
Removal of Crop Ion Components in Relation to Mollisol Acidification under Long-Term Management

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sites
2.2. Experimental Design and Crop Management
- Continuous Corn (CC): continuous corn with chemical fertilizer.
- Continuous Soybean (CS): continuous soybean with chemical fertilizer.
- CornCF–SoybeanCF (CCFSCF): corn–soybean rotation with chemical fertilizer.
- CornCF–Soybean1/2CF (CCFS1/2CF): corn–soybean rotation with chemical fertilizer for corn, but 1/2 chemical fertilizer for soybean.
- CornCF–SoybeanNOF (CCFSNOF): corn–soybean rotation with chemical fertilizer for corn, but no chemical fertilizer for soybean.
- CornCR–SoybeanNOR (CCRSNOR): corn–soybean rotation with chemical fertilizer plus straw return for corn, no chemical fertilizer but straw return for soybean.
- CornCF–SoybeanDM (CCFSDM): corn–soybean rotation with chemical fertilizer for corn, and dairy manure only for soybean.
2.3. Sampling and Chemical Analysis
2.4. Data Calculation
2.5. Data Analysis
3. Results
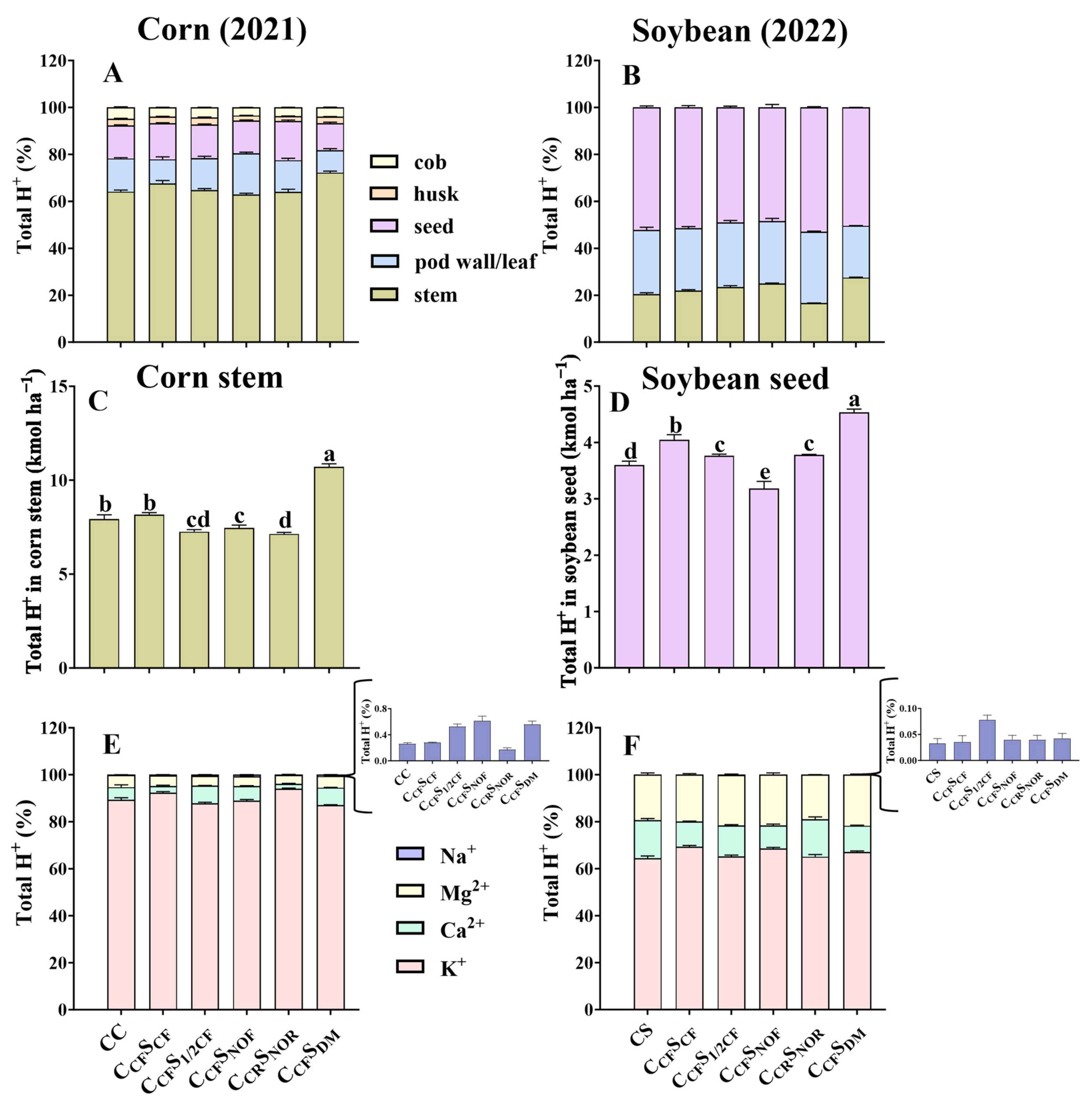
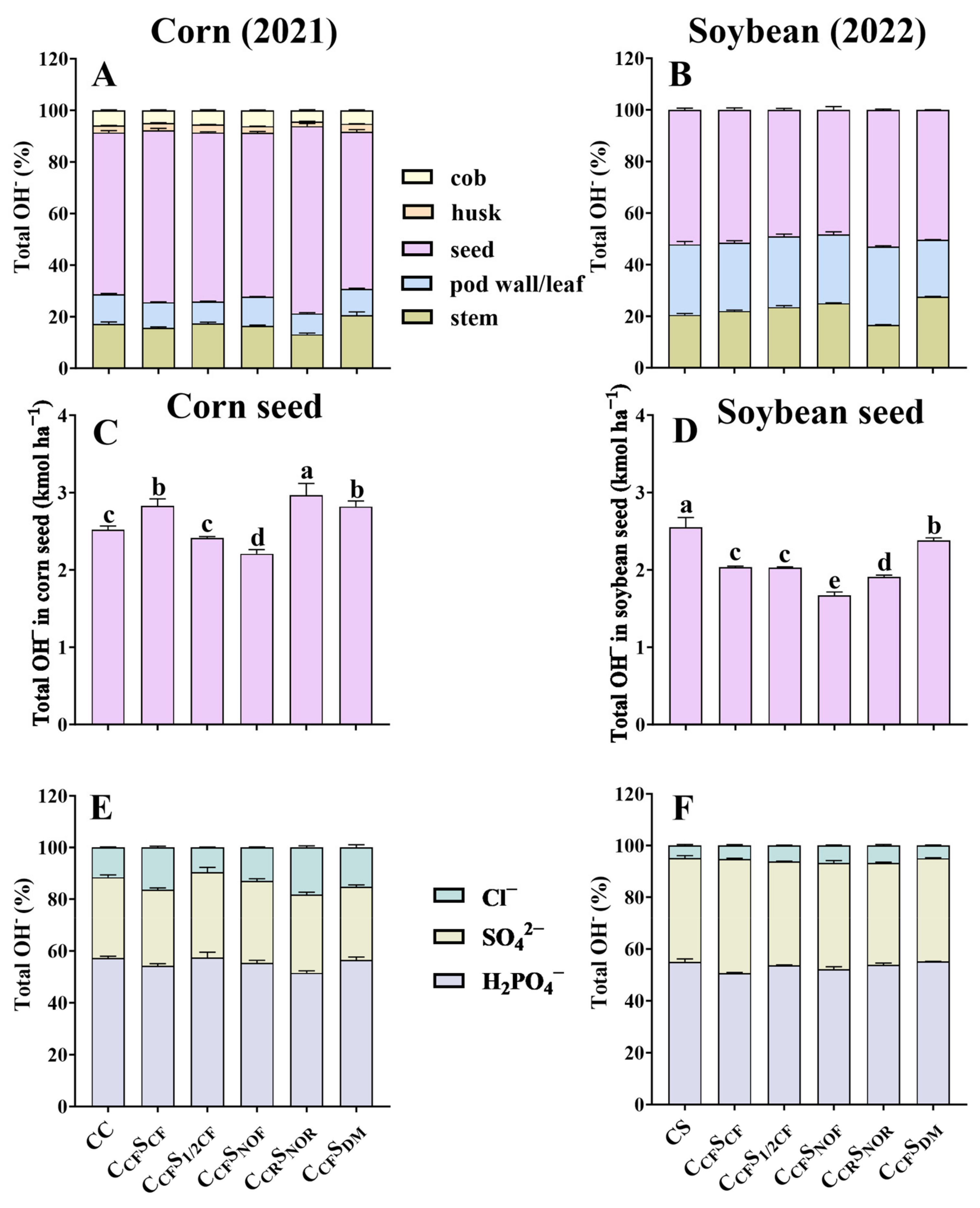
3.1. Crop Biomass, Seed Yield, Total H+, OH−, and Net H+ Production
3.2. Total Amount of Cations and Anions Removed in Crops
3.3. Percentage of Different Tissues for Total H+ and OH− and Contribution Rate of Ion Components
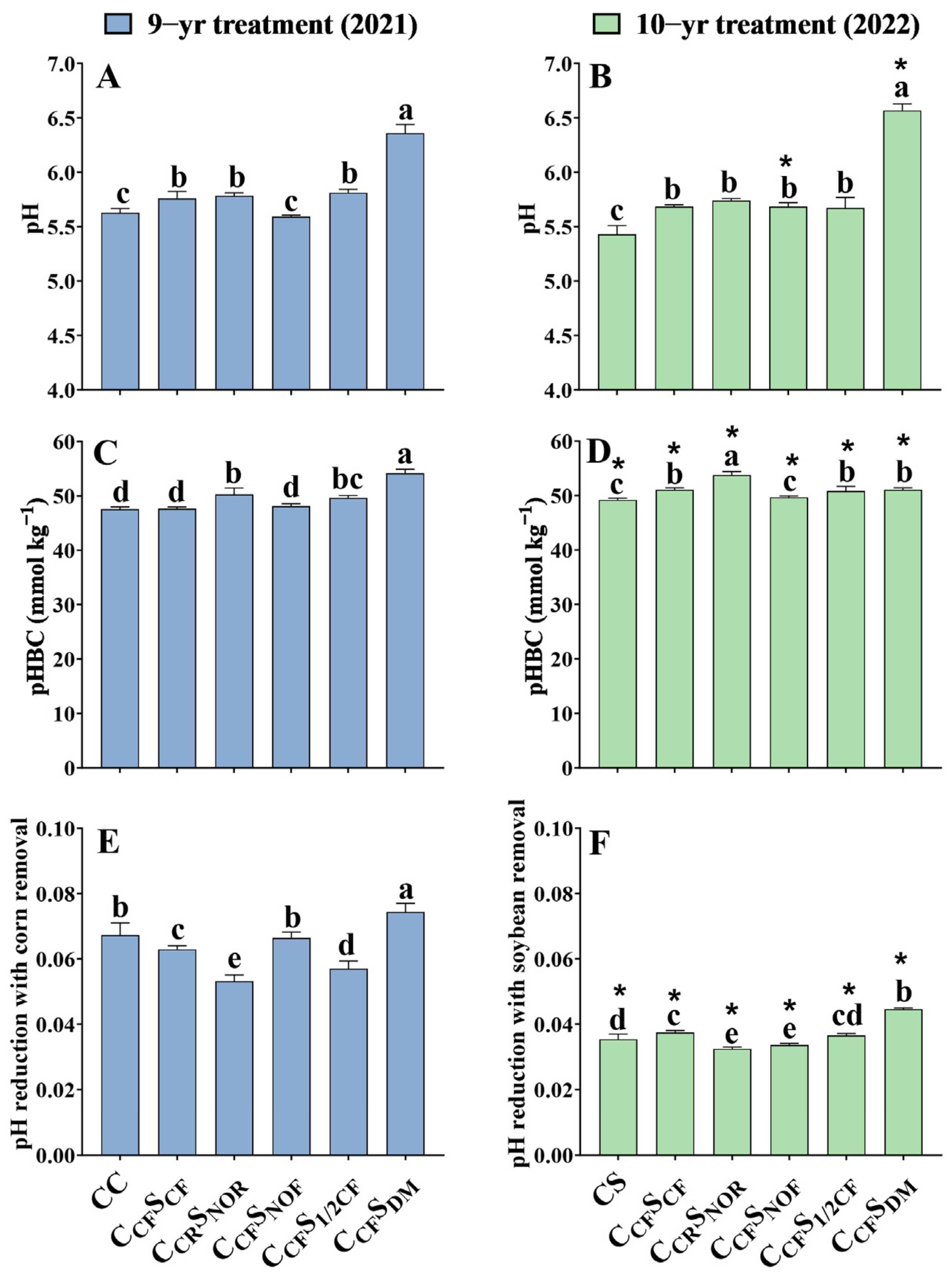
3.4. Soil pH, pHBC, and pH Reduction with Crop Removal
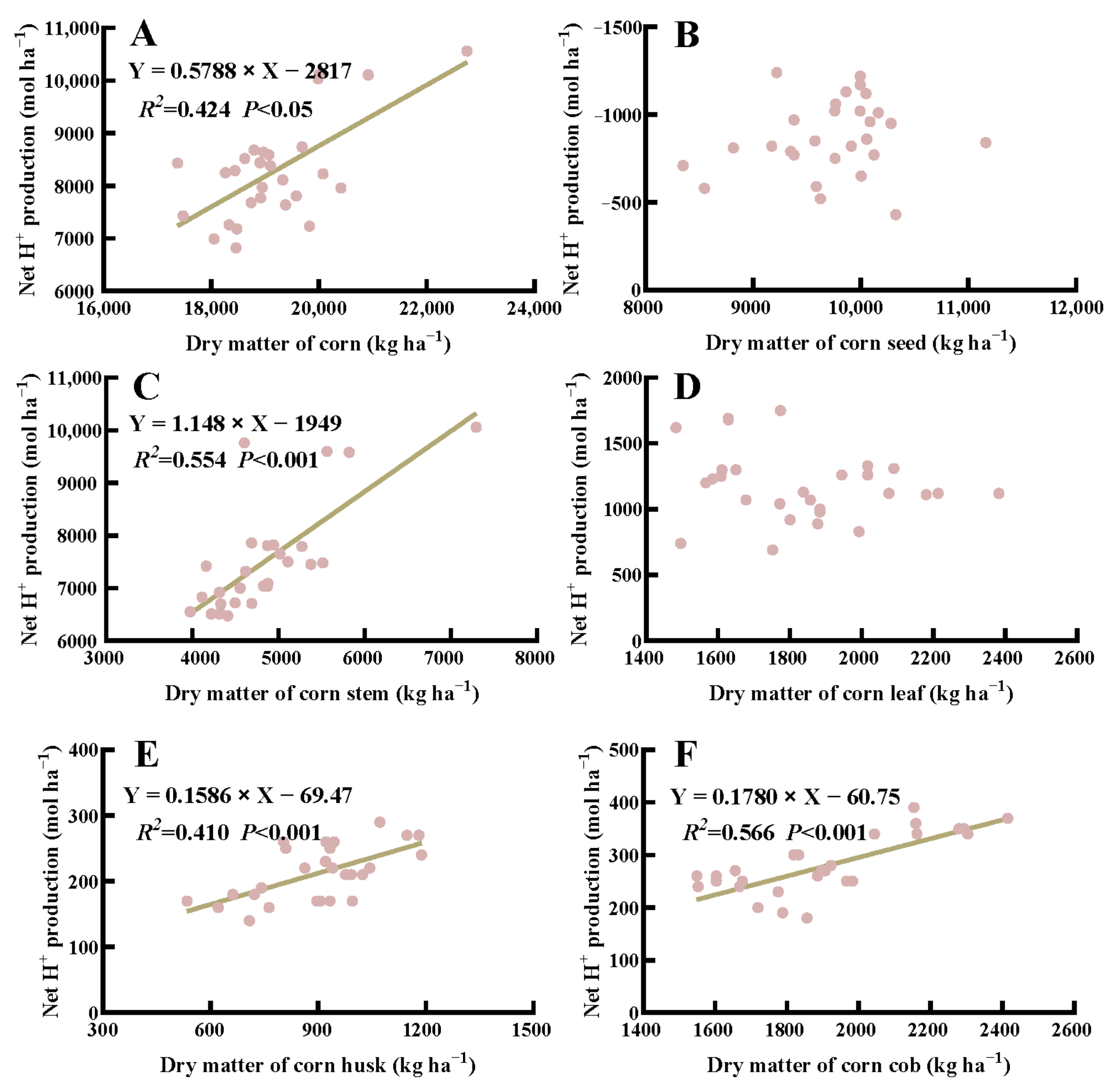
3.5. Correlations between Dry Weight and Net H+ Production in Crops
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Breemen, N.; Driscoll, C.T.; Mulder, J. Acidic deposition and internal proton sources in acidification of soils and waters. Nature 1984, 307, 599–604. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, J.; Zhao, X.; Yang, S.; Zhang, G. Contribution of different proton sources to the acidification of red soil with maize cropping in subtropical China. Geoderma 2021, 392, 114995. [Google Scholar] [CrossRef]
- De Vries, W.; Breeuwsma, A. The relation between soil acidification and element cycling. Water Air Soil Pollut. 1987, 35, 293–310. [Google Scholar] [CrossRef]
- Berner, R.A. Weathering, plants, and the long-term carbon cycle. Geochim. Cosmochim. Acta 1992, 56, 3225–3231. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.; Zhang, Y.; Shen, J.; Han, W.; Zhang, W.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1009. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sun, X.; Sun, Y.; Yan, J.; Zhao, Y.; Chen, J. Soil acidification and factors controlling topsoil pH shift of cropland in central China from 2008 to 2018. Geoderma 2021, 408, 115586. [Google Scholar] [CrossRef]
- Zhu, Q.; Liu, X.; Hao, T.; Zeng, M.; Shen, J.; Zhang, F.; De Vries, W. Modeling soil acidification in typical Chinese cropping systems. Sci. Total Environ. 2018, 613, 1339–1348. [Google Scholar] [CrossRef]
- Zhang, Y.; Ye, C.; Su, Y.; Peng, W.; Lu, R.; Liu, Y.; Huang, H.; He, X.; Yang, M.; Zhu, S. Soil Acidification caused by excessive application of nitrogen fertilizer aggravates soil-borne diseases: Evidence from literature review and field trials. Agric. Ecosyst. Environ. 2022, 340, 108176. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, B.; Xu, M.; Zhang, H.; He, X.; Zhang, L.; Gao, S. Intensified soil acidification from chemical N fertilization and prevention by manure in an 18-year field experiment in the red soil of southern China. J. Soils Sediments 2015, 15, 260–270. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, B.; Xu, M.; Zhang, H.; Zhang, L.; Gao, S. Nitrification and acidification from urea application in red soil (Ferralic Cambisol) after different long-term fertilization treatments. J. Soils Sediments 2014, 14, 1526–1536. [Google Scholar] [CrossRef]
- Schroder, J.L.; Zhang, H.; Girma, K.; Raun, W.R.; Penn, C.J.; Payton, M.E. Soil acidification from long-term use of nitrogen fertilizers on winter wheat. Soil Sci. Soc. Am. J. 2011, 75, 957–964. [Google Scholar] [CrossRef]
- Vieira, F.C.B.; Bayer, C.; Mielniczuk, J.; Zanatta, J.; Bissani, C.A. Long-term acidification of a Brazilian Acrisol as affected by no till cropping systems and nitrogen fertiliser. Soil Res. 2008, 46, 17–26. [Google Scholar] [CrossRef]
- Barak, P.; Jobe, B.O.; Krueger, A.R.; Peterson, L.A.; Laird, D.A. Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Plant Soil 1997, 197, 61–69. [Google Scholar] [CrossRef]
- Tarkalson, D.D.; Payero, J.O.; Hergert, G.W.; Cassman, K.G. Acidification of soil in a dry land winter wheat-sorghum/corn-fallow rotation in the semiarid US Great Plains. Plant Soil 2006, 283, 367–379. [Google Scholar] [CrossRef]
- Zhang, F.; Cui, Z.; Fan, M.; Zhang, W.; Chen, X.; Jiang, R. Integrated soil-crop system management, reducing environmental risk while increasing crop productivity and improving nutrient use efficiency in China. J. Environ. Qual. 2011, 40, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. A plan for efficient use of nitrogen fertilizers. Nature 2017, 543, 322–323. [Google Scholar] [CrossRef]
- Hassan, F.; Ali, E.F.; Mahfouz, S. Comparison between different fertilization sources, irrigation frequency and their combinations on the growth and yield of coriander plant. Aust. J. Basic Appl. Sci. 2012, 6, 600–615. [Google Scholar]
- Xu, R.; Zhao, A.; Yuan, J.; Jiang, J. pH buffering capacity of acid soils from tropical and subtropical regions of China as influenced by incorporation of crop straw biochars. J. Soils Sediments 2012, 12, 494–502. [Google Scholar] [CrossRef]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints, A review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, J.; Zhao, X.; Yang, S.; Mulder, J.; Dorsch, P.; Peng, X.; Zhang, G. Soil acidification and loss of base cations in a subtropical agricultural watershed. Sci. Total Environ. 2022, 827, 154338. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.X.; Zhu, Q.; Zeng, M.; Shen, J.; Shi, X.; Liu, X.; Zhang, F.; de Vries, W. Quantification of the contribution of nitrogen fertilization and crop harvesting to soil acidification in a wheat-maize double cropping system. Plant Soil 2018, 434, 167–184. [Google Scholar] [CrossRef]
- Hao, T.; Liu, X.; Zhu, Q.; Zeng, M.; Chen, X.; Yang, L.; Shen, J.; Shi, X.; Zhang, F.; de Vries, W. Quantifying drivers of soil acidification in three Chinese cropping systems. Soil Tillage Res. 2022, 215, 105230. [Google Scholar] [CrossRef]
- Liu, X.; Burras, C.L.; Kravchenko, Y.S.; Duran, A.; Huffman, T.; Morras, H.; Studdert, G.; Zhang, X.; Cruse, R.; Yuan, X. Overview of Mollisols in the world: Distribution, land use and management. Can. J. Soil Sci. 2012, 92, 383–402. [Google Scholar] [CrossRef]
- Ding, J.; Ma, M.; Jiang, X.; Liu, Y.; Zhang, J.; Suo, L.; Wang, L.; Wei, D.; Li, J. Effects of applying inorganic fertilizer and organic manure for 35 years on the structure and diversity of ammonia-oxidizing archaea communities in a Chinese Mollisols field. Microbiol. Open 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Ma, M.; Jiang, X.; Wang, Q.; Ongena, M.; Wei, D.; Ding, J.; Guan, D.; Cao, F.; Zhao, B.; Li, J. Responses of fungal community composition to long-term chemical and organic fertilization strategies in Chinese Mollisols. Microbiol. Open 2018, 7, e597. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, X.; Huang, B.; Zheng, H.; Deng, W. Uncertainty assessment for mapping changes in soil organic matter using sparse legacy soil data and dense new-measured data in a typical black soil region of China. Environ. Earth Sci. 2015, 73, 197–207. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.; Zhou, B.; Zhao, B.; Ma, M.; Qin, J.; Jiang, X.; Chen, S.; Cao, F.; Shen, D.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.; Cao, F.; Shen, D.; et al. Thirty-four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in northeast China. Soil Biol. Biochem. 2016, 95, 135–143. [Google Scholar] [CrossRef]
- Dabessa, A.; Debala, C. Long-term soybean-maize rotation experiments in cereal-based farming systems at Bako, Western Ethiopia. Food Energy Secur. 2023, 12, 1–13. [Google Scholar] [CrossRef]
- Posch, M.; Reinds, G.J. A very simple dynamic soil acidification model for scenario analyses and target load calculations. Environ. Model. Softw. 2009, 24, 329–340. [Google Scholar] [CrossRef]
- Verstraten, J.M.; Dopheide, J.C.R.; Duysings, J.J.H.M.; Tietema, A.; Bouten, W. The proton cycle of a deciduous forest ecosystem in the Netherlands and its implications for soil acidification. Plant Soil 1990, 127, 61–69. [Google Scholar] [CrossRef]
- Lesturgez, G.; Poss, R.; Noble, A.; Grünberger, O.; Chintachao, W.; Tessier, D. Soil acidification without pH drop under intensive cropping systems in Northeast Thailand. Agric. Ecosyst. Environ. 2006, 114, 239–248. [Google Scholar] [CrossRef]
- Houx, J.H.; Wiebold, W.J.; Fritschi, F.B. Rotation and tillage affect soybean grain composition, yield, and nutrient removal. Field Crop. Res. 2014, 164, 12–21. [Google Scholar] [CrossRef]
- Osaki, M.; Morikawa, K.; Shinano, T.; Urayama, M.; Tadano, T. Productivity of high-yielding crops. II. Comparison of N, P, K, Ca, and Mg accumulation and distribution among high-yielding crops. Soil Sci. Plant Nutr. 1991, 37, 445–454. [Google Scholar] [CrossRef]
- Xu, Z.; Lai, T.; Li, S.; Si, D.; Zhang, C.; Cui, Z.; Chen, X. Promoting potassium allocation to stalk enhances stalk bending resistance of maize (Zea mays L.). Field Crop. Res. 2018, 215, 200–206. [Google Scholar] [CrossRef]
- Noble, A.D.; Suzuki, S.; Soda, W.; Ruaysoongnern, S.; Berthelsen, S. Soil acidification and carbon storage in fertilized pastures of Northeast Thailand. Geoderma 2008, 144, 248–255. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, J.; Wang, Q.; Mulder, J.; Liu, C.; He, N. Transformation from NHx to NOy deposition aggravated China’s forest soil acidification. Glob. Biogeochem. Cycle 2023, 37, 1–10. [Google Scholar] [CrossRef]
- Duan, L.; Chen, X.; Ma, X.; Zhao, B.; Larssen, T.; Wang, S.; Ye, Z. Atmospheric S and N deposition relates to increasing riverine transport of S and N in southwest China, Implications for soil acidification. Environ. Pollut. 2016, 218, 1191–1199. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, C.; Ma, W.; Wang, S.; Wang, S.; Han, W.; Mohammat, A.; Robinson, D.; Smith, P. Significant soil acidification across northern China’s grasslands during 1980s–2000s. Glob. Chang. Biol. 2012, 18, 2292–2300. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Q.; Zhu, J.; Xu, L.; Li, M.; Rengel, Z.; Xiao, J.; Hobbie, E.A.; Piao, S.; Luo, W.; et al. Higher soil acidification risk in southeastern Tibetan Plateau. Sci. Total Environ. 2021, 755, 143372. [Google Scholar] [CrossRef]
- Randall, P.J.; Abaidoo, R.C.; Hocking, P.J.; Sanginga, N. Mineral nutrient uptake and removal by cowpea, soybean and maize cultivars in West Africa, and implications for carbon cycle effects on soil acidification. Exp. Agric. 2006, 42, 475–494. [Google Scholar] [CrossRef]
- Slattery, W.J.; Ridley, A.M.; Windsor, S.M. Ash alkalinity of animal and plant products. Aust. J. Exp. Agric. 1991, 31, 321–324. [Google Scholar] [CrossRef]
- Hickman, M.V. Long-term tillage and crop rotation effects on soil chemical and mineral properties. J. Plant Nutr. 2002, 25, 1457–1470. [Google Scholar] [CrossRef]
- White, A.; Darby, H.; Ruhl, L.; Sands, B. Long-term influence of alternative corn cropping practices and corn-hay rotations on soil health, yields and forage quality. Front. Environ. Sci. 2023, 11, 1–19. [Google Scholar] [CrossRef]
- Behnke, G.D.; Zuber, S.M.; Pittelkow, C.M.; Nafziger, E.D.; Villamil, M.B. Long-term crop rotation and tillage effects on soil greenhouse gas emissions and crop production in Illinois, USA. Agric. Ecosyst. Environ. 2018, 261, 62–70. [Google Scholar] [CrossRef]
- Kazula, M.J.; Lauer, J.G. The influence of crop rotation on corn total biomass production. J. Soil Water Conserv. 2018, 73, 541–548. [Google Scholar] [CrossRef]
- Sainju, U.M.; Lenssen, A.W.; Allen, B.L.; Jabro, J.D.; Stevens, W.B. Crop water and nitrogen productivity in response to long-term diversified crop rotations and management systems. Agric. Water Manag. 2021, 257, 107149. [Google Scholar] [CrossRef]
- Dal Molin, S.J.; Ernani, P.R.; Gerber, J.M. Soil acidification and nitrogen release following application of nitrogen fertilizers. Commun. Soil Sci. Plant Anal. 2020, 51, 2551–2558. [Google Scholar] [CrossRef]
- Fujii, K.; Toma, T.; Sukartiningsih. Comparison of soil acidification rates under different land uses in Indonesia. Plant Soil 2021, 465, 1–17. [Google Scholar] [CrossRef]
- Wang, Z.; Tao, T.; Wang, H.; Chen, J.; Small, G.E.; Johnson, D.; Chen, J.; Zhang, Y.; Zhu, Q.; Zhang, S.; et al. Forms of nitrogen inputs regulate the intensity of soil acidification. Glob. Chang. Biol. 2023, 29, 4044–4055. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Zhao, Y.; Xu, R. Soil acidification of alfisols influenced by nitrate and ammonium nitrogen level in tea plantation. Int. J. Agric. Biol. Eng. 2018, 11, 168–172. [Google Scholar] [CrossRef]
- Bolan, N.S.; Hedley, M.J.; White, R.E. Processes of soil acidification during nitrogen cycling with emphasis on legume-based pastures. Plant Soil 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Vieira, F.C.B.; Bayer, C.; Zanatta, J.; Ernani, P.R. Organic matter kept Al toxicity low in a subtropical no-tillage soil under long-term (21-year) legume-based crop systems and N fertilisation. Soil Res. 2009, 47, 707–714. [Google Scholar] [CrossRef]
- Liang, F.; Li, B.; Vogt, R.D.; Mulder, J.; Song, H.; Chen, J.; Guo, J. Straw return exacerbates soil acidification in major Chinese croplands. Resour. Conserv. Recycl. 2023, 198, 107176. [Google Scholar] [CrossRef]
- Wang, N.; Xu, R.; Li, J. Amelioration of an acid Ultisol by agricultural by-products. Land Degrad. Dev. 2011, 22, 513–518. [Google Scholar] [CrossRef]
- Noble, A.D.; Randall, P.J. Alkalinity effects of different tree litters incubated in an acid soil of NSW, Australia. Agrofor. Syst. 1999, 46, 147–160. [Google Scholar] [CrossRef]
- Tang, C.; Sparling, G.P.; McLay, C.D.A.; Raphael, C. Effect of short-term legume residue decomposition on soil acidity. Aust. J. Soil Res. 1999, 37, 561–573. [Google Scholar]
- Naramabuye, F.X.; Haynes, R.J. Effect of organic amendments on soil pH and Al solubility and use of laboratory indices to predict their liming effect. Soil Sci. 2006, 171, 754–763. [Google Scholar] [CrossRef]
- Hue, N.V. Alleviating soil acidity with crop residues. Soil Sci. 2011, 176, 543–549. [Google Scholar] [CrossRef]
- Ano, A.O.; Ubochi, C.I. Neutralization of soil acidity by animal manures, mechanism of reaction. Afr. J. Biotechnol. 2007, 6, 364–368. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | K+ | Na+ | Ca2+ | Mg2+ | SO42− | Cl− | H2PO4− | |
|---|---|---|---|---|---|---|---|---|
| kg ha−1 | ||||||||
| Corn (2021) | CC | 368 c | 1.59 c | 16.1 b | 24.4 b | 20.5 b | 16.4 c | 70.2 b |
| CCFSCF | 381 b | 2.16 a | 7.69 d | 21.9 c | 20.4 b | 22.3 b | 72.3 b | |
| CCFS1/2CF | 329 e | 1.34 d | 18.9 a | 21.2 c | 19.2 bc | 13.6 d | 65.2 c | |
| CCFSNOF | 337 d | 1.65 c | 18.7 a | 26.5 a | 18.2 c | 15.7 c | 58.5 d | |
| CCRSNOR | 342 d | 1.83 b | 9.70 c | 21.6 c | 20.7 b | 23.6 a | 66.0 c | |
| CCFSDM | 463 a | 2.26 a | 19.8 a | 22.2 c | 22.4 a | 22.4 b | 80.2 a | |
| Soybean (2022) | CS | 128 d | 0.22 b | 30.4 e | 25.2 d | 16.3 c | 6.87 e | 38.9 e |
| CCFSCF | 149 b | 0.30 a | 31.8 d | 29.4 b | 20.4 a | 7.42 cd | 42.4 c | |
| CCFS1/2CF | 133 c | 0.24 b | 34.8 c | 30.1 a | 18.2 b | 7.79 bc | 43.3 b | |
| CCFSNOF | 109 e | 0.17 c | 28.9 f | 28.1 c | 14.0 d | 7.26 de | 34.3 f | |
| CCRSNOR | 134 c | 0.22 b | 35.8 b | 22.9 e | 15.9 c | 7.97 b | 40.4 d | |
| CCFSDM | 183 a | 0.17 c | 38.8 a | 28.1 c | 20.4 a | 9.70 a | 51.8 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Yu, Z.; Li, Y.; Jin, J.; Zhang, X.; Wang, G.; Liu, X. Removal of Crop Ion Components in Relation to Mollisol Acidification under Long-Term Management. Agronomy 2024, 14, 191. https://doi.org/10.3390/agronomy14010191
Xu Y, Yu Z, Li Y, Jin J, Zhang X, Wang G, Liu X. Removal of Crop Ion Components in Relation to Mollisol Acidification under Long-Term Management. Agronomy. 2024; 14(1):191. https://doi.org/10.3390/agronomy14010191
Chicago/Turabian StyleXu, Ying, Zhenhua Yu, Yansheng Li, Jian Jin, Xingyi Zhang, Guanghua Wang, and Xiaobing Liu. 2024. "Removal of Crop Ion Components in Relation to Mollisol Acidification under Long-Term Management" Agronomy 14, no. 1: 191. https://doi.org/10.3390/agronomy14010191
APA StyleXu, Y., Yu, Z., Li, Y., Jin, J., Zhang, X., Wang, G., & Liu, X. (2024). Removal of Crop Ion Components in Relation to Mollisol Acidification under Long-Term Management. Agronomy, 14(1), 191. https://doi.org/10.3390/agronomy14010191