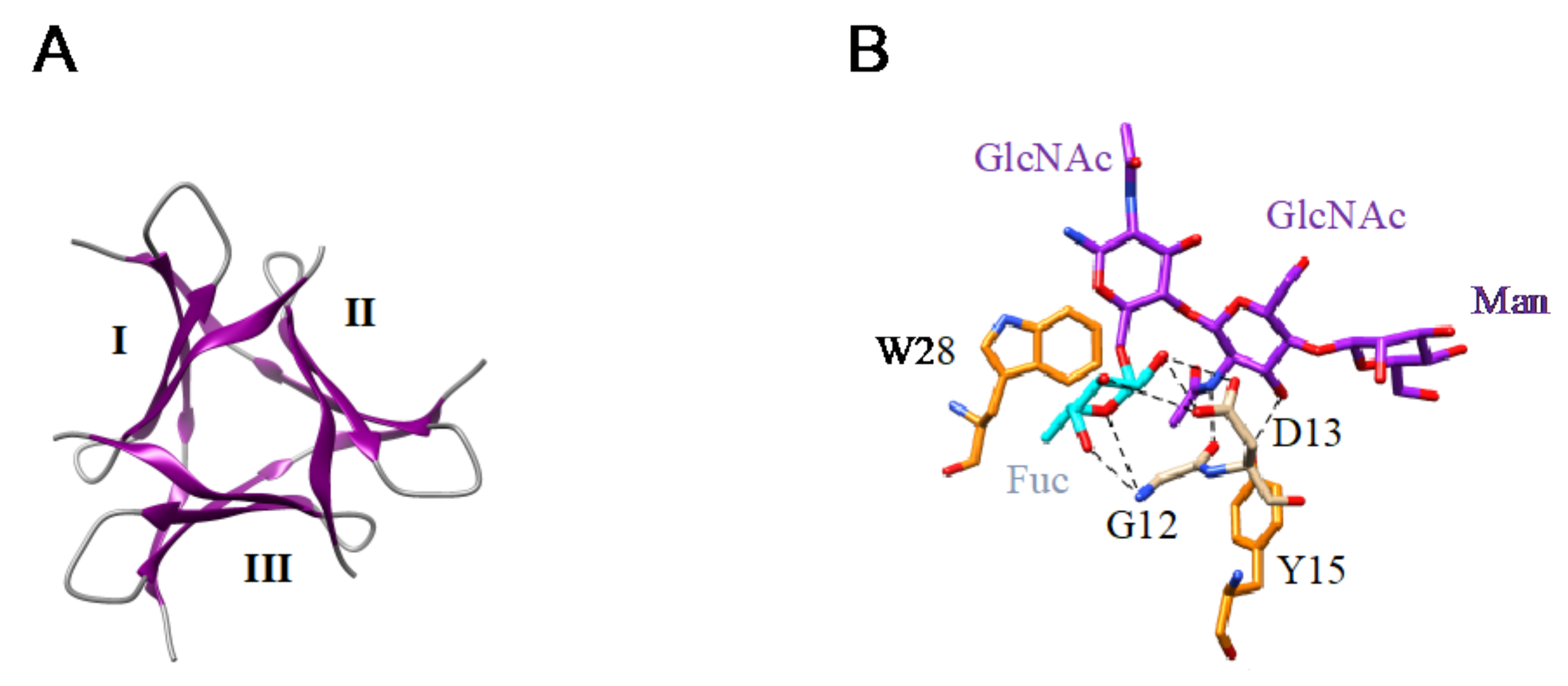
- The β-prism II structure (β-trefoil structural scaffold), that builds the GNA-related lectin protomer, also occurs in Marpola, the Man-specific lectin isolated from the liverwort Marchantia polymorpha, and in mushrooms (Abmb from Agaricus bisporus, MOA from Marasmius oreades). Special mention should be made to the apparent β-trefoil structure that was found in the mini-lectin PhosL from the mushroom Pholiota squarrosa. In fact, PhosL results from the trimeric association of three antiparallel β-sheets, each possessing a CBS, that mimics a β-trefoil structure with three identical CBSs. This newly identified lectin fold is involved in the specific recognition of the core α1,6-fucosylation of fucosylated N-glycans [100] (Figure 12A,B).
- -
- A rather unusual 6-bladded β-propeller organization was identified in tectonin 2, the Man-specific lectin from the basidiomycete Laccaria bicolor [109]. As commonly observed in other lectins exhibiting a β-propeller structure, tectonin 2 contains 6 identical Man-specific CBSs located at the top of each of the six blades forming the β-propeller.
- -
- Another quite unusual structural scaffold, which consists of an antiparallel five-stranded β-sheet associated to two α-helices (α + β fold), occurs in the monomeric ginkbilobin 2, a Man-specific lectin from Ginkgo biloba [103]. Hydrophilic residues occurring on the loops connecting the β-strands associated with charged residues of two β-strands, to form a Man-binding pocket located at the top of the monomer [103].
2.3. Man-Specific Lectins from Algae and Bacteria
- -
- The β-prism I or β-barrel scaffold occurs predominantly in red algae (Rhodophyta) but is less common in other classes of algae (Figure 18A–M). Algae of the genera Agardhiella, Eucheuma, Kappaphycus, Meristiella, Meristotheca, and Solieria exhibit this type of molecular organization, in which the CBS is located at the top of the barrel structure.
- -
- A less common β−prism II organization occurs in red algae (Grateloupia chiangii) and green algae (Boodlea coacta), in which CBSs are located in the grooves delineated by the bundles of β-strands. The MFP2-like structural scaffold, related to the β-prism II organization, occurs in the green alga Bryopsis plumosa.
- -
- A rather uncommon β-sandwich organization was found in the red alga Hydropuntia (Gracilaria) fisheri, and in the green alga Ostreococcus tauri.
- -
- It is interesting to note that the structural scaffolds of many other Man-specific lectins, identified in red algae (Carpopeltis flabellata, Gracilaria bursa-pastoris) and green algae (Enteromopha prolifera, Halimeda renschii), still remain unknown and are not apparently related to any of the known structural scaffolds reported in algae.
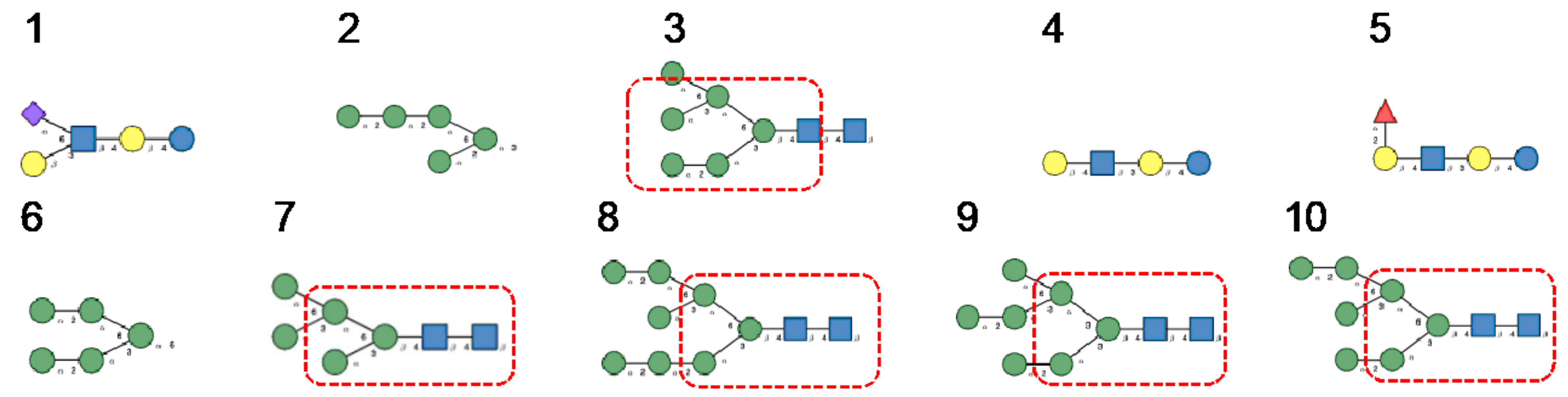
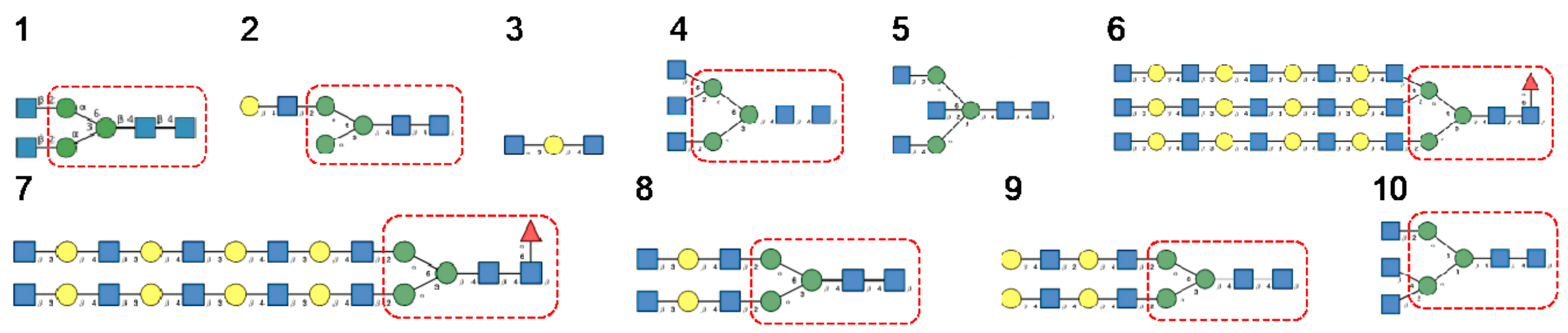
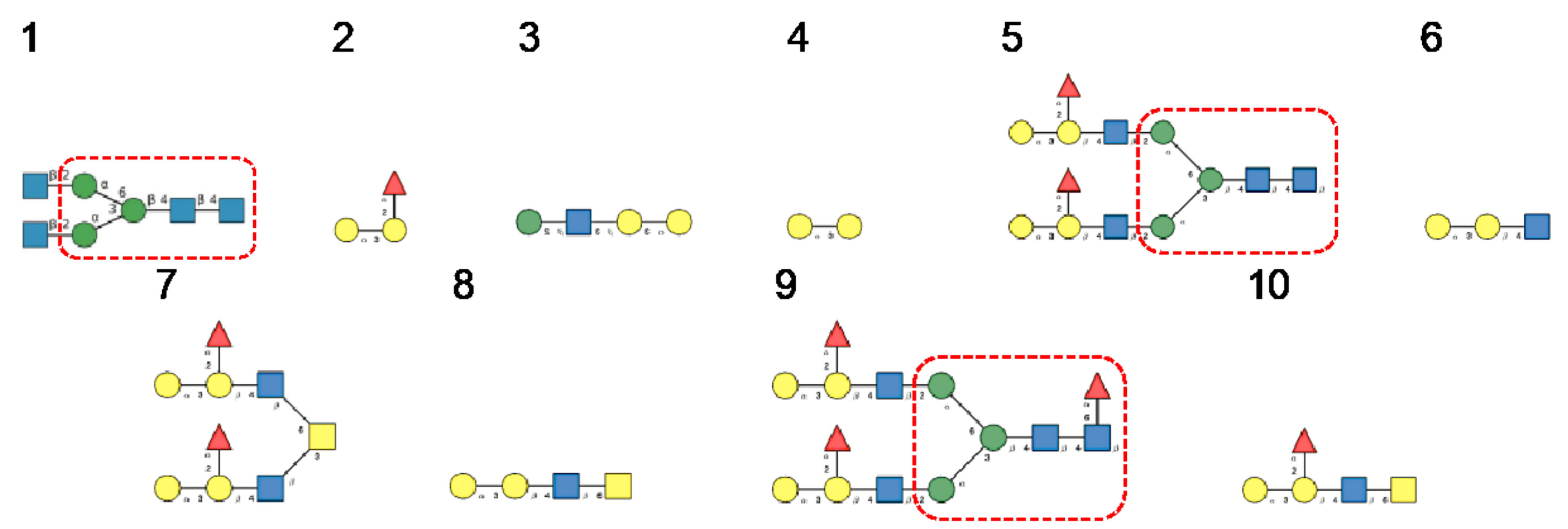
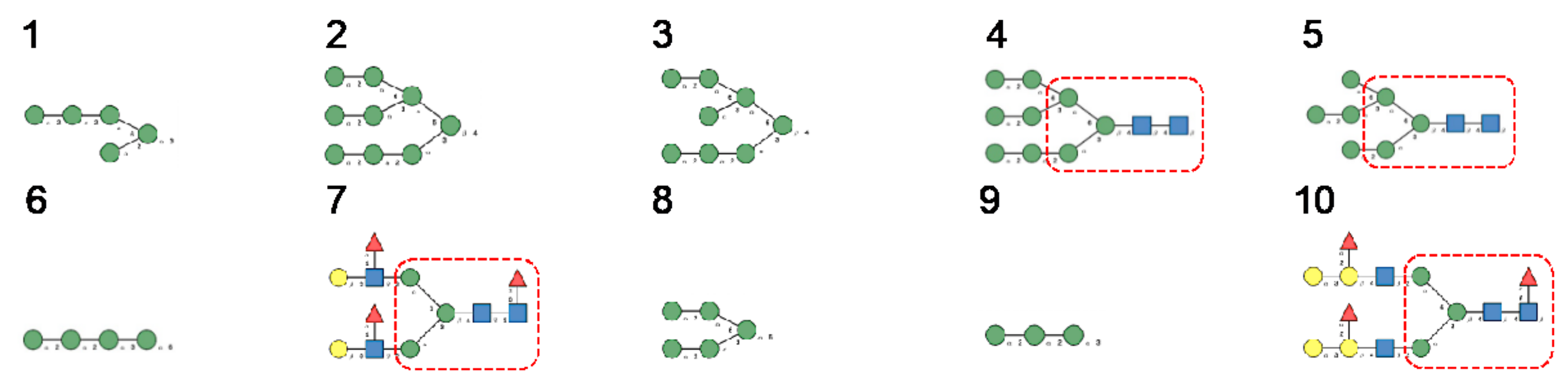
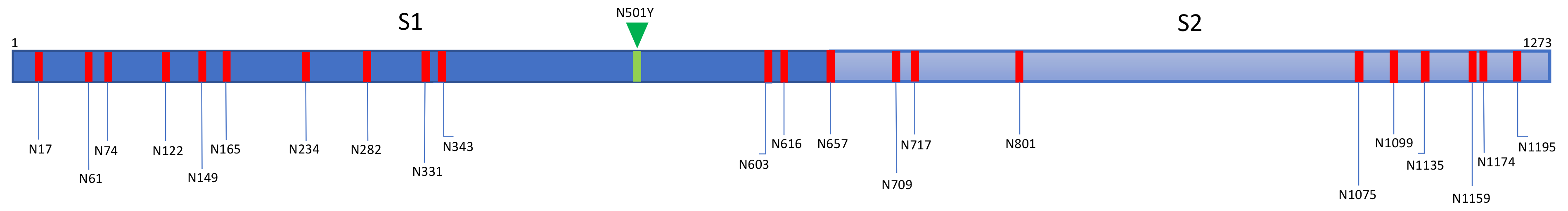
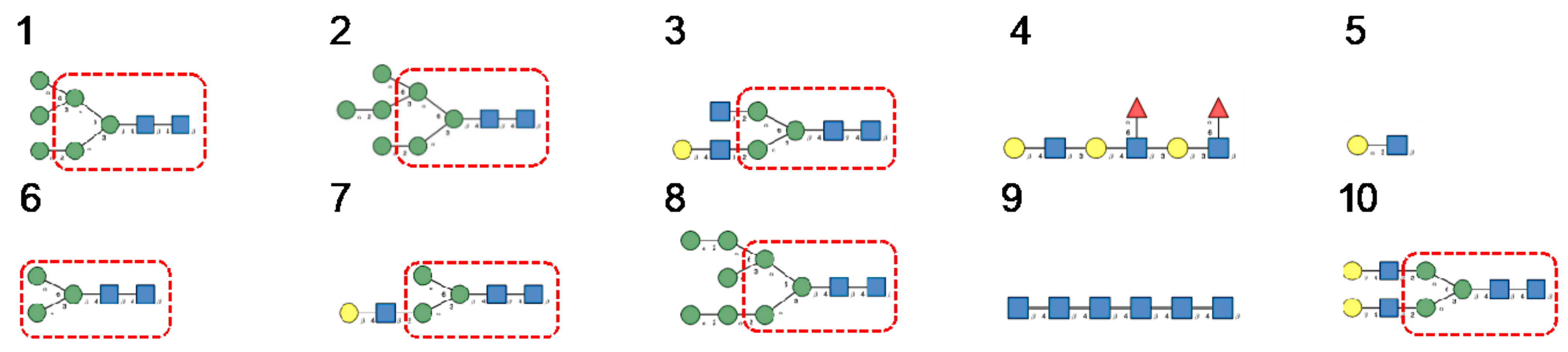
3. Diversity of Glycans Decorating the Spike-Forming S Glycoproteins of Coronaviruses
- -
- Hybrid type glycans, often bisected, have been detected at different N-glycosylation sites but with the exception of the last C-terminal N-glycosylation site occurring in S-glycoprotein of MERS-CoV which is exclusively occupied by hybrid-type glycans. All other N-glycosylation sites are occupied by a mixture of high-mannose, complex, and hybrid glycans (Table 1).
- -
- Scarce potential O-glycosylation sites T/S, actually occupied by O-glycans, have been identified essentially in the S-glycoprotein of SARS-CoV-2 (T323, S325, and T678) [12].
4. Structural Organization and Glycosylation Pattern of S Glycoproteins Forming the Spikes of Coronaviruses
- -
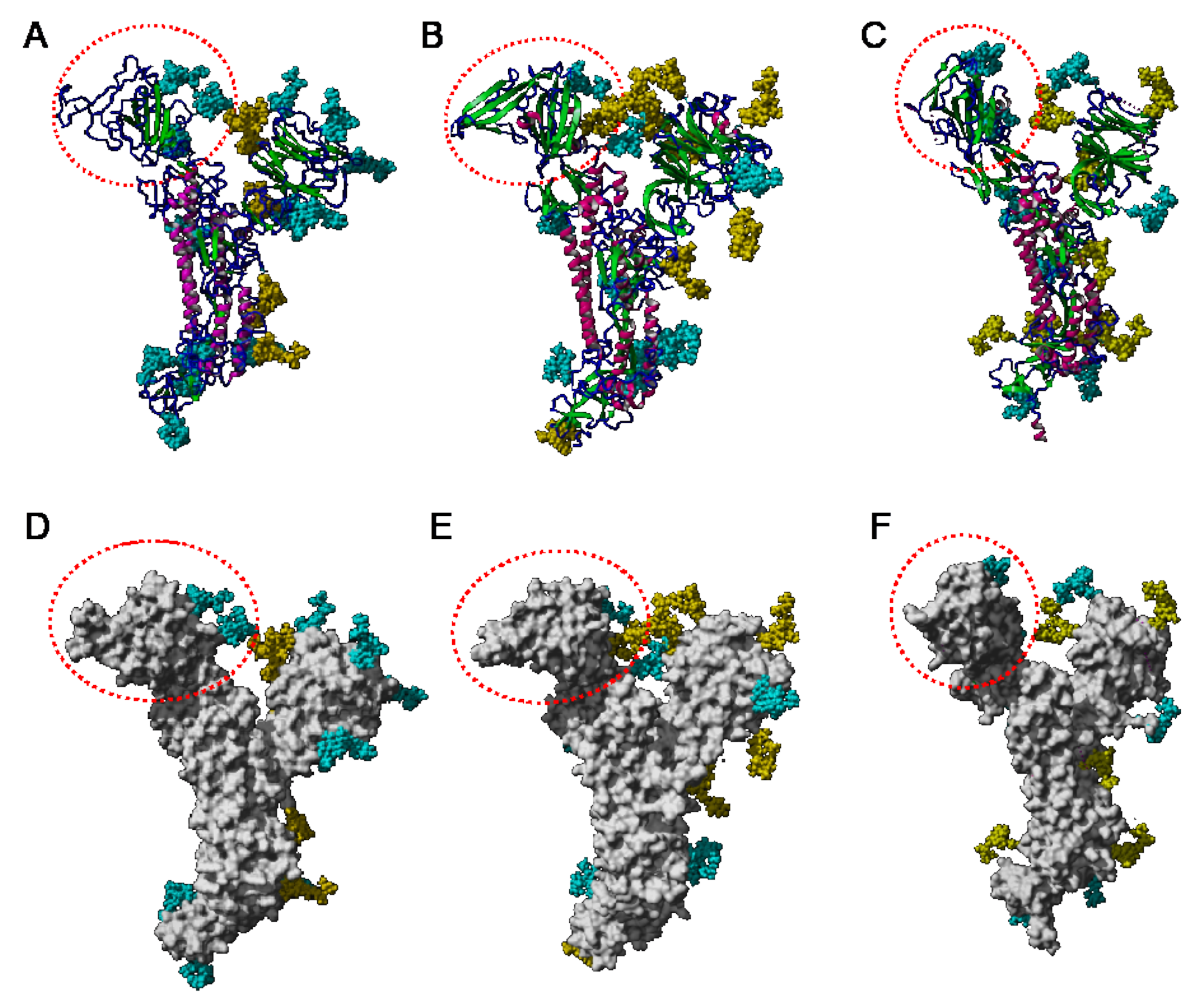
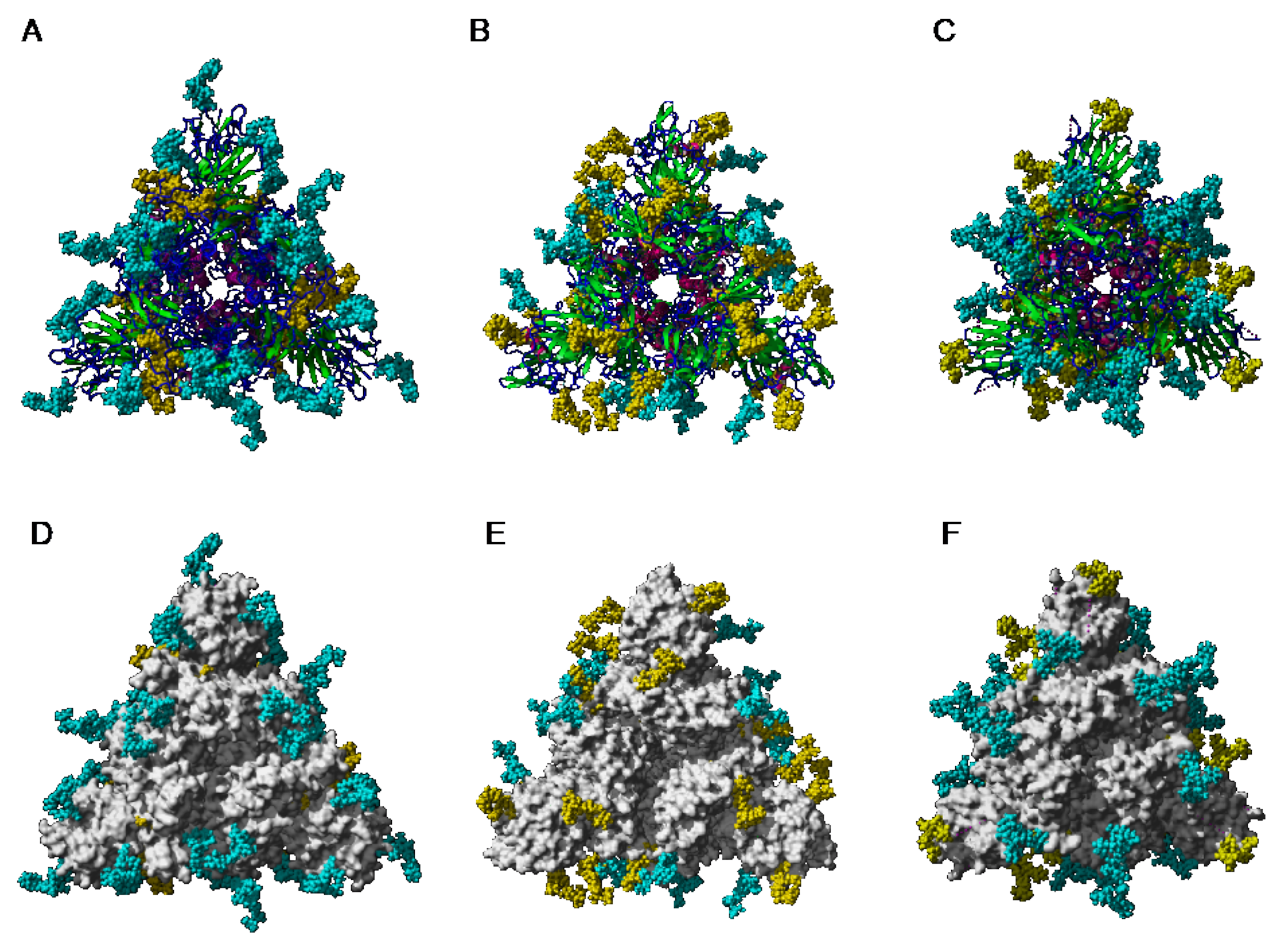
- high-mannose glycans are predominantly distributed in the upper part of the S-glycoprotein from MERS-CoV, whereas complex glycans mainly occur along the lower part of the S-glycoprotein.
- -
- complex glycans predominate in the upper part and lower part of S-glycoprotein from SARS-CoV, while the less abundant high-mannose glycans are equally distributed along with the glycoprotein.
- -
- in all three coronaviruses, the RBDs only contain complex N-glycans.
5. Man-Specific Lectins from Higher Plants, Fungi, Algae and Cyanobacteria, Specifically Interact with the Highly Glycosylated S Glycoprotein from Coronaviruses
- -
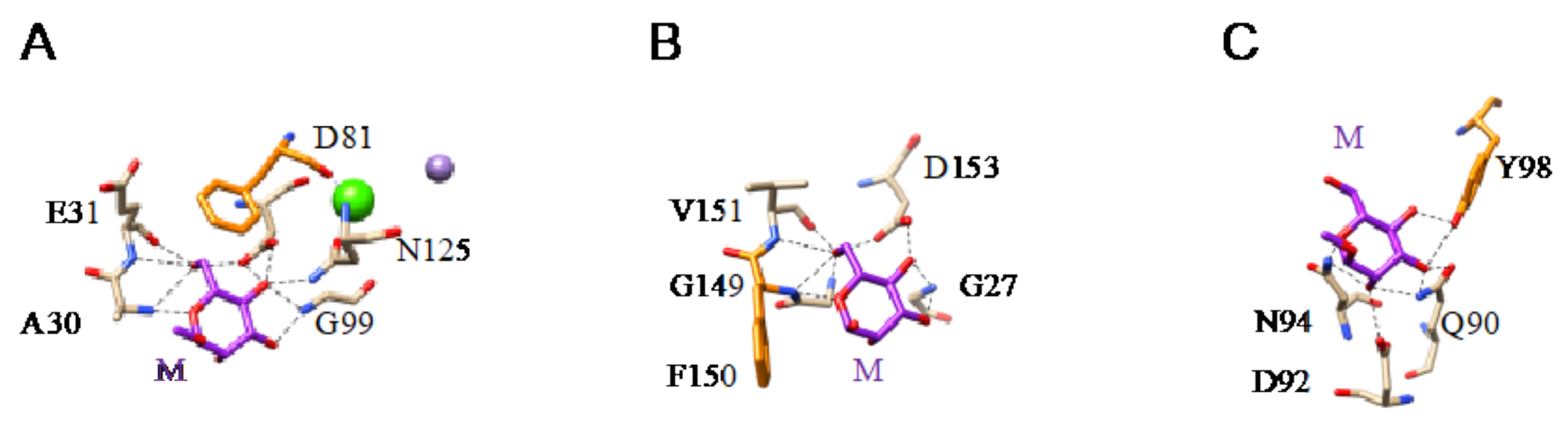
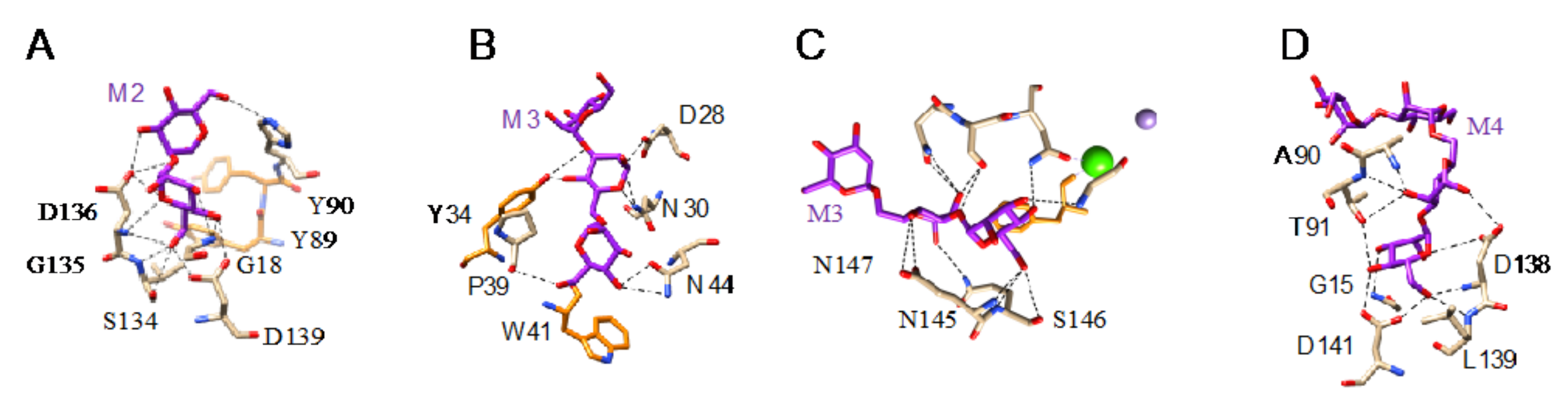
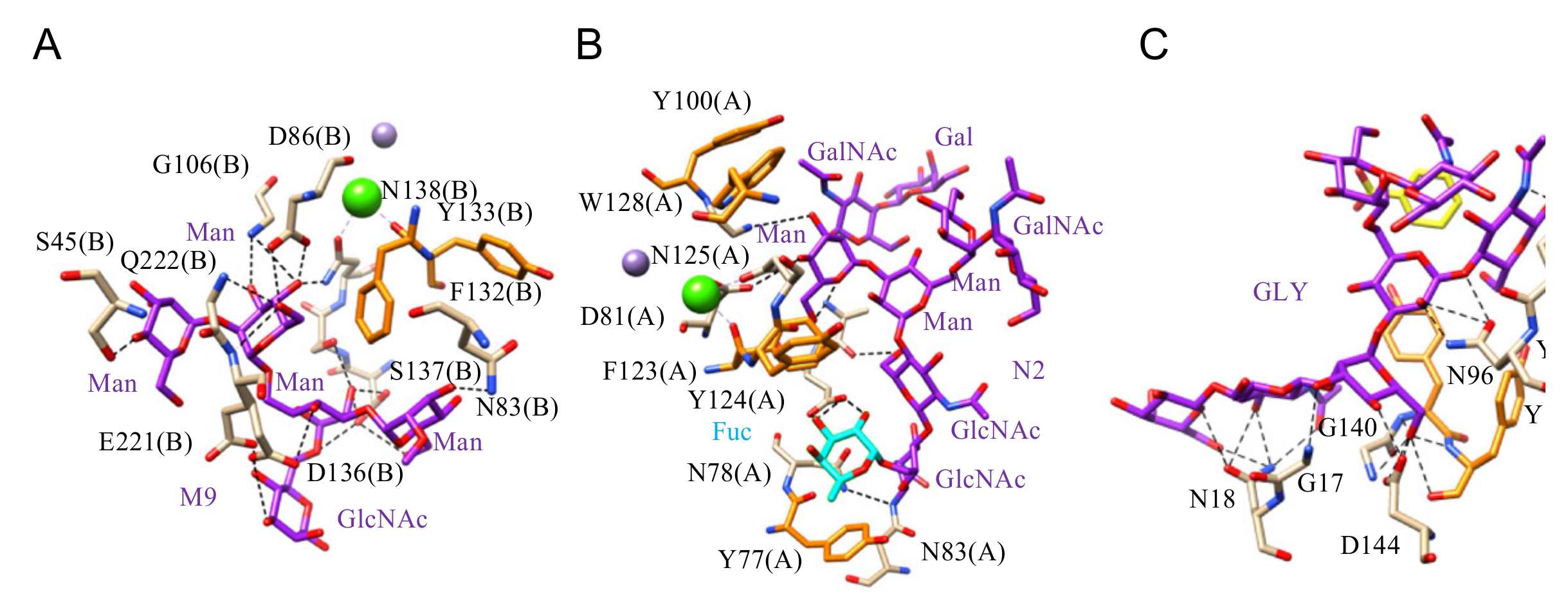
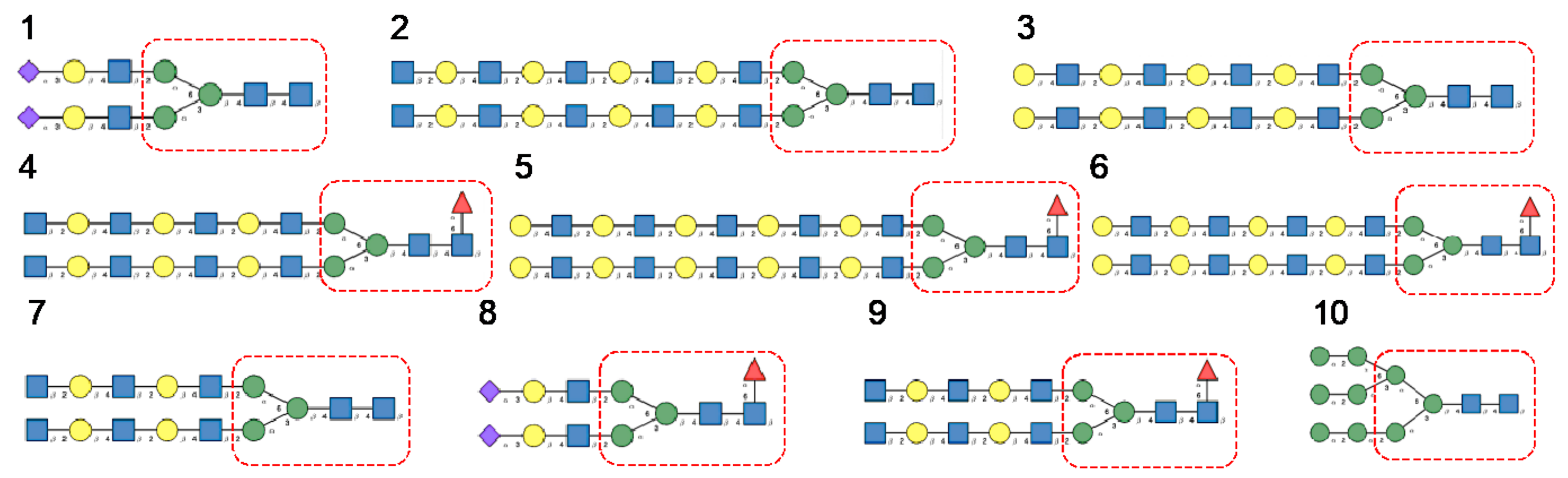
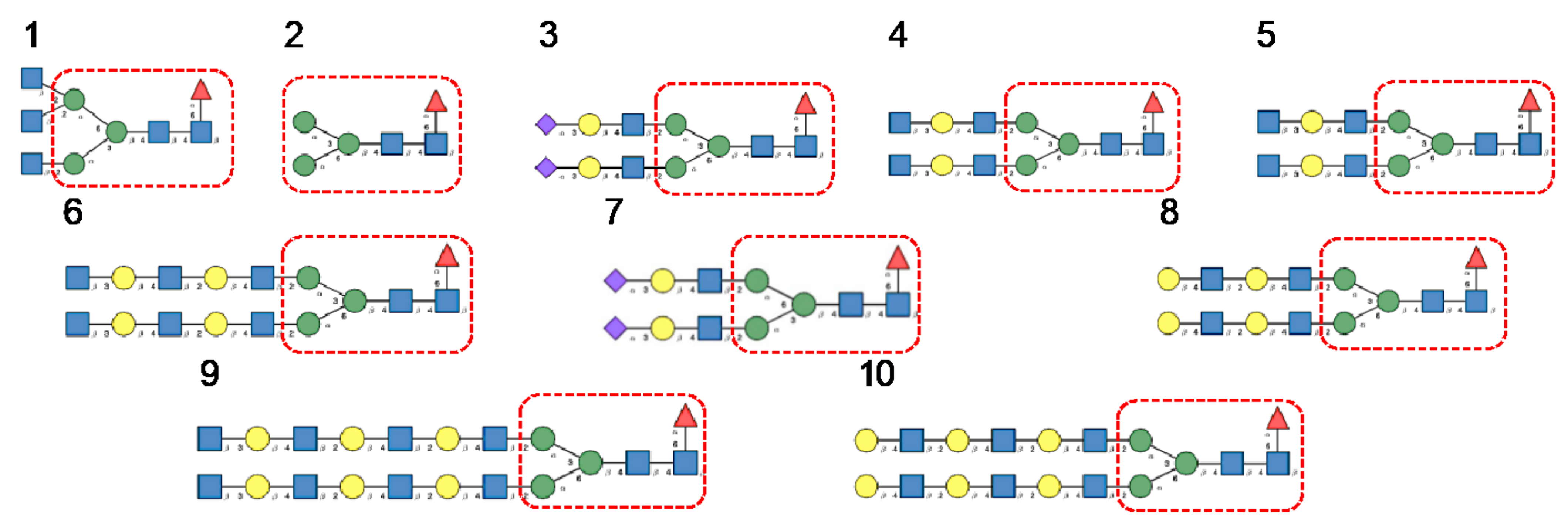
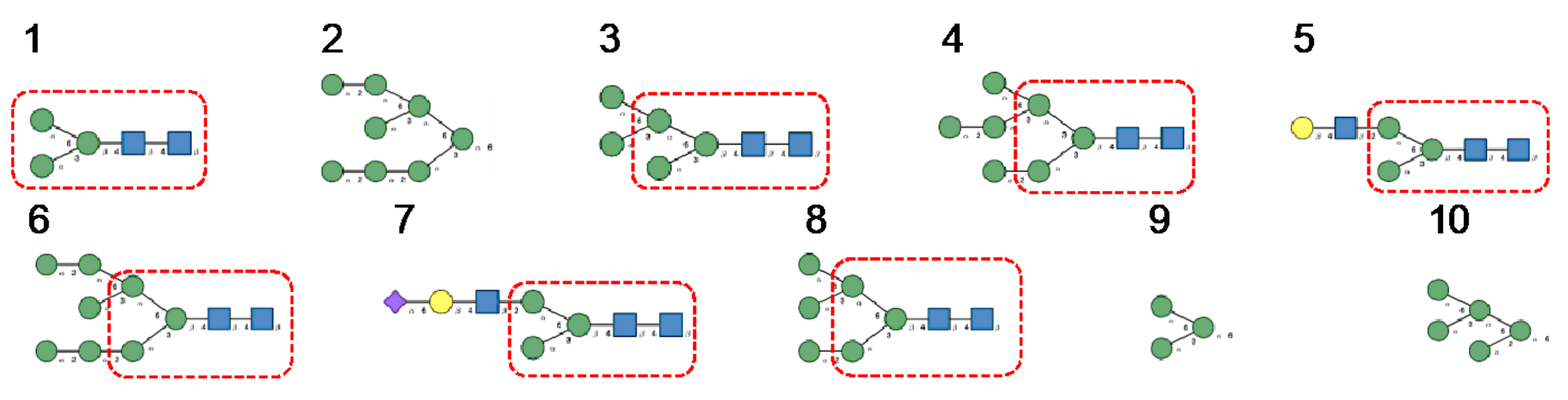
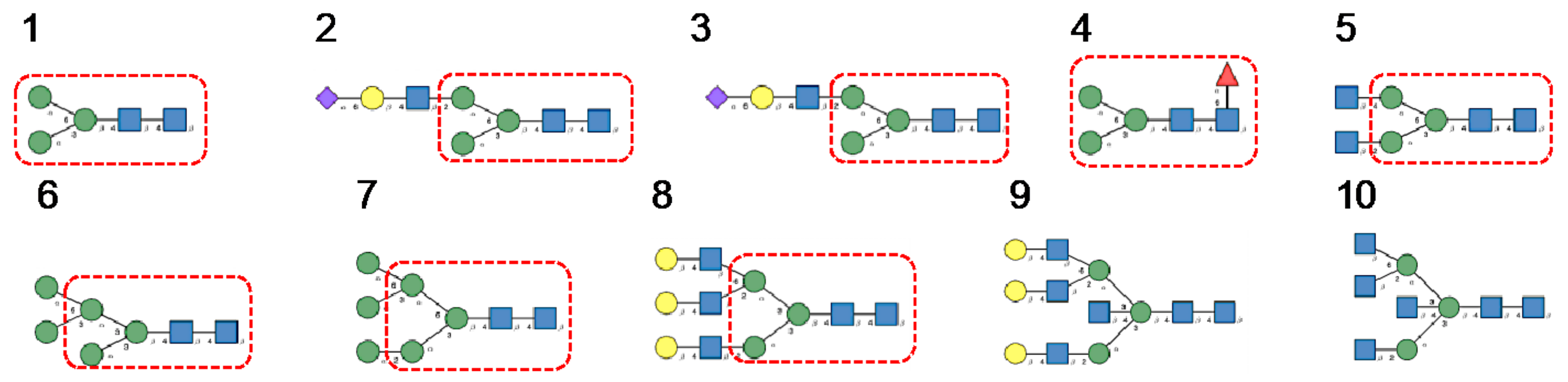
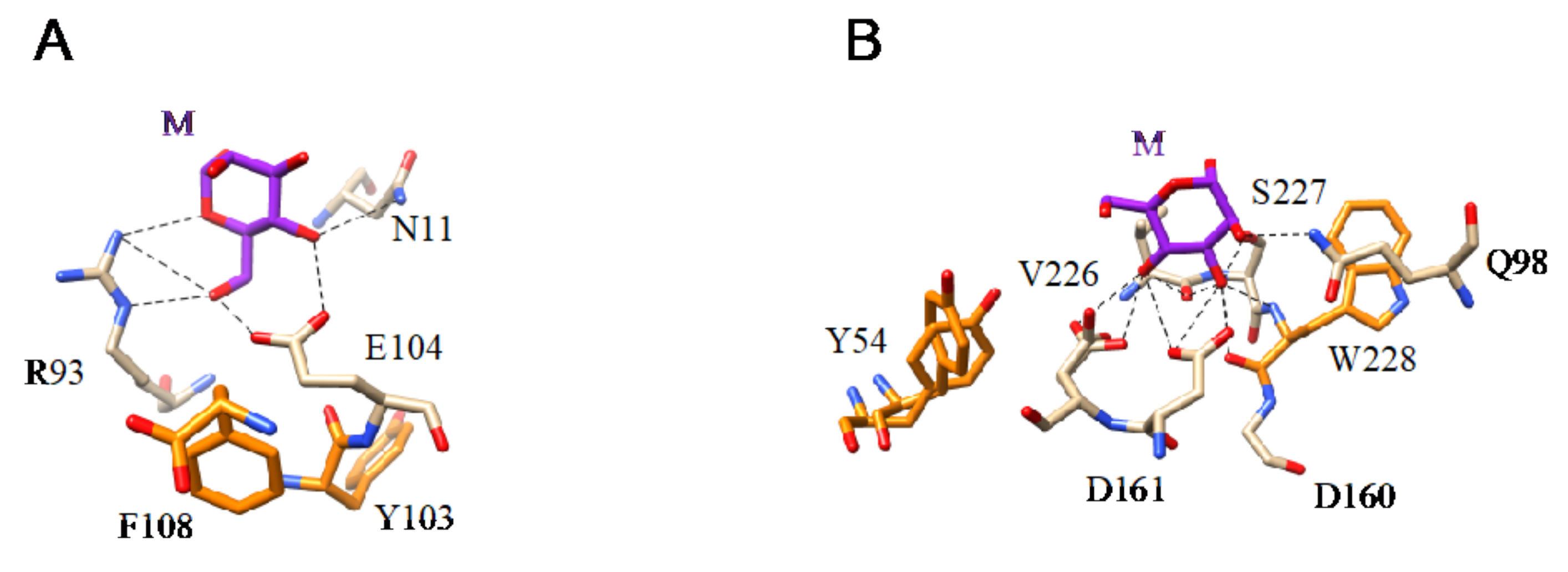
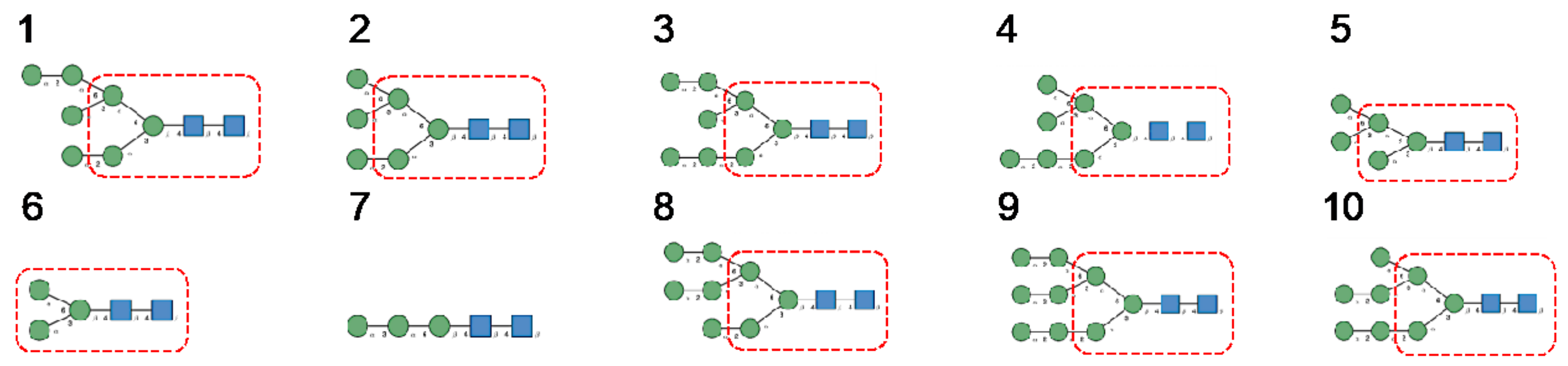
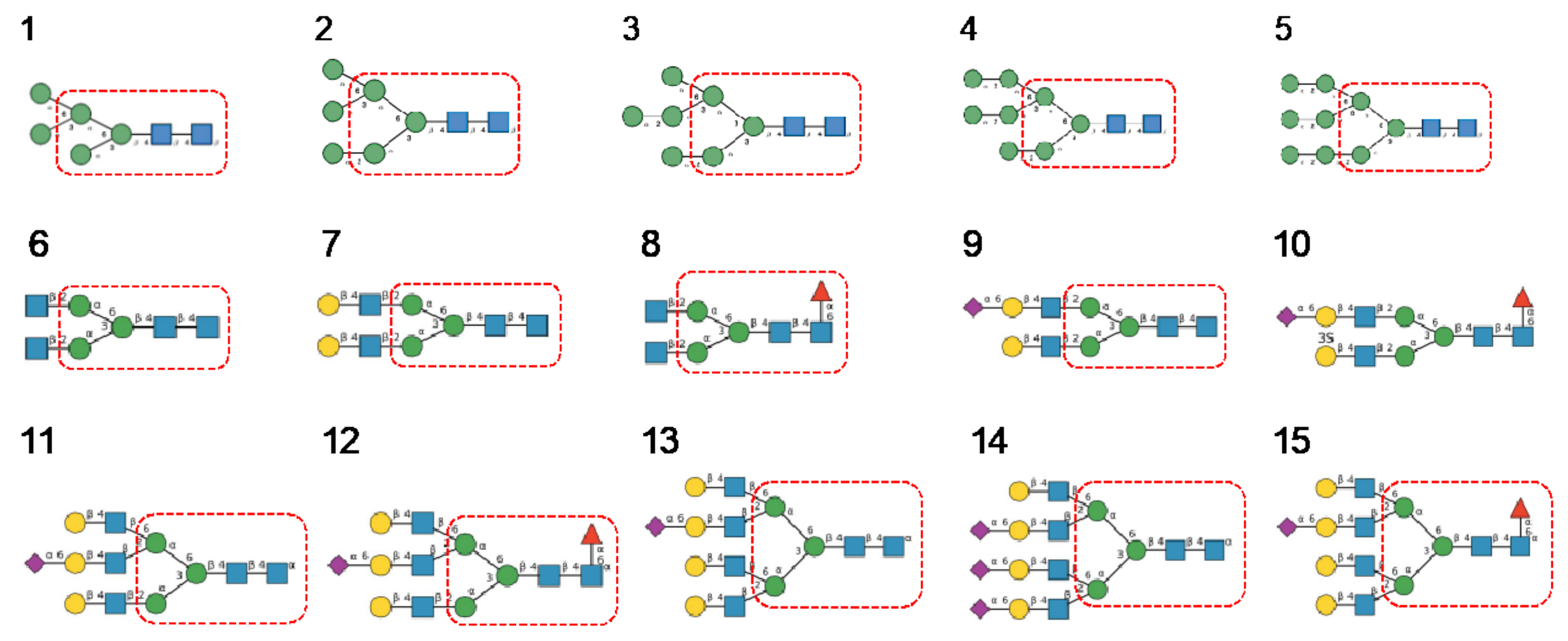
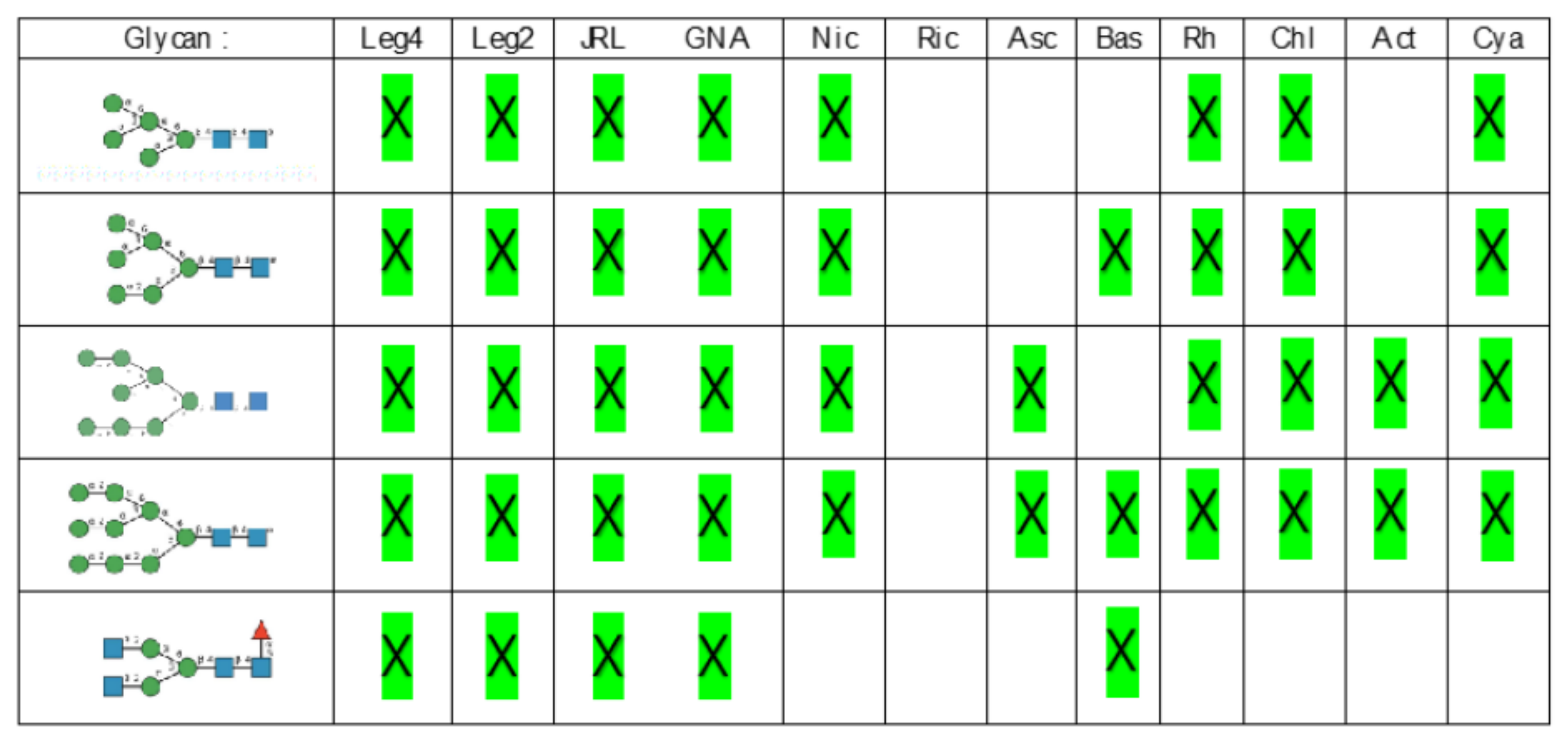
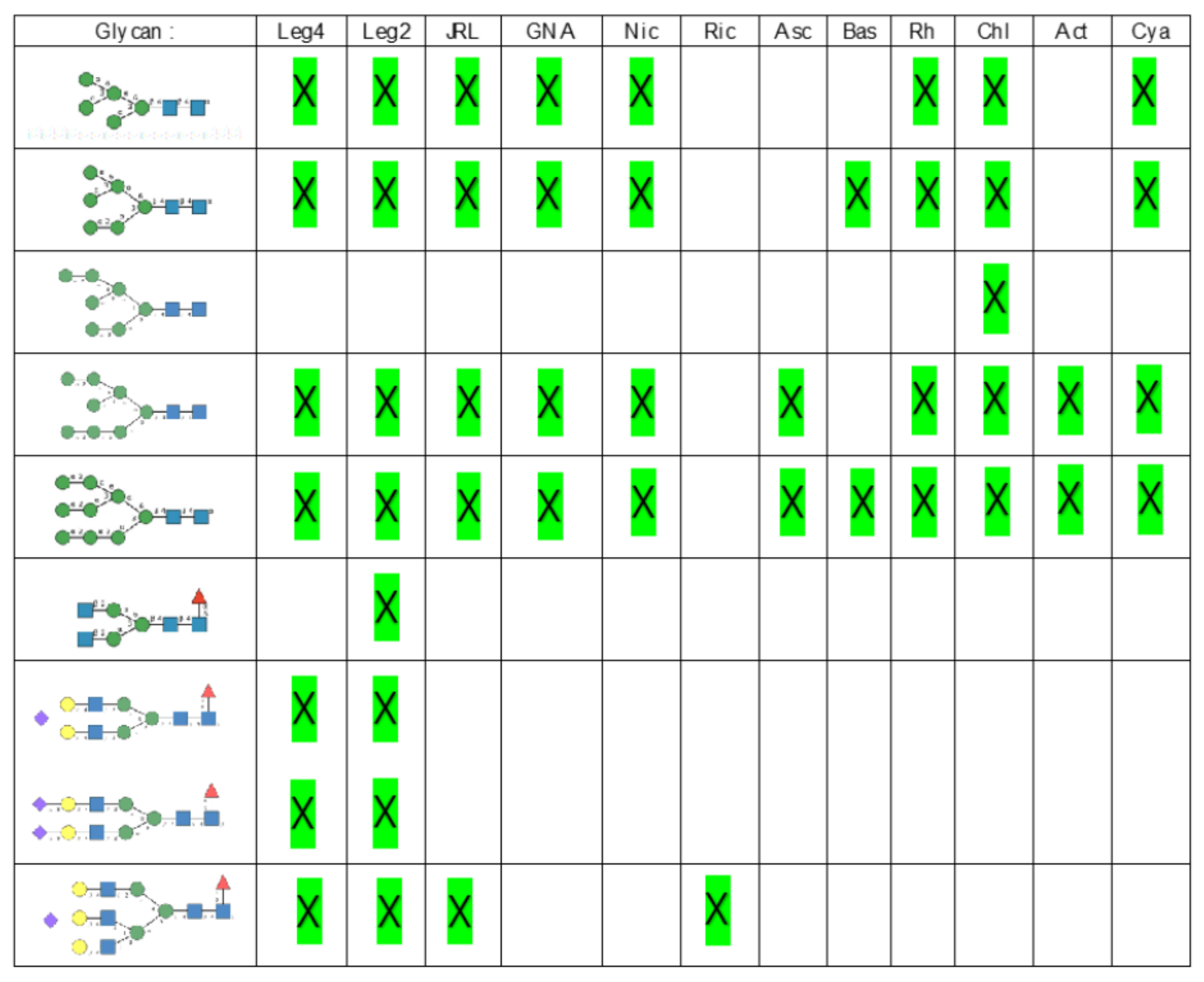
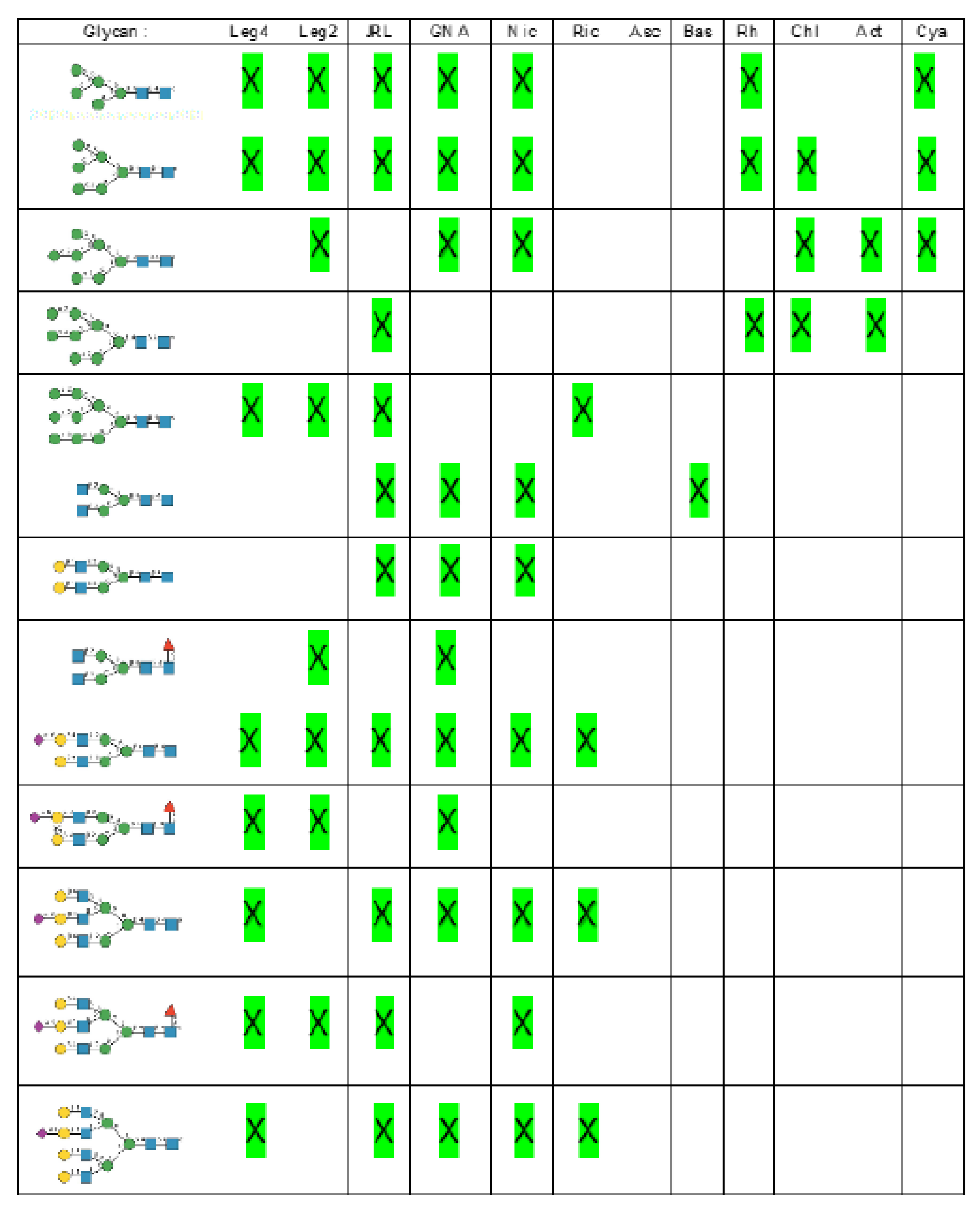
- Man-specific single- and two-chain lectins from the Fabaceae, which primarily recognize the (α3,α6)mannoside core of the branched GlcNAc2Man3 oligosaccharides, readily interact with the complex glycans and, to a lesser extent, with high-mannose glycans of the spikes from SARS-CoV (Figure 31), MERS-CoV (Figure 32) and SARS-CoV-2 (Figure 33). In this respect, two-chain lectins from the Viciae, e.g., pea lectin PsA and lentil lectin LcA, differ from single-chain lectins from other families of Fabaceae, e.g., Con A from Canavalia ensiformis or DGL from Dioclea grandiflora, by a much more pronounced affinity for α6-fucosylated complex glycans [21]. In addition, due to the particularly well-exposed distribution of complex glycans at the top of the SARS-CoV and SARS-CoV-2 spikes, both types of coronaviruses are predicted to better interact with lectins from the Fabaceae, compared to MERS-CoV spikes (Figure 29).
- -
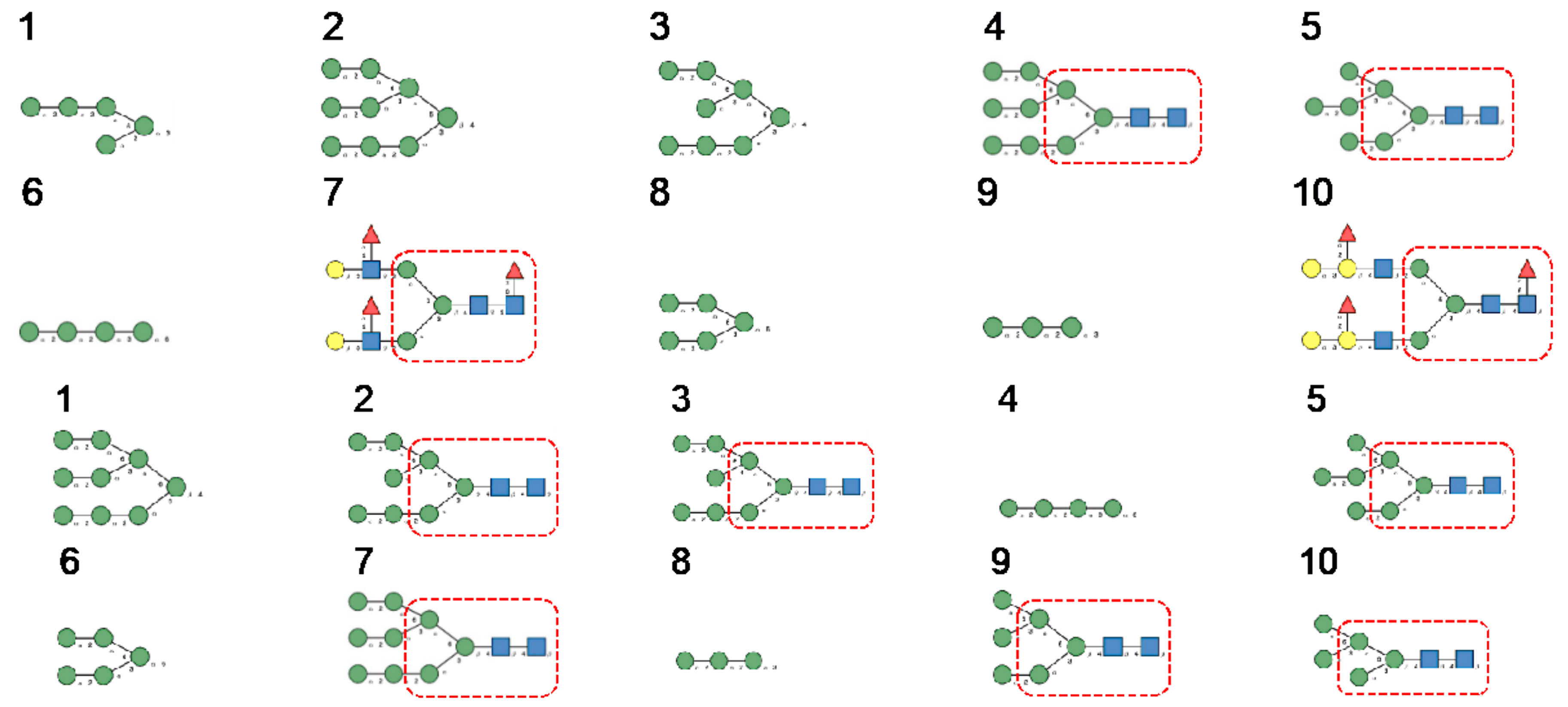
- The Man-specific jacalin-related lectins (JRL), such as Morniga-M from Morus nigra, which also recognizes N-glycans of the hybrid type in addition to complex glycans and high-mannose glycans, could serve as glycan probes for coronaviruses, particularly for binding to MERS-CoV, which exhibits hybrid glycans at the N1288 glycosylation site (Figure 26).
- -
- The GNA-related lectins, which specifically interact with branched Man5-Man9 oligomannosides, would predominantly recognize high-mannose glycans (Figure 31, Figure 32 and Figure 33) and, especially, the well-exposed high-mannose glycans protruding at the top of the MERS-CoV spikes (Figure 29). The less exposed high-mannose glycans located at the bottom of the SARS-CoV and SARS-CoV-2 spikes (Figure 28), should also be recognized, but to a lesser extent.
- -
- Nictaba lectin from Nicotiana tabacum interacts with the following glycans arranged in decreasing order of affinity according to glycan array experiments performed at the Consortium for Functional Glycomics, and should therefore recognize the complex glycans and high-mannose glycans forming the glycan shield of the spikes from coronaviruses (Figure 34):
- -
- The ricin-B lectin from the Iris hollandica RIP-II was also shown to interact with complex (α6-fucosylated) glycans, high-mannose glycans, and hybrid glycans in glycan arrays experiments. Accordingly, it should readily interact with the spikes covering all types of coronaviruses (Figure 31, Figure 32 and Figure 33).
- -
- Man-specific lectins from mushrooms belonging to the Ascomycota, essentially recognize high-mannose glycans (Nc-CVN from Neurospora crassa, and Tb-CVN from Tuber borchii, Flo1p from Saccharomyces pasteurianus, and Flo5A from S. cerevisiae), whereas other Man-specific lectins from Basidiomycota, e.g Abmb from Agaricus bisporus, and MOA from Marasmius oreades, preferentially recognize complex glycans. Mushroom lectins should recognize the high-mannose glycans and the complex glycans from coronaviruses.
- -
- Man-specific lectins from red algae and green algae, both recognize exclusively high-mannose glycans containing Man5-Man9 mannose residues. They should be nicely adapted for the recognition of the high-mannose glycan shield exposed at the top of the MERS-CoV spikes (Figure 28 and Figure 29). Obviously, they should also recognize the high-mannose glycans associated with the SARS-CoV and SARS-CoV-2 spikes (Figure 31, Figure 32 and Figure 33).
- -
- Man-specific lectins from algae, actinobacteria (actinohivin) and cyanobacteria (cyanovirin, microvirin, scytovirin), which predominantly possess a β−barrel structure and display an exclusive specificity for branched Man5-Man9 oligomannosides, should recognize the high-mannose glycan shield covering the coronavirus spikes and, especially, the MERS-CoV virus (Figure 28 and Figure 29).
6. Biomedical Perspectives
7. Bioinformatics
8. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Belouard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef]
- Lu, G.; Wang, Q.; Gao, G.F. Bat-to-human: Spike features determining ‘host jump’ of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol. 2015, 23, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Structure, function, and evolution of coronavirus spike proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Tortorici, M.A.; Bosch, B.J.; Frenz, B.; Rottier, P.J.M.; DiMaio, F.; Rey, F.A.; Veesler, D. Cryo-electron microscopy structure of a coronavirus spike glycoprotein trimer. Nature 2016, 531, 114–117. [Google Scholar] [CrossRef]
- Tortorici, M.A.; Veesler, D. Structural insights into coronavirus entry. Adv. Virus Res. 2019, 105, 93–116. [Google Scholar]
- Glowacka, I.; Bertram, S.; Herzog, P.; Pfefferle, S.; Steffen, I.; Muench, M.O.; Simmons, G.; Hofmann, H.; Kuru, T.; Wever, F.; et al. Differential downregulation of ACE2 by the spike proteins of severe acute respiratory syndrome coronavirus and human coronavirus NL63. J. Virol. 2010, 84, 1198–1205. [Google Scholar] [CrossRef]
- Li, F. Receptor recognition and cross-species infections of SARS coronavirus. Antivir. Res. 2013, 100, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Mou, H.; Smits, S.; Dekkers, D.H.W.; Müller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.A.; Zaki, A.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Kee Wong, S.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.M.; Rottier, P.J.M. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Berndsen, Z.; Raghwani, J.; Seabright, G.; Allen, J.D.; Pybus, O.G.; McLellan, J.S.; Wilson, I.A.; Bowden, T.A.; Ward, A.B.; et al. Vulnerabilities in coronavirus glycan shields despite extensive glycosylation. Nat. Commun. 2020, 11, 2688. [Google Scholar] [CrossRef] [PubMed]
- Shajahan, A.; Supekar, N.T.; Gleinich, A.S.; Azadi, P. Deducing the N- and O-glycosylation profile of the spike protein of novel coronavirus SARS-CoV-2. Glycobiology 2020, 30, 981–988. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; Van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; Van Ranst, M. Plant lectins are potent inhibitors of coronaviruses by interfering with two targets in the viral replication cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef]
- Balzarini, J. Carbohydrate-binding agents: A potential future cornerstone for the chemotherapy of enveloped viruses? Antivir. Chem. Chemother. 2007, 18, 1–11. [Google Scholar] [CrossRef]
- Barre, A.; Van Damme, E.J.M.; Simplicien, M.; Benoist, H.; Rougé, P. Man-specific, GalNAc/T-Tn-specific and Neu5Ac-specific seaweed lectins as glycan probes for the SARS-CoV-2 (COVID19) coronavirus. Mar. Drugs 2019, 18, 543. [Google Scholar]
- Sohrab, S.S.; Suhail, M.; Kamal, M.A.; Ahmad, F.; Azhar, E.I. Emergence of human pathogenic coronaviruses: Lectins as antivirals for SARS-CoV-2. Curr. Pharm. Des. 2020, 26, 5286–5292. [Google Scholar] [CrossRef]
- Kumar Gupta, R.; Apte, G.R.; Bharat Lokhande, K.; Mishra, S.; Pal, J.K. Carbohydrate-binding agents: Potential of repurposing for COVID-19 therapy. Curr. Protein Pept. Sci. 2020, 21, 1085–1096. [Google Scholar] [CrossRef]
- Tsaneva, M.; Van Damme, E.J.M. 130 years of plant lectin research. Glycoconj. J. 2020, 37, 533–551. [Google Scholar] [CrossRef] [PubMed]
- Liener, I.E.; Sharon, N.; Goldstein, I.J. The Lectins, Properties, Functions, and Applications in Biology and Medicine; Academic Press Inc.: Orlando, FL, USA; San Diego, CA, USA; New York, NY, USA; Austin, TX, USA, 1986; pp. 1–600. [Google Scholar]
- Peumans, W.J.; Van Damme, E.J.M.; Barre, A.; Rougé, P. Classification of plant lectins in families of structurally and evolutionary related proteins. Adv. Exp. Med. Biol. 2001, 491, 27–54. [Google Scholar] [PubMed]
- Debray, H.; Decout, D.; Strecker, G.; Spik, G.; Montreuil, J. Specificity of twelve lectins towards oligosaccharides and glycopeptides related to N-glycosylproteins. Eur. J. Biochem. 1981, 117, 41–55. [Google Scholar] [CrossRef]
- Bourne, Y.; Anguille, C.; Fontecilla-Camps, J.C.; Rougé, P.; Cambillau, C. Co-crystallization and preliminary X-ray diffraction studies of Lathyrus ochrus isolectin I with di- and trisaccharides, and a biantennary octasaccharide. J. Mol. Biol. 1990, 213, 211–213. [Google Scholar] [CrossRef]
- Barre, A.; Bourne, Y.; Van Damme, E.J.M.; Peumans, W.J.; Rougé, P. Mannose-binding plant lectins: Different structural scaffolds for a common sugar-recognition process. FEBS Lett. 2001, 83, 645–651. [Google Scholar] [CrossRef]
- Akkouh, O.; Ng, T.B.; Singh, S.S.; Yin, C.; Dan, X.; Chan, Y.S.; Pan, W.; Cheung, R.C.F. Lectins with anti-HIV activity: A review. Molecules 2015, 20, 648–668. [Google Scholar] [CrossRef] [PubMed]
- Levendosky, K.; Mizenina, O.; Martinelli, E.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Kleinbeck, K.; Bonnaire, T.; Robbiani, M.; Zydowsky, T.M.; et al. Griffithsin and carrageenan combination to target Herpes simplex virus 2 and human papilloma virus. Antimicrob. Agents Chemother. 2015, 59, 7290–7298. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, G. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R.; et al. Activity of and effect of subcutaneous treatment with the broad-spectrum antiviral lectin griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef]
- Chiba, H.; Inokoshi, J.; Okamoto, M.; Asanuma, S.; Matsuzaki, K.I.; Iwama, M.; Mizumoto, K.; Tanaka, H.; Oheda, M.; Fujita, K.; et al. Actinohivin, a novel anti-HIV protein from actinomycete that inhibits syncytium formation: Isolation, characterization, and biological activities. Biochem. Biophys. Res. Commun. 2001, 282, 595–601. [Google Scholar] [CrossRef]
- Huskens, D.; Férir, G.; Vermeire, K.; Kehr, J.-C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel α(1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef]
- Balzarini, J.; Schols, D.; Neyts, J.; Van Damme, E.; Peumans, W.; De Clercq, E. α-(1-3)- and α-(1–6)-D-mannose-specific plant lectins are markedly inhibitory to human immunodeficiency virus and cytomegalovirus infections in vitro. Antimicrob. Agents Chemother. 1991, 35, 410–416. [Google Scholar] [CrossRef]
- Balzarini, J.; Neyts, J.; Schols, D.; Hosoya, M.; Van Damme, E.; Peumans, W.; De Clercq, E. The mannose-specific plant lectins from Cymbidium hybrid and Epipactis helleborine and the (N-acetylglucosamine)n-specific plant lectin from Urtica dioica are potent and selective inhibitors of human immunodeficiency virus and cytomegalovirus replication in vitro. Antivir. Res. 1992, 18, 191–207. [Google Scholar]
- Balzarini, J.; Hatse, S.; Vermeire, K.; Princen, K.; Aquaro, S.; Perno, C.F.; De Clercq, E.; Egberink, H.; Vanden Mooter, G.; Peumans, W.; et al. Mannose-specific plant lectins from the Amaryllidaceae family qualify as efficient microbicides for prevention of human immunodeficiency virus infection. Antimicrob. Agents Chemother. 2004, 48, 3858–3870. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Simplicien, M.; Benoist, H.; Van Damme, E.J.M.; Rougé, P. Mannose-specific lectins from marine algae: Diverse structural scaffolds associated to common virucidal and anti-cancer properties. Mar. Drugs 2019, 17, 440. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, K.B.; Gray, E.S.; Lambson, B.E.; nMoore, P.L.; Choge, I.A.; Mlisana, K.; Karim, S.S.; McMahon, J.; O’Keefe, B.; Chikwamba, R.; et al. Mannose-rich glycosylation patterns on HIV-1 subtype C gp120 and sensitivity to lectins, griffithsin, cyanovirin-N and scytovirin. Virology 2010, 402, 187–196. [Google Scholar] [CrossRef]
- Sato, Y.; Morimoto, K.; Kubo, T.; Sakaguchi, T.; Nishizono, A.; Hirayama, M.; Hori, K. Entry inhibition of influenza viruses with high mannose binding lectin ESA-2 from the red alga Eucheuma serra through the recognition of viral hemagglutinin. Mar. Drugs 2015, 13, 3454–3465. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tang, G.; Tang, J.; Wang, L.-X. Targeting N-glycan cryptic sugar moieties for broad-spectrum virus neutralization: Progress in identifying conserved molecular targets in viruses of distinct phylogenetic origins. Molecules 2015, 20, 4610–4622. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antivir. Res. 2017, 142, 37–54. [Google Scholar] [CrossRef]
- Gupta, A.; Gupta, G.S. Status of mannose-binding lectin (MBL) and complement system in COVID-19 patients and therapeutic applications of antiviral plant MBLs. Mol. Cell Biochem. 2021, 21, 1–26. [Google Scholar]
- Animashaun, T.; Hughes, R.C. Bowringia milbraedii agglutinin. Specificity of binding to early processing intermediates of asparagine-linked oligosaccharide and use as a marker of endoplasmic reticulum glycoproteins. J. Biol. Chem. 1989, 264, 4657–4663. [Google Scholar] [CrossRef]
- Souza Teixeira, C.; da Silva, H.C.; de Moura, T.R.; Pereira-Júnior, F.N.; do Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; Delatorre, P.; Rocha, B.A.; Cavada, B.S. Crystal structure of the lectin of Camptosema pedicellatum: Implication of a conservative substitution at the hydrophobic subsite. J. Biochem. 2012, 152, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, B.B.; Goldstein, I.J. Physical and chemical characterization of concanavalin A, the hemagglutinin from jack bean (Canavalia ensiformis). Biochim. Biophys. Acta 1967, 133, 376–379. [Google Scholar] [CrossRef]
- Del Sol, F.G.; Cavada, B.S.; Calvete, J.J. Crystal structures of Cratylia floribunda seed lectin at acidic and basic pHs. Insights into the structural basis of the pH-dependent dimer-tetramer transition. J. Struct. Biol. 2007, 158, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rocha, B.A.; Delatorre, P.; Oliveira, T.M.; Benevides, R.G.; Pires, A.F.; Sousa, A.A.; Souza, L.A.; Assreuy, A.M.; Debray, H.; de Azevedo, X.W.F.; et al. Structural basis for both pro- and anti-inflammatory response induced by mannose-specific legume lectin from Cymbosema roseum. Biochimie 2011, 93, 806–816. [Google Scholar] [CrossRef]
- Rozwarski, D.A.; Swami, B.M.; Brewer, C.F.; Sacchetini, J.C. Crystal structure of the lectin from Dioclea grandiflora complexed with core trimannoside of asparagine-linked carbohydrates. J. Biol. Chem. 1998, 273, 32818–32825. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.C.; Osterne, V.J.; Santiago, M.Q.; Pinto-Junior, V.R.; Silva-Filho, J.C.; Lossio, C.F.; Nascimento, F.L.; Almeida, R.P.; Teixeira, C.S.; Leal, R.B.; et al. Structural analysis of Centrolobium tomentosum seed lectin with inflammatory activity. Arch. Biochem. Biophys. 2016, 596, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Abergel, C.; Cambillau, C.; Frey, M.; Rougé, P.; Fontecilla-Camps, J.C. X-ray crystal structure determination and refinement at 1.9 Å resolution of isolectin I from the seeds of Lathyrus ochrus. J. Mol. Biol. 1990, 214, 571–584. [Google Scholar] [CrossRef]
- Foriers, A.; Van Driessche, E.; De Neve, R.; Kanarek, L.; Strosberg, A.D. The subunit structure and N-terminal sequences of the α- and β-subunits of the lentil lectin (Lens culinaris). FEBS Lett. 1977, 75, 237–240. [Google Scholar] [CrossRef]
- Gao, S.; An, J.; Wu, C.F.; Gu, Y.; Chen, F.; Yu, Y.; Wu, Q.Q.; Bao, J.K. Effect of amino acid residue and oligosaccharide chain chemical modifications on spectral and hemagglutinating activity of Millettia dielsiana Harms. ex Diels. lectin. Acta Biochim. Biophys. Sin. 2005, 37, 47–54. [Google Scholar] [CrossRef]
- Young, N.M.; Williams, R.E.; Roy, C.; Yaguchi, M. Structural comparison of the lectin from sainfoin (Onobrychis viciifolia) with concanavalin A and other D-mannose specific lectins. Can. J. Biochem. 1982, 60, 933–941. [Google Scholar] [CrossRef]
- Einspahr, H.; Pareks, E.H.; Suguna, K.; Subramanian, E.; Suddath, F.L. The crystal structure of pea lectin at 3.0-Å resolution. J. Biol. Chem. 1986, 261, 16518–16527. [Google Scholar] [CrossRef]
- Cavada, B.S.; Araripe, D.A.; Silva, I.B.; Pinto-Junior, V.R.; Osterne, V.J.S.; Neco, A.H.B.; Laranjeira, E.P.P.; Lossio, C.F.; Correia, J.L.A.; Pires, A.F.; et al. Structural studies and nociceptive activity of a native lectin from Platypodium elegans seeds (nPELa). Int. J. Biol. Macromol. 2018, 107, 236–246. [Google Scholar] [CrossRef]
- Pereira-Junior, F.N.; Silva, H.C.; Freitas, B.T.; Rocha, B.A.; Nascimento, K.S.; Nagano, C.S.; Leal, R.B.; Sampaio, A.H.; Cavada, B.S. Purification and characterization of a mannose/N-acetyl-D-glucosamine-specific lectin from the seeds of Platymiscium floribundum Vogel. J. Mol. Recognit. 2012, 25, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Loris, R.; Imberty, A.; Beeckmans, S.; Van Driessche, E.; Read, J.S.; Bouckaert, J.; De Greve, H.; Buts, L.; Wyns, L. Crystal structure of Pterocarpus angolensis lectin in complex with glucose, sucrose, and turanose. J. Biol. Chem. 2003, 278, 16297–16303. [Google Scholar] [CrossRef]
- Naeem, A.; Ahmad, E.; Ashraf, M.T.; Khan, R.H. Purification and characterization of mannose/glucose-specific lectin from seeds of Trigonella foenumgraecum. Biochemistry 2007, 72, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Reeke, G.N., Jr.; Becker, J.W. Three-dimensional structure of favin: Saccharide binding-cyclic permutation in leguminous lectins. Science 1986, 234, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peumans, W.J.; Hause, B.; Bras, J.; Kumar, M.; Proost, P.; Barre, A.; Rougé, P.; Van Damme, E.J.M. Jasmonic acid methyl ester induces the synthesis of a cytoplasmic/nuclear chito-oligosaccharide binding lectin in tobacco leaves. FASEB J. 2002, 16, 905–907. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Briké, F.; Winter, H.C.; Van Leuven, F.; Goldstein, I.J.; Peumans, W.J. Molecular cloning of two different mannose-binding lectins from tulip bulbs. Eur. J. Biochem. 1996, 236, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Meagher, J.L.; Winter, H.C.; Ezell, P.; Goldstein, I.J.; Stuckey, J.A. Crystal structure of banana lectin reveals a novel second sugar binding site. Glycobiology 2005, 15, 1033–1042. [Google Scholar] [CrossRef]
- Bourne, Y.; Roig-Zamboni, V.; Barre, A.; Peumans, W.J.; Astoul, C.H.; Van Damme, E.J.; Rougé, P. The crystal structure of the Calystegia sepium agglutinin reveals a novel quaternary arrangement of lectin subunits with a β-prism fold. J. Biol. Chem. 2004, 279, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K.; Ashida, H.; Uemura, N.; Kushibe, S.; Ozaki, T.; Yoshida, M. Purification and characterization of a mannose/glucose-specific lectin from Castanea crenata. Phytochemistry 1998, 49, 667–673. [Google Scholar] [CrossRef]
- Nakamura-Tsuruta, S.; Uchiyama, N.; Peumans, W.J.; Van Damme, E.J.M.; Totani, K.; Ito, Y.; Hirabayashi, J. Analysis of the sugar-binding specificity of mannose-binding-type jacalin-related lectins by frontal affinity chromatography—An approach to functional classification. FEBS J. 2008, 275, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- Azarkan, M.; Feller, G.; Vandenameele, J.; Herman, R.; El Mahyaoui, R.; Sauvage, E.; Vanden Broeck, A.; Matagne, A.; Charlier, P.; Kerff, F. Biochemical and structural characterization of a mannose-binding jacalin-related lectin with two-sugar binding sites from pineapple (Anana comosus) stem. Sci. Rep. 2018, 8, 11508. [Google Scholar] [CrossRef]
- Misquith, S.; Rani, P.G.; Surolia, A. Carbohydrate binding specificity of the B-cell maturation mitogen from Artocarpus integrifolia seeds. J. Biol. Chem. 1994, 269, 30393–30401. [Google Scholar] [CrossRef]
- Barre, A.; Peumans, W.J.; Rossignol, M.; Borderies, M.; Culerrier, R.; Van Damme, E.J.M.; Rougé, P. Artocarpin is a polyspecific jacalin-related lectin with a monosaccharide preference for mannose. Biochimie 2004, 86, 685–691. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, F.D.; Bezerra da Silva, B.; Pessoa Furtado, G.; de Sa Carneiro, I.; Lobo, M.D.P.; Guan, Y.; Guo, J.; Coker, A.R.; Lourenzoni, M.R.; Florindo Guedes, M.I.; et al. Frutapin, a lectin from Artocarpus incisa (breadfruit): Cloning, expression and molecular analysis. Biosci. Rep. 2017, 37, BSR20170969. [Google Scholar] [CrossRef]
- Chang, W.C.; Liu, K.L.; Hsu, F.C.; Jeng, S.T.; Cheng, Y.S. Ipomoelin, a jacalin-related lectin with a compact tetrameric association and versatile carbohydrate binding properties regulated by its N terminus. PLoS ONE 2012, 7, e40618. [Google Scholar] [CrossRef]
- Rabijns, A.; Barre, A.; Van Damme, E.J.M.; Peumans, W.J.; De Ranter, C.J.; Rougé, P. Structural analysis of the jacalin-related lectin Morniga-M from the black mulberry (Morus nigra) in complex with mannose. FEBS J. 2005, 272, 3725–3732. [Google Scholar] [CrossRef]
- Mann, K.; Farias, C.M.; Del Sol, F.G.; Santos, C.F.; Grangeiro, T.B.; Nagano, C.S.; Cavada, B.S.; Calvete, J.J. The amino acid sequence of the glucose/mannose-specific lectin isolated from Parkia platycephala seeds reveals three tandemly arranged jacalin-related domains. Eur. J. Biochem. 2001, 268, 4414–4422. [Google Scholar] [CrossRef]
- Bourne, Y.; Zamboni, V.; Barre, A.; Peumans, W.J.; Van Damme, E.J.; Rougé, P. Helianthus tuberosus lectin reveals a widespread scaffold for mannose-binding lectins. Structure 1999, 7, 1473–1482. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Smeets, K.; Torrekens, S.; van Leuven, F.; Goldstein, I.J.; Peumans, W.J. The closely related homomeric and heterodimeric mannose-binding lectins from garlic are encoded by one-domain and two-domain lectin genes, respectively. Eur. J. Biochem. 1992, 206, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-F.; An, J.; He, X.-J.; Deng, J.; Hong, Z.-X.; Liu, C.; Hong-Zhou, L.; Li, Y.-J.; Wang, C.-J.; Chen, F.; et al. Molecular cloning of a novel mannose-binding lectin gene from bulbs of Amaryllis vittata (Amaryllidaceae). Acta Bot. Sin. 2004, 46, 1301–1306. [Google Scholar]
- Lin, J.; Zhou, X.; Pang, Y.; Gao, H.; Fei, J.; Shen, G.A.; Wang, J.; Li, X.; Sun, X.; Tang, K. Cloning and characterization of an agglutinin gene from Arisaema lobatum. Biosci. Rep. 2005, 25, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Goossens, K.; Smeets, K.; Van Leuven, F.; Verhaert, P.; Peumans, W.J. The major tuber storage protein of Araceae species is a lectin. Characterization and molecular cloning of the lectin from Arum maculatum L. Plant Physiol. 1995, 107, 1147–1158. [Google Scholar] [CrossRef]
- Xu, X.; Wu, C.; Liu, C.; Luo, C.; Luo, Y.; Li, J.; Zhao, X.; Van Damme, E.; Bao, J. Purification and characterization of a mannose-binding lectin from the rhizomes of Aspidistra elatior Blume with antiproliferative activity. Acta Biochim. Biophys. Sin. 2007, 39, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.R.; Winter, H.C.; Vericimo, M.A.; Meagher, J.L.; Stckey, J.A.; Goldstein, I.J.; Paschoalin, V.M.; Silva, J.T. Structural analysis and binding properties of isoforms of tarin, the GNA-related lectin from Colocasia esculenta. Biochim. Biophys. Acta 2015, 1854, 20–30. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Smeets, K.; Torrekens, S.; Van Leuven, F.; Peumans, W.J. Characterization and molecular cloning of mannose-binding lectins from the Orchidaceae species Listera ovata, Epipactis helleborine and Cymbidium hybrid. Eur. J. Biochem. 1994, 221, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, X.; Tang, K. Cloning and expression of a novel cDNA encoding a mannose-binding lectin from Dendrobium officinale. Toxicon 2005, 45, 535–540. [Google Scholar] [CrossRef]
- Kaku, H.; Van Damme, E.J.; Peumans, W.J.; Goldstein, I.J. Carbohydrate-binding specificity of the daffodil (Narcissus pseudonarcissus) and amaryllis (Hippeastrum hybr.) bulb lectins. Arch. Biochem. Biophys. 1990, 279, 298–304. [Google Scholar] [CrossRef]
- Liu, J.; Xu, X.; Liu, J.; Balzarini, J.; Luo, Y.; Kong, Y.; Li, J.; Chen, F.; Van Damme, E.J.; Bao, J. A novel tetrameric lectin from Lycoris aurea with four mannose binding sites per monomer. Acta Biochim. Pol. 2007, 54, 159–166. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakamoto, H.; Tateno, H.; Hirabayashi, J.; Oguri, S. Purification, characterization, and molecular cloning of lectin from winter buds of Lysichiton camtschatcensis (L.) Schott. Biosci. Biotechnol. Biochem. 2012, 76, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.; Sun, S.S.; Ooi, V.E. Purification and characterization of a new antiviral protein from the leaves of Pandanus amaryllifolius (Pandanaceae). Int. J. Biochem. Cell Biol. 2004, 36, 1440–1446. [Google Scholar] [CrossRef]
- Yao, J.H.; Zhao, X.Y.; Liao, Z.H.; Lin, J.; Chen, Z.H.; Chen, F.; Song, J.; Sun, X.F.; Tang, K.X. Cloning and molecular characterization of a novel lectin gene from Pinellia ternata. Cell Res. 2003, 13, 301–308. [Google Scholar] [CrossRef]
- Van Damme, E.J.; Barre, A.; Rougé, P.; Van Leuven, F.; Balzarini, J.; Peumans, W.J. Molecular cloning of the lectin and a lectin-related protein from common Solomon’s seal (Polygonatum multiflorum). Plant Mol. Biol. 1996, 31, 657–672. [Google Scholar] [CrossRef]
- Ding, J.; Bao, J.; Zhu, D.; Zhang, Y.; Wang, D.C. Crystal structure of a novel anti-HIV mannose-binding lectin from Polygonatum cyrtonema Hua with unique ligand-binding property and super-structure. J. Struct. Biol. 2010, 171, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Shetty, K.N.; Bhat, G.G.; Inamdar, S.R.; Swamy, B.M.; Suguna, K. Crystal structure of a β-prism II lectin from Remusatia vivipara. Glycobiology 2012, 22, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.M.; Wood, S.D.; Reynolds, C.D.; Rizkallah, P.J.; Peumans, W.J.; Van Damme, E.J.; Allen, A.K. Purification, crystallization and preliminary X-ray analysis of a mannose-binding lectin from bluebell (Scilla campanulata) bulbs. Acta Crystallogr. D Biol. Crystallogr. 1996, 52, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.S.; Sun, S.S.; Wang, H.; Ooi, V.E. New mannose-binding lectin isolated from the rhizome of Sarsaparilla Smilax glabra Roxb. (Liliaceae). J. Agric. Food Chem. 2004, 52, 6091–6095. [Google Scholar] [CrossRef]
- Luo, Y.; Xu, X.; Liu, J.; Li, J.; Sun, Y.; Liu, Z.; Liu, J.; Van Damme, E.; Balzarini, J.; Bao, J. A novel mannose-binding tuber lectin from Typhonium divaricatum (L.) Decne (family Araceae) with antiviral activity against HSV-II and anti-proliferative effect on human cancer cell lines. J. Biochem. Mol. Biol. 2007, 40, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Pang, Y.; Liu, X.; Wang, X.; Deng, Z.; Sun, X.; Tang, K. Molecular cloning and characterization of a novel mannose-binding lectin cDNA from Zantedeschia aethiopica. Biocell 2005, 29, 187–193. [Google Scholar] [CrossRef]
- Koike, T.; Titani, K.; Suzuki, M.; Beppu, H.; Kuzuya, H.; Maruta, K.; Shimpo, K.; Fujita, K. The complete amino acid sequence of a mannose-binding lectin from “Kidachi Aloe” (Aloe arborescens Miller var. natalensis Berger). Biochem. Biophys. Res. Commun. 1995, 214, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Astoul, C.H.; Barre, A.; Rougé, P.; Peumans, W.J. Cloning and characterization of a monocot mannose-binding lectin from Crocus vernus (family Iridaceae). Eur. J. Biochem. 2000, 267, 5067–5077. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.; Kaku, H.; Perini, F.; Goldstein, I.J.; Peeters, B.; Yagi, F.; Decock, B.; Peumans, W.J. Biosynthesis, primary structure and molecular cloning of snowdrop (Galanthus nivalis L.) lectin. Eur. J. Biochem. 1991, 202, 23–30. [Google Scholar] [CrossRef]
- Wu, C.-F.; Li, J.; An, J.; Chang, L.-Q.; Che, F.; Bao, J.-K. Purification, biological activities, and molecular cloning of a novel mannose-binding lectin from bulbs of Zephyranthes candida herb (Amaryllidaceae). J. Integr. Plant Biol. 2006, 48, 223–231. [Google Scholar] [CrossRef]
- Lannoo, N.; Peumans, W.J.; Van Pamel, E.V.; Alvarez, R.; Xiong, T.C.; Hause, G.; Mazars, C.; Van Damme, E.J. Localization and in vitro binding studies suggest that the cytoplasmic/nuclear tobacco lectin can interact with high-mannose and complex N-glycans. FEBS Lett. 2006, 580, 6329–6337. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Van Damme, E.J.; Hause, B.; Barre, A.; Chen, Y.; Rougé, P.; Peumans, W.J. Iris bulbs express type 1 and type 2 ribosome-inactivating proteins with unusual properties. Plant Physiol. 2001, 125, 866–876. [Google Scholar] [CrossRef]
- Schouppe, D.; Rougé, P.; Lasanajak, Y.; Barre, A.; Smith, D.F.; Proost, P.; Van Damme, E.J.M. Mutational analysis of the carbohydrate binding activity of the tobacco lectin. Glycoconj. J. 2010, 27, 613–623. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peumans, W.J.; Barre, A.; Bras, J.; Rougé, P.; Proost, P.; Van Damme, E.J. The liverwort contains a lectin that is structurally and evolutionary related to the monocot mannose-binding lectins. Plant Physiol. 2002, 129, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Yamasaki, T.; Tateno, H. The trimeric solution structure and fucose-binding mechanism of the core fucosylation-specific lectin PhoSL. Sci. Rep. 2018, 8, 7740. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Winter, H.C.; Petryniak, J.; Goldstein, I.J. Purification, characterization, molecular cloning, and expression of novel members of jacalin-related lectins from rhizomes of the true fern Phlebodium aureum (L) J. Smith (Polypodiaceae). J. Biol. Chem. 2003, 278, 10891–10899. [Google Scholar] [CrossRef]
- Yagi, F.; Iwaya, T.; Haraguchi, T.; Goldstein, I.J. The lectin from leaves of Japanese cycad, Cycas revoluta Thunb. (gymnosperm) is a member of the jacalin-related family. Eur. J. Biochem. 2002, 269, 4335–4341. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, T.; Hatano, K.; Miyauchi, Y.; Suwa, Y.; Sawano, Y.; Tanokura, M. A secreted protein with plant-specific cysteine-rich motif functions as a mannose-binding lectin that exhibits antifungal activity. Plant Physiol. 2014, 166, 766–778. [Google Scholar] [CrossRef]
- Francis, F.; Jaber, K.; Colinet, F.; Portetelle, D.; Haubruge, E. Purification of a new mannose-specific lectin from Penicillium chrysogenum and its aphicidal properties. Fungal Biol. 2011, 115, 1093–1099. [Google Scholar] [CrossRef]
- Goossens, K.V.Y.; Ielasi, F.S.; Nookaew, I.; Stals, I.; Alonso-Sarduy, L.; Daenen, L.; Van Mulders, S.E.; Stassen, C.; van Eijsden, R.G.E.; Siewers, V.; et al. Molecular mechanism of flocculation self-recognition in yeast and its role in mating and survival. mBio 2015, 6, e00427-15. [Google Scholar] [CrossRef] [PubMed]
- Veelders, M.; Brückner, S.; Ott, D.; Unverzagt, C.; Mösch, H.U.; Essen, L.O. Structural basis of flocculin-mediated social behavior in yeast. Proc. Natl. Acad. Sci. USA 2010, 107, 22511–22516. [Google Scholar] [CrossRef] [PubMed]
- Rachmawati, H.; Sundari, S.; Nabila, N.; Tandrasasmita, O.M.; Amalia, R.; Siahaan, T.J.; Tjandrawinata, R.T.; Ismaya, W. Orf239342 from the mushroom Agaricus bisporus is a mannose binding protein. Biochem. Biophys. Res. Commun. 2019, 515, 99–103. [Google Scholar] [CrossRef]
- Suzuki, T.; Sugiyama, K.; Hirai, H.; Ito, H.; Morita, T.; Dohra, H.; Murata, T.; Usui, T.; Tateno, H.; Hirabayashi, J.; et al. Mannose-specific lectin from the mushroom Hygrophorus russula. Glycobiology 2012, 22, 616–629. [Google Scholar] [CrossRef]
- Wohlschlager, T.; Titz, A.; Künzler, M.; Varrot, A. Expression, purification, and functional characterization of tectonin 2 from Laccaria bicolor: A six-bladed b-propeller lectin specific for O-methylated glycans. Methods Mol. Biol. 2020, 2132, 669–682. [Google Scholar] [PubMed]
- Sommer, R.; Makshakova, O.N.; Wohlschlager, T.; Hutin, S.; Marsh, M.; Titz, A.; Künzler, M.; Varrot, A. Crystal structure of fungal tectonin in complex with O-methylated glycans suggest key role in innate immune defense. Structure 2018, 26, 391–402. [Google Scholar] [CrossRef]
- Shimokawa, M.; Fukudome, A.; Yamashita, R.; Minami, Y.; Yagi, F.; Tateno, H.; Hirabayashi, J. Characterization and cloning of GNA-like lectin from the mushroom Marasmius oreades. Glycoconj. J. 2012, 29, 457–465. [Google Scholar] [CrossRef]
- Koharudin, L.M.; Viscomi, A.R.; Jee, J.G.; Ottonello, S.; Gronenborn, A.M. The evolutionary conserved family of cyanovirin-N homologs: Structures and carbohydrate specificity. Structure 2008, 16, 570–584. [Google Scholar] [CrossRef] [PubMed]
- Chaves, R.P.; da Silva, S.R.; Nascimento Neto, L.G.; Carneiro, R.F.; Coelho da Silva, A.L.; Sampaio, A.H.; Lopes de Sousa, B.; Cabral, M.G.; Videira, P.A.; Teixeira, E.H.; et al. Structural characterization of two isolectins from the marine red alga Solieria filiformis (Kützing) P.W. Gabrielson and their anticancer effect on MCF-7 breast cancer cells. Int. J. Biol. Macromol. 2018, 107, 1320–1329. [Google Scholar] [CrossRef] [PubMed]
- Kawakubo, A.; Makino, H.; Ohnishi, J.; Hirohara, H.; Kanji, H. Occurrence of highly yielded lectins homologous within genus Eucheuma. J. Appl. Phycol. 1999, 11, 149–156. [Google Scholar] [CrossRef]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.D.; Sato, Y.; Hori, K. High-mannose N-glycan-specific lectin from the red alga Kappaphycus striatum (Carrageenophyte). Phytochemistry 2011, 72, 855–861. [Google Scholar] [CrossRef]
- Chaves, R.P.; da Silva, S.R.; da Silva, J.P.F.A.; Carneiro, R.F.; de Sousa, B.L.; Abreu, J.O.; de Carvalho, F.C.T.; Rocha, C.R.C.; Farias, W.R.L.; de Sousa, O.V.; et al. Meristiella echinocarpa lectin (MEL): A new member of the OAAH-lectin family. J. Appl. Phycol. 2018, 30, 2629–2638. [Google Scholar] [CrossRef]
- Van Holle, S.; Van Damme, E.J.M. Messages from the past: New insights in plant lectin evolution. Front. Plant Sci. 2019, 10, 36. [Google Scholar] [CrossRef]
- Suttisrisung, S.; Senapin, S.; Withyachumnarnkul, B.; Wongprasert, K. Identification and characterization of a novel legume-like lectin cDNA sequence from the red marine algae Gracilaria fisheri. J. Biosci. 2011, 36, 833–843. [Google Scholar] [CrossRef]
- Hori, K.; Miyazawa, K.; Ito, K. Isolation and characterization of glycoconjugate-specific isoagglutinins from a marine green alga Boodlea coacta (Dickie) Murray et De Toni. Bot. Mar. 1986, XXIX, 323–328. [Google Scholar] [CrossRef]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High-mannose-binding lectin with preference for the cluster of α1-2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef]
- Derelle, E.; Ferraz, C.; Rombauts, S.; Rouze, P.; Worden, A.Z.; Robbens, S.; Partensky, F.; Degroeve, S.; Echeynie, S.; Cooke, R.; et al. Genome analysis of the smallest free-living eukaryote Ostreococcus tauri unveils many unique features. Proc. Natl. Acad. Sci. USA 2006, 103, 11647–11652. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga Halimeda renschii exhibits a potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, 255. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, A.L.; Sanz, L.; Sánchez, E.L.; Wolfenstein-Todel, C.; Calvete, J.J. Isolation of two novel mannan- and L-fucose-binding lectins from the green alga Enteromorpha prolifera: Biochemical characterization of EPL-2. Arch. Biochem. Biophys. 2003, 415, 245–250. [Google Scholar] [CrossRef]
- Hwang, H.J.; Han, J.W.; Jeon, H.; Han, J.W. Induction of recombinant lectin expression by an artificially constructed tandem repeat structure: A case study using Bryopsis plumosa mannose-binding lectin. Biomolecules 2018, 8, 146. [Google Scholar] [CrossRef]
- Corteggiani Carpinelli, E.; Telatin, A.; Vitulo, N.; Forcato, C.; D’Angelo, M.; Schiavon, R.; Vezzi, A.; Giacometti, G.M.; Morosinotto, T.; Valle, G. Chromosome scale genome assembly and transcriptome profiling of Nannochloropsis gaditana in nitrogen depletion. Mol. Plant. 2014, 7, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Cai, M.; Ray, S.; Ghirlando, R.; Yamaguchi, M.; Muramoto, K. New carbohydrate specificity and HIV-1 fusion blocking activity of the cyanobacterial protein MVL: NMR, ITC and sedimentation equilibrium studies. J. Mol. Biol. 2004, 339, 901–914. [Google Scholar] [CrossRef]
- Sato, Y.; Okuyama, S.; Hori, K. Primary structure and carbohydrate binding specificity of a potent anti-HIV lectin isolated from the filamentous cyanobacterium, Oscillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.; Gardella, R.S.; Sowder, R.C.; Turmin, J.; Watson, K.; Buckheit, R.W., Jr.; et al. A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef]
- Hori, K.; Sato, Y.; Ito, K.; Fujiwara, Y.; Iwamoto, Y.; Makino, H.; Kawakubo, A. Strict specificity for high-mannose type N-glycans and primary structure of a red alga Eucheuma serrata lectin. Glycobiology 2007, 17, 479–491. [Google Scholar] [CrossRef]
- Wang, P.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; Graham, B.S.; et al. Increased resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7 to antibody neutralization. Cell Host Microbe 2021, 29, 1–5. [Google Scholar] [CrossRef]
- van der Meer, F.J.U.M.; de Haan, C.A.M.; Schuurman, N.M.P.; Haijema, B.J.; Peumans, W.J.; Van Damme, E.J.M.; Delputte, P.L.; Balzarini, J.; Egberink, H.F. Antiviral activity of carbohydrate-binding agents against Nidovirales in cell cultures. Antivir. Res. 2007, 76, 21–29. [Google Scholar] [CrossRef]
- Liu, Y.-M.; Shahed-Al-Mahmud, M.; Chen, X.; Chen, T.-H.; Liao, K.-S.; Lo, J.M.; Wu, Y.-M.; Ho, M.-C.; Wu, C.-Y.; Wong, C.-H.; et al. A carbohydrate-binding protein from the edible lablab beans effectively blocks the infections of influenza viruses and SARS-CoV-2. Cell Rep. 2020, 32, 108016. [Google Scholar] [CrossRef] [PubMed]
- Debray, H.; Rougé, P.; Debray, H.; Rougé, P. The fine sugar specificity of the Lathyrus ochrus seed lectin and isolectins. FEBS Lett. 1994, 176, 120–124. [Google Scholar] [CrossRef]
- Bourne, Y.; Mazurier, J.; Legrand, D.; Rougé, P.; Montreuil, J.; Spik, G.; Cambillau, C. Structure of a legume lectin complexed with the human lactotransferrin N2 fragment, and with an isolated biantennary glycopeptide: Role of the fucose moiety. Structure 1994, 2, 209–219. [Google Scholar] [CrossRef]
- Zlółkowska, N.E.; O’Keefe, B.R.; Mori, T.; Zhu, C.; Giomarelli, B.; Vojdani, F.; Palmer, K.E.; McMahon, J.B.; Wlodawer, A. Domain-swapped structure of the potent antiviral protein griffithsin and its mode of carbohydrate binding. Structure 2006, 14, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Vohlford-Lenane, C.L.; et al. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Séron, K.; Labitt, R.N.; Danneels, A.; Palmer, K.E.; Whittaker, G.R.; Dubuisson, J.; Belouzard, S. Middle East respiratory syndrome coronavirus infection is inhibited by griffithsin. Antivir. Res. 2016, 133, 1–8. [Google Scholar] [CrossRef]
- Rahimi, N. C-type lectin CD209L/L-SIGN and CD209/DC-SIGN; Cell adhesion molecules turned to pathogen recognition receptors. Biology 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Roehrich, H.; Yuan, C.; Hou, J.H. Immunohistochemical study of SARS-CoV-2 viral entry factors in the cornea and ocular surface. Cornea 2020, 39, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Lenza, M.P.; Oyenarte, I.; Diercks, T.; Quintana, J.I.; Gimeno, A.; Coelho, H.; Diniz, A.; Peccati, F.; Delgado, S.; Bosch, A.; et al. Structural characterization of the N-linked glycans in the receptor binding domain of the SARS-CoV-2 spike protein and their interactions with human lectins using NMR spectroscopy. Angew. Chem. Int. Ed. 2020, 59, 23763–23771. [Google Scholar] [CrossRef] [PubMed]
- Caniglia, J.L.; Guda, M.R.; Asuthkar, S.; Tsung, A.J.; Velpula, K.K. A potential role for galectin-3 inhibitors in the treatment of COVID-19. PeerJ 2020, 8, e9392. [Google Scholar] [CrossRef] [PubMed]
- van der Meer, F.J.U.M.; de Haan, C.A.M.; Schuurman, N.M.P.; Haijema, B.J.; Verheije, M.H.; Bosch, B.J.; Balzarini, J.; Egberink, H.F. The carbohydrate-binding plant lectins and the non-peptidic antibiotic pradimicin a target the glycans of the coronavirus envelope glycoproteins. J. Antimicrob. Chemother. 2007, 60, 741–749. [Google Scholar] [CrossRef]
- Casalino, L.; Gaieb, Z.; Goldsmith, J.A.; Hjorth, C.K.; Dommer, A.C.; Harbison, A.M.; Fogarty, C.A.; Barros, E.P.; Taylor, B.C.; McLellan, J.S.; et al. Beyond shielding: The roles of glycans in SARS-CoV-2 spike protein. ACS Cent. Sci. 2020, 6, 1722–1734. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.L.; Hume, S.D.; Bratcher, B.; et al. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical micrbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef]
- Leblanc, J.-F.; Germain, M.; Delage, G.; O’Brien, S.; Drews, S.J.; Lewin, A. Risk of transmission of severe acute respiratory syndrome coronavirus 2 by transfusion: A literature review. Transfusion 2020, 60, 3046–3054. [Google Scholar] [CrossRef] [PubMed]
- Gaussen, A.; Hornby, L.; Rockl, G.; O’Brien, S.; Delage, G.; Sapir-Pichhadze, R.; Drews, S.J.; Weiss, M.J.; Lewin, A. Evidence of SARS-CoV-2 infection in cells, tissues and organs and the risk of transmission through transplantation. Transplantation 2021, 105, 1405–1422. [Google Scholar] [CrossRef]
- Koch, B.; Schult-Dietrich, P.; Büttner, S.; Dilmaghani, B.; Lohmann, D.; Baer, P.C.; Dietrich, U.; Geiger, H. Lectin affinity plasmapheresis for middle east respiratory syndrome-coronavirus and Marburg virus glycoprotein elimination. Blood Purif. 2018, 46, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 244, 108693. [Google Scholar] [CrossRef]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the precision of comparative models with YASARA NOVA—A self-parametrizing force field. Proteins 2002, 47, 393–402. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemistry of protein structures. J. Appl. Crystallogr. 1993, 126, 283–291. [Google Scholar] [CrossRef]
- Melo, F.; Feytmans, E. Assessing protein structures with a non-local atomic interaction energy. J. Mol. Biol. 1998, 277, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasiin, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. Fast docking using the CHARMM force field with EADock DSS. J. Comput. Chem. 2011, 32, 2149–2159. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Böhm, M.; Bohne-Lang, A.; Frank, M.; Loss, A.; Rojas-Macias, M.A.; Lütteke, T. Glycosciences.DB: An annotated data collection linking glycomics and proteomics data (2018 update). Nucleic Acids Res. 2019, 47, D1195–D1201. [Google Scholar] [CrossRef]
- Cheng, K.; Zhou, Y.; Neelamegham, S. DrawGlycan-SNFG: A robust tool to render glycans and glycopeptides with fragmentation information. Glycobiology 2017, 27, 200–205. [Google Scholar] [CrossRef]
- Kumaki, Y.; Wandersee, M.K.; Smith, A.J.; Zhou, Y.; Simmons, G.; Nelson, N.M.; Bailey, K.W.; Vest, Z.G.; Li, J.K.-K.; Kay-Sheung Chan, P.; et al. Inhibition of severe acute respiratory syndrome coronavirus replication in a lethal SARS-CoV BALB/c mouse model by stinging nettle lectin, Urtica dioica agglutinin. Antivir. Res. 2011, 90, 22–32. [Google Scholar] [CrossRef]
- Gordts, S.C.; Renders, M.; Férir, G.; Huskens, D.; Van Damme, E.J.M.; Peumans, W.J.; Balzarini, J.; Schols, D. NICTABA and UDA, two GlcNAc-binding lectins with unique antiviral activity profiles. J. Antimicrob. Chemother. 2015, 70, 1674–1685. [Google Scholar] [CrossRef] [PubMed]






































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin Family | Plant Species | Lectin | Structural Scaffold | EC50 (μg/mL) |
|---|---|---|---|---|
| GNA-related | Allium porum | APA | β-trefoil | 0.45 ± 0.08 |
| Epipactis helleborine | EHA | β-trefoil | 1.8 ± 0.3 | |
| Listera ovata | LOA | β-trefoil | 2.2 ± 1.3 | |
| Hyppeastrum hybrid | HHA | β-trefoil | 3.2 ± 2.8 | |
| Cymbidium hybrid | CA | β-trefoil | 4.9 ± 0.8 | |
| Narcissus pseudonarcissus | NPA | β-trefoil | 5.7 ± 4.4 | |
| Galanthus nivalis | GNA | β-trefoil | 6.2 ± 0.6 | |
| Allium ursinum | AUA | β-trefoil | 18 ± 4 | |
| Tulipa hybrid | TL MI | β-trefoil | 22 ± 6 | |
| Lycoris radiata | LRA | β-trefoil | 48 | |
| Colocasia esculenta | CEA | β-trefoil | >60 | |
| Allium sativum | ASA | β-trefoil | >100 | |
| JRL-related | Morus nigra | Morniga-M | β-barrel | 1.6 ± 0.5 |
| Nictaba-related | Nicotiana tabacum | Nictaba | β-sandwich | 1.7 ± 0.3 |
| Legume lectins | Cladrastis lutea | Cladrastis | β-sandwich | 7.4 ±0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barre, A.; Van Damme, E.J.M.; Simplicien, M.; Le Poder, S.; Klonjkowski, B.; Benoist, H.; Peyrade, D.; Rougé, P. Man-Specific Lectins from Plants, Fungi, Algae and Cyanobacteria, as Potential Blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) Coronaviruses: Biomedical Perspectives. Cells 2021, 10, 1619. https://doi.org/10.3390/cells10071619
Barre A, Van Damme EJM, Simplicien M, Le Poder S, Klonjkowski B, Benoist H, Peyrade D, Rougé P. Man-Specific Lectins from Plants, Fungi, Algae and Cyanobacteria, as Potential Blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) Coronaviruses: Biomedical Perspectives. Cells. 2021; 10(7):1619. https://doi.org/10.3390/cells10071619
Chicago/Turabian StyleBarre, Annick, Els J. M. Van Damme, Mathias Simplicien, Sophie Le Poder, Bernard Klonjkowski, Hervé Benoist, David Peyrade, and Pierre Rougé. 2021. "Man-Specific Lectins from Plants, Fungi, Algae and Cyanobacteria, as Potential Blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) Coronaviruses: Biomedical Perspectives" Cells 10, no. 7: 1619. https://doi.org/10.3390/cells10071619
APA StyleBarre, A., Van Damme, E. J. M., Simplicien, M., Le Poder, S., Klonjkowski, B., Benoist, H., Peyrade, D., & Rougé, P. (2021). Man-Specific Lectins from Plants, Fungi, Algae and Cyanobacteria, as Potential Blockers for SARS-CoV, MERS-CoV and SARS-CoV-2 (COVID-19) Coronaviruses: Biomedical Perspectives. Cells, 10(7), 1619. https://doi.org/10.3390/cells10071619