
Crosstalk between the mTOR and DNA Damage Response Pathways in Fission Yeast




{kind=link}
{kind=link}
Abstract
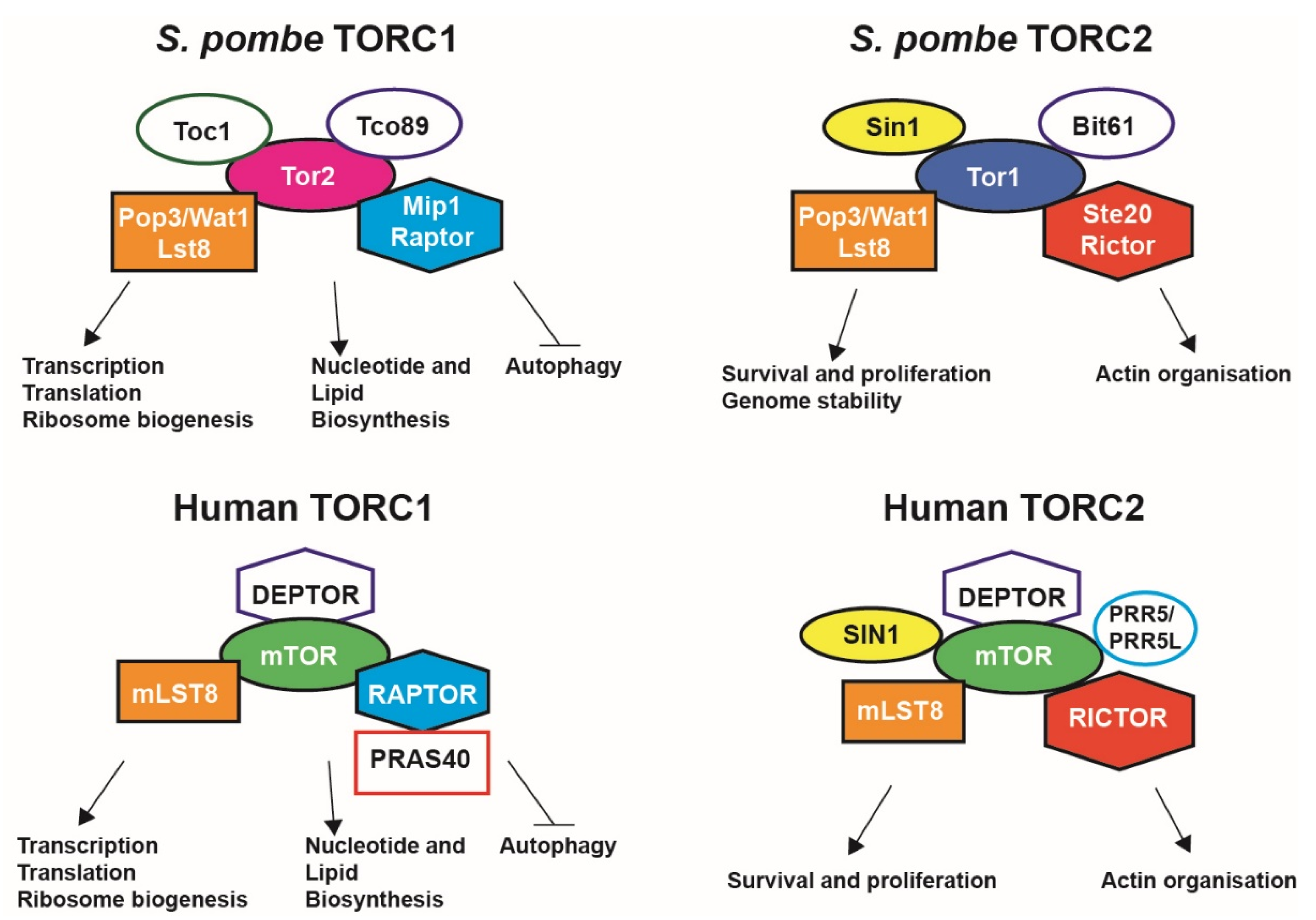
1. The Mechanistic Target of Rapamycin Pathway in Cell Growth and Metabolism
2. DNA Damage Response (DDR) Signaling: Cdc25, Wee1, and DNA Damage CheckPoint Activation
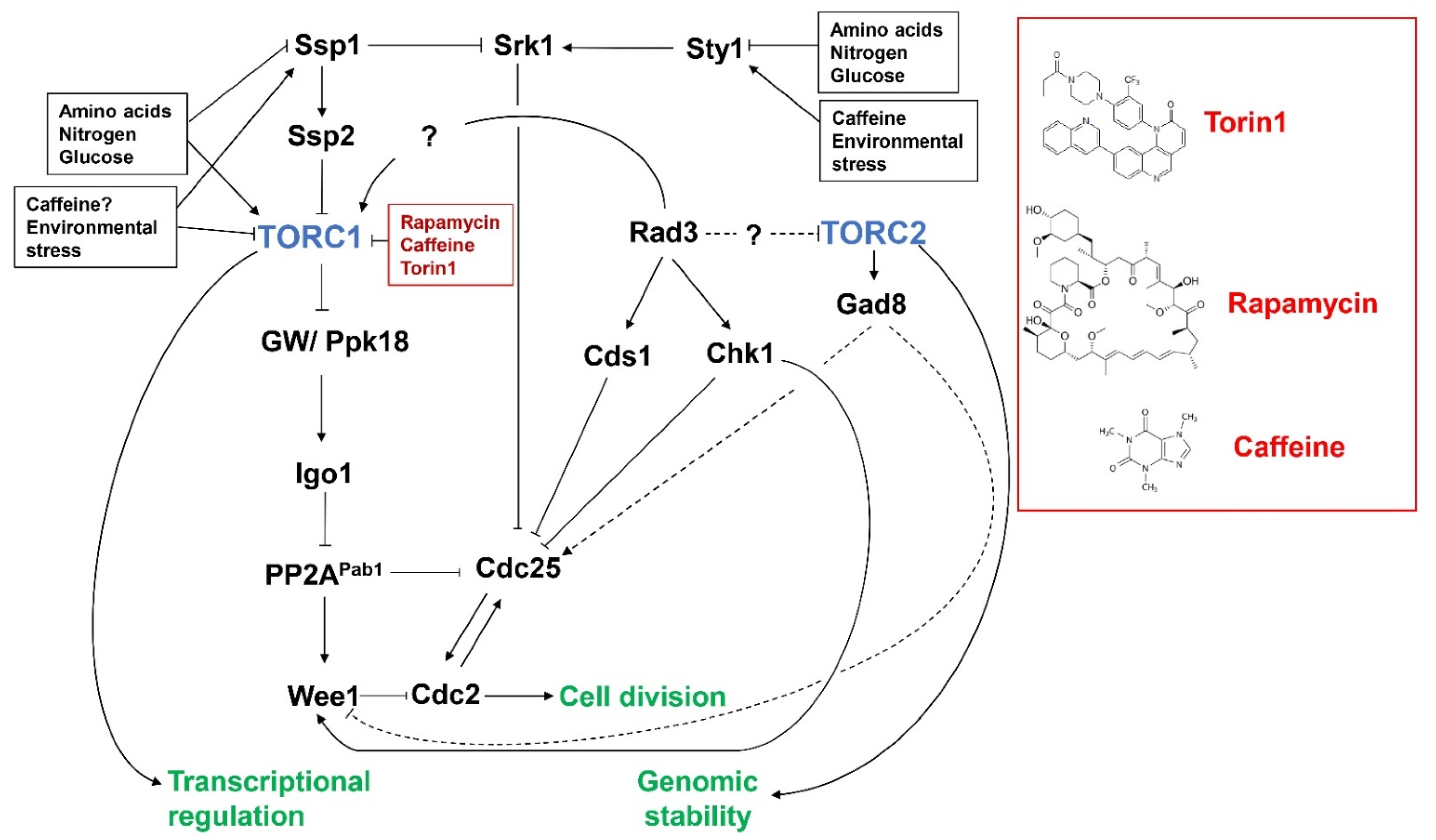
3. TORC1, Caffeine and the DNA Damage Response Pathway
4. Crosstalk between TORC2 and DNA Damage Response Pathways
5. Prospects for mTOR as a Chemo- and Radio-Sensitisation Therapeutic Target
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef]
- Kapahi, P.; Chen, D.; Rogers, A.N.; Katewa, S.D.; Li, P.W.-L.; Thomas, E.L.; Kockel, L. With TOR, Less Is More: A Key Role for the Conserved Nutrient-Sensing TOR Pathway in Aging. Cell Metab. 2010, 11, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for Cell Cycle Arrest by the Immunosuppressant Rapamycin in Yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef]
- Brown, E.J.; Albers, M.W.; Shin, T.B.; Ichikawa, K.; Keith, C.T.; Lane, W.S.; Schreiber, S.L. A Mammalian Protein Targeted by G1-Arresting Rapamycin-Receptor Complex. Nature 1994, 369, 756–758. [Google Scholar] [CrossRef] [PubMed]
- Oldham, S.; Montagne, J.; Radimerski, T.; Thomas, G.; Hafen, E. Genetic and Biochemical Characterization of DTOR, the Drosophila Homolog of the Target of Rapamycin. Genes Dev. 2000, 14, 2689–2694. [Google Scholar] [CrossRef] [PubMed]
- Sabers, C.J.; Martin, M.M.; Brunn, G.J.; Williams, J.M.; Dumont, F.J.; Wiederrecht, G.; Abraham, R.T. Isolation of a Protein Target of the FKBP12-Rapamycin Complex in Mammalian Cells. J. Biol. Chem. 1995, 270, 815–822. [Google Scholar] [CrossRef]
- Sabatini, D.M.; Erdjument-Bromage, H.; Lui, M.; Tempst, P.; Snyder, S.H. RAFT1: A Mammalian Protein That Binds to FKBP12 in a Rapamycin-Dependent Fashion and Is Homologous to Yeast TORs. Cell 1994, 78, 35–43. [Google Scholar] [CrossRef]
- Long, X.; Spycher, C.; Han, Z.S.; Rose, A.M.; Müller, F.; Avruch, J. TOR Deficiency in C. Elegans Causes Developmental Arrest and Intestinal Atrophy by Inhibition of MRNA Translation. Curr. Biol. 2002, 12, 1448–1461. [Google Scholar] [CrossRef]
- Long, X.; Lin, Y.; Ortiz-Vega, S.; Yonezawa, K.; Avruch, J. Rheb Binds and Regulates the MTOR Kinase. Curr. Biol. 2005, 15, 702–713. [Google Scholar] [CrossRef]
- Matsumoto, S.; Bandyopadhyay, A.; Kwiatkowski, D.J.; Maitra, U.; Matsumoto, T. Role of the Tsc1-Tsc2 Complex in Signaling and Transport across the Cell Membrane in the Fission Yeast Schizosaccharomyces pombe. Genetics 2002, 161, 1053–1063. [Google Scholar] [PubMed]
- Huang, J.; Manning, B.D. The TSC1-TSC2 Complex: A Molecular Switchboard Controlling Cell Growth. Biochem. J. 2008, 412, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, M. Cellular Quiescence: Are Controlling Genes Conserved? Trends Cell Biol. 2009, 19, 705–715. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. MTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Nakashima, A.; Ueno, M.; Ushimaru, T.; Aiba, K.; Doi, H.; Uritani, M. Fission Yeast Tor1 Functions in Response to Various Stresses Including Nitrogen Starvation, High Osmolarity, and High Temperature. Curr. Genet. 2001, 39, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Otsubo, Y.; Nakashima, A.; Yamamoto, M.; Yamashita, A. TORC1-Dependent Phosphorylation Targets in Fission Yeast. Biomolecules 2017, 7, 50. [Google Scholar] [CrossRef]
- Weisman, R.; Choder, M. The Fission Yeast TOR Homolog, tor1+, Is Required for the Response to Starvation and Other Stresses via a Conserved Serine. J. Biol. Chem. 2001, 276, 7027–7032. [Google Scholar] [CrossRef]
- Loewith, R.; Hall, M.N. Target of Rapamycin (TOR) in Nutrient Signaling and Growth Control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef]
- Guertin, D.A.; Sabatini, D.M. Defining the Role of MTOR in Cancer. Cancer Cell 2007, 12, 9–22. [Google Scholar] [CrossRef]
- Rallis, C.; Bähler, J. Inhibition of TORC1 Signaling and Increased Lifespan: Gained in Translation? Aging (Albany NY) 2013, 5, 335–336. [Google Scholar] [CrossRef][Green Version]
- Rallis, C.; Codlin, S.; Bähler, J. TORC1 Signaling Inhibition by Rapamycin and Caffeine Affect Lifespan, Global Gene Expression, and Cell Proliferation of Fission Yeast. Aging Cell 2013, 12, 563–573. [Google Scholar] [CrossRef]
- Wanke, V.; Cameroni, E.; Uotila, A.; Piccolis, M.; Urban, J.; Loewith, R.; De Virgilio, C. Caffeine Extends Yeast Lifespan by Targeting TORC1. Mol. Microbiol. 2008, 69, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Kuo, C.J.; Crabtree, G.R.; Blenis, J. Rapamycin-FKBP Specifically Blocks Growth-Dependent Activation of and Signaling by the 70 Kd S6 Protein Kinases. Cell 1992, 69, 1227–1236. [Google Scholar] [CrossRef]
- Ma, Y.; Vassetzky, Y.; Dokudovskaya, S. MTORC1 Pathway in DNA Damage Response. Biochim. Biophys. Acta BBA Mol. Cell Res. 2018, 1865, 1293–1311. [Google Scholar] [CrossRef]
- Ma, X.M.; Blenis, J. Molecular Mechanisms of MTOR-Mediated Translational Control. Nat. Rev. Mol. Cell Biol. 2009, 10, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Tokunaga, C.; Eguchi, S.; Oshiro, N.; Hidayat, S.; Yoshino, K.; Hara, K.; Tanaka, N.; Avruch, J.; Yonezawa, K. The Mammalian Target of Rapamycin (MTOR) Partner, Raptor, Binds the MTOR Substrates P70 S6 Kinase and 4E-BP1 through Their TOR Signaling (TOS) Motif. J. Biol. Chem. 2003, 278, 15461–15464. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.-H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. Rictor, a Novel Binding Partner of MTOR, Defines a Rapamycin-Insensitive and Raptor-Independent Pathway That Regulates the Cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar] [CrossRef]
- Xu, S.; Li, L.; Li, M.; Zhang, M.; Ju, M.; Chen, X.; Gu, H. Impact on Autophagy and Ultraviolet B Induced Responses of Treatment with the MTOR Inhibitors Rapamycin, Everolimus, Torin 1, and Pp242 in Human Keratinocytes. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Gaubitz, C.; Prouteau, M.; Kusmider, B.; Loewith, R. TORC2 Structure and Function. Trends Biochem. Sci. 2016, 41, 532–545. [Google Scholar] [CrossRef]
- Matsuo, T.; Otsubo, Y.; Urano, J.; Tamanoi, F.; Yamamoto, M. Loss of the TOR Kinase Tor2 Mimics Nitrogen Starvation and Activates the Sexual Development Pathway in Fission Yeast. Mol. Cell. Biol. 2007, 27, 3154–3164. [Google Scholar] [CrossRef]
- Du, W.; Halova, L.; Kirkham, S.; Atkin, J.; Petersen, J. TORC2 and the AGC Kinase Gad8 Regulate Phosphorylation of the Ribosomal Protein S6 in Fission Yeast. Biol. Open 2012, 1, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Schonbrun, M.; Laor, D.; López-Maury, L.; Bähler, J.; Kupiec, M.; Weisman, R. TOR Complex 2 Controls Gene Silencing, Telomere Length Maintenance, and Survival under DNA-Damaging Conditions. Mol. Cell. Biol. 2009, 29, 4584–4594. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, S.; Rallis, C. The TOR Signaling Pathway in Spatial and Temporal Control of Cell Size and Growth. Front. Cell Dev. Biol. 2017, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.; Kirkham, S.; Halova, L.; Atkin, J.; Franz-Wachtel, M.; Cobley, D.; Krug, K.; Maček, B.; Mulvihill, D.P.; Petersen, J. TOR Complex 2 Localises to the Cytokinetic Actomyosin Ring and Controls the Fidelity of Cytokinesis. J. Cell Sci. 2016, 129, 2613–2624. [Google Scholar] [CrossRef] [PubMed]
- Ikai, N.; Nakazawa, N.; Hayashi, T.; Yanagida, M. The Reverse, but Coordinated, Roles of Tor2 (TORC1) and Tor1 (TORC2) Kinases for Growth, Cell Cycle and Separase-Mediated Mitosis in Schizosaccharomyces pombe. Open Biol. 2011, 1. [Google Scholar] [CrossRef] [PubMed]
- Weisman, R. Target of Rapamycin (TOR) Regulates Growth in Response to Nutritional Signals. In The Fungal Kingdom; Heitman, J., Howlett, B.J., Crous, P.W., Strukenbrock, E.H., James, T.Y., Grow, N.A.R., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 535–548. [Google Scholar] [CrossRef]
- Schonbrun, M.; Kolesnikov, M.; Kupiec, M.; Weisman, R. TORC2 Is Required to Maintain Genome Stability during S Phase in Fission Yeast. J. Biol. Chem. 2013, 288, 19649–19660. [Google Scholar] [CrossRef]
- Shaltiel, I.A.; Krenning, L.; Bruinsma, W.; Medema, R.H. The Same, Only Different-DNA Damage Checkpoints and Their Reversal throughout the Cell Cycle. J. Cell Sci. 2015, 128, 607–620. [Google Scholar] [CrossRef]
- Jimenez, G.; Yucel, J.; Rowley, R.; Subramani, S. The Rad3+ Gene of Schizosaccharomyces pombe is Involved in Multiple Checkpoint Functions and in DNA Repair. Proc. Natl. Acad. Sci. USA 1992, 89, 4952–4956. [Google Scholar] [CrossRef]
- Bentley, N.J.; Holtzman, D.A.; Flaggs, G.; Keegan, K.S.; DeMaggio, A.; Ford, J.C.; Hoekstra, M.; Carr, A.M. The Schizosaccharomyces pombe Rad3 Checkpoint Gene. EMBO J. 1996, 15, 6641–6651. [Google Scholar] [CrossRef]
- Carr, A.M. DNA Structure Dependent Checkpoints as Regulators of DNA Repair. DNA Repair 2002, 1, 983–994. [Google Scholar] [CrossRef]
- Awasthi, P.; Foiani, M.; Kumar, A. ATM and ATR Signaling at a Glance. J. Cell Sci. 2015, 128, 4255–4262. [Google Scholar] [CrossRef]
- Corcoles-Saez, I.; Dong, K.; Cha, R.S. Versatility of the Mec1ATM/ATR Signaling Network in Mediating Resistance to Replication, Genotoxic, and Proteotoxic Stresses. Curr. Genet. 2019, 65, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Cussiol, J.R.R.; Soares, B.L.; de Oliveira, F.M.B. From Yeast to Humans: Understanding the Biology of DNA Damage Response (DDR) Kinases. Genet. Mol. Biol. 2020, 43, e20190071. [Google Scholar] [CrossRef] [PubMed]
- Crncec, A.; Hochegger, H. Triggering Mitosis. FEBS Lett. 2019, 593, 2868–2888. [Google Scholar] [CrossRef]
- Moseley, J.B. Wee1 and Cdc25: Tools, Pathways, Mechanisms, Questions. Cell Cycle 2017, 16, 599–600. [Google Scholar] [CrossRef]
- O’Connell, M.J.; Walworth, N.C.; Carr, A.M. The G2-Phase DNA-Damage Checkpoint. Trends Cell Biol. 2000, 10, 296–303. [Google Scholar] [CrossRef]
- Karlsson-Rosenthal, C.; Millar, J.B.A. Cdc25: Mechanisms of Checkpoint Inhibition and Recovery. Trends Cell Biol. 2006, 16, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Stracker, T.H.; Roig, I.; Knobel, P.A.; Marjanović, M. The ATM Signaling Network in Development and Disease. Front. Genet. 2013, 4, 37. [Google Scholar] [CrossRef]
- Rhind, N.; Russell, P. Chk1 and Cds1: Linchpins of the DNA Damage and Replication Checkpoint Pathways. J. Cell Sci. 2000, 113 Pt 22, 3889–3896. [Google Scholar]
- Paparatto, D.; Fletcher, D.; Piwowar, K.; Baldino, K.; Morel, C.; Dunaway, S. The Schizosaccharomyces pombe Checkpoint Kinases Chk1 and Cds1 Are Important for Cell Survival in Response to Cisplatin. PLoS ONE 2009, 4, e6181. [Google Scholar] [CrossRef]
- Qu, M.; Yang, B.; Tao, L.; Yates, J.R.; Russell, P.; Dong, M.-Q.; Du, L.-L. Phosphorylation-Dependent Interactions between Crb2 and Chk1 Are Essential for DNA Damage Checkpoint. PLoS Genet. 2012, 8, e1002817. [Google Scholar] [CrossRef] [PubMed]
- Rhind, N.; Russell, P. Mitotic DNA Damage and Replication Checkpoints in Yeast. Curr. Opin. Cell Biol. 1998, 10, 749–758. [Google Scholar] [CrossRef]
- Boddy, M.N.; Furnari, B.; Mondesert, O.; Russell, P. Replication Checkpoint Enforced by Kinases Cds1 and Chk1. Science 1998, 280, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.U.; Bentley, N.J.; Martinho, R.G.; Nielsen, O.; Carr, A.M. Mik1 Levels Accumulate in S Phase and May Mediate an Intrinsic Link between S Phase and Mitosis. Proc. Natl. Acad. Sci. USA 2000, 97, 2579–2584. [Google Scholar] [CrossRef]
- Zeng, Y.; Forbes, K.C.; Wu, Z.; Moreno, S.; Piwnica-Worms, H.; Enoch, T. Replication Checkpoint Requires Phosphorylation of the Phosphatase Cdc25 by Cds1 or Chk1. Nature 1998, 395, 507–510. [Google Scholar] [CrossRef]
- Zeng, Y.; Piwnica-Worms, H. DNA Damage and Replication Checkpoints in Fission Yeast Require Nuclear Exclusion of the Cdc25 Phosphatase via 14-3-3 Binding. Mol. Cell. Biol. 1999, 19, 7410–7419. [Google Scholar] [CrossRef]
- Lopez-Girona, A.; Furnari, B.; Mondesert, O.; Russell, P. Nuclear Localization of Cdc25 Is Regulated by DNA Damage and a 14-3-3 Protein. Nature 1999, 397, 172–175. [Google Scholar] [CrossRef]
- Furnari, B.; Blasina, A.; Boddy, M.N.; McGowan, C.H.; Russell, P. Cdc25 Inhibited in vivo and in vitro by Checkpoint Kinases Cds1 and Chk1. Mol. Biol. Cell 1999, 10, 833–845. [Google Scholar] [CrossRef]
- Frazer, C.; Young, P.G. Carboxy-Terminal Phosphorylation Sites in Cdc25 Contribute to Enforcement of the DNA Damage and Replication Checkpoints in Fission Yeast. Curr. Genet. 2012, 58, 217–234. [Google Scholar] [CrossRef]
- Frazer, C.; Young, P.G. Redundant Mechanisms Prevent Mitotic Entry Following Replication Arrest in the Absence of Cdc25 Hyper-Phosphorylation in Fission Yeast. PLoS ONE 2011, 6, e21348. [Google Scholar] [CrossRef]
- Alao, J.P.; Sjölander, J.J.; Baar, J.; Özbaki-Yagan, N.; Kakoschky, B.; Sunnerhagen, P. Caffeine Stabilizes Cdc 25 Independently of Rad 3 in Schizosaccharomyces pombe Contributing to Checkpoint Override. Mol. Microbiol. 2014, 92, 777–796. [Google Scholar] [CrossRef] [PubMed]
- Kovelman, R.; Russell, P. Stockpiling of Cdc25 during a DNA Replication Checkpoint Arrest in Schizosaccharomyces pombe. Mol. Cell. Biol. 1996, 16, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, J.M.; O’Connell, M.J. The G(2) DNA Damage Checkpoint Targets Both Wee1 and Cdc25. J. Cell Sci. 2000, 113 Pt 10, 1727–1736. [Google Scholar]
- López-Avilés, S.; Lambea, E.; Moldón, A.; Grande, M.; Fajardo, A.; Rodríguez-Gabriel, M.A.; Hidalgo, E.; Aligue, R. Activation of Srk1 by the Mitogen-Activated Protein Kinase Sty1/Spc1 Precedes Its Dissociation from the Kinase and Signals Its Degradation. Mol. Biol. Cell 2008, 19, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Toone, W.M.; Chen, D.; Bähler, J.; Jones, N.; Morgan, B.A.; Quinn, J. The Srk1 Protein Kinase Is a Target for the Sty1 Stress-Activated MAPK in Fission Yeast. J. Biol. Chem. 2002, 277, 33411–33421. [Google Scholar] [CrossRef] [PubMed]
- Atkin, J.; Halova, L.; Ferguson, J.; Hitchin, J.R.; Lichawska-Cieslar, A.; Jordan, A.M.; Pines, J.; Wellbrock, C.; Petersen, J. Torin1-Mediated TOR Kinase Inhibition Reduces Wee1 Levels and Advances Mitotic Commitment in Fission Yeast and HeLa Cells. J. Cell Sci. 2014, 127, 1346–1356. [Google Scholar] [CrossRef]
- Shiozaki, K.; Russell, P. Cell-Cycle Control Linked to Extracellular Environment by MAP Kinase Pathway in Fission Yeast. Nature 1995, 378, 739–743. [Google Scholar] [CrossRef]
- Degols, G.; Russell, P. Discrete Roles of the Spc1 Kinase and the Atf1 Transcription Factor in the UV Response of Schizosaccharomyces pombe. Mol. Cell. Biol. 1997, 17, 3356–3363. [Google Scholar] [CrossRef]
- Coulthard, L.R.; White, D.E.; Jones, D.L.; McDermott, M.F.; Burchill, S.A. P38MAPK: Stress Responses from Molecular Mechanisms to Therapeutics. Trends Mol. Med. 2009, 15, 369–379. [Google Scholar] [CrossRef]
- Rupeš, I.; Jia, Z.; Young, P.G. Ssp1 Promotes Actin Depolymerization and Is Involved in Stress Response and New End Take-Off Control in Fission Yeast. Mol. Biol. Cell 1999, 10, 1495–1510. [Google Scholar] [CrossRef]
- Yanagida, M.; Ikai, N.; Shimanuki, M.; Sajiki, K. Nutrient Limitations Alter Cell Division Control and Chromosome Segregation through Growth-Related Kinases and Phosphatases. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3508–3520. [Google Scholar] [CrossRef] [PubMed]
- Freitag, S.I.; Wong, J.; Young, P.G. Genetic and Physical Interaction of Ssp1 CaMKK and Rad24 14-3-3 during Low PH and Osmotic Stress in Fission Yeast. Open Biol. 2014, 4, 130127. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gómez-Hierro, A.; Lambea, E.; Giménez-Zaragoza, D.; López-Avilés, S.; Yance-Chávez, T.; Montserrat, M.; Pujol, M.J.; Bachs, O.; Aligue, R. Ssp1 CaMKK: A Sensor of Actin Polarization That Controls Mitotic Commitment through Srk1 in Schizosaccharomyces pombe. PLoS ONE 2015, 10, e0143037. [Google Scholar] [CrossRef] [PubMed]
- Davie, E.; Forte, G.M.A.; Petersen, J. Nitrogen Regulates AMPK to Control TORC1 Signaling. Curr. Biol. 2015, 25, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Hartmuth, S.; Petersen, J. Fission Yeast Tor1 Functions as Part of TORC1 to Control Mitotic Entry through the Stress MAPK Pathway Following Nutrient Stress. J. Cell Sci. 2009, 122, 1737–1746. [Google Scholar] [CrossRef]
- Alao, J.P.; Huis in ’t Veld, P.J.; Buhse, F.; Sunnerhagen, P. Hyperosmosis Enhances Radiation and Hydroxyurea Resistance of Schizosaccharomyces pombe Checkpoint Mutants through the Spindle Checkpoint and Delayed Cytokinesis. Mol. Microbiol. 2010, 77, 143–157. [Google Scholar] [CrossRef]
- Alao, J.P.; Sunnerhagen, P. Caffeine as a Tool for Investigating the Integration of Cdc25 Phosphorylation, Activity and Ubiquitin-Dependent Degradation in Schizosaccharomyces pombe. Cell Div. 2020, 15. [Google Scholar] [CrossRef]
- Weisman, R.; Cohen, A.; Gasser, S.M. TORC 2—A New Player in Genome Stability. EMBO Mol. Med. 2014, 6, 995–1002. [Google Scholar] [CrossRef]
- Fletcher, J.; Griffiths, L.; Caspari, T. Nutrient Limitation Inactivates Mrc1-to-Cds1 Checkpoint Signalling in Schizosaccharomyces pombe. Cells 2018, 7, 15. [Google Scholar] [CrossRef]
- Castro, A.; Lorca, T. Greatwall Kinase at a Glance. J. Cell Sci. 2018, 131, jcs222364. [Google Scholar] [CrossRef]
- Pérez-Hidalgo, L.; Moreno, S. Coupling TOR to the Cell Cycle by the Greatwall–Endosulfine–PP2A-B55 Pathway. Biomolecules 2017, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Lopez-Aviles, S. Express Yourself: How PP2A-B55Pab1 Helps TORC1 Talk to TORC2. Curr. Genet. 2018, 64, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Lucena, R.; Alcaide-Gavilán, M.; Anastasia, S.D.; Kellogg, D.R. Wee1 and Cdc25 Are Controlled by Conserved PP2A-Dependent Mechanisms in Fission Yeast. Cell Cycle 2017, 16, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, K.; Nemoto, T.; Nabeshima, K.; Kondoh, H.; Niwa, H.; Yanagida, M. The Regulatory Subunits of Fission Yeast Protein Phosphatase 2A (PP2A) Affect Cell Morphogenesis, Cell Wall Synthesis and Cytokinesis. Genes Cells 1996, 1, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. The Enigmatic Effects of Caffeine in Cell Cycle and Cancer. Cancer Lett. 2007, 247, 26–39. [Google Scholar] [CrossRef]
- Osman, F.; McCready, S. Differential Effects of Caffeine on DNA Damage and Replication Cell Cycle Checkpoints in the Fission Yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 1998, 260, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Sarkaria, J.N.; Busby, E.C.; Tibbetts, R.S.; Roos, P.; Taya, Y.; Karnitz, L.M.; Abraham, R.T. Inhibition of ATM and ATR Kinase Activities by the Radiosensitizing Agent, Caffeine. Cancer Res. 1999, 59, 4375–4382. [Google Scholar]
- Blasina, A.; Price, B.D.; Turenne, G.A.; McGowan, C.H. Caffeine Inhibits the Checkpoint Kinase ATM. Curr. Biol. 1999, 9, 1135–1138. [Google Scholar] [CrossRef]
- Moser, B.A.; Brondello, J.M.; Baber-Furnari, B.; Russell, P. Mechanism of Caffeine-Induced Checkpoint Override in Fission Yeast. Mol. Cell. Biol. 2000, 20, 4288–4294. [Google Scholar] [CrossRef][Green Version]
- Cortez, D. Caffeine Inhibits Checkpoint Responses without Inhibiting the Ataxia-Telangiectasia-Mutated (ATM) and ATM- and Rad3-Related (ATR) Protein Kinases. J. Biol. Chem. 2003, 278, 37139–37145. [Google Scholar] [CrossRef]
- Calvo, I.A.; Gabrielli, N.; Iglesias-Baena, I.; García-Santamarina, S.; Hoe, K.-L.; Kim, D.U.; Sansó, M.; Zuin, A.; Pérez, P.; Ayté, J.; et al. Genome-Wide Screen of Genes Required for Caffeine Tolerance in Fission Yeast. PLoS ONE 2009, 4, e6619. [Google Scholar] [CrossRef]
- Campos, A.; Clemente-Blanco, A. Cell Cycle and DNA Repair Regulation in the Damage Response: Protein Phosphatases Take Over the Reins. Int. J. Mol. Sci. 2020, 21, 446. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Norbury, C.; Harris, A.L.; Toda, T. Caffeine Can Override the S-M Checkpoint in Fission Yeast. J. Cell Sci. 1999, 112 Pt 6, 927–937. [Google Scholar]
- Reinke, A.; Chen, J.C.-Y.; Aronova, S.; Powers, T. Caffeine Targets TOR Complex I and Provides Evidence for a Regulatory Link between the FRB and Kinase Domains of Tor1p. J. Biol. Chem. 2006, 281, 31616–31626. [Google Scholar] [CrossRef]
- Alao, J.P.; Johansson-Sjölander, J.; Rallis, C.; Sunnerhagen, P. Caffeine Stabilises Fission Yeast Wee1 in a Rad24-Dependent Manner but Attenuates Its Expression in Response to DNA Damage. Microorganisms 2020, 8, 1512. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-López, M.; Gonzalez, S.; Hillson, O.; Tunnacliffe, E.; Codlin, S.; Tallada, V.A.; Bähler, J.; Rallis, C. The GATA Transcription Factor Gaf1 Represses TRNAs, Inhibits Growth, and Extends Chronological Lifespan Downstream of Fission Yeast TORC1. Cell Rep. 2020, 30, 3240–3249.e4. [Google Scholar] [CrossRef] [PubMed]
- Pataki, E.; Simhaev, L.; Engel, H.; Cohen, A.; Kupiec, M.; Weisman, R. TOR Complex 2- Independent Mutations in the Regulatory PIF Pocket of Gad8AKT1/SGK1 Define Separate Branches of the Stress Response Mechanisms in Fission Yeast. PLoS Genet. 2020, 16, e1009196. [Google Scholar] [CrossRef] [PubMed]
- Ochotorena, I.L.; Hirata, D.; Kominami, K.; Potashkin, J.; Sahin, F.; Wentz-Hunter, K.; Gould, K.L.; Sato, K.; Yoshida, Y.; Vardy, L.; et al. Conserved Wat1/Pop3 WD-Repeat Protein of Fission Yeast Secures Genome Stability through Microtubule Integrity and May Be Involved in MRNA Maturation. J. Cell Sci. 2001, 114, 2911–2920. [Google Scholar]
- Verma, S.K.; Ranjan, R.; Kumar, V.; Siddiqi, M.I.; Ahmed, S. Wat1/Pop3, a Conserved WD Repeat Containing Protein Acts Synergistically with Checkpoint Kinase Chk1 to Maintain Genome Ploidy in Fission Yeast S. pombe. PLoS ONE 2014, 9, e89587. [Google Scholar] [CrossRef]
- Tenzer, A.; Pruschy, M. Potentiation of DNA-Damage-Induced Cytotoxicity by G2 Checkpoint Abrogators. Curr. Med. Chem. Anti Cancer Agents 2003, 3, 35–46. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Iliakis, G.; Wang, Y. Caffeine-Induced Radiosensitization Is Independent of Nonhomologous End Joining of DNA Double-Strand Breaks. Radiat. Res. 2003, 159, 426–432. [Google Scholar] [CrossRef]
- Sinn, B.; Tallen, G.; Schroeder, G.; Grassl, B.; Schulze, J.; Budach, V.; Tinhofer, I. Caffeine Confers Radiosensitization of PTEN-Deficient Malignant Glioma Cells by Enhancing Ionizing Radiation-Induced G1 Arrest and Negatively Regulating Akt Phosphorylation. Mol. Cancer Ther. 2010, 9, 480–488. [Google Scholar] [CrossRef] [PubMed]
- van Dam, R.M.; Hu, F.B.; Willett, W.C. Coffee, Caffeine, and Health. N. Engl. J. Med. 2020, 383, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Saud, S.M.; Young, M.R.; Chen, G.; Hua, B. Targeting AMPK for Cancer Prevention and Treatment. Oncotarget 2015, 6, 7365–7378. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, S.; Higurashi, T.; Nakajima, A. AMPK: Therapeutic Target for Diabetes and Cancer Prevention. Curr. Pharm. Des. 2017, 23, 3629–3644. [Google Scholar] [CrossRef]
- Vara-Ciruelos, D.; Russell, F.M.; Hardie, D.G. The Strange Case of AMPK and Cancer: Dr Jekyll or Mr Hyde? Open Biol. 2019, 9. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alao, J.-P.; Legon, L.; Rallis, C. Crosstalk between the mTOR and DNA Damage Response Pathways in Fission Yeast. Cells 2021, 10, 305. https://doi.org/10.3390/cells10020305
Alao J-P, Legon L, Rallis C. Crosstalk between the mTOR and DNA Damage Response Pathways in Fission Yeast. Cells. 2021; 10(2):305. https://doi.org/10.3390/cells10020305
Chicago/Turabian StyleAlao, John-Patrick, Luc Legon, and Charalampos Rallis. 2021. "Crosstalk between the mTOR and DNA Damage Response Pathways in Fission Yeast" Cells 10, no. 2: 305. https://doi.org/10.3390/cells10020305
APA StyleAlao, J.-P., Legon, L., & Rallis, C. (2021). Crosstalk between the mTOR and DNA Damage Response Pathways in Fission Yeast. Cells, 10(2), 305. https://doi.org/10.3390/cells10020305