Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification




Abstract
1. Introduction
2. Common Dietary Deficiencies in Minerals and Vitamins and the Corresponding Biofortification Strategies
2.1. High Iron (Fe) Acquisition Can Be Achieved through Soil Microbes
2.2. Zinc (Zn) Deficiency Can Be Eradicated Using Soil Microbes
2.3. Vitamins Are Popular Targets of Biofortification
3. Microbes Enhance the Level of Beneficial Secondary Metabolites
3.1. Flavonoids
3.2. Saponins
4. Endophytes as the Emerging Participant of Microbe-Mediated Biofortification
5. The Mechanisms of Microbe-Mediated Biofortification
5.1. By Improving the Availability of Nutrients to Be Absorbed by Plants
5.2. By Directly Synthesizing and Releasing Micronutrients Either in the Soil or as Endophytes
5.3. By Inducing the Plants to Synthesize the Micronutrients
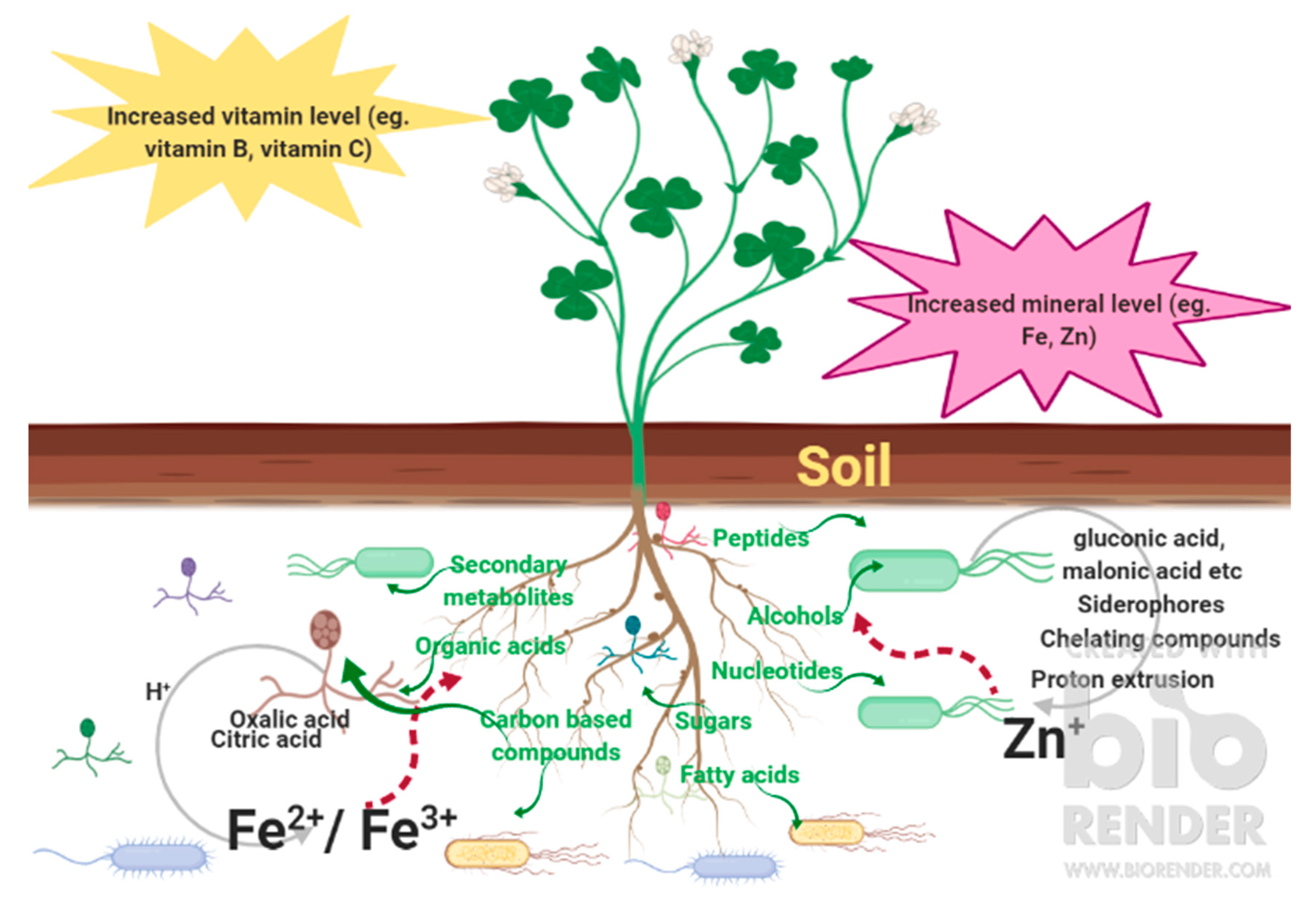
6. How Root Exudates Help to Shape the Rhizobiome
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Müller, C.; Elliott, J.; Levermann, A. Fertilizing hidden hunger. Nat. Publ. Gr. 2014, 4, 540–541. [Google Scholar] [CrossRef]
- Smith, M.R.; Golden, C.D.; Myers, S.S. Potential rise in iron deficiency due to future anthropogenic carbon dioxide emissions. GeoHealth 2017, 1, 248–257. [Google Scholar] [CrossRef]
- Umbreit, J. Iron deficiency: A concise review. Am. J. Hematol. 2005, 231, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Beal, T.; Massiot, E.; Arsenault, J.E.; Smith, M.R. Global trends in dietary micronutrient supplies and estimated prevalence of inadequate intakes. PLoS ONE 2017, 12, e0175554. [Google Scholar] [CrossRef] [PubMed]
- Murgia, I.; Arosio, P.; Tarantino, D.; Soave, C. Biofortification for combating ‘ hidden hunger ’ for iron. Trends Plant Sci. 2012, 17, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.M.; Cooper, J.W. Legume biofortification is an underexploited strategy for combatting hidden hunger. Plant Cell Environ. 2018, 42, 52–70. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Neelam, K.; Tiwari, V.K.; Dhaliwal, H.S. Review Biofortification of cereals to overcome hidden hunger. Plant Breed. 2013, 132, 437–445. [Google Scholar]
- Food, S.; Nestel, P.; Bouis, H.E.; Meenakshi, J.V.; Pfeiffer, W. Biofortification of staple food crops. J. Nutr. 2006, 136, 1064–1067. [Google Scholar]
- Bouis, H.E.; Welch, R.M. Biofortification—a sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Sci. 2010, 50, S-20–S-32. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Garg, M. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef]
- Carvalho, S.M.P.; Vasconcelos, M.W. Producing more with less: strategies and novel technologies for plant-based food biofortification. Food Res. Int. 2013, 54, 961–971. [Google Scholar] [CrossRef]
- Anderson, J.C.; Appleton, E.; Densmore, D.; Endy, D.; Galdzicki, M.; Gennari, J.H.; Grünberg, R.; Huynh, L.; David, J.; Kahl, L.J.; et al. Status and market potential of transgenic biofortified crops. Nat. Biotechnol. 2015, 33, 25–29. [Google Scholar]
- FAO Human vitamin and mineral requirements. In Proceedings of the FAO/WHO expert consultation on human vitamin and mineral requirements, Bangkok, Thailand, 21–30 September 1998.
- García-seco, D.; Bonilla, A.; Algar, E.; García-villaraco, A.; Mañero, J.G.; Ramos-solano, B. Enhanced blackberry production using Pseudomonas fluorescens as elicitor. Agron. Sustain. Dev. 2013, 33, 385–392. [Google Scholar] [CrossRef]
- Garcia-seco, D.; Zhang, Y.; Gutierrez-ma, F.J.; Martin, C.; Ramos-solano, B. Application of Pseudomonas fluorescens to blackberry under field conditions improves fruit quality by modifying flavonoid metabolism. PLoS ONE 2015, 10, e0142639. [Google Scholar] [CrossRef]
- Jinal, H.N.; Gopi, K.; Prittesh, P.; Kartik, V.P.; Amaresan, N. Phytoextraction of iron from contaminated soils by inoculation of iron-tolerant plant growth-promoting bacteria in Brassica juncea L. Czern. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Vadlamudi, S.; Samineni, S.; Kumar, C.V.S. Plant growth- promotion and biofortification of chickpea and pigeonpea through inoculation of biocontrol potential bacteria, isolated from organic soils. Springerplus 2016, 5, 1882. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S. Enhancing ecosystem services in sustainable agriculture: Biofertilization and biofortification of chickpea (Cicer arietinum L.) by arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2014, 68, 429–439. [Google Scholar] [CrossRef]
- Sathya, A.; Vijayabharathi, R.; Srinivas, V.; Gopalakrishnan, S. Plant growth-promoting actinobacteria on chickpea seed mineral density: An upcoming complementary tool for sustainable biofortification strategy. 3 Biotech 2016, 6, 138. [Google Scholar] [CrossRef]
- Baslam, M.; Garmendia, I.; Goicoechea, N.; Unidad, V.; Icvv, Z. Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse-grown lettuce. J. Agric. Food Chem. 2011, 59, 5504–5515. [Google Scholar] [CrossRef]
- Kamarudin, A.N.; Seman, I.A.; Palm, M.; Board, O.; Norhana, Z.; Yusof, B. Thiamine biosynthesis gene expression analysis in Elaeis guineensis during interactions with Hendersonia toruloidea. J. Oil Palm Res. 2017, 29, 218–226. [Google Scholar] [CrossRef][Green Version]
- Kamarudin, A.N.; Lai, K.S.; Lamasudin, D.U.; Idris, A.S. Enhancement of thiamine biosynthesis in oil palm seedlings by colonization of endophytic fungus Hendersonia toruloidea. Front. Plant Sci. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Manjunath, M.; Kanchan, A.; Ranjan, K.; Venkatachalam, S.; Prasanna, R.; Ramakrishnan, B.; Hossain, F.; Nain, L.; Shivay, Y.S.; Rai, A.B.; et al. Beneficial cyanobacteria and eubacteria synergistically enhance bioavailability of soil nutrients and yield of okra. Heliyon 2016, 2, e00066. [Google Scholar] [CrossRef] [PubMed]
- Rozpądek, P.; Rąpała-Kozik, M.; Wężowicz, K.; Grandin, A.; Karlsson, S.; Ważny, R.; Anielska, T.; Turnau, K. Arbuscular mycorrhiza improves yield and nutritional properties of onion (Allium cepa). Plant Physiol. Biochem. 2016, 107, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, W.; Zhou, S.; Wu, S. Mycorrhizal inoculation affects Pb and Cd accumulation and translocation in pakchoi (Brassica chinensis L.). Pedosphere 2016, 26, 13–26. [Google Scholar] [CrossRef]
- Ismail, Y.; McCormick, S.; Hijri, M. The arbuscular mycorrhizal fungus, Glomus irregulare, controls the mycotoxin production of Fusarium sambucinum in the pathogenesis of potato. FEMS Microbiol. Lett. 2013, 348, 46–51. [Google Scholar] [CrossRef]
- Shakeel, M.; Rais, A.; Hassan, M.N.; Hafeez, F.Y. Root associated Bacillus sp. improves growth, yield and zinc translocation for basmati rice (Oryza sativa) varieties. Front. Microbiol. 2015, 6, 1286. [Google Scholar] [CrossRef]
- Adak, A.; Prasanna, R.; Babu, S.; Bidyarani, N.; Verma, S.; Pal, M.; Shivay, Y.S.; Nain, L. Micronutrient enrichment mediated by plant-microbe interactions and rice cultivation practices. J. Plant Nutr. 2016, 39, 1216–1232. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Zhang, X.; Dong, L.; Zhang, J.; Wei, Y.; Feng, Y.; Lu, L. Improved plant growth and Zn accumulation in grains of rice (Oryza sativa L.) by inoculation of endophytic microbes isolated from a Zn hyperaccumulator, Sedum alf redii H. J. Agric. Food Chem. 2014, 62, 1783–1791. [Google Scholar] [CrossRef]
- Rana, A.; Kabi, S.R.; Verma, S.; Adak, A.; Pal, M.; Shivay, Y.S.; Prasanna, R.; Nain, L. Prospecting plant growth promoting bacteria and cyanobacteria as options for enrichment of macro- and micronutrients in grains in rice–wheat cropping sequence. Cogent Food Agric. 2015, 1. [Google Scholar] [CrossRef]
- Dave, S.; Tarafdar, J.C. Stimulatory synthesis of saponin by mycorrhizal fungi in safed musli (Chlorophytum borivilianum) tubers. Int. Res. J. Agric. Sci. Soil Sci. 2011, 1, 137–141. [Google Scholar]
- Gosal, S.K.; Karlupia, A.; Gosal, S.S.; Chhibba, I.M.; Varma, A. Biotization with Piriformospora indica and Pseudomonas fluorescens improves survival rate, nutrient acquisition, field performance and saponin content of micropropagated Chlorophytum sp. Indian J. Biotechnol. 2010, 9, 289–297. [Google Scholar]
- Salinas-Ramírez, N.; Escalante-Estrada, J.A.; Rodríguez-González, M.T.; Sosa-Montes, E. Yield and nutritional quality of snap bean in terms of biofertilization. Trop. Subtrop. Agroecosyst. 2011, 13, 347–355. [Google Scholar]
- Khalid, M.; Hassani, D.; Bilal, M.; Asad, F.; Huang, D. Influence of bio-fertilizer containing beneficial fungi and rhizospheric bacteria on health promoting compounds and antioxidant activity of Spinacia oleracea L. Bot. Stud. 2017, 58, 35. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; Agostino, G.D.; Massa, N.; Avidano, L.; et al. AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef]
- Flores-félix, J.D.; Silva, L.R.; Rivera, L.P.; Marcos-garcía, M. Plants probiotics as a tool to produce highly functional fruits: The Case of Phyllobacterium and vitamin C in strawberries. PLoS ONE 2015, 10, e0122281. [Google Scholar] [CrossRef]
- Erturk, Y.; Ercisli, S.; Cakmakci, R. Yield and growth response of strawberry to plant growth-promoting Rhizobacteria inoculation. J. Plant Nutr. 2012, 35, 817–826. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; Agostino, G.D.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Hart, M.; Ehret, D.L.; Krumbein, A.; Leung, C.; Murch, S.; Turi, C.; Franken, P. Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 2015, 25, 359–376. [Google Scholar] [CrossRef]
- Ochoa-velasco, C.E.; Valadez-blanco, R.; Salas-coronado, R.; Sustaita-rivera, F.; Hernández-carlos, B.; García-ortega, S.; Santos-sánchez, N.F. Effect of nitrogen fertilization and Bacillus licheniformis biofertilizer addition on the antioxidants compounds and antioxidant activity of greenhouse cultivated tomato fruits (Solanum lycopersicum L. var. Sheva). Sci. Hortic. (Amsterdam) 2016, 201, 338–345. [Google Scholar] [CrossRef]
- Mäder, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A.; et al. Inoculation of root microorganisms for sustainable wheat-rice and wheat-black gram rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Yasin, M.; El-Mehdawi, A.F.; Pilon-Smits, E.A.; Faisal, M. Selenium-fortified wheat: Potential of microbes for biofortification of selenium and other essential nutrients. Int. J. Phytoremed. 2015, 17, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Ingraffia, R.; Amato, G.; Frenda, A.S.; Giambalvo, D. Impacts of arbuscular mycorrhizal fungi on nutrient uptake, N2 fixation, N transfer, and growth in a wheat/faba bean intercropping system. PLoS ONE 2019, 14, e0213672. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Acuña, J.J.; Jorquera, M.A.; Azcón, R.; Borie, F.; Cornejo, P.; Mora, M.L. Enhanced selenium content in wheat grain by co-inoculation of selenobacteria and arbuscular mycorrhizal fungi: A preliminary study as a potential Se biofortification strategy. J. Cereal Sci. 2013, 57, 275–280. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron Uptake, Translocation, and Regulation in Higher Plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Jin, C.W.; Ye, Y.Q.; Zheng, S.J. An underground tale: Contribution of microbial activity to plant iron acquisition via ecological processes. Ann. Bot. 2014, 113, 7–18. [Google Scholar] [CrossRef]
- Jin, C.W.; Li, G.X.; Yu, X.H.; Zheng, S.J. Plant Fe status affects the composition of siderophore-secreting microbes in the rhizosphere. Ann. Bot. 2010, 105, 835–841. [Google Scholar] [CrossRef]
- Ivanov, R.; Brumbarova, T.; Bauer, P. Fitting into the harsh reality: Regulation of iron-deficiency responses in dicotyledonous plants. Mol. Plant 2012, 5, 27–42. [Google Scholar] [CrossRef]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef]
- Jin, C.W.; He, Y.F.; Tang, C.X.; Wu, P.; Zheng, S.J. Mechanisms of microbially enhanced Fe acquisition in red clover (Trifolium pratense L.). Plant Cell Environ. 2006, 29, 888–897. [Google Scholar] [CrossRef]
- De Santiago, A.; García-López, A.M.; Quintero, J.M.; Avilés, M.; Delgado, A. Effect of Trichoderma asperellum strain T34 and glucose addition on iron nutrition in cucumber grown on calcareous soils. Soil Biol. Biochem. 2013, 57, 598–605. [Google Scholar] [CrossRef]
- Philpott, C.C. Iron uptake in fungi: A system for every source. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Hannauer, M.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Bonnefoy, V.; Holmes, D.S. Genomic insights into microbial iron oxidation and iron uptake strategies in extremely acidic environments. Environ. Microbiol. 2012, 14, 1597–1611. [Google Scholar] [CrossRef] [PubMed]
- Curie, C.; Panaviene, Z.; Loulergue, C.; Dellaporta, S.L.; Briat, J.F.; Walker, E.L. Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 2001, 409, 346–349. [Google Scholar] [CrossRef]
- Guerinot, M.L.; Yi, Y. Iron: Nutritious, noxious, and not readily available. Plant Physiol. 1994, 104, 815–820. [Google Scholar] [CrossRef]
- Jetten, M.S.; Logemann, S.; Muyzer, G.; Robertson, L.A.; De Vries, S.; van Loosdrecht, M.C.; Kuenen, J.G. Novel principles in the microbial conversion of nitrogen compounds. Antonie Van Leeuwenhoek 1997, 71, 75–93. [Google Scholar] [CrossRef]
- Illmer, P.; Schinner, F. Solubilization of inorganic calcium phosphates—Solubilization mechanisms. Soil Biol. Biochem. 1995, 27, 257–263. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Y.; Xie, X.; Kim, M.S.; Dowd, S.E.; Pare, P.W. A soil bacterium regulates plant acquisition of iron via deficiency-inducible mechanisms. Plant J. 2009, 58, 568–577. [Google Scholar] [CrossRef]
- Jin, C.W.; You, G.Y.; Zheng, S.J. The iron deficiency-induced phenolics secretion plays multiple important roles in plant iron acquisition underground. Plant Signal Behav. 2008, 3, 60–61. [Google Scholar] [CrossRef]
- Chen, W.W.; Yang, J.L.; Qin, C.; Jin, C.W.; Mo, J.H.; Ye, T.; Zheng, S.J. Nitric oxide acts downstream of auxin to trigger root ferric-chelate reductase activity in response to iron deficiency in Arabidopsis. Plant Physiol. 2010, 154, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Graziano, M.; Lamattina, L. Nitric oxide accumulation is required for molecular and physiological responses to iron deficiency in tomato roots. Plant J. 2007, 52, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.J.; Suarez, V.; Romera, F.J.; Alcantara, E.; Perez-Vicente, R. A new model involving ethylene, nitric oxide and Fe to explain the regulation of Fe-acquisition genes in Strategy I plants. Plant Physiol. Biochem. 2011, 49, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Seguela, M.; Briat, J.F.; Vert, G.; Curie, C. Cytokinins negatively regulate the root iron uptake machinery in Arabidopsis through a growth-dependent pathway. Plant J. 2008, 55, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Li, Y.; Zhang, W.H. Brassinosteroids are involved in response of cucumber (Cucumis sativus) to iron deficiency. Ann. Bot. 2012, 110, 681–688. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Kim, D.G.; Vargas, R.; Bond-Lamberty, B.; Turetsky, M.R. Effects of soil rewetting and thawing on soil gas fluxes: A review of current literature and suggestions for future research. Biogeosciences 2012, 9, 2459–2483. [Google Scholar] [CrossRef]
- Jin, C.W.; Du, S.T.; Shamsi, I.H.; Luo, B.F.; Lin, X.Y. NO synthase-generated NO acts downstream of auxin in regulating Fe-deficiency-induced root branching that enhances Fe-deficiency tolerance in tomato plants. J. Exp. Bot. 2011, 62, 3875–3884. [Google Scholar] [CrossRef]
- Meiser, J.; Lingam, S.; Bauer, P. Posttranslational regulation of the iron deficiency basic helix-loop-helix transcription factor FIT is affected by iron and nitric oxide. Plant Physiol. 2011, 157, 2154–2166. [Google Scholar] [CrossRef]
- Jäckel, U.; Schnell, S.; Conrad, R. Microbial ethylene production and inhibition of methanotrophic activity in a deciduous forest soil. Soil Biol. Biochem. 2004, 36, 835–840. [Google Scholar] [CrossRef]
- Wu, J.; Wang, C.; Zheng, L.; Wang, L.; Chen, Y.; Whelan, J.; Shou, H. Ethylene is involved in the regulation of iron homeostasis by regulating the expression of iron-acquisition-related genes in Oryza sativa. J. Exp. Bot. 2011, 62, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Terpolilli, J.J.; Hood, G.A.; Poole, P.S. What determines the efficiency of N2-fixing Rhizobium-legume symbioses? Adv. Microb. Physiol. 2012, 60, 325–389. [Google Scholar] [PubMed]
- Slatni, T.; Dell’Orto, M.; Ben Salah, I.; Vigani, G.; Smaoui, A.; Gouia, H.; Zocchi, G.; Abdelly, C. Immunolocalization of H+-ATPase and IRT1 enzymes in N2-fixing common bean nodules subjected to iron deficiency. J. Plant Physiol. 2012, 169, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Bisht, S.C.; Ruwari, P.; Joshi, G.K.; Singh, G.; Bisht, J.K.; Bhatt, J.C. Bioassociative effect of cold tolerant Pseudomonas spp. and Rhizobium leguminosarum-PR1 on iron acquisition, nutrient uptake and growth of lentil (Lens culinaris L.). Eur. J. Soil Biol. 2011, 47, 35–43. [Google Scholar] [CrossRef]
- Mishra, P.K.; Bisht, S.C.; Mishra, S.; Selvakumar, G.; Bisht, J.K.; Gupta, H.S. Coinoculation of Rhizobium Leguminosarum-PR1 with a cold tolerant Pseudomonas sp. improves iron acquisition, nutrient uptake and growth of field pea (Pisum Sativum L.). J. Plant Nutr. 2012, 35, 243–256. [Google Scholar] [CrossRef]
- Welch, R.M. Breeding strategies for biofortified staple plant foods to reduce micronutrient malnutrition globally. J. Nutr. 2002, 132, 495s–499s. [Google Scholar] [CrossRef]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef]
- Alloway, B.J. Micronutrients and crop production: An introduction. In Micronutrient Deficiencies in Global Crop Production; Alloway, B.J., Ed.; Springer: Dordrecht, the Netherlands, 2008; pp. 1–39. ISBN 978-1-4020-6860-7. [Google Scholar]
- Tavallali, V.; Rahemi, M.; Maftoun, M.; Panahi, B.; Karimi, S.; Ramezanian, A.; Vaezpour, M. Scientia horticulturae zinc influence and salt stress on photosynthesis, water relations, and carbonic anhydrase activity in pistachio. Sci. Hortic. 2009, 123, 272–279. [Google Scholar] [CrossRef]
- Cakamk, I.; Marschener, H. Enhanced Superoxide Radical Production in Roots of Zinc-Deficient Plants. J. Exp. Bot. 1988, 39, 1449–1460. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Hussain, A.; Zahir, Z.A.; Asghar, H.N.; Ahmad, M.; Jamil, M.; Naveed, M.; Zaman Akhtar, M.F.U. Zinc solubilizing bacteria for zinc biofortification in cereals: a step toward sustainable nutritional security. In Role of Rhizospheric Microbes in Soil: Volume 2: Nutrient Management and Crop Improvement; Meena, V.S., Ed.; Springer: Singapore, 2018; pp. 203–227. ISBN 978-981-13-0044-8. [Google Scholar]
- Imran, M.; Arshad, M.; Khalid, A.; Kanwal, S.; Crowley, D.E. Perspectives of rhizosphere microflora for improving Zn bioavailability and acquisition by higher plants. Int. J. Agric. Biol. 2014, 16, 653–662. [Google Scholar]
- Saravanan, V.S.; Madhaiyan, M.; Thangaraju, M. Solubilization of zinc compounds by the diazotrophic, plant growth promoting bacterium Gluconacetobacter diazotrophicus. Chemosphere 2007, 66, 1794–1798. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A. Soil microbiology and biochemistry in perspective. In Soil Microbiology and Biochemistry; Paul, E.A., Clark, F.E., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 1–10. ISBN 978-0-12-546805-3. [Google Scholar]
- Ahmad, M.; Nadeem, S.M.; Naveed, M.; Zahir, Z.A. Potassium-solubilizing bacteria and their application in agriculture. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 293–313. ISBN 978-81-322-2776-2. [Google Scholar]
- Biari, A.; Gholami, A.; Rahmani, H.A. Growth promotion and enhanced nutrient uptake of maize (Zea mays L.) by application of plant growth promoting Rhizobacteria in arid region of Iran. J. Biol. Sci. 2008, 8, 1015–1020. [Google Scholar]
- Goteti, P.K.; Emmanuel, L.D.A.; Desai, S.; Shaik, M.H.A. Prospective zinc solubilising bacteria for enhanced nutrient uptake and growth promotion in maize (Zea mays L.). Int. J. Microbiol. 2013, 1, 869697. [Google Scholar] [CrossRef]
- Hafeez, F.Y.; Hameed, S.; Zaidi, A.H.; Malik, K.A. Techniques for sustainable agriculture. In Biofertilizers for Sustainable Agriculture; Azam, F., Iqbal, M.M., Inayatullah, C., Malik, K.A., Eds.; NIAB: Faisalabad, Pakistan, 2002; pp. 67–73. [Google Scholar]
- Tariq, M.; Hameed, S.; Malik, K.A.; Hafeez, F.Y. Plant root associated bacteria for zinc mobilization in rice. Pakistan J. Bot. 2007, 39, 245. [Google Scholar]
- Ramesh, A.; Sharma, S.K.; Sharma, M.P.; Yadav, N.; Joshi, O.P. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in Vertisols of central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Ullah, M.A.; Yusuf, F. Biofertilizer/biopesticide potentiality of zinc solubilizing Pseudomonas aeruginosa FA-9 and Enterobactersp. FA-11 isolated from the wheat rhizosphere grown in arid zone. Access Microbiol. 2019, 1. [Google Scholar] [CrossRef]
- Sultana, U.; Desai, S.; Reddy, G.; Prasad, T. Zinc solubilizing plant growth promoting microbes produce zinc nanoparticles. bioRxiv 2019, 602219. [Google Scholar] [CrossRef]
- Costerousse, B.; Schönholzer-Mauclaire, L.; Frossard, E.; Thonar, C. Identification of heterotrophic zinc mobilization processes among bacterial strains isolated from wheat rhizosphere (Triticum aestivum L.). Appl. Environ. Microbiol. 2017, 84, e01715-17. [Google Scholar] [CrossRef]
- Rose, M.T.; Pariasca-Tanaka, J.; Rose, T.J.; Wissuwa, M. Bicarbonate tolerance of Zn-efficient rice genotypes is not related to organic acid exudation, but to reduced solute leakage from roots. Funct. Plant Biol. 2011, 38, 493–504. [Google Scholar] [CrossRef]
- Impa, S.M.; Johnson-Beebout, S.E. Mitigating zinc deficiency and achieving high grain Zn in rice through integration of soil chemistry and plant physiology research. Plant Soil 2012, 361, 3–41. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management, 7th ed.; Pearson Education: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Wu, S.C.; Cheung, K.C.; Luo, Y.M.; Wong, M.H. Effects of inoculation of plant growth-promoting rhizobacteria on metal uptake by Brassica juncea. Environ. Pollut. 2006, 140, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Fasim, F.; Ahmed, N.; Parsons, R.; Gadd, G.M. Solubilization of zinc salts by a bacterium isolated from the air environment of a tannery. FEMS Microbiol. Lett. 2002, 213, 1–6. [Google Scholar] [CrossRef]
- Saravanan, V.S.; Subramoniam, S.R.; Raj, S.A. Assessing in vitro solubilization potential of different zinc solubilizing bacterial (zsb) isolates. Brazilian J. Microbiol. 2004, 35, 121–125. [Google Scholar] [CrossRef]
- Martino, E.; Perotto, S.; Parsons, R.; Gadd, G.M. Solubilization of insoluble inorganic zinc compounds by ericoid mycorrhizal fungi derived from heavy metal polluted sites. Soil Biol. Biochem. 2003, 35, 133–141. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Jayalakshmi, K.; Ramachandran, V.T. Role of arbuscular mycorrhizal fungus (Glomus intraradices) (fungus aided) in zinc nutrition of maize. J. Agric. Biotechnol. Sustain. Dev. 2009, 1, 29–38. [Google Scholar]
- Obrador, A.; Novillo, J.; Alvarez, J.M. Mobility and availability to plants of two zinc sources applied to a calcareous Soil. Soil Sci. Soc. Am. J. 2003, 67, 564–572. [Google Scholar] [CrossRef]
- Whiting, S.N.; de Souza, M.P.; Terry, N. Rhizosphere bacteria mobilize Zn for hyperaccumulation by Thlaspicaerulescens. Environ. Sci. Technol. 2001, 35, 3144–3150. [Google Scholar] [CrossRef]
- The World Food Prize- Improving The Quality, Quantity and Availability of Food in the World. Available online: https://www.worldfoodprize.org/en/laureates/2010__2017_laureates/2016__andrade_mwanga_low_and_bouis/ (accessed on 29 September 2019).
- Giuliano, G. Provitamin A biofortification of crop plants: A gold rush with many miners. Curr. Opin. Biotechnol. 2017, 44, 169–180. [Google Scholar] [CrossRef]
- Ye, X.; Al-babili, S.; Klӧti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–306. [Google Scholar] [CrossRef]
- Golden Rice- International Rice Research Institute. Available online: https://www.irri.org/golden-rice (accessed on 29 September 2019).
- Roje, S. Vitamin B biosynthesis in plants. Phytochemistry 2007, 68, 1904–1921. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Conklin, P.L.; Loewus, F.A. Biosynthesis of ascorbic acid in plants: A renaissance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 437–467. [Google Scholar] [CrossRef] [PubMed]
- Mozafar, A. Enrichment of some B-vitamins in plants with application of organic fertilizers. Plant Soil 1994, 167, 305–311. [Google Scholar] [CrossRef]
- Goyer, A. Thiamin biofortification of crops. Curr. Opin. Biotechnol. 2017, 44, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Moulin, M.; Fitzpatrick, T.B. Examining strategies to facilitate vitamin B 1 biofortification of plants by genetic engineering. Front. Plant Sci. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Thomas, N.; Ronald, P.C.; Goyer, A. Overexpression of thiamin biosynthesis genes in rice increases leaf and unpolished grain thiamin content but not resistance to Xanthomonas oryzae pv. oryzae. Front. Plant Sci. 2016, 7, 616. [Google Scholar] [CrossRef] [PubMed]
- Antoniani, C.; Monzini, A. The vitamin content of produce and products on the farm. III. Vitamin B i content of fodder from meadows irrigated by sewage as compared with that irrigated by clean water. Ann. Sper. Agrar. 1950, 4, 625–630. [Google Scholar]
- Wilberg, E. Über die Qualitfit von Spinat aus “biologischem Anban”. Landwirtsch. Forsch 1972, 25, 167–169. [Google Scholar]
- Singh, R.D.; Dhar, N.R. Effect of organic matter without and with rockphosphate on crop yield, quality and soil characteristics. Indian J. Agric. Sci. 1986, 56, 539–543. [Google Scholar]
- Mozafar, A.; Oertli, J.J. Uptake of a microbially-produced vitamin (B12) by soybean roots. Plant Soil 1992, 1, 23–30. [Google Scholar] [CrossRef]
- Gray, L.F.; Daniel, L.J. Studies of vitamin B12 in turnip greens. J. Nutr. 1959, 10, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Robbins, W.J.; Hervey, A.; Stebbins, M.E. Studies on euglena and Vitamin B12. Bull. Torrey Bot. 1950, 77, 423–441. [Google Scholar] [CrossRef]
- Chadare, F.J.; Idohou, R.; Nago, E.; Affonfere, M.; Hounhouigan, D.J.; Agossadou, J.; Kévin, T.; Christel, F.; Sewanou, K.; Azokpota, P.; et al. Conventional and food-to-food fortification: An appraisal of past practices and lessons learned. Food Sci. Nutr. 2019, 7, 2781–2795. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Cimini, S.; De Gara, L. Strategies to increase vitamin C in plants: From plant defense perspective to food biofortification. Front. Plant Sci. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Olowokere, F.A. Effect of inorganic and organo-mineral fertilizers on soil properties and nutrient composition of pepper (Capsicum spp.). J. Agric. Sci. Environ. 2014, 14, 82–96. [Google Scholar]
- Zhang, C.; Liu, J.; Zhang, Y. Overexpression of SlGMEs leads to ascorbate accumulation with enhanced oxidative stress, cold, and salt tolerance in tomato. Plant Cell Rep. 2011, 30, 389–398. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Celador-Lera, L.; Fradejas-Bayón, M.; Rivas, R. Plant probiotic bacteria enhance the quality of fruit and horticultural crops. AIMS Microbiol. 2017, 3, 483–501. [Google Scholar]
- Baslam, M.; Garmendia, I.; Goicoechea, N. Enhanced accumulation of vitamins, nutraceuticals and minerals in lettuces associated with arbuscular mycorrhizal fungi (AMF): A question of interest for both vegetables and humans. Agriculture 2013, 3, 188–209. [Google Scholar] [CrossRef]
- Kaushik, B.D.; Venkatarman, G.S. Effect of algal inoculation on the yield and vitamin C content of two varieties of tomato. Plant Soil 1979, 52, 135–137. [Google Scholar] [CrossRef]
- Prlak, L.; Köse, M. Effects of plant growth promoting Rhizobacteria on yield and some fruit properties of strawberry. J. Plant Nutr. 2009, 32. [Google Scholar] [CrossRef]
- Birt, D.F. Flavonoids. Adv. Nutr. 2013, 4, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cruz, S.; Chaparro-Hernández, S.; Hernández-Ruiz, K.L.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Gassos Ortega, L.E.; Ornelas-Paz, J.d.J.; Lopez Mata, M.A. Flavonoids: Important biocompounds in food. In Flavonoids—from Bosynthesis To Human Health; IntechOpen: London, UK, 2017; pp. 353–369. [Google Scholar]
- Scarano, A.; Chieppa, M.; Santino, A. Looking at flavonoid biodiversity in horticultural crops: A colored mine with nutritional benefits. Plants 2018, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Bovy, A.; De Vos, R.; Kemper, M.; Schijlen, E.; Pertejo, A.; Muir, S.; Collins, G.; Robinson, S.; Verhoeyen, M.; Hughes, S.; et al. High-flavonol tomatoes resulting from the heterologous expression of the maize transcription factor genes LC and C1. Plant Cell 2002, 14, 2509–2526. [Google Scholar] [CrossRef] [PubMed]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Güçlü-Üstündağ, Ö.; Mazza, G. Saponins: Properties, applications and processing. Crit. Rev. Food Sci. Nutr. 2007, 47, 231–258. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.; Kang, S.; Kang, H.; Oh, H.; Bae, C. Hypocholesterolemic property of Yucca schidigera and Quillaja saponaria extracts in human body. Arch. Pharm. Res. 2003, 26, 1042–1046. [Google Scholar] [CrossRef]
- Huang, Z.; Lin, J.; Cheng, Z.; Xu, M.; Guo, M.; Huang, X.; Yang, Z. Production of oleanane-type sapogenin in transgenic rice via expression of β-amyrin synthase gene from Panax japonicus C. A. Mey. BMC Biotechnol. 2015, 15, 45. [Google Scholar] [CrossRef]
- Wu, H.; Yang, H.; You, X.; Li, Y. Diversity of endophytic fungi from roots of Panax ginseng and their saponin yield capacities. Springerplus 2013, 2, 107. [Google Scholar] [CrossRef]
- Microbial Root Endophytes; Schulz, B., Boyle, C., Sieber, T., Eds.; Springer: New York, NY, USA, 2006; ISBN 9783540335252. [Google Scholar]
- Bacon, C.W.; White, J.F., Jr. Functions, mechanisms and regulation of endophytic and epiphytic microbial communities of plants. Symbiosis 2016, 68, 87–98. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef]
- Luginbuehl, L.H.; Menard, G.N.; Kurup, S.; Van Erp, H.; Radhakrishnan, G.V.; Breakspear, A.; Oldroyd, G.E.D.; Eastmond, P.J. Fatty acids in arbuscular mycorrhizal fungi are synthesized by the host plant. Science 2017, 356, 1175–1178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Blaylock, L.A.; Harrison, M.J. Two Medicago truncatula half-ABC transporters are essential for arbuscule development in arbuscular mycorrhizal symbiosis. Plant Cell 2010, 22, 1483–1497. [Google Scholar] [CrossRef] [PubMed]
- Fourcroy, P.; Siso-Terraza, P.; Sudre, D.; Saviron, M.; Reyt, G.; Gaymard, F.; Abadia, A.; Abadia, J.; Alvarez-Fernandez, A.; Briat, J.F. Involvement of the ABCG37 transporter in secretion of scopoletin and derivatives by Arabidopsis roots in response to iron deficiency. New Phytol. 2014, 201, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Commoner, B. Cost-risk-benefit analysis of nitrogen fertilization: A case history. Ambio 1977, 6, 157–161. [Google Scholar]
- Carlsson, G.; Huss-Danell, K. Nitrogen fixation in perennial forage legumes in the field. Plant Soil 2003, 253, 353–372. [Google Scholar] [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Alves, B.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; Sampet, C.; Rerkasem, B.; et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Hestrin, R.; Hammer, E.C.; Mueller, C.W.; Lehmann, J. Synergies between mycorrhizal fungi and soil microbial communities increase plant nitrogen acquisition. Commun. Biol. 2019, 2, 233. [Google Scholar] [CrossRef]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science 2012, 336, 1576–1578. [Google Scholar] [CrossRef]
- Singh, D.; Rajawat, M.V.S.; Kaushik, R.; Prasanna, R.; Saxena, A.K. Beneficial role of endophytes in biofortification of Zn in wheat genotypes varying in nutrient use efficiency grown in soils sufficient and deficient in Zn. Plant Soil 2017, 416, 107–116. [Google Scholar] [CrossRef]
- Bharadwaj, D.P.; Alstrom, S.; Lundquist, P.O. Interactions among Glomus irregulare, arbuscular mycorrhizal spore-associated bacteria, and plant pathogens under in vitro conditions. Mycorrhiza 2012, 22, 437–447. [Google Scholar] [CrossRef]
- Saridis, G.; Chorianopoulou, S.N.; Ventouris, Y.E.; Sigalas, P.P.; Bouranis, D.L. An Exploration of the Roles of Ferric Iron Chelation-Strategy Components in the Leaves and Roots of Maize Plants. Plants (Basel, Switzerland) 2019, 8, 133. [Google Scholar] [CrossRef] [PubMed]
- Chorianopoulou, S.N.; Saridis, Y.I.; Dimou, M.; Katinakis, P.; Bouranis, D.L. Arbuscular mycorrhizal symbiosis alters the expression patterns of three key iron homeostasis genes, ZmNAS1, ZmNAS3, and ZmYS1, in S deprived maize plants. Front. Plant Sci. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Garrido-Oter, R.; Gonzalez, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and host nutrition across plant and animal kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [PubMed]
- El Zahar Haichar, F.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Samain, E.; van Tuinen, D.; Jeandet, P.; Aussenac, T.; Selim, S. Biological control of septoria leaf blotch and growth promotion in wheat by Paenibacillus sp. strain B2 and Curtobacterium plantarum strain EDS. Biol. Control 2017, 114, 87–96. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Knee, E.M.; Gong, F.C.; Gao, M.; Teplitski, M.; Jones, A.R.; Foxworthy, A.; Mort, A.J.; Bauer, W.D. Root mucilage from pea and its utilization by rhizosphere bacteria as a sole carbon source. Mol. Plant Microbe Interact. 2001, 14, 775–784. [Google Scholar] [CrossRef]
- Hennion, N.; Durand, M.; Vriet, C.; Doidy, J.; Maurousset, L.; Lemoine, R.; Pourtau, N. Sugars en route to the roots. Transport, metabolism and storage within plant roots and towards microorganisms of the rhizosphere. Physiol. Plant 2019, 165, 44–57. [Google Scholar] [CrossRef]
- Jaeger, C.H., 3rd; Lindow, S.E.; Miller, W.; Clark, E.; Firestone, M.K. Mapping of sugar and amino acid availability in soil around roots with bacterial sensors of sucrose and tryptophan. Appl. Environ. Microbiol. 1999, 65, 2685–2690. [Google Scholar]
- Carvalhais, L.C.; Dennis, P.G.; Fedoseyenko, D.; Hajirezaei, M.-R.; Borriss, R.; von Wirén, N. Root exudation of sugars, amino acids, and organic acids by maize as affected by nitrogen, phosphorus, potassium, and iron deficiency. J. Plant Nutr. Soil Sci. 2011, 174, 3–11. [Google Scholar] [CrossRef]
- Flugge, U.I.; Hausler, R.E.; Ludewig, F.; Gierth, M. The role of transporters in supplying energy to plant plastids. J. Exp. Bot. 2011, 62, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Allard-Massicotte, R.; Tessier, L.; Lecuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involves multiple chemotaxis receptors. MBio 2016, 7, 1616–1664. [Google Scholar] [CrossRef] [PubMed]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef]
- Xu, W.H.; Wang, H.X.; Liu, H.; Xiong, Z.T.; Singh, B. Sigle and combined pollution of zinc and Cadmium influence on root exudates and Zn, Cd forms in rygrass. Acta Sci. Circumstantiae 2007, 28, 2089–2095. [Google Scholar]
- Ahmadzadeh, F.; Khoshgoftarmanesh, A.H. Release of phytosiderophores from roots of wheat and triticale under nickel-deficient conditions. J. Plant Nutr. Soil Sci. 2019, 182, 708–714. [Google Scholar] [CrossRef]
- Gillissen, B.; Burkle, L.; Andre, B.; Kuhn, C.; Rentsch, D.; Brandl, B.; Frommer, W.B. A new family of high-affinity transporters for adenine, cytosine, and purine derivatives in Arabidopsis. Plant Cell 2000, 12, 291–300. [Google Scholar] [CrossRef]
- Desimone, M.; Catoni, E.; Ludewig, U.; Hilpert, M.; Schneider, A.; Kunze, R.; Tegeder, M.; Frommer, W.B.; Schumacher, K. A novel superfamily of transporters for allantoin and other oxo derivatives of nitrogen heterocyclic compounds in Arabidopsis. Plant Cell 2002, 14, 847–856. [Google Scholar] [CrossRef][Green Version]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Dennis, P.G.; Fan, B.; Fedoseyenko, D.; Kierul, K.; Becker, A.; von Wiren, N.; Borriss, R. Linking plant nutritional status to plant-microbe interactions. PLoS ONE 2013, 8, e68555. [Google Scholar] [CrossRef]
- Ziegler, J.; Schmidt, S.; Chutia, R.; Muller, J.; Bottcher, C.; Strehmel, N.; Scheel, D.; Abel, S. Non-targeted profiling of semi-polar metabolites in Arabidopsis root exudates uncovers a role for coumarin secretion and lignification during the local response to phosphate limitation. J. Exp. Bot. 2016, 67, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Hirano, Y.; Brunner, I. Exudation of organic acid anions from poplar roots after exposure to Al, Cu and Zn. Tree Physiol. 2007, 27, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, D.S.; Teixeira, P.J.P.L. Root-exuded coumarin shapes the root microbiome. Proc. Natl. Acad. Sci. USA 2018, 115, 5629. [Google Scholar] [CrossRef]
- Micallef, S.A.; Shiaris, M.P.; Colon-Carmona, A. Influence of Arabidopsis thaliana accessions on rhizobacterial communities and natural variation in root exudates. J. Exp. Bot. 2009, 60, 1729–1742. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Garrido-Oter, R.; Munch, P.C.; Weiman, A.; Droge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef]
- Van Nuland, M.E.; Ware, I.M.; Bailey, J.K.; Schweitzer, J.A. Ecosystem feedbacks contribute to geographic variation in plant–soil eco-evolutionary dynamics across a fertility gradient. Funct. Ecol. 2019, 33, 95–106. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Correction: Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time. Isme J. 2017, 11, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.M.; Ji, P.; Firestone, M.K.; Lindow, S.E. Two novel bacterial biosensors for detection of nitrate availability in the rhizosphere. Appl. Environ. Microbiol. 2005, 71, 8537–8547. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, K.M.; Brodie, E.L.; DeSantis, T.Z.; Andersen, G.L.; Lindow, S.E.; Firestone, M.K. Selective progressive response of soil microbial community to wild oat roots. Isme J. 2009, 3, 168–178. [Google Scholar] [CrossRef]
- Massalha, H.; Korenblum, E.; Malitsky, S.; Shapiro, O.H.; Aharoni, A. Live imaging of root-bacteria interactions in a microfluidics setup. Proc. Natl. Acad. Sci. USA 2017, 114, 4549–4554. [Google Scholar] [CrossRef]
- Kawasaki, A.; Donn, S.; Ryan, P.R.; Mathesius, U.; Devilla, R.; Jones, A.; Watt, M. Microbiome and exudates of the root and rhizosphere of Brachypodium distachyon, a model for wheat. PLoS ONE 2016, 11, e0164533. [Google Scholar] [CrossRef]
- Watson, B.S.; Bedair, M.F.; Urbanczyk-Wochniak, E.; Huhman, D.V.; Yang, D.S.; Allen, S.N.; Li, W.; Tang, Y.; Sumner, L.W. Integrated metabolomics and transcriptomics reveal enhanced specialized metabolism in Medicago truncatula root border cells. Plant Physiol. 2015, 167, 1699–1716. [Google Scholar] [CrossRef]
- Vicre, M.; Santaella, C.; Blanchet, S.; Gateau, A.; Driouich, A. Root border-like cells of Arabidopsis. Microscopical characterization and role in the interaction with rhizobacteria. Plant Physiol. 2005, 138, 998–1008. [Google Scholar] [CrossRef]
- Neumann, G.; Martinoia, E. Cluster roots—an underground adaptation for survival in extreme environments. Trends Plant Sci. 2002, 7, 162–167. [Google Scholar] [CrossRef]
- Weisskopf, L.; Heller, S.; Eberl, L. Burkholderia species are major inhabitants of white lupin cluster roots. Appl. Environ. Microbiol. 2011, 77, 7715–7720. [Google Scholar] [CrossRef]
- Weisskopf, L.; Abou-Mansour, E.; Fromin, N.; Tomasi, N.; Santelia, D.; Edelkott, I.; Neumann, G.; Aragno, M.; Tabacchi, R.; Martinoia, E. White lupin has developed a complex strategy to limit microbial degradation of secreted citrate required for phosphate acquisition. Plant Cell Environ. 2006, 29, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Dobies, T.; Cesarz, S.; Hobbie, S.E.; Meyer, R.J.; Worm, K.; Reich, P.B. Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment. Proc. Natl. Acad. Sci. USA 2013, 110, 6889–6894. [Google Scholar] [CrossRef] [PubMed]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root exudate cocktails: The link between plant diversity and soil microorganisms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets-iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]


{kind=link}
| Crop | Microbe | Type of microbe | Tissue | Impact | Reference |
|---|---|---|---|---|---|
| Blackberry | Pseudomonas fluorescens N21.4 | Rhizospheric bacterium | Fruit | Improved flavonoid level | [14,15] |
| Brassica juncea | Bacillus toyonensis (MG430287), Rhodococcus hoagii (MG432495), Lysinibacillus mangiferihumi (MG432492), and Lysinibacillus fusiformis (MG430290) | Rhizospheric bacteria | Root, stem, and leaf | Increased Fe contents | [16] |
| Chickpea and pigeonpea | Pseudomonas plecoglossicida SRI-156, Brevibacterium antiquum SRI-158, Bacillus altitudinis SRI-178, Enterobacter ludwigii SRI-211, E. ludwigii SRI-229, Acinetobacter tandoii SRI-305, and Pseudomonas monteilii SRI-360 | Rhizospheric bacteria | Grain | Improved contents of Fe (up to 18% and 12%), Zn (up to 23% and 5%), Cu (up to 19% and 8%), Mn (up to 2% and 39%) and Ca (up to 22% and 11%) | [17] |
| Chickpea | Funneliformis mosseae, Rhizophagus irregularis | Arbuscular mycorrhizal fungi | Grain | Increased protein, Fe, and Zn contents | [18] |
| Isolates of actinobacteria | Rhizospheric bacteria | Seeds | Increased Fe (10%–38%), Zn (13%–30%), Ca (14%–26%), Cu (11%–54%) and Mn (18%–35%) and Mg (14%–21%) | [19] | |
| Lettuce | Glomus fasciculatum | Arbuscular mycorrhizal fungus | Leaf | Improved ascorbate level and fresh weight | [20] |
| Glomus intraradices (Schenck and Smith) and Glomus mosseae [(Nicol. and Gerd.) Gerd. and Trappe] | Arbuscular mycorrhizal fungi | Leaf | Improved ascorbate level and fresh weight | [20] | |
| Oil palm | Hendersonia toruloidea | Endophytic fungus | Leaf | Improvement of thiamine biosynthesis | [21,22] |
| Okra | Azotobacter sp. and cyanobacterium (e.g., Calothrix sp.) | Rhizospheric bacteria | Fruit | Increased concentration of Zn 60%–70% | [23] |
| Onion | Rhizophagus irregularis | Arbuscular mycorrhizal fungus | Bulb | Improved biomass production, the abundance of vitamin B1 and its analogues, and organic acids concentration | [24] |
| Pak choi | Funneliformis mosseae, Glomus versiforme, and Rhizophagus intraradices | Arbuscular mycorrhizal fungi | Shoot | Reduced concentrations of Pb (by 20.6%–67.5%) and Cd (by 14.3%–54.1%) in shoots | [25] |
| Potato | Glomus irregulare | Arbuscular mycorrhizal fungus | Roots and tubers | Controlled the growth of Fusarium sambucinum and inhibited the production of the mycotoxin trichothecene 4, 15-diacetoxyscirpenol | [26] |
| Rice | Bacillus sp. SH-10 and B. cereus SH-17 | Rhizospheric bacteria | Grain | Increased Zn concentration in grain (to ~31 from ~18 mg kg−1 in control) | [27] |
| Cyanobacteria-based inoculants | Rhizospheric bacteria | Grain | 13%–46% enhancement in iron and 15%-41% enhancement in zinc | [28] | |
| Enterobacter sp. ScCS20 | Rhizospheric bacterium | Grain | Improved Zn level | [29] | |
| Sphingomonas sp. SaMR12 | Rhizospheric bacterium | Grain | Improved Zn level | [29] | |
| Rice-wheat cropping system | Anabaena oscillarioides, Brevundimonas diminuta, and Ochrobactrum anthropi | Rhizospheric bacteria | Grain | Micronutrient enrichment: Fe, Zn in rice, and Cu, Mn in wheat. Increased N-P-K contents and improved rice yield by 21.2% (compared to using the recommended dosage of chemical fertilizers) | [30] |
| Safed musli | Glomus fasciculatum, Glomus intraradices, or Glomus mosseae | Arbuscular mycorrhizal fungi | Tuber | Improved saponin level and improved biomass of roots and shoots | [31] |
| Piriformospora indica and Pseudomonas fluorescens | Arbuscular mycorrhizal fungus and rhizospheric bacterium respectively | Micropropagated plants | P. indica enhanced the level of saponin in the plants. The co-inoculation of P. indica and P. fluorescens further enhanced the saponin level and also the contents of micronutrients including Fe, Zn, and Mn. | [32] | |
| Snap bean | Rhizobium etli and Glomus intraradices | Rhizospheric bacterium and arbuscular mycorrhizal fungus | Seed | Increased yield, number of snap beans, and percentage of protein after N fertilization | [33] |
| Spinach | Glomus fasciculatum, Glomus mosseae, N-fixer (Azotobacter chroococcum), K solubilizer (Bacillus mucilaginous), and P solubilizer (Bacillus megaterium) | Arbuscular mycorrhizal fungi and rhizospheric bacteria | Leaf | Augmented the concentrations of total phenolic compounds, flavonoids, and phenolic acids | [34] |
| Strawberry | Rhizophagus intraradices, Glomus aggregatum, Glomus viscosum, Claroideoglomus etunicatum, and Claroideoglomus claroideum, and Pseudomonas fluorescens strain Pf4 (Pf4) and Pseudomonas sp. 5Vm1K (5Vm1K) | Arbuscular mycorrhizal fungi and Rhizospheric bacteria | Fruit | Co-inoculation increased fruit production, fruit size, and concentrations of sugars and ascorbic and folic acids | [35] |
| Phyllobacterium endophyticum PEPV15 | Rhizospheric bacterium | Fruit | Improved vitamin C level and yield | [36] | |
| Paenibacillus polymyxa RC05 | Rhizospheric bacterium | Fruit | Improved vitamin C level and yield | [37] | |
| Bacillus simplex RC19 | Rhizospheric bacterium | Fruit | Improved vitamin C level and yield | [38] | |
| Pseudomonas sp. 5Vm1K | Rhizospheric bacterium | Fruit | Improved vitamin C level | [35] | |
| Tomato | Rhizophagus irregularis, Funneliformis mosseae | Arbuscular mycorrhizal fungus | Fruit | Increased N, P, and Cu concentrations, higher antioxidant concentrations and carotenoid contents | [39] |
| Pseudomonas sp. 19Fv1T | Rhizospheric bacterium | Fruit | Improved vitamin C level and fructose level | [38] | |
| Bacillus licheniformis | Rhizospheric bacterium | Fruit | Improved flavonoid level | [40] | |
| Wheat | Pseudomonas jessenii (R62) and Pseudomonas synxantha (R81) and AMF consortium (Mnat) and (Mss2) | Rhizospheric bacteria and arbuscular mycorrhizal fungi | Grain | Significantly improved selenium content | [41] |
| Pseudomonas strain (Providencia sp. PW5) and Anabaena sp., Calothrix sp. and Anabaena sp. | Rhizospheric bacteria and arbuscular mycorrhizal fungi | Grain | increased wheat yield and mineral nutrient concentrations of P, K, Cu, Fe, Zn, and Mn | [41] | |
| Providencia sp. combined with N60P60K60 chemical fertilizer | Rhizospheric bacterium | Grain | Increases grain Zn accumulation to ~42 mg kg−1compared to 31.60 mg kg−1 in control, along with a three-fold increase in the concentration of Fe. | [42] | |
| Bacillus cereus YAP6 and Bacillus licheniformis YAP7 | Rhizospheric bacteria | Grain | Increased Se concentrations in the grain by up to 375% | [43] | |
| Eight AMF species (Gigaspora margarita, Funneliformis mosseae, Rhizophagus irregulare, Glomus clarum, Glomus deserticola, Glomus monosporum, Glomus brasilianum, Glomus aggregatum) | Arbuscular mycorrhizal fungus | Root | increased the uptake of P, Fe, and Zn with improved root lengths and density | [44] | |
| Glomus clarideum and Pseudomonas sp. R8 | Arbuscular mycorrhizal fungus and rhizospheric bacterium, respectively | Grain | Improved selenium level | [45] | |
| Glomus clarideum and Bacillus sp. R12 | Arbuscular mycorrhizal fungus and rhizospheric bacterium, respectively | Grain | Improved selenium level | [45] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, Y.-S.; Rehman, H.M.; Lam, H.-M. Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification. Agronomy 2019, 9, 764. https://doi.org/10.3390/agronomy9110764
Ku Y-S, Rehman HM, Lam H-M. Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification. Agronomy. 2019; 9(11):764. https://doi.org/10.3390/agronomy9110764
Chicago/Turabian StyleKu, Yee-Shan, Hafiz Mamoon Rehman, and Hon-Ming Lam. 2019. "Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification" Agronomy 9, no. 11: 764. https://doi.org/10.3390/agronomy9110764
APA StyleKu, Y.-S., Rehman, H. M., & Lam, H.-M. (2019). Possible Roles of Rhizospheric and Endophytic Microbes to Provide a Safe and Affordable Means of Crop Biofortification. Agronomy, 9(11), 764. https://doi.org/10.3390/agronomy9110764