
KLF5 Is Activated by Gene Amplification in Gastric Cancer and Is Essential for Gastric Cell Proliferation

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Public Datasets
2.3. RNA Preparation and Real-Time PCR
2.4. siRNA Transfection and Western Blot
2.5. Cell Proliferation Assay, Clone Formation Assay
2.6. Statistical Analysis
3. Results
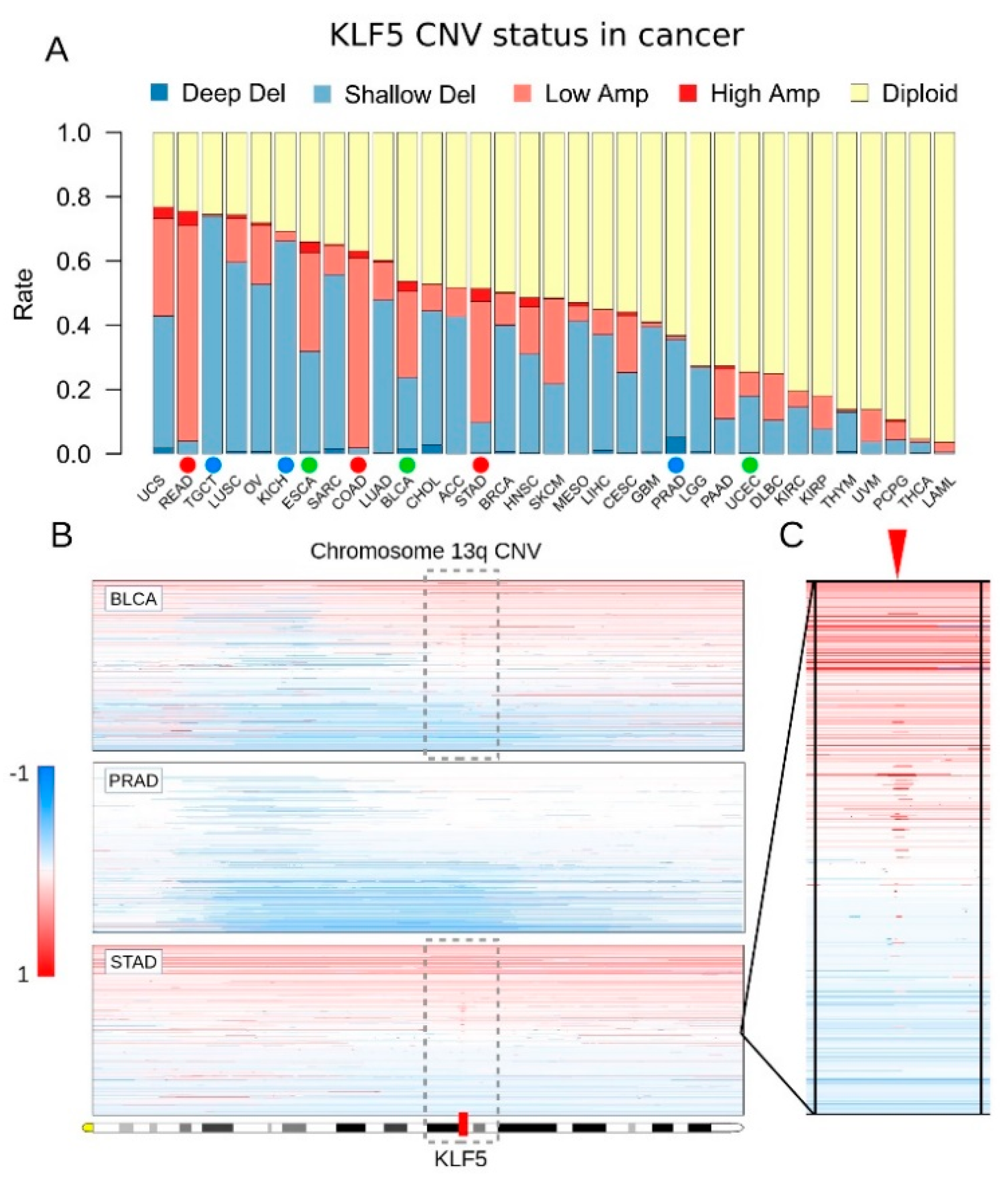
3.1. KLF5 Showed Both Gene Amplifications and Deletions in Pan-Cancer
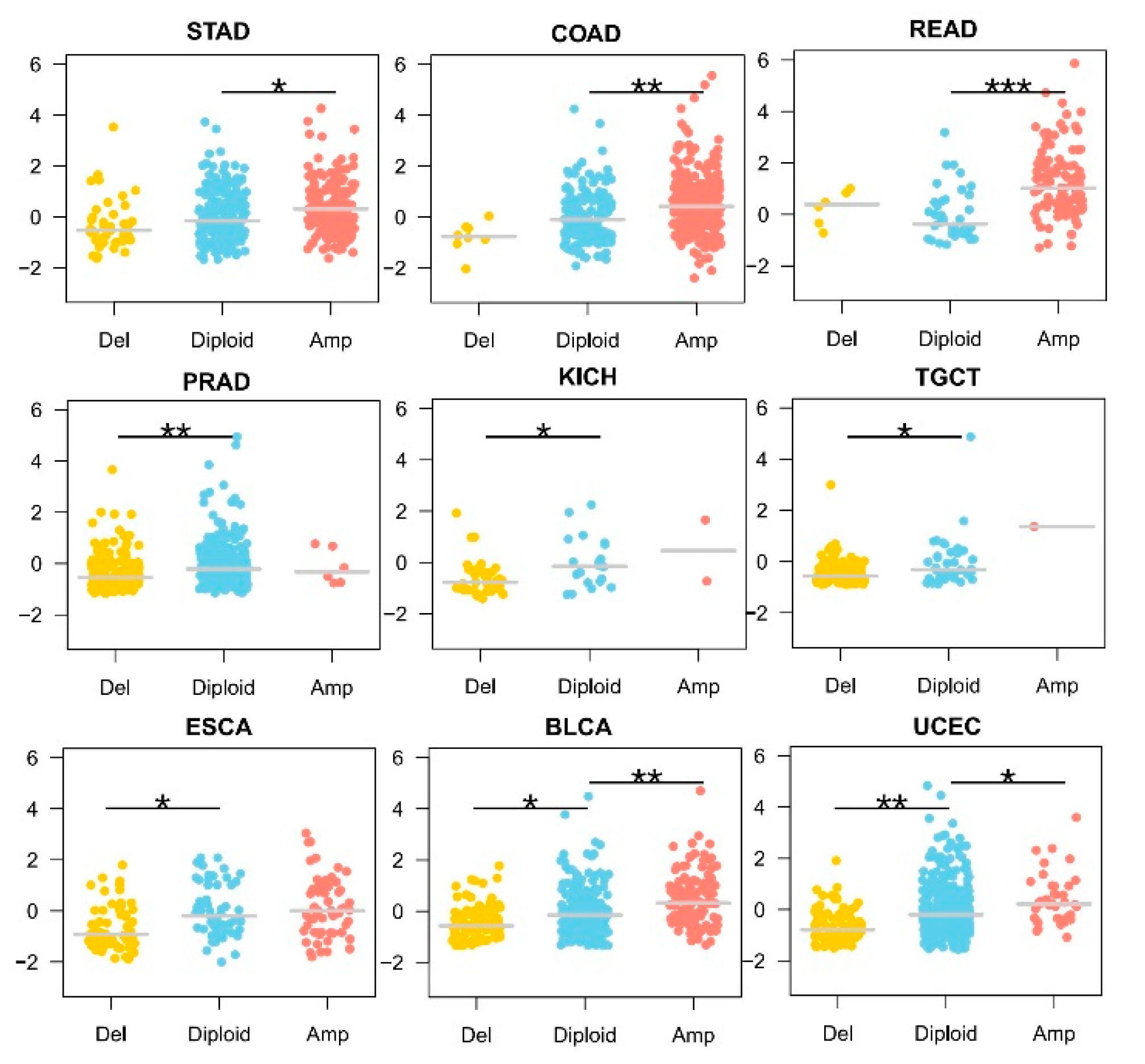
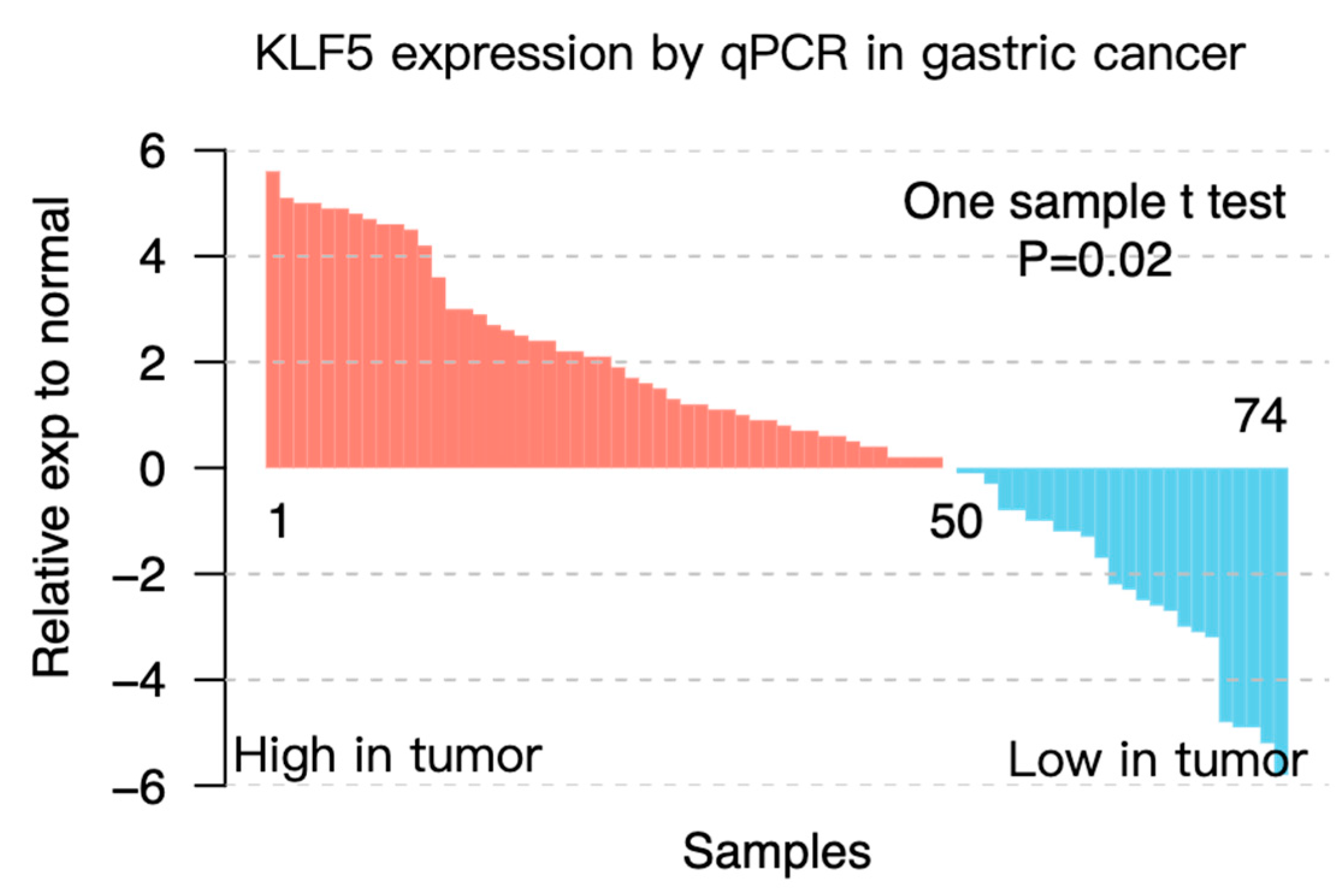
3.2. KLF5 Copy Number Showed a Dose-Response Effect on Its mRNA Expression
3.3. KLF5 Correlated with More Locally Invasive Gastric Cancer
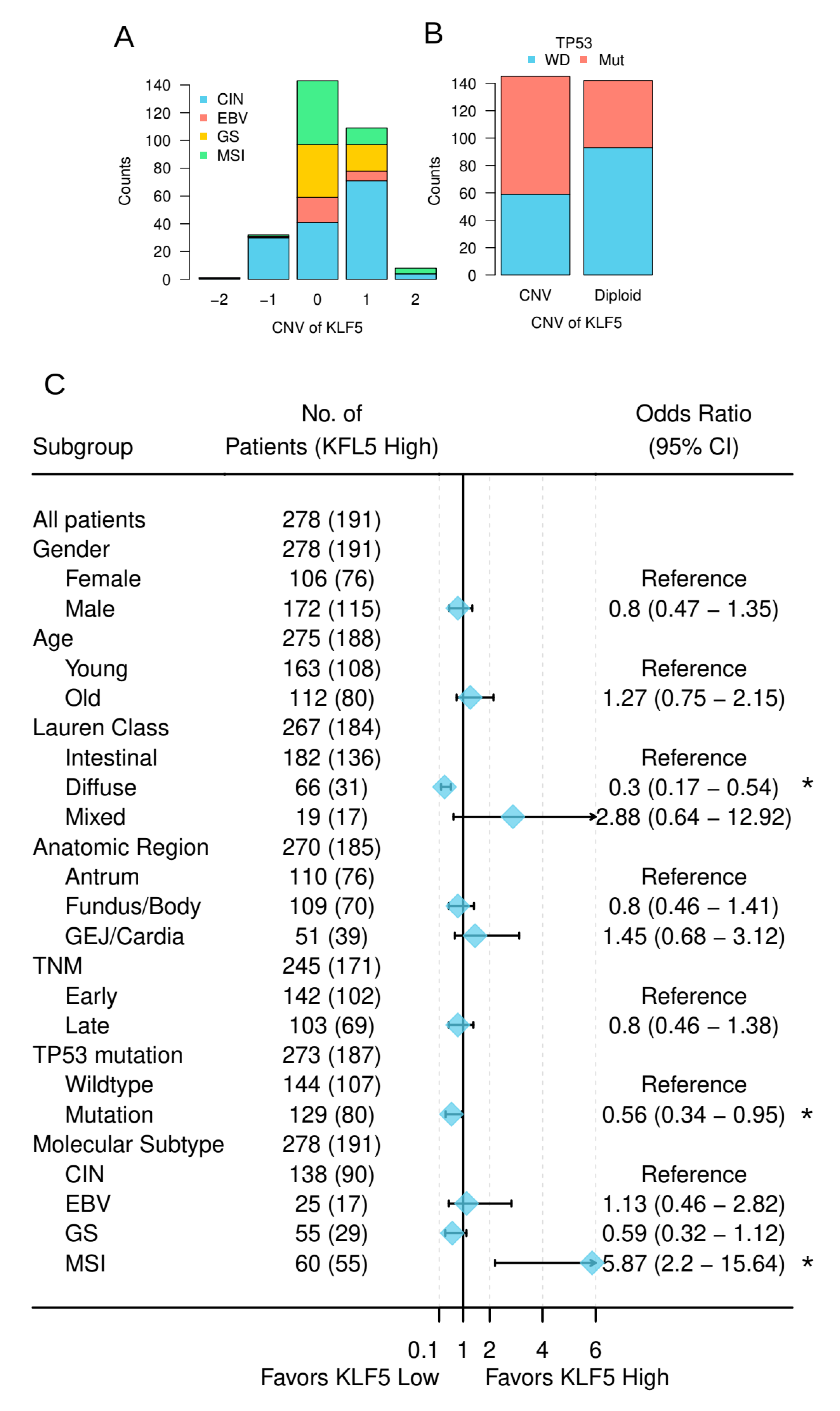
3.4. KLF5 Was Associated with the Molecular Features of Gastric Cancer
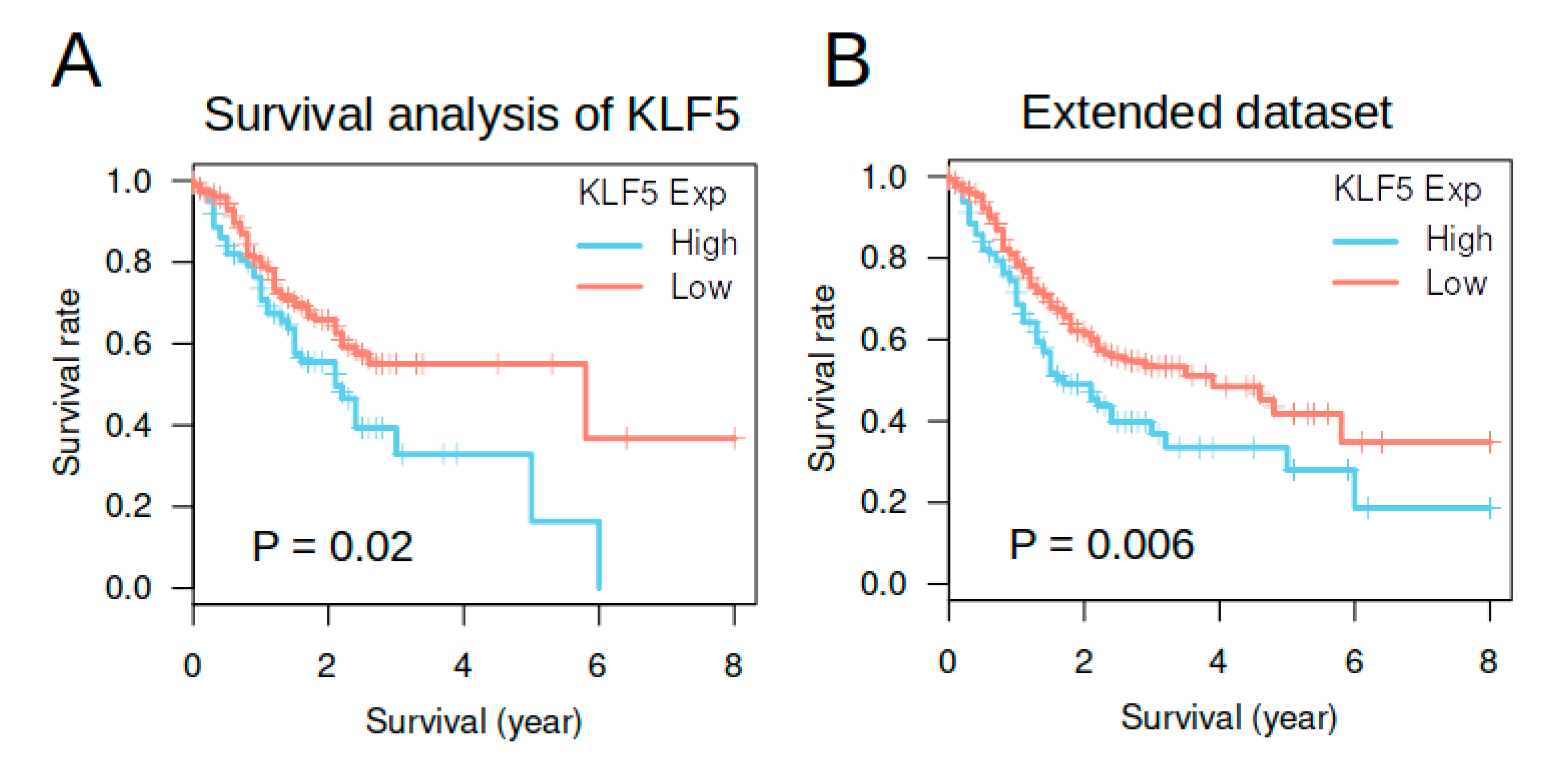
3.5. KLF5 Expression Was Not an Independent Prognosis Factor in TCGA Gastric Cancer Cohort
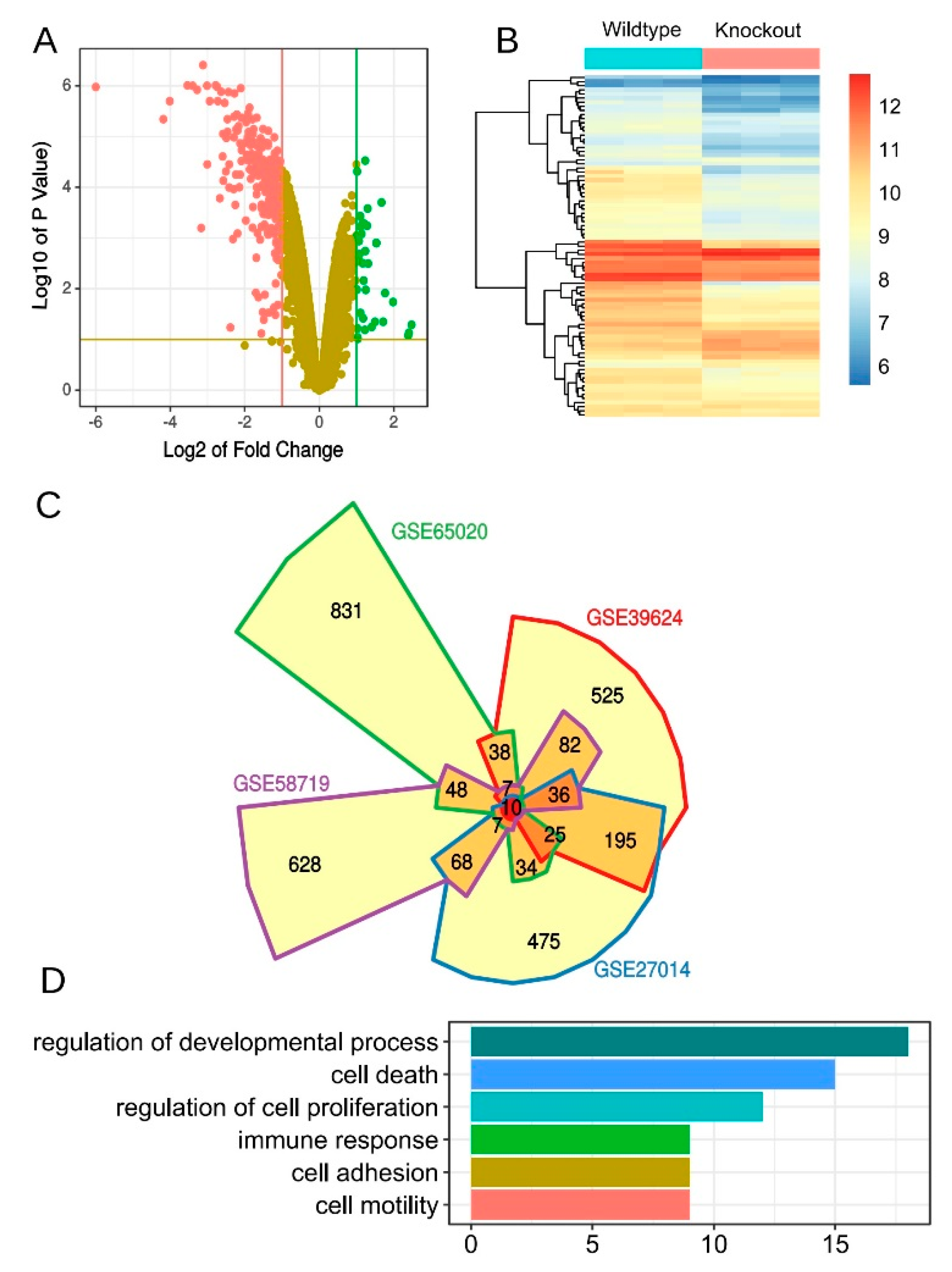
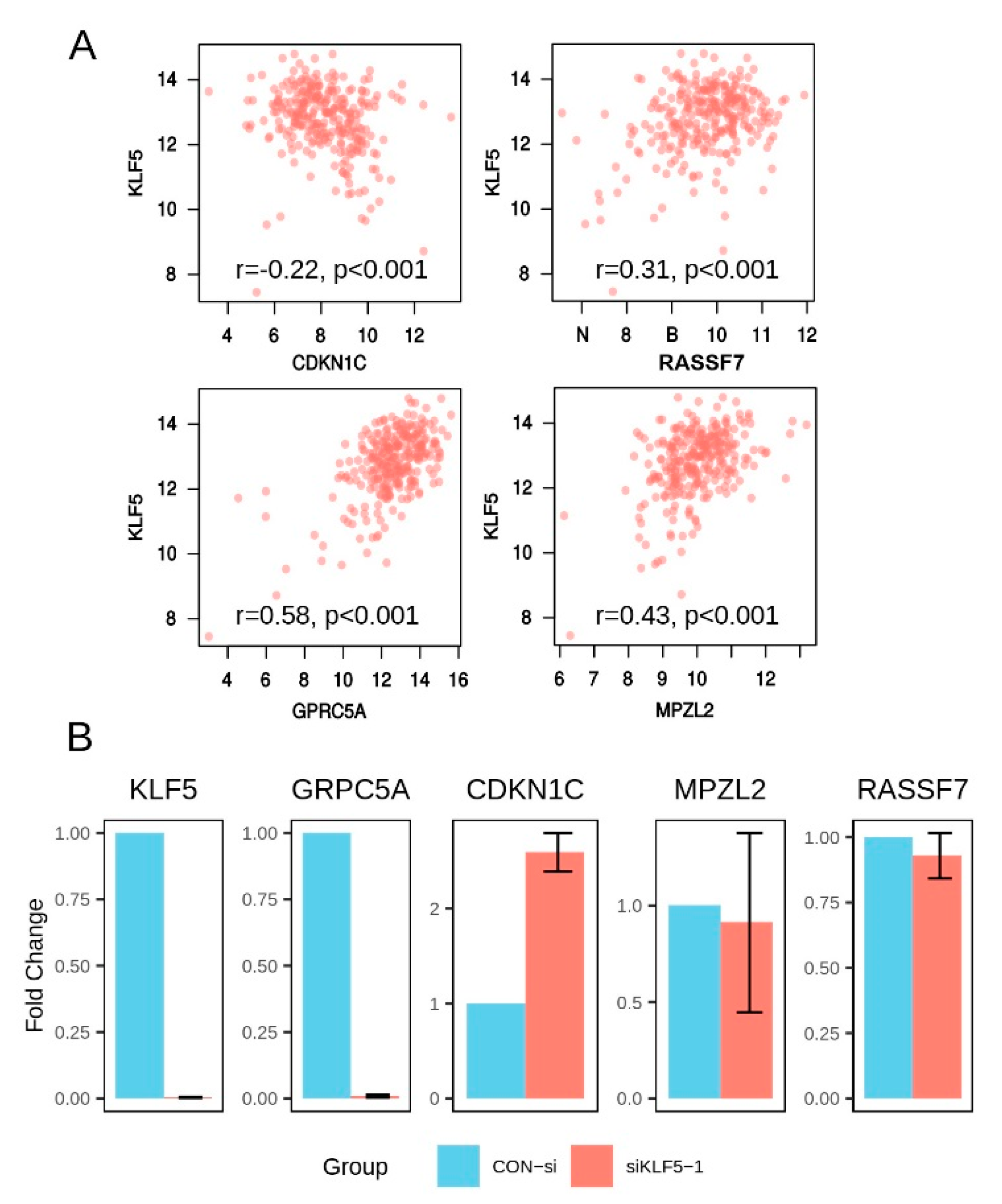
3.6. Derivation of an Aonsensus Gene Expression Signature Associated with KLF5
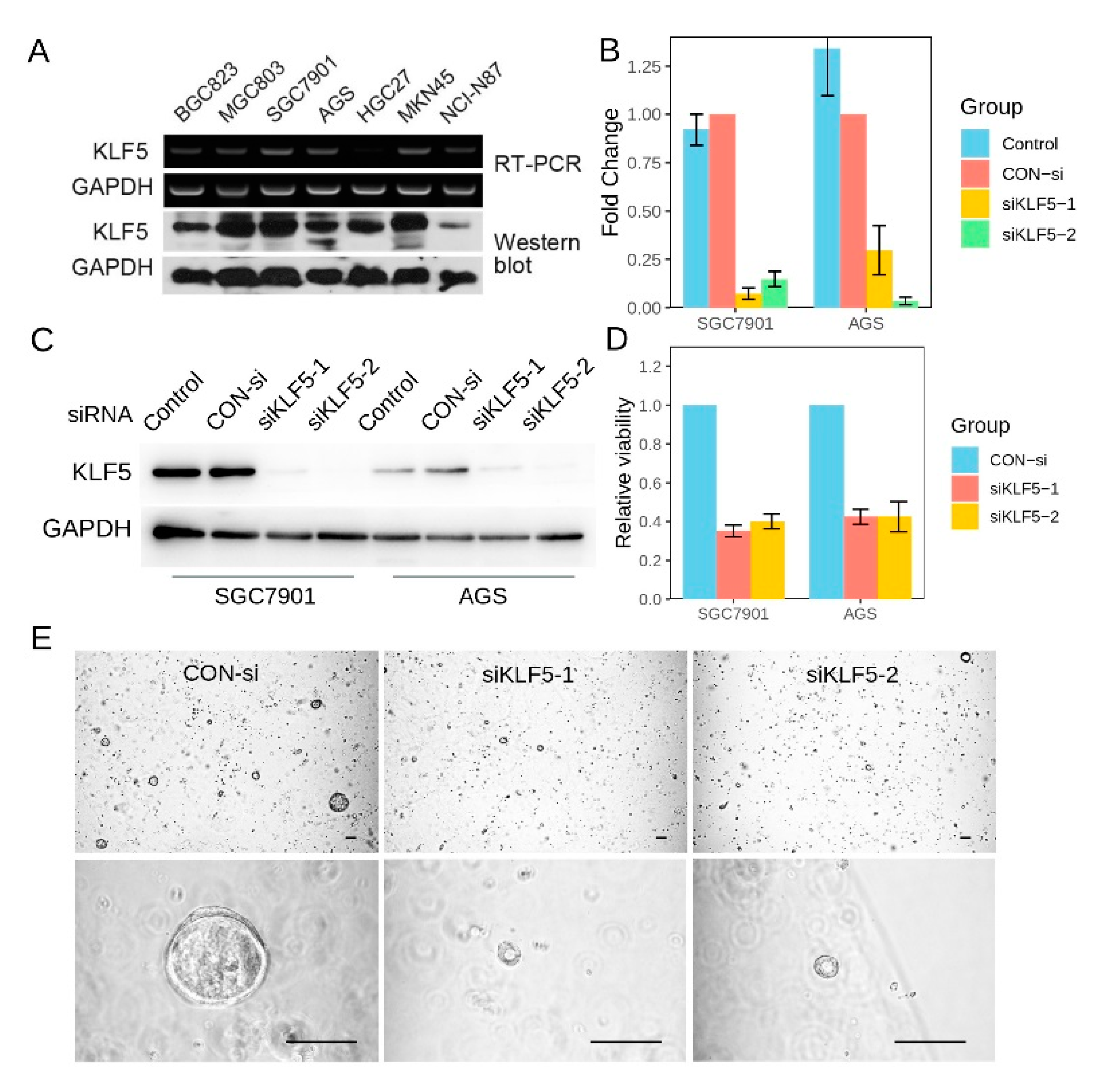
3.7. KLF5 Regulated the Proliferation of Gastric Cancer CSells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Smyth, E.C.; Cunningham, D. Targeted Therapy for Gastric Cancer. Curr. Treat. Options Oncol. 2012, 13, 377–389. [Google Scholar] [CrossRef]
- Bang, Y.-J.; Van Cutsem, E.; Feyereislova, A.; Chung, H.C.; Shen, L.; Sawaki, A.; Lordick, F.; Ohtsu, A.; Omuro, Y.; Satoh, T.; et al. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label, randomised controlled trial. Lancet 2010, 376, 687–697. [Google Scholar] [CrossRef]
- Deng, N.; Goh, L.K.; Wang, H.; Das, K.; Tao, J.; Tan, I.B.; Zhang, S.; Lee, M.; Wu, J.; Lim, K.H.; et al. A comprehensive survey of genomic alterations in gastric cancer reveals systematic patterns of molecular exclusivity and co-occurrence among distinct therapeutic targets. Gut 2012, 61, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Choi, P.S.; Francis, J.M.; Imielinski, M.; Watanabe, H.; Cherniack, A.D.; Meyerson, M. Identification of focally amplified lineage-specific super-enhancers in human epithelial cancers. Nat. Genet. 2016, 48, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Conkright, M.D.; Wani, M.A.; Anderson, K.P.; Lingrel, J.B. A gene encoding an intestinal-enriched member of the Krüppel-like factor family expressed in intestinal epithelial cells. Nucleic Acids Res. 1999, 27, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.-T.; Chen, C. Essential role of KLF5 transcription factor in cell proliferation and differentiation and its implications for human diseases. Cell. Mol. Life Sci. 2009, 66, 2691–2706. [Google Scholar] [CrossRef]
- Zhu, N.; Gu, L.; Findley, H.W.; Chen, C.; Dong, J.-T.; Yang, L.; Zhou, M. KLF5 Interacts with p53 in regulating survivin expression in acute lymphoblastic leukemia. J. Biol. Chem. 2006, 281, 14711–14718. [Google Scholar] [CrossRef]
- Kwak, M.K.; Lee, H.-J.; Hur, K.; Park, D.J.; Lee, H.S.; Kim, W.H.; Lee, K.U.; Choe, K.J.; Guilford, P.; Yang, H.-K. Expression of Krüppel-like factor 5 in human gastric carcinomas. J. Cancer Res. Clin. Oncol. 2007, 134, 163–167. [Google Scholar] [CrossRef]
- Soon, M.-S.; Hsu, L.-S.; Chen, C.-J.; Chu, P.-Y.; Liou, J.-H.; Lin, S.-H.; Hsu, J.-D.; Yeh, K.-T. Expression of Krűppel-like factor 5 in gastric cancer and its clinical correlation in Taiwan. Virchows Arch. 2011, 459, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Azami, T.; Waku, T.; Matsumoto, K.; Jeon, H.; Muratani, M.; Kawashima, A.; Yanagisawa, J.; Manabe, I.; Nagai, R.; Kunath, T.; et al. Klf5 maintains the balance of primitive endoderm versus epiblast specification during mouse embryonic development by suppression of Fgf4. Development 2017, 144, 3706–3718. [Google Scholar] [CrossRef]
- Xing, C.; Ci, X.; Sun, X.; Fu, X.; Zhang, Z.; Dong, E.N.; Hao, Z.-Z.; Dong, J.-T. Klf5 deletion promotes Pten deletion-initiated luminal-type mouse prostate tumors through multiple oncogenic signaling pathways. Neoplasia 2014, 16, 883–899. [Google Scholar] [CrossRef]
- Bell, S.M.; Zhang, L.; Mendell, A.; Xu, Y.; Haitchi, H.M.; Lessard, J.L.; Whitsett, J.A. Kruppel-like factor 5 is required for formation and differentiation of the bladder urothelium. Dev. Biol. 2011, 358, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.-Q.; Zhou, Z.; Huang, J.; Chaudhury, L.; Dong, J.-T.; Chen, C. Krüppel-like factor 5 promotes breast cell proliferation partially through upregulating the transcription of fibroblast growth factor binding protein 1. Oncogene 2009, 28, 3702–3713. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Chen, W.; Wei, Z.W.; Chu, W.W.; Lu, X.F.; Li, B.; Chen, H.; Meng, S.J.; Hao, T.F.; Wei, J.T.; et al. The anti-tumor effect of nab-paclitaxel proven by patient-perived organoids. Onco Targets Ther. 2020, 13, 6017–6025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Choi, P.S.; Francis, J.M.; Gao, G.F.; Campbell, J.D.; Ramachandran, A.; Mitsuishi, Y.; Ha, G.; Shih, J.; Vazquez, F.; et al. Somatic Superenhancer Duplications and Hotspot Mutations Lead to Oncogenic Activation of the KLF5 Transcription Factor. Cancer Discov. 2018, 8, 108–125. [Google Scholar] [CrossRef]
- Matsuoka, S.; Edwards, M.C.; Bai, C.; Parker, S.; Zhang, P.; Baldini, A.; Harper, J.W.; Elledge, S.J. p57KIP2, a structurally distinct member of the p21CIP1 Cdk inhibitor family, is a candidate tumor suppressor gene. Genes Dev. 1995, 9, 650–662. [Google Scholar] [CrossRef]
- Weitzel, J.N.; Kasperczyk, A.; Mohan, C.; Krontiris, T.G. The HRAS1 gene cluster: Two upstream regions recognizing transcripts and a third encoding a gene with a leucine zipper domain. Genomics 1992, 14, 309–319. [Google Scholar] [CrossRef]
- Takahashi, S.; Ebihara, A.; Kajiho, H.; Kontani, K.; Nishina, H.; Katada, T. RASSF7 negatively regulates pro-apoptotic JNK signaling by inhibiting the activity of phosphorylated-MKK7. Cell Death Differ. 2011, 18, 645–655. [Google Scholar] [CrossRef]
- Sherwood, V.; Manbodh, R.; Sheppard, C.; Chalmers, A.D. RASSF7 is a member of a new family of RAS association domain-containing proteins and is required for completing mitosis. Mol. Biol. Cell 2008, 19, 1772–1782. [Google Scholar] [CrossRef]
- Cheng, Y.; Lotan, R. Molecular cloning and characterization of a novel retinoic acid-inducible gene that encodes a putative G protein-coupled receptor. J. Biol. Chem. 1998, 273, 35008–35015. [Google Scholar] [CrossRef]
- Tao, Q.; Fujimoto, J.; Men, T.; Ye, X.; Deng, J.; Lacroix, L.; Clifford, J.L.; Mao, L.; Van Pelt, C.S.; Lee, J.J.; et al. Identification of the retinoic acid-inducible Gprc5a as a new lung tumor suppressor gene. J. Natl. Cancer Inst. 2007, 99, 1668–1682. [Google Scholar] [CrossRef] [PubMed]
- Guttinger, M.; Sutti, F.; Panigada, M.; Porcellini, S.; Merati, B.; Mariani, M.; Teesalu, T.; Consalez, G.G.; Grassi, F. Epithelial V-like antigen (EVA), a novel member of the immunoglobulin superfamily, expressed in embryonic epithelia with a potential role as homotypic adhesion molecule in thymus histogenesis. J. Cell Biol. 1998, 141, 1061–1071. [Google Scholar] [CrossRef]
- Sun, R.; Chen, X.; Yang, V.W. Intestinal-enriched Krüppel-like Factor (Krüppel-like Factor 5) Is a Positive Regulator of Cellular Proliferation. J. Biol. Chem. 2001, 276, 6897–6900. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Benjamin, M.S.; Sun, X.; Otto, K.B.; Guo, P.; Dong, X.-Y.; Bao, Y.; Zhou, Z.; Cheng, X.; Simons, J.W.; et al. KLF5 promotes cell proliferation and tumorigenesis through gene regulation and the TSU-Pr1 human bladder cancer cell line. Int. J. Cancer 2006, 118, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.E.; Hasenstein, J.R.; Baktula, A.; Velu, C.S.; Xu, Y.; Wan, H.; Whitsett, J.A.; Gilks, C.B.; Grimes, H.L. Krüppel-Like Factor 5 Is Not Required for K-RasG12D Lung Tumorigenesis, but Represses ABCG2 Expression and Is Associated with Better Disease-Specific Survival. Am. J. Pathol. 2010, 177, 1503–1513. [Google Scholar] [CrossRef]
- Tong, D.; Czerwenka, K.; Heinze, G.; Ryffel, M.; Schuster, E.; Witt, A.; Leodolter, S.; Zeillinger, R. Expression of KLF5 is a Prognostic Factor for Disease-Free Survival and Overall Survival in Patients with Breast Cancer. Clin. Cancer Res. 2006, 12, 2442–2448. [Google Scholar] [CrossRef]
- Chen, C.; Bhalala, H.V.; Qiao, H.; Dong, J.-T. A possible tumor suppressor role of the KLF5 transcription factor in human breast cancer. Oncogene 2002, 21, 6567–6572. [Google Scholar] [CrossRef]
- Chen, C.; Bhalala, H.V.; Vessella, R.L.; Dong, J.-T. KLF5 is frequently deleted and down-regulated but rarely mutated in prostate cancer. Prostate 2003, 55, 81–88. [Google Scholar] [CrossRef]
- Gao, Y.; Ding, Y.; Chen, H.; Chen, H.; Zhou, J. Targeting Krüppel-like factor 5 (KLF5) for cancer therapy. Curr. Top. Med. Chem. 2015, 15, 699–713. [Google Scholar] [CrossRef]
- Goldstein, B.G.; Chao, H.-H.; Yang, Y.; Yermolina, Y.A.; Tobias, J.W.; Katz, J.P. Overexpression of Kruppel-like factor 5 in esophageal epithelia in vivo leads to increased proliferation in basal but not suprabasal cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1784–G1792. [Google Scholar] [CrossRef]
- McConnell, B.B.; Klapproth, J.-M.A.; Sasaki, M.; Nandan, M.O.; Yang, V.W. Krüppel-like factor 5 mediates transmissible murine colonic hyperplasia caused by Citrobacter rodentium infection. Gastroenterology 2008, 134, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Nandan, M.O.; Ghaleb, A.M.; McConnell, B.B.; Patel, N.V.; Robine, S.; Yang, V.W. Krüppel-like factor 5 is a crucial mediator of intestinal tumorigenesis in mice harboring combined ApcMin and KRASV12 mutations. Mol. Cancer 2010, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Chia, N.-Y.; Deng, N.; Das, K.; Huang, D.; Hu, L.; Zhu, Y.; Lim, K.H.; Lee, M.-H.; Wu, J.; Sam, X.X.; et al. Regulatory crosstalk between lineage-survival oncogenes KLF5, GATA4 and GATA6 cooperatively promotes gastric cancer development. Gut 2015, 64, 707–719. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | KLF5 Expression (Low) | KLF5 Expression (High) | P | |
|---|---|---|---|---|
| Cases | 74 | 24 | 50 | |
| Sex | 0.296 | |||
| Male | 55 (74.3%) | 16 (66.7%) | 39 (78.0%) | |
| Female | 19 (25.7%) | 8 (33.3%) | 11 (22.0%) | |
| Age (year) | 0.909 | |||
| ≤55 | 24 (32.4%) | 8 (33.3%) | 16 (32.0%) | |
| >55 | 50 (67.6%) | 16 (66.7%) | 34 (68.0%) | |
| Length (cm) | 0.008 ** | |||
| ≤5 | 39 (52.7%) | 18 (75.0%) | 21 (42.0%) | |
| >5 | 35 (47.3%) | 6 (25.0%) | 29 (58.0%) | |
| Serosa Invasion | 0.054 | |||
| Negative | 58 (78.4%) | 22 (91.7%) | 36 (72.0%) | |
| Positive | 16 (21.6%) | 2 (8.3%) | 14 (28.0%) | |
| Primary Tumor (T) | 0.029 * | |||
| T1/T2 | 19 (25.7%) | 10 (41.7%) | 9 (18.0%) | |
| T3/T4 | 55 (74.3%) | 14 (58.3%) | 41 (82.0%) | |
| Regional Lymph Nodes (N) | 0.619 | |||
| N0/N1 | 37 (50.0%) | 13 (54.2%) | 24 (48.0%) | |
| N2/N3 | 37 (50.0%) | 11 (45.8%) | 26 (52.0%) | |
| Distant Metastasis (M) | 0.329 | |||
| M0 | 60 (81.1%) | 21 (87.5%) | 39 (78.0%) | |
| M1 | 14 (18.9%) | 3 (12.5%) | 11 (22.0%) | |
| TNM | 0.189 | |||
| Ⅰ/Ⅱ | 32 (43.2%) | 13 (54.2%) | 19 (38.0%) | |
| Ⅲ/Ⅳ | 42 (56.8%) | 11 (45.8%) | 31 (62.0%) | |
| Histological Grade (G) | 0.762 | |||
| Well/Moderate | 29 (39.2%) | 10 (41.7%) | 19 (38.0%) | |
| Poor/Undifferentiated | 45 (60.8%) | 14 (58.3%) | 31 (62.0%) | |
| Univariate | Multivariate | |||
|---|---|---|---|---|
| HR (95% CI) | P | HR (95% CI) | P | |
| KLF5 expression (high vs. low) | 0.62 (0.41, 0.94) | 0.02 * | 0.72 (0.43, 1.22) | 0.22 |
| TNM stage (late vs. early) | 2.24 (1.44, 3.5) | 0.0004 *** | 1.8 (1.12, 2.89) | 0.02 * |
| Molecular subtype (EBV vs. CIN) | 0.85 (0.4, 1.8) | 0.67 | 0.91 (0.38, 2.14) | 0.82 |
| Molecular subtype (GS vs. CIN) | 1.11 (0.68, 1.83) | 0.67 | 0.87 (0.45, 1.69) | 0.69 |
| Molecular subtype (MSI vs. CIN) | 0.74 (0.43, 1.27) | 0.27 | 0.69 (0.34, 1.4) | 0.3 |
| Lauren (Diffused vs. Intestinal) | 1.61 (1.02, 2.55) | 0.04 * | 1.29 (0.68, 2.47) | 0.44 |
| Lauren (Mixed vs. Intestinal) | 2.5 (1.33, 4.7) | 0.004 ** | 2.37 (1.14, 4.92) | 0.02 * |
| Gene | Correlation | p-Value |
|---|---|---|
| AIM1L | 0.41 | 1.57 × 10−12 |
| CDKN1C | −0.22 | 1.64 × 10−4 |
| CMBL | 0.17 | 5.84 × 10−3 |
| DGAT2 | 0.10 | 1.02 × 10−1 |
| GPRC5A | 0.58 | 3.02 × 10−26 |
| GPX2 | 0.20 | 1.05 × 10−3 |
| JAKMIP1 | −0.10 | 9.61 × 10−2 |
| KRT7 | 0.11 | 7.44 × 10−2 |
| MPZL2 | 0.43 | 3.80 × 10−14 |
| PGLYRP1 | −0.02 | 7.85 × 10−1 |
| PRR13 | 0.29 | 1.28 × 10−6 |
| RASSF7 | 0.31 | 2.08 × 10−7 |
| SH2D4A | 0.48 | 2.27 × 10−17 |
| SH3BGRL2 | 0.42 | 1.95 × 10−13 |
| SLC25A10 | 0.30 | 5.58 × 10−7 |
| TINAGL1 | 0.06 | 2.92 × 10−1 |
| TJP3 | 0.51 | 2.31 × 10−19 |
| TSPAN8 | 0.44 | 1.58 × 10−14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Zhang, J.; Fu, H.; Hou, X.; Su, Q.; He, Y.; Yang, D. KLF5 Is Activated by Gene Amplification in Gastric Cancer and Is Essential for Gastric Cell Proliferation. Cells 2021, 10, 1002. https://doi.org/10.3390/cells10051002
Chen W, Zhang J, Fu H, Hou X, Su Q, He Y, Yang D. KLF5 Is Activated by Gene Amplification in Gastric Cancer and Is Essential for Gastric Cell Proliferation. Cells. 2021; 10(5):1002. https://doi.org/10.3390/cells10051002
Chicago/Turabian StyleChen, Wei, Jian Zhang, Huafeng Fu, Xun Hou, Qiao Su, Yulong He, and Dongjie Yang. 2021. "KLF5 Is Activated by Gene Amplification in Gastric Cancer and Is Essential for Gastric Cell Proliferation" Cells 10, no. 5: 1002. https://doi.org/10.3390/cells10051002
APA StyleChen, W., Zhang, J., Fu, H., Hou, X., Su, Q., He, Y., & Yang, D. (2021). KLF5 Is Activated by Gene Amplification in Gastric Cancer and Is Essential for Gastric Cell Proliferation. Cells, 10(5), 1002. https://doi.org/10.3390/cells10051002