
Bacterial Community Dynamics in Oil-Contaminated Soils in the Hyper-Arid Arava Valley

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling
2.3. Soil Moisture (SM) Content and Electrical Conductivity (EC)
2.4. Organic Matter (OM) Content
2.5. Soil pH
2.6. Microbial Community Diversity Determination
2.7. Data Analysis
3. Results
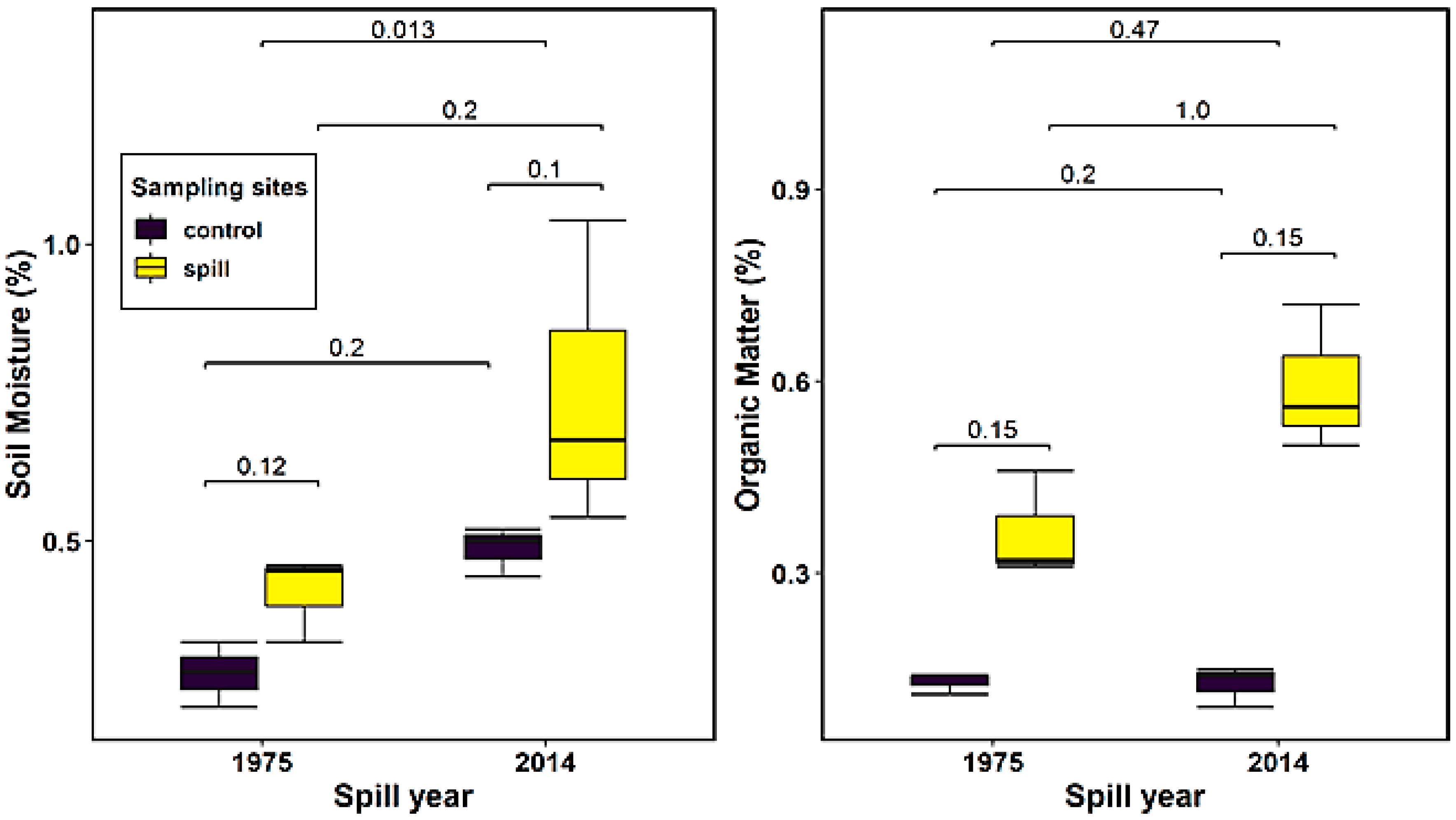
3.1. Soil Physicochemical Properties
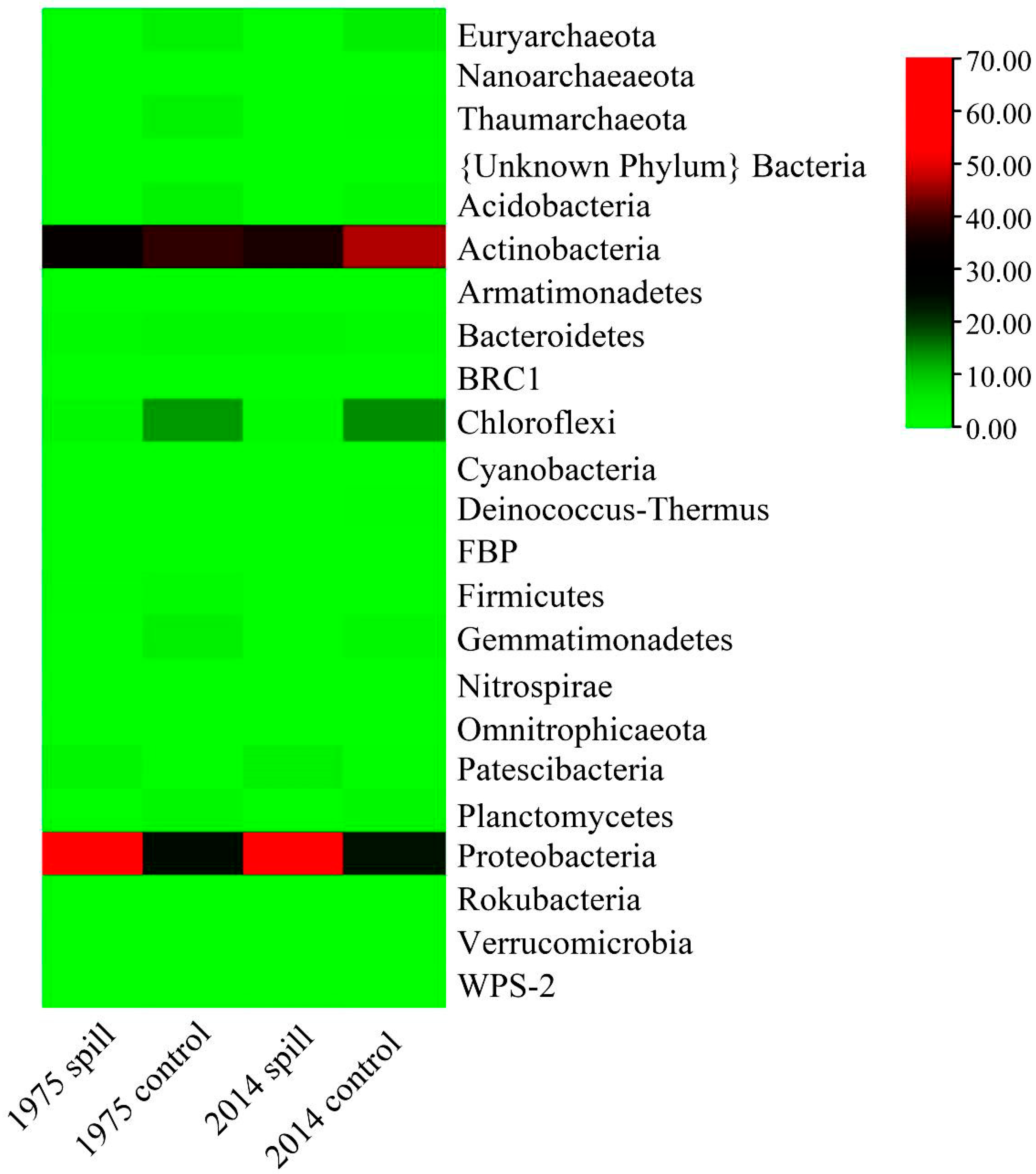
3.2. Miseq Sequencing Results and Bacterial Community Structures
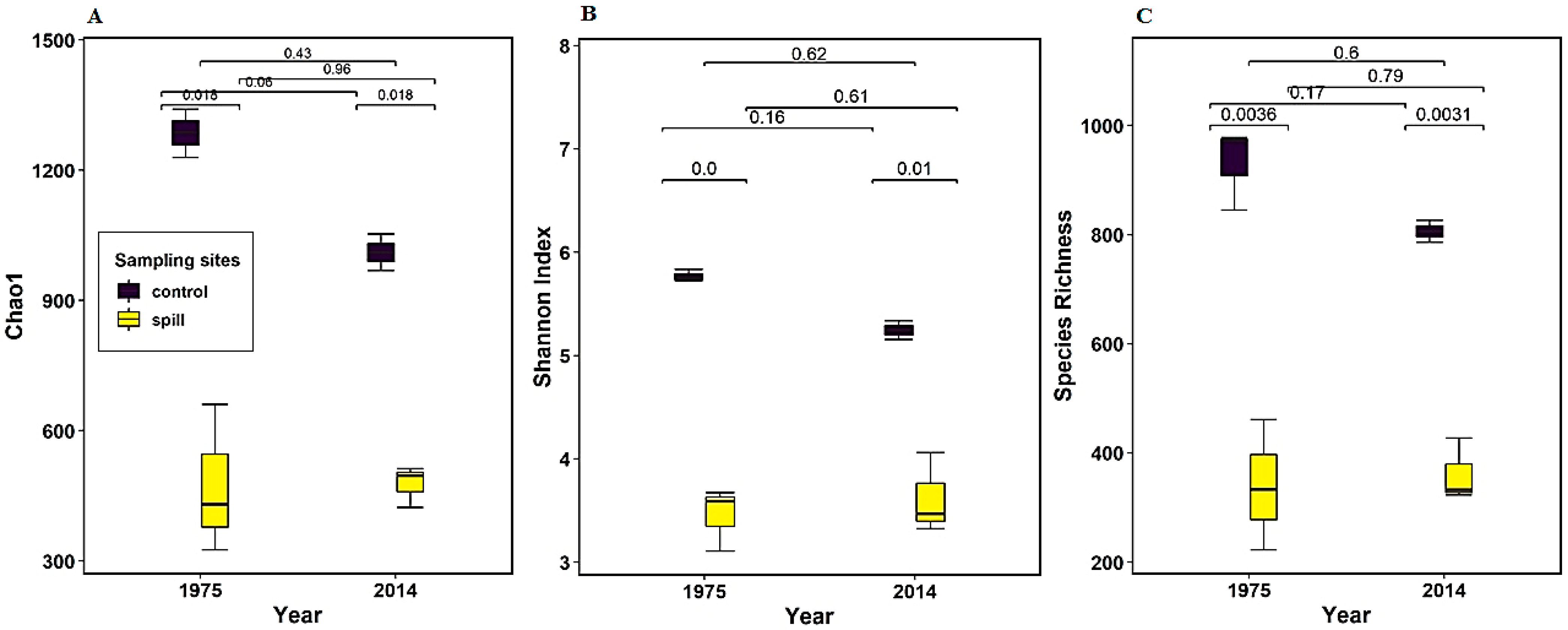
3.3. Alpha Diversity Within Samples and Rarefaction Curves
3.4. Beta Diversity (Pairwise Sample Dissimilarity) Among Samples, and Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuppusamy, S.; Maddela, N.R.; Megharaj, M.; Venkateswarlu, K. Impact of Total Petroleum Hydrocarbons on Human Health. In Total Petroleum Hydrocarbons; Springer: Cham, Switzerland, 2020; pp. 139–165. [Google Scholar] [CrossRef]
- Tedesco, P.; Balzano, S.; Coppola, D.; Esposito, F.P.; de Pascale, D.; Denaro, R. Bioremediation for the recovery of oil polluted marine environment, opportunities and challenges approaching the Blue Growth. Mar. Pollut. Bull. 2024, 200, 116157. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, C.; Xin, Y.; Huang, T.; Liu, J. Petroleum pollution affects soil chemistry and reshapes the diversity and networks of microbial communities. Ecotoxicol. Environ. Saf. 2022, 246, 114129. [Google Scholar] [CrossRef]
- Sui, X.; Wang, X.; Li, Y.; Ji, H. Remediation of Petroleum-Contaminated Soils with Microbial and Microbial Combined Methods: Advances, Mechanisms, and Challenges. Sustainability 2021, 13, 9267. [Google Scholar] [CrossRef]
- Oruabena, B.; Elechi, O. Clean-up of crude oil-contaminated soils: Bioremediation option. Int. J. Environ. Sci. Technol. 2019, 17, 1185–1198. [Google Scholar]
- Maciejczyk, M.; Tyrpień-Golder, K.; Janoszka, B.; Gierat, B.; Muzyka, R. Mutagenic and carcinogenic polycyclic aromatic hydrocarbons (PAHs) in food—Occurrence, human health effects, and assessment methods of exposure. Environ. Med. 2023, 26, 8–15. [Google Scholar] [CrossRef]
- Elgazali, A.; Althalb, H.; Elmusrati, I.; Ahmed, H.M.; Banat, I.M. Remediation Approaches to Reduce Hydrocarbon Contamination in Petroleum-Polluted Soil. Microorganisms 2023, 11, 2577. [Google Scholar] [CrossRef]
- Adedeji, J.A.; Tetteh, E.K.; Amankwa, M.O.; Asante-Sackey, D.; Ofori-Frimpong, S.; Armah, E.K.; Rathilal, S.; Mohammadi, A.H.; Chetty, M. Microbial Bioremediation and Biodegradation of Petroleum Products—A Mini Review. Appl. Sci. 2022, 12, 12212. [Google Scholar] [CrossRef]
- Cai, P.; Ning, Z.; Liu, Y.; He, Z.; Shi, J.; Niu, M. Diagnosing bioremediation of crude oil-contaminated soil and related geochemical processes at the field scale through microbial community and functional genes. Ann. Microbiol. 2020, 70, 36. [Google Scholar] [CrossRef]
- Chen, G.; Yuan, M.; Ma, B.; Ren, Y. Responses of Petroleum Contamination at Different Sites to Soil Physicochemical Properties and Indigenous Microbial Communities. Water Air Soil Pollut. 2023, 234, 494. [Google Scholar] [CrossRef]
- Borah, G.; Deka, H. Crude oil associated heavy metals (HMs) contamination in agricultural land: Understanding risk factors and changes in soil biological properties. Chemosphere 2022, 310, 136890. [Google Scholar] [CrossRef]
- Ullah, Z.; Rashid, A.; Ghani, J.; Nawab, J.; Zeng, X.-C.; Shah, M.; Alrefaei, A.F.; Kamel, M.; Aleya, L.; Abdel-Daim, M.M.; et al. Groundwater contamination through potentially harmful metals and its implications in groundwater management. Front. Environ. Sci. 2022, 10, 1021596. [Google Scholar] [CrossRef]
- McKew, B.A.; Coulon, F.; Yakimov, M.M.; Denaro, R.; Genovese, M.; Smith, C.J.; Osborn, A.M.; Timmis, K.N.; McGenity, T.J. Efficacy of intervention strategies for bioremediation of crude oil in marine systems and effects on indigenous hydrocarbonoclastic bacteria. Environ. Microbiol. 2007, 9, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Kebede, G.; Tafese, T.; Abda, E.M.; Kamaraj, M.; Assefa, F. Factors influencing the bacterial bioremediation of hydrocarbon contaminants in the soil: Mechanisms and impacts. J. Chem. 2021, 2021, 9823362. [Google Scholar] [CrossRef]
- Shi, L.; Liu, Z.; Yang, L.; Fan, W. Effects of oil pollution on soil microbial diversity in the Loess hilly areas, China. Ann. Microbiol. 2022, 72, 26. [Google Scholar] [CrossRef]
- Maestre, F.T.; Eldridge, D.J.; Soliveres, S.; Kéfi, S.; Delgado-Baquerizo, M.; Bowker, M.A.; García-Palacios, P.; Gaitán, J.; Gallardo, A.; Lázaro, R.; et al. Structure and functioning of dryland ecosystems in a changing world. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 215–237. [Google Scholar] [CrossRef]
- Abbasian, F.; Lockington, R.; Megharaj, M.; Naidu, R. The Biodiversity Changes in the Microbial Population of Soils Contaminated with Crude Oil. Curr. Microbiol. 2016, 72, 663–670. [Google Scholar] [CrossRef]
- Paul, S.; Ingti, B.; Das, D.; Joshi, S.R. Microbial Responses to Hydrocarbon Contamination and Its Bioremediation. In Mineral Transformation and Bioremediation by Geo-Microbes; Joshi, S.R., Verma, P., Banerjee, S., Eds.; Interdisciplinary Biotechnological Advances; Springer: Singapore, 2025. [Google Scholar] [CrossRef]
- Widdel, F.; Knittel, K.; Galushko, A. Anaerobic Hydrocarbon-Degrading Microorganisms: An Overview. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Wang, W.; Tian, S.; Jiang, P.; Qi, Q.; Li, F.; Li, H.; Wang, Q.; Li, H.; et al. Potential biodegradation of phenanthrene by isolated halotolerant bacterial strains from petroleum oil polluted soil in Yellow River Delta. Sci. Total. Environ. 2019, 664, 1030–1038. [Google Scholar] [CrossRef]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. petroleum hydrocarbon contamination in terrestrial ecosystems—Fate and microbial responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Ye, J.; Jiang, K.; Wang, Y.; Li, Y. Oil contamination drives the transformation of soil microbial communities: Co-occurrence pattern, metabolic enzymes and culturable hydrocarbon-degrading bacteria. Ecotoxicol. Environ. Saf. 2021, 225, 112740. [Google Scholar] [CrossRef]
- Yerulker, G.; Patel, P.; Chafale, A.; Rathod, V.; Das, S.; Pandey, P.; Khan, N.A.; Devi, A.; Munshi, N.S.; Dhodapkar, R.; et al. Comparative assessment of soil microbial community in crude oil contaminated sites. Environ. Pollut. 2023, 328, 121578. [Google Scholar] [CrossRef]
- Fuentes, S.; Barra, B.; Caporaso, J.G.; Seeger, M. From rare to dominant: A fine-tuned soil bacterial bloom during petroleum hydrocarbon bioremediation. Appl. Environ. Microbiol. 2016, 82, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Gnansounou, E. Microbial dynamics in petroleum oilfields and their relationship with physiological properties of petroleum oil reservoirs. Bioresour. Technol. 2017, 245, 1258–1265. [Google Scholar] [CrossRef]
- Varjani, S.; Upasani, V.N.; Pandey, A. Bioremediation of oily sludge polluted soil employing a novel strain of Pseudomonas aeruginosa and phytotoxicity of petroleum hydrocarbons for seed germination. Sci. Total. Environ. 2020, 737, 139766. [Google Scholar] [CrossRef]
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Lee, K.; Greer, C.W. Chemical dispersants enhance the activity of oil- and gas condensate-degrading marine bacteria. ISME J. 2017, 11, 2793–2808. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhai, Z.; Li, H.; Wang, Q.; Han, X.; Yu, H. Synergetic effect of bio-photocatalytic hybrid system: G-C3N4 and Acinetobacter sp. JLS1 for enhanced degradation of C16 alkane. Chem. Eng. J. 2017, 323, 520–529. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Dane, J.H.; Topp, G.C. Methods of Soil Analysis. In Part 4: Physical Methods; Soil Science Society of America, Inc.: Madison, WI, USA, 2002. [Google Scholar]
- Rhoades, J.D. Salinity: Electrical Conductivity and Total Dissolved Solids. In Methods for Soil Analysis, Part 3: Chemical Methods; Sparks, R.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 417–435. [Google Scholar]
- Girsowicz, R.; Koryachenko, O.; Sherman, C.; Mayzlish-Gati, E.; Doniger, T.; Steinberger, Y. Impact of oil-spill contamination on a soil bacterial community: A 40-year history of rehabilitation in the arava valley. Soil Sediment Contam. Int. J. 2018, 27, 175–185. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Sys-Tematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–147. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Quail, M.A.; Smith, M.; Coupland, P.; Otto, T.D.; Harris, S.R.; Connor, T.R.; Bertoni, A.; Swerdlow, H.P.; Gu, Y. A tale of three next generation sequencing platforms: Comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq Sequencers. BMC Genom. 2012, 13, 341. [Google Scholar] [CrossRef]
- Lou, J.; Liu, Y.; Li, Y. Review of high-throughput sequencing techniques in studies of soil microbial diversity. Chin. Agri-Cult. Sci. Bull. 2014, 30, 256–260. [Google Scholar]
- Smith, D.P.; Peay, K.G. Sequence depth, not pcr replication, improves ecological inference from next generation DNA sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, C.; Duraipandiyan, V.; Balakrishna, K.; Ignacimuthu, S. Petroleum and polycyclic aromatic hydrocarbons (PAHs) degradation and naphthalene metabolism in Streptomyces sp. (ERI-CPDA-1) isolated from oil contaminated soil. Bioresour. Technol. 2012, 112, 83–90. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Q.; Xie, S. Bacterial and archaeal community structures in phenanthrene amended aquifer sediment microcosms under oxic and anoxic conditions. Int. J. Environ. Res. 2012, 6, 1077–1088. [Google Scholar]
- Grządziel, J.; Gałązka, A. Fungal biodiversity of the most common types of polish soil in a long-term microplot experiment. Front. Microbiol. 2019, 10, 6. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Zhang, Y.; Pan, Y.-R.; Li, L.; Liu, J.; Butler, D. Stepwise pH control to promote synergy of chemical and biological processes for augmenting short-chain fatty acid production from anaerobic sludge fermentation. Water Res. 2019, 155, 193–203. [Google Scholar] [CrossRef]
- Kocaoba, S. Adsorption of Fe(II) and Fe(III) from aqueous solution by using sepiolite: Speciation studies with MINEQL+ computer program. Sep. Sci. Technol. 2019, 55, 896–906. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Samira, A.; Omar, S.; Bhat, N.R.; Asem, A. Critical assessment of the environmental consequences of the invasion of Kuwait, the Gulf War, and the aftermath. In Environmental Consequences of War and Aftermath—The Handbook of Environmental Chemistry; Kassim, T.A., Barcelo, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 141–170. [Google Scholar]
- Kostka, J.E.; Prakash, O.; Overholt, W.A.; Green, S.J.; Freyer, G.; Canion, A.; Delgardio, J.; Norton, N.; Hazen, T.C.; Huettel, M. Hydrocarbon-degrading bacteria and the bacterial community response in gulf of mexico beach sands impacted by the deepwater horizon oil spill. Appl. Environ. Microbiol. 2011, 77, 7962–7974. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, Q.; Zhu, H.; Reich, P.B.; Banerjee, S.; A van der Heijden, M.G.; Sadowsky, M.J.; Ishii, S.; Jia, X.; Shao, M.; et al. Erosion reduces soil microbial diversity, network complexity and multifunctionality. ISME J. 2021, 15, 2474–2489. [Google Scholar] [CrossRef]
- Liang, Y.; Van Nostrand, J.D.; Deng, Y.; He, Z.; Wu, L.; Zhang, X.; Li, G.; Zhou, J. Functional gene diversity of soil microbial communities from five oil-contaminated fields in China. ISME J. 2010, 5, 403–413. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Moisture (%) | Organic Matter (%) | pH | Electrical Conductivity (µS cm−1) | |
|---|---|---|---|---|
| 1975 Control | 0.28 ± 0.04 | 0.13 ± 0.01 | 8.12 ± 0.11 | 111.60 ± 16.57 |
| 1975 Spill | 0.41 ± 0.06 | 0.28 ± 0.04 | 7.95 ± 0.05 | 87.80 ± 23.90 |
| 2014 Control | 0.49 ± 0.03 | 0.13 ± 0.03 | 7.91 ± 0.05 | 272.67 ± 67.20 |
| 2014 Spill | 0.75 ± 0.21 | 0.59 ± 0.09 | 7.93 ± 0.14 | 348.67 ± 158.60 |
| Mean No. of OTUs ± SD | Evenness | Observed Species | Chao1 | Shannon–Weaver | |
|---|---|---|---|---|---|
| 1975 control | 7373.3 ± 1090.5 | 0.6 ± 0.03 | 931.7 ± 75.1 | 1285.4 ± 55.7 | 5.8 ± 0.06 |
| 1975 spill | 11,234.7 ± 2585.2 | 0.8 ± 0.01 | 338.7 ± 119.6 | 472.2 ± 171.8 | 3.5 ± 0.31 |
| 2014 control | 8334.0 ± 524.7 | 0.6 ± 0.05 | 806.0 ± 28.3 | 1010.72 ± 59.1 | 5.2 ± 0.13 |
| 2014 spill | 11,608.7 ± 981.6 | 0.8 ± 0.02 | 360.7 ± 57.6 | 477.4 ± 47.9 | 3.6 ± 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martirosyan, V.; Stavi, I.; Doniger, T.; Applebaum, I.; Sherman, C.; Levi, M.; Steinberger, Y. Bacterial Community Dynamics in Oil-Contaminated Soils in the Hyper-Arid Arava Valley. Agronomy 2025, 15, 1198. https://doi.org/10.3390/agronomy15051198
Martirosyan V, Stavi I, Doniger T, Applebaum I, Sherman C, Levi M, Steinberger Y. Bacterial Community Dynamics in Oil-Contaminated Soils in the Hyper-Arid Arava Valley. Agronomy. 2025; 15(5):1198. https://doi.org/10.3390/agronomy15051198
Chicago/Turabian StyleMartirosyan, Varsik, Ilan Stavi, Tirza Doniger, Itaii Applebaum, Chen Sherman, May Levi, and Yosef Steinberger. 2025. "Bacterial Community Dynamics in Oil-Contaminated Soils in the Hyper-Arid Arava Valley" Agronomy 15, no. 5: 1198. https://doi.org/10.3390/agronomy15051198
APA StyleMartirosyan, V., Stavi, I., Doniger, T., Applebaum, I., Sherman, C., Levi, M., & Steinberger, Y. (2025). Bacterial Community Dynamics in Oil-Contaminated Soils in the Hyper-Arid Arava Valley. Agronomy, 15(5), 1198. https://doi.org/10.3390/agronomy15051198