
Analysis of Circulating microRNA Signatures and Preeclampsia Development



Abstract
1. Introduction
2. Materials and Methods
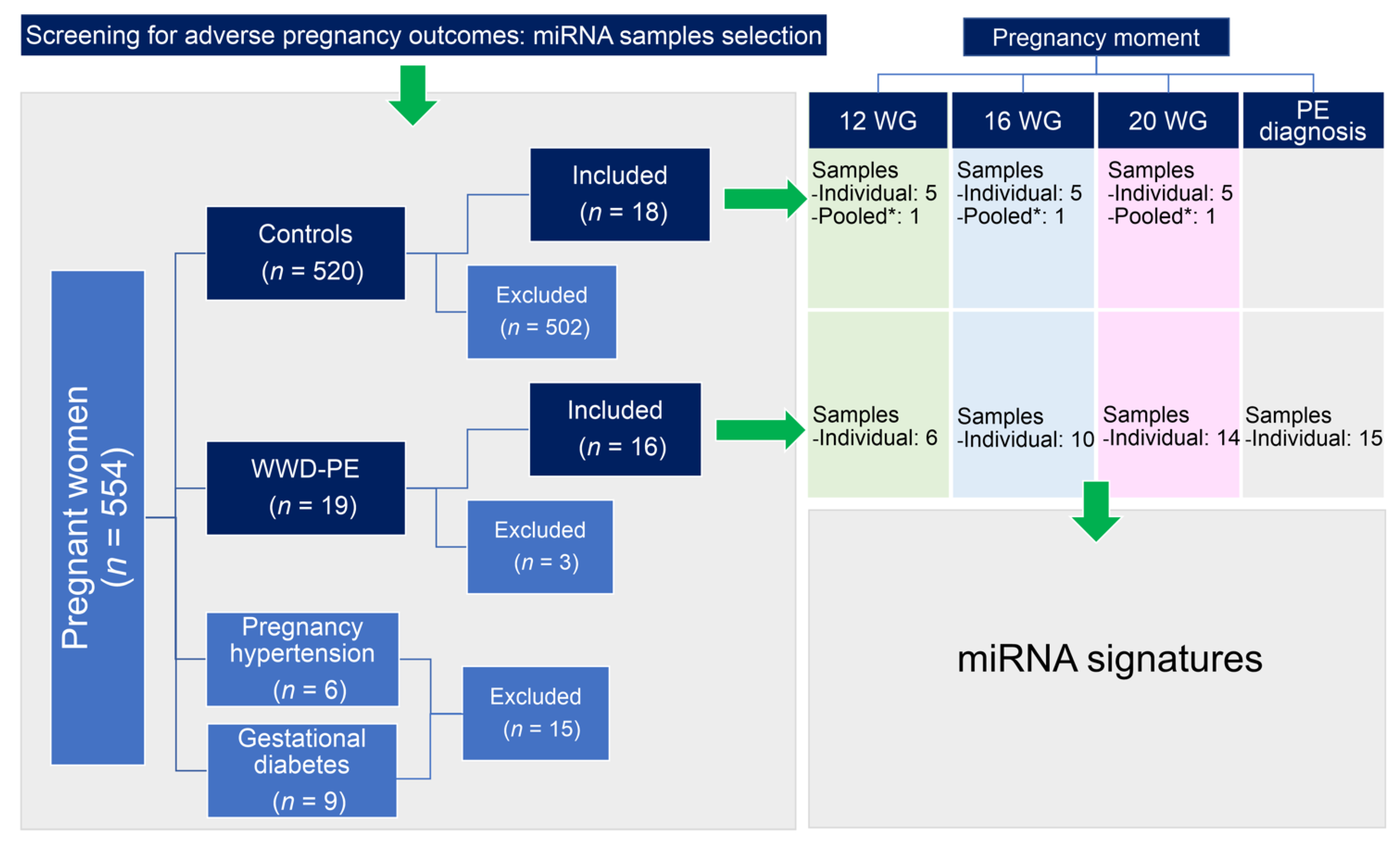
2.1. Patients and Study Design
2.2. Biological Samples
2.3. Maternal Serum miRNA Profiling
2.4. Data Analysis
3. Results
3.1. General Characteristics of the Study Participants
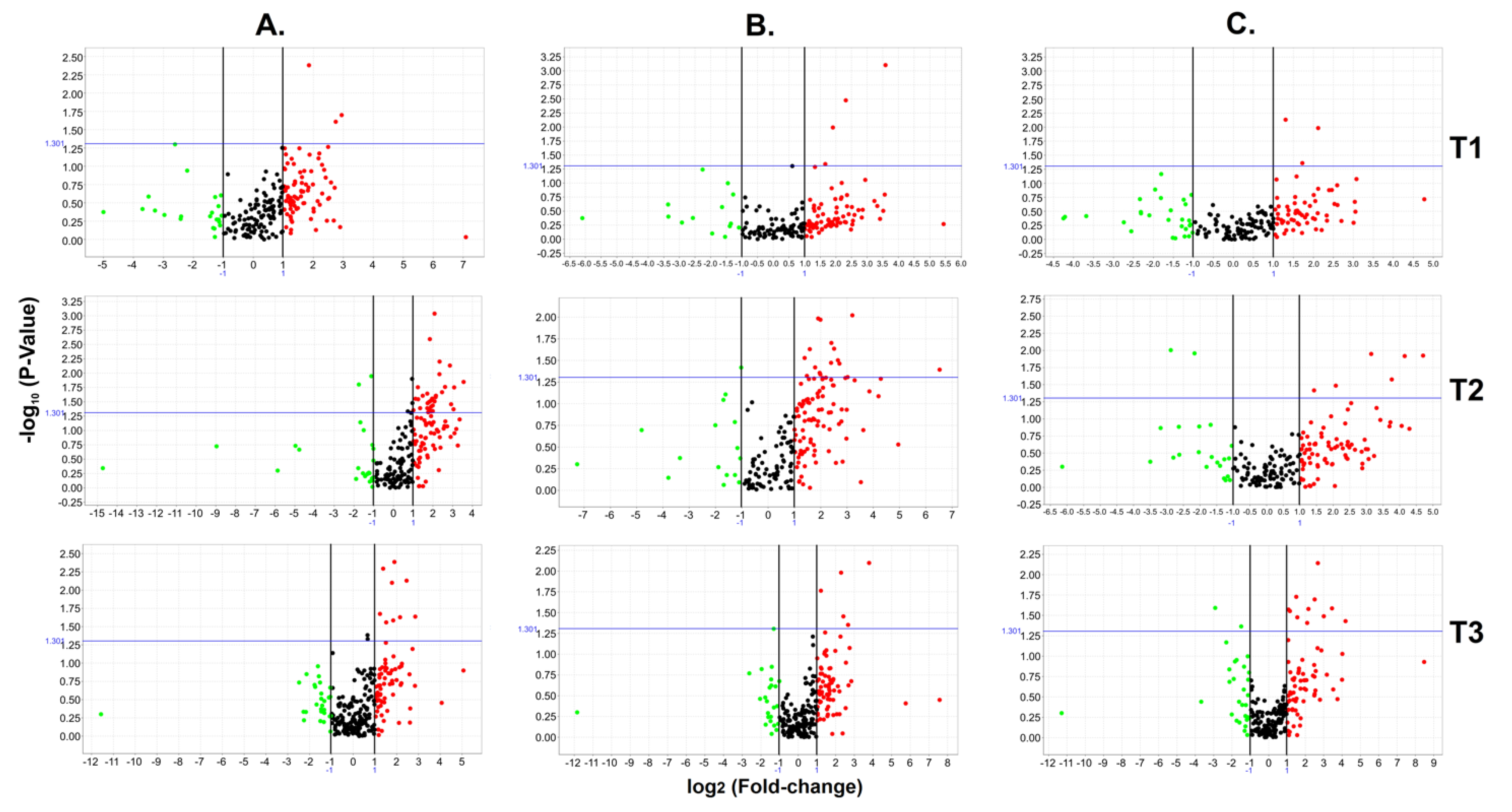
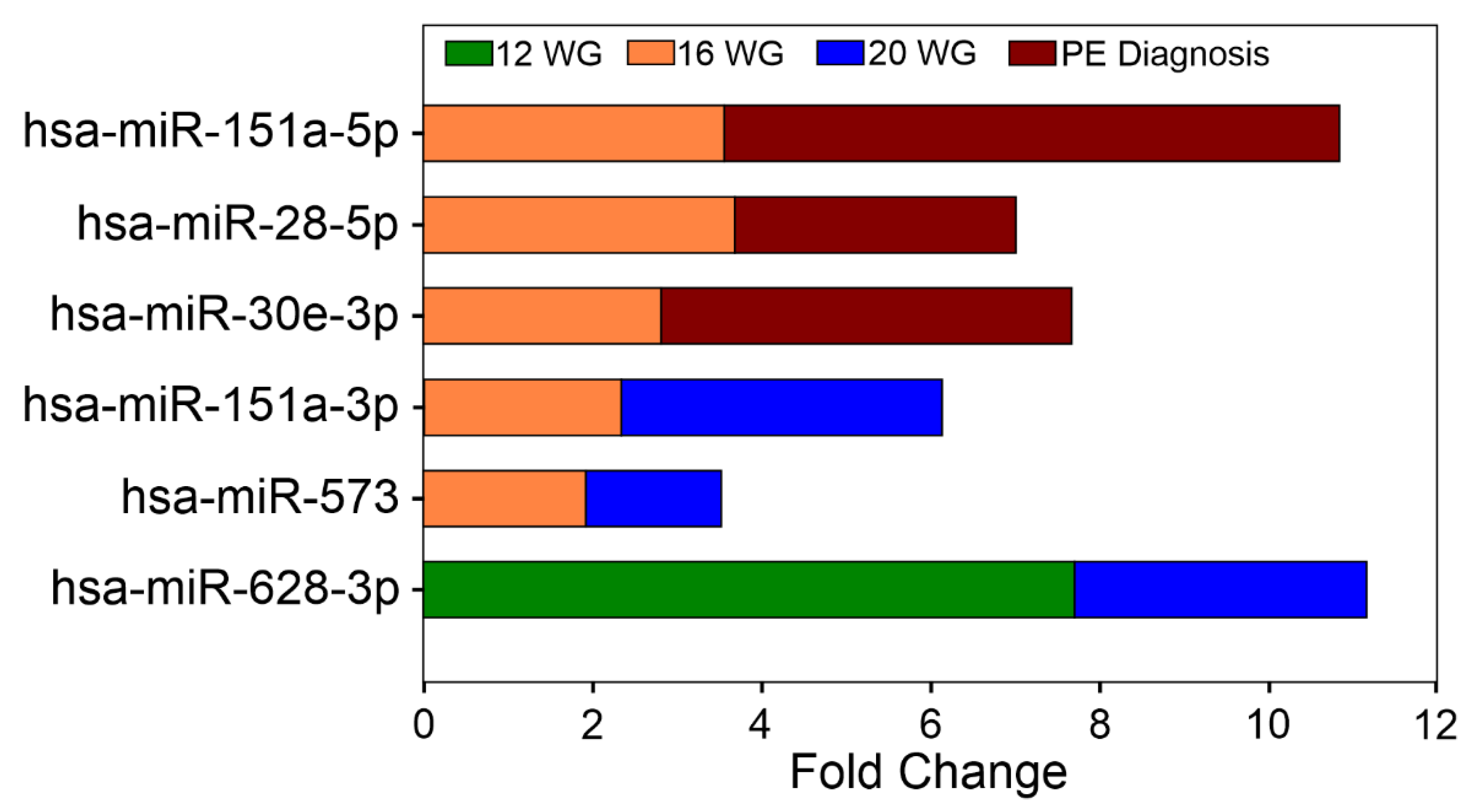
3.2. Early Circulating miRNA Expression Profiles and PE Development
Circulating miRNA Expression Signatures and PE Severity
3.3. Signaling Pathways Targeted by miRNA Clusters and the Biological Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aslam, M.I. MicroRNAs are novel biomarkers of colorectal cancer. Br. J. Surg. 2009, 96, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Maccani, M.A. Epigenetics in the placenta. Am. J. Reprod. Immunol. 2009, 62, 78–89. [Google Scholar] [CrossRef]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Brkic, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.L. Placental trophoblast cell differentiation: Physiological regulation and pathological relevance to preeclampsia. Mol. Asp. Med. 2013, 34, 981–1023. [Google Scholar] [CrossRef] [PubMed]
- Enquobahrie, D.A.; Abetew, D.F.; Sorensen, T.K.; Willoughby, D.; Chidambaram, K.; Williams, M.A. Placental microRNA expression in pregnancies complicated by preeclampsia. Am. J. Obs. Gynecol. 2011, 204, 178.e112–178.e121. [Google Scholar] [CrossRef]
- Fu, G.; Brkic, J.; Hayder, H.; Peng, C. MicroRNAs in Human Placental Development and Pregnancy Complications. Int. J. Mol. Sci. 2013, 14, 5519–5544. [Google Scholar] [CrossRef]
- Pineles, B.L.; Romero, R.; Montenegro, D.; Tarca, A.L.; Han, Y.M.; Kim, Y.M.; Draghici, S.; Espinoza, J.; Kusanovic, J.P.; Mittal, P.; et al. Distinct subsets of microRNAs are expressed differentially in the human placentas of patients with preeclampsia. Am. J. Obs. Gynecol. 2007, 196, 261.e1–261.e6. [Google Scholar] [CrossRef]
- Chen, D.B.; Wang, W. Human placental microRNAs and preeclampsia. Biol. Reprod. 2013, 88, 130. [Google Scholar] [CrossRef]
- Doridot, L.; Miralles, F.; Barbaux, S.; Vaiman, D. Trophoblasts, invasion, and microRNA. Front. Genet. 2013, 4, 248. [Google Scholar] [CrossRef]
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists. Hypertension in pregnancy. Report of the American College of Obstetricians and Gynecologists’ task force on hypertension in pregnancy. Obs. Gynecol. 2013, 122, 1122–1131. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Hernandez-Delgadillo, G.P.; Flores-Morales, V.; Cardenas-Vargas, E.; Mercado-Reyes, M.; Rodriguez-Sanchez, I.P.; Delgado-Enciso, I.; Galvan-Tejada, C.E.; Galvan-Tejada, J.I.; Celaya-Padilla, J.M.; et al. Current model systems for the study of preeclampsia. Exp. Biol. Med. 2018, 243, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Luque, A.; Farwati, A.; Crovetto, F.; Crispi, F.; Figueras, F.; Gratacos, E.; Aran, J.M. Usefulness of circulating microRNAs for the prediction of early preeclampsia at first-trimester of pregnancy. Sci. Rep. 2014, 4, 4882. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zhao, Y.; Liu, M.; Wang, Y.; Wang, H.; Li, Y.X.; Zhu, X.; Yao, Y.; Qiao, J.; Ji, L.; et al. Variations of microRNAs in human placentas and plasma from preeclamptic pregnancy. Hypertension 2014, 63, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ge, Q.; Guo, L.; Lu, Z. Maternal plasma miRNAs expression in preeclamptic pregnancies. BioMed Res. Int. 2013, 2013, 970265. [Google Scholar] [CrossRef]
- Yang, Q.; Lu, J.; Wang, S.; Li, H.; Ge, Q.; Lu, Z. Application of next-generation sequencing technology to profile the circulating microRNAs in the serum of preeclampsia versus normal pregnant women. Clin. Chim. Acta Int. J. Clin. Chem. 2011, 412, 2167–2173. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, H.; Lin, H.; Qi, J.; Zhu, C.; Gao, Z.; Wang, H. Circulating microRNAs are elevated in plasma from severe preeclamptic pregnancies. Reproduction 2012, 143, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Ura, B.; Feriotto, G.; Monasta, L.; Bilel, S.; Zweyer, M.; Celeghini, C. Potential role of circulating microRNAs as early markers of preeclampsia. Taiwan. J. Obstet. Gynecol. 2014, 53, 232–234. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Garza-Veloz, I.; Gutierrez-Arteaga, C.; Delgado-Enciso, I.; Barbosa-Cisneros, O.Y.; Flores-Morales, V.; Hernandez-Delgadillo, G.P.; Rocha-Pizana, M.R.; Rodriguez-Sanchez, I.P.; Badillo-Almaraz, J.I.; et al. Circulating levels of specific members of chromosome 19 microRNA cluster are associated with preeclampsia development. Arch. Gynecol. Obstet. 2018, 297, 365–371. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Garza-Veloz, I.; Castruita-Dela Rosa, C.; Ortiz-Castro, Y.; Aceves-Medina, M.C.; Vazquez-Castro, R.; Delgado-Enciso, I.; Castaneda-Lopez, M.E. Plasma cancer biomarker multiplex screening and the risk of subsequent preeclampsia. Int. J. Cardiol. 2015, 179, 58–60. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Castruita-De La Rosa, C.; Garza-Veloz, I.; Cardiel-Hernandez, R.M.; Espinoza-Juarez, M.A.; Delgado-Enciso, I.; Castaneda-Lopez, M.E.; Cardenas-Vargas, E.; Trejo-Vazquez, F.; Sotelo-Ham, E.I.; et al. Early pregnancy protein multiplex screening reflects circulating and urinary divergences associated with the development of preeclampsia. Hypertens. Pregnancy 2018, 37, 37–50. [Google Scholar] [CrossRef]
- Brown, M.A.; Lindheimer, M.D.; de Swiet, M.; Van Assche, A.; Moutquin, J.M. The classification and diagnosis of the hypertensive disorders of pregnancy: Statement from the International Society for the Study of Hypertension in Pregnancy (ISSHP). Hypertens. Pregnancy 2001, 20, IX–XIV. [Google Scholar] [CrossRef]
- Garza-Veloz, I.; Castruita-De la Rosa, C.; Cortes-Flores, R.; Martinez-Gaytan, V.; Rivera-Munoz, J.E.; Garcia-Mayorga, E.A.; Meza-Lamas, E.; Rojas-Martinez, A.; Ortiz-Lopez, R.; Martinez-Fierro, M.L. No association between polymorphisms/haplotypes of the vascular endothelial growth factor gene and preeclampsia. BMC Pregnancy Childbirth 2011, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fierro, M.L.; Garza-Veloz, I.; Carrillo-Sanchez, K.; Martinez-Gaytan, V.; Cortes-Flores, R.; Ochoa-Torres, M.A.; Guerrero, G.G.; Rodriguez-Sanchez, I.P.; Cancela-Murrieta, C.O.; Zamudio-Osuna, M.; et al. Expression levels of seven candidate genes in human peripheral blood mononuclear cells and their association with preeclampsia. Hypertens Pregnancy 2014, 33, 191–203. [Google Scholar] [CrossRef]
- von Dadelszen, P.; Magee, L.A.; Roberts, J.M. Subclassification of preeclampsia. Hypertens Pregnancy 2003, 22, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Dawson, A.; Lohsoonthorn, V.; Williams, M.A. Risk Factors of Early and Late Onset Preeclampsia among Thai Women. Asian Biomed 2009, 3, 477–486. [Google Scholar]
- Louwen, F.; Muschol-Steinmetz, C.; Reinhard, J.; Reitter, A.; Yuan, J. A lesson for cancer research: Placental microarray gene analysis in preeclampsia. Oncotarget 2012, 3, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Moslehi, R.; Mills, J.L.; Signore, C.; Kumar, A.; Ambroggio, X.; Dzutsev, A. Integrative transcriptome analysis reveals dysregulation of canonical cancer molecular pathways in placenta leading to preeclampsia. Sci. Rep. 2013, 3, 2407. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, C.; Bruni, L.; Dangles-Marie, V.; Pecking, A.P.; Bellet, D. Molecular circuits shared by placental and cancer cells, and their implications in the proliferative, invasive and migratory capacities of trophoblasts. Hum. Reprod. Update 2007, 13, 121–141. [Google Scholar] [CrossRef]
- Huang, Y.S.; Dai, Y.; Yu, X.F.; Bao, S.Y.; Yin, Y.B.; Tang, M.; Hu, C.X. Microarray analysis of microRNA expression in hepatocellular carcinoma and non-tumorous tissues without viral hepatitis. J. Gastroenterol. Hepatol. 2008, 23, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.S.; Co, W.S.; Donat, C.; Wang, M.; Che, W.; Prabhala, P.; Schuster, F.; Schulz, V.; Martin, J.L.; Ammit, A.J. Repression of breast cancer cell growth by proteasome inhibitors in vitro: Impact of mitogen-activated protein kinase phosphatase 1. Cancer Biol. Ther. 2015. [Google Scholar] [CrossRef] [PubMed]
- Garza-Veloz, I.; Martinez-Fierro, M.L.; Jaime-Perez, J.C.; Carrillo-Sanchez, K.; Hoyo, R.-D.; Guadalupe, M.; Lugo-Trampe, A.; Rojas-Martinez, A.; Gutierrez-Aguirre, C.H.; Gonzalez-Llano, O. Identification of differentially expressed genes associated with prognosis of B acute lymphoblastic leukemia. Dis. Markers 2015, 2015, 828145. [Google Scholar] [CrossRef] [PubMed]
- Bielskiene, K.; Bagdoniene, L.; Mozuraitiene, J.; Kazbariene, B.; Janulionis, E. E3 ubiquitin ligases as drug targets and prognostic biomarkers in melanoma. Medicina 2015, 51, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Zhang, L.; Zhou, Y.; Xiao, J.; Li, S.; Chen, Y.; Qiao, Z.; Niu, J.; Gu, Y. Angiotensin-(1-7) attenuates damage to podocytes induced by preeclamptic serum through MAPK pathways. Int. J. Mol. Med. 2014, 34, 1057–1064. [Google Scholar] [CrossRef]
- Knofler, M.; Pollheimer, J. Human placental trophoblast invasion and differentiation: A particular focus on Wnt signaling. Front. Genet. 2013, 4, 190. [Google Scholar] [CrossRef] [PubMed]
- Chin, L.J.; Slack, F.J. A truth serum for cancer--microRNAs have major potential as cancer biomarkers. Cell Res. 2008, 18, 983–984. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | WWD-PE (n = 16) | Control (n = 18) | p-Value |
|---|---|---|---|
| Maternal Age (years) | 23.5 ± 5.1 | 23.4 ± 5.8 | 0.913 |
| Body mass index | 28.1 ± 5.5 | 27.2 ± 4.9 | 0.639 |
| Number of pregnancies | 2 (1–5) | 1 (1–4) | 0.387 |
| First pregnancy | 6 (37.5) | 9 (50.0) | 0.699 |
| Weeks of gestation at PE diagnosis | 34.6 + 5.4 | - | - |
| Early preeclampsia n (%) | 4 (25.0) | - | - |
| Severe preeclampsia n (%) | 5 (31.25) | - | - |
| Gestational Age | miRNAs in the Analysis | KEGG Pathway | Enrichment p-Value | No. of Target Genes | No. of miRNAs in the Gene Intersection | Gene Name |
|---|---|---|---|---|---|---|
| 12 WG | 3 | Adherens junction | 0.0007 | 1 | 3 | NLK |
| Wnt signaling pathway | 0.0009 | 1 | 3 | |||
| MAPK signaling pathway | 0.0013 | 1 | 3 | |||
| 16 WG | 19 | Ubiquitin mediated proteolysis | 0.0011 | 1 | 7 | UBE2W TAOK1 JMJD1C |
| MAPK signaling pathway | 0.0013 | 1 | 7 | |||
| Transcriptional misregulation in cancer | 0.0013 | 1 | 7 | |||
| 20 WG | 9 | Fanconi anemia pathway | 0.0009 | 1 | 6 | REV3L ELK4 |
| MAPK signaling pathway | 0.0009 | 1 | 6 | |||
| Metabolic pathways | 0.0055 | 1 | 6 | |||
| Transcriptional misregulation in cancer | 0.0068 | 1 | 6 | |||
| HTLV-I infection | 0.0079 | 1 | 6 | |||
| PE diagnosis time 1 | 14 | Lysine degradation | 1.62 × 10−5 | 1 | 5 | KMT2C NFAT5 JMJD1C NR3C1 |
| VEGF signaling pathway | 0.0072 | 1 | 5 | |||
| B cell receptor signaling pathway | 0.0072 | 1 | 5 | |||
| T cell receptor signaling pathway | 0.0105 | 1 | 5 | |||
| Wnt signaling pathway | 0.0139 | 1 | 5 | |||
| Axon guidance | 0.0139 | 1 | 5 | |||
| Natural killer cell-mediated cytotoxicity | 0.0139 | 1 | 5 | |||
| Hepatitis B | 0.0139 | 1 | 5 | |||
| Transcriptional misregulation in cancer | 0.0191 | 1 | 5 | |||
| Neuroactive ligand-receptor interaction | 0.0229 | 1 | 5 | |||
| HTLV-I infection | 0.0229 | 1 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Fierro, M.L.; Garza-Veloz, I. Analysis of Circulating microRNA Signatures and Preeclampsia Development. Cells 2021, 10, 1003. https://doi.org/10.3390/cells10051003
Martinez-Fierro ML, Garza-Veloz I. Analysis of Circulating microRNA Signatures and Preeclampsia Development. Cells. 2021; 10(5):1003. https://doi.org/10.3390/cells10051003
Chicago/Turabian StyleMartinez-Fierro, Margarita L., and Idalia Garza-Veloz. 2021. "Analysis of Circulating microRNA Signatures and Preeclampsia Development" Cells 10, no. 5: 1003. https://doi.org/10.3390/cells10051003
APA StyleMartinez-Fierro, M. L., & Garza-Veloz, I. (2021). Analysis of Circulating microRNA Signatures and Preeclampsia Development. Cells, 10(5), 1003. https://doi.org/10.3390/cells10051003