


Characterization of Local Mediterranean Grapevine Varieties for Their Resilience to Semi-Arid Conditions under a Rain-Fed Regime
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Varietal Identification, and Growing Conditions
2.2. Climatic Conditions
2.3. Ampelographic Characterization of Varieties
2.4. Vigour and Other Related Agronomical and Quality Traits
2.5. Stomatal Conductance and Other Gas Exchange Measurements
2.6. Stomatal Characterization
2.7. Statistical Analysis
3. Results
3.1. Ampelographic and Agronomical Related Traits
3.2. Evaluation of Stomatal Conductance (GS) and Related Parameters
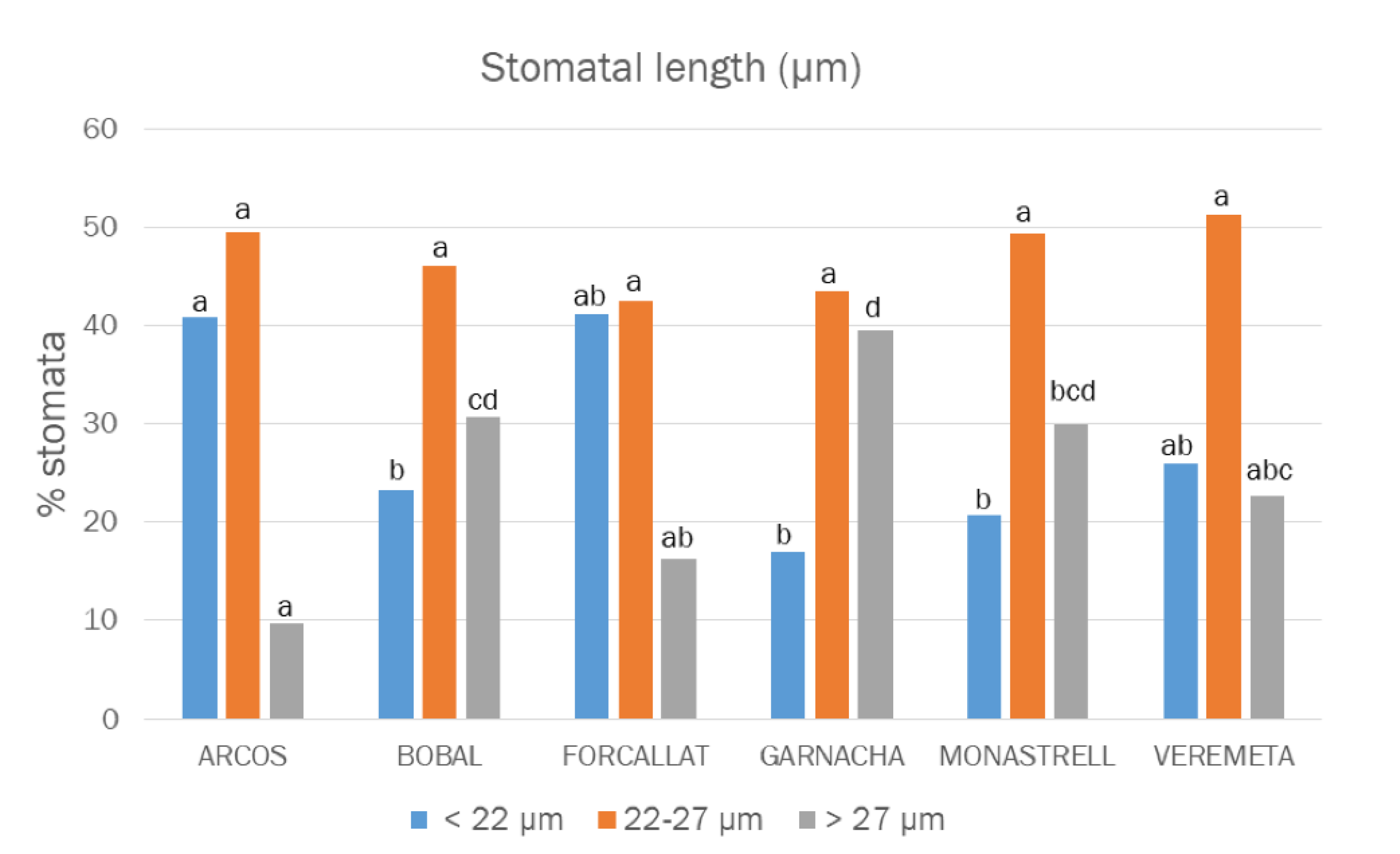
3.3. Stomata Densities and Size
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Leeuwen, C.; Destrac-Irvine, A. Modified Grape Composition under Climate Change Conditions Requires Adaptations in the Vineyard. OENO One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of Drought on Photosynthesis in Grapevines under Field Conditions: An Evaluation of Stomatal and Mesophyll Limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S. The Physiology of Drought Stress in Grapevine: Towards an Integrative Definition of Drought Tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate Change and Interconnected Risks to Sustainable Development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulias, J.; Pou, A.; Ribas-Carbo, M.; Tomàs, M.; Medrano, H. Improving Water Use Efficiency in Grapevines: Potential Physiological Targets for Biotechnological Improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving Water Use Efficiency of Vineyards in Semi-Arid Regions. A review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef]
- Medrano, H.; Tortosa, I.; Montes, E.; Pou, A.; Balda, P.; Bota, J.; Escalona, J.M. Genetic improvement of grapevine (Vitis vinifera L.) water use efficiency. In Water Scarcity and Sustainable Agriculture in Semiarid Environment: Tools, Strategies, and Challenges for Woody Crops, 1st ed.; García, I.F., Durán, V.H., Eds.; Academic Press: London, UK, 2018; Volume 1, pp. 377–401. [Google Scholar]
- Antolín, M.C.; Izurdiaga, D.; Urmeneta, L.; Pascual, I.; Irigoyen, J.J.; Goicoechea, N. Dissimilar Responses of Ancient Grapevines Recovered in Navarra (Spain) to Arbuscular Mycorrhizal Symbiosis in Terms of Berry Quality. Agronomy 2020, 10, 473. [Google Scholar] [CrossRef]
- Peiró, R.; Jiménez, C.; Perpiñà, G.; Soler, J.X.; Gisbert, C. Evaluation of the Genetic Diversity and Root Architecture under Osmotic Stress of Common Grapevine Rootstocks and Clones. Sci. Hortic. 2020, 266, 109283. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of Photosynthesis of C3 Plants in Response to Progressive Drought: Stomatal Conductance as a Reference Parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Boso, S.; Gago, P.; Alonso-Villaverde, V.; Santiago, J.L.; Martínez, M.C. Density and Size of Stomata in the Leaves of Different Hybrids (Vitis sp.) and Vitis vinifera Varieties. Vitis 2016, 55, 17–22. [Google Scholar]
- Serra, I.; Strever, A.; Myburgh, P.; Schmeisser, M.; Deloire, P.A. Grapevine (Vitis vinifera L. ‘Pinotage’) Leaf Stomatal Size and Density as Modulated by Different Rootstocks and Scion Water Status. Acta Hortic. 2017, 1157, 177–181. [Google Scholar] [CrossRef]
- MacMillan, P.; Teixeira, G.; Lopes, C.M.; Monteiro, A. The role of grapevine leaf morphoanatomical traits in determining capacity for coping with abiotic stresses: A review. Cienc Tec Vitivinic 2021, 36, 75–88. [Google Scholar] [CrossRef]
- Peiró, R.; Soler, J.; Crespo, A.; Jiménez, C.; Cabello, F.; Gisbert, C. Genetic Variability Assessment in ‘Muscat’ Germplasm Including ‘Muscat of Alexandria’ Clones from Selection Programs. Span. J. Agric. Res. 2018, 16, e0702. [Google Scholar] [CrossRef]
- OIV Protocol for Identification of Varieties. Available online: https://www.oiv.int/public/medias/6886/oiv-viti-609-2019-en.pdf (accessed on 5 July 2022).
- Iland, P.; Bruer, N.; Edwards, G.; Weeks, S.; Wilkes, E. Chemical Analysis of Grapes and Wine Techniques and Concepts, 2nd ed.; Wine Promotions Pty Ltd.: Athelstone, SA, Australia, 2004; p. 120. [Google Scholar]
- Fischer, R.A.; Turner, N.C. Plant productivity in the arid and semiarid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Miller, N.A.; Ashby, W.C. Studying Stomates with Polish. Turtox News 1968, 46, 322–324. [Google Scholar]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- OIV Focus 2017-Vine Varieties in the World. Available online: https://www.oiv.int/public/medias/5336/infographie-focus-oiv-2017-new.pdf (accessed on 5 July 2022).
- Resco, P.; Iglesias, A.; Bardají, I.; Sotés, V. Exploring Adaptation Choices for Grapevine Regions in Spain. Reg. Environ. Chang. 2016, 16, 979–993. [Google Scholar] [CrossRef]
- Audrey, N.; Christian, G.; Laurent, P.; Laure, H. Evaluating Strategies for Adaptation to Climate Change in Grapevine Production–A Systematic Review. Front. Plant Sci. 2021, 11, 607859. [Google Scholar]
- Dirección General de Agricultura, Industria y Comercio (DGAIC). Avance Estadístico sobre Cultivo y Producción de la Vid en España Formado por la Junta Consultiva Agronómica, 1st ed.; Péant e Hijos: Madrid, Spain, 1891; pp. 70–73. [Google Scholar]
- State of the World Vitivinicultural Sector in 2020. Available online: https://www.oiv.int/public/medias/7909/oiv-state-of-the-world-vitivinicultural-sector-in-2020.pdf (accessed on 5 July 2022).
- Valcárcel, J.A. Agricultura General, 1st ed.; Ediciones Josep Esteban y Cervera: Spain, Valencia, 1791; pp. 290–295. [Google Scholar]
- Evaluation Support Study on Geographical Indications and Traditional Specialities Guaranteed Protected in the EU. Available online: https://www.mapa.gob.es/es/alimentacion/temas/calidad-diferenciada/estudioapoyoevaluacioniiggyetgenlaue_tcm30-560200.pdf (accessed on 5 July 2022).
- Jiménez, C.; Peiró, R.; Yuste, A.; García, J.; Martínez-Gil, F.; Gisbert, C. Looking for Old Grapevine Varieties. Vitis 2019, 58, 59–60. [Google Scholar]
- Janini, R. Resumen de Trabajos Realizados desde el 1 de Abril de 1921 al 31 de Mayo de 1922 en la Estación Enológica de Requena, 1st ed.; de Imprenta Hijo, F., Ed.; Vives Mora: Spain, Valencia, 1922. [Google Scholar]
- García de los Salmones, N. Dos Conferencias sobre Vinos y Uvas de Mesa de España: Ciclo de Conferencias Desarrollado en la Escuela Especial de Ingenieros Agrónomos, 1st ed.; Instituto Nacional Agronómico: Spain, Madrid, 1935; p. 13. [Google Scholar]
- Tello, J.; Ibáñez, J. What do we know about grapevine bunch compactness? A state-of-the-art review. Aust. J. Grape Wine Res. 2017, 24, 6–23. [Google Scholar] [CrossRef]
- Cortiñas, J.A.; González-Fernández, E.; Fernández–González, M.; Vázquez–Ruiz, R.A.; Aira, M.J. Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model. Agronomy 2020, 10, 219. [Google Scholar] [CrossRef]
- Gavara, A.; Navarro-Llopis, V.; Primo, J.; Vacas, S. Influence of weather conditions on Lobesia botrana (Lepidoptera: Tortricidae) mating disruption dispensers’ emission rates and efficacy. Crop. Prot. 2022, 155, 105926. [Google Scholar] [CrossRef]
- Lavoie-Lamoureux, A.; Sacco, D.; Risse, P.A.; Lovisolo, C. Factors influencing stomatal conductance in response to water availability in grapevine: A meta-analysis. Physiol. Plant 2017, 159, 468–482. [Google Scholar] [CrossRef]
- Toro, G.; Flexas, J.; Escalona, J.M. Contrasting leaf porometer and infra-red gas analyser methodologies: An old paradigm about the stomatal conductance measurement. Theor. Exp. Plant Physiol. 2019, 31, 483–492. [Google Scholar] [CrossRef]
- Jones, H.G. Interaction and integration of adaptive responses to water stress: The implications of an unpredictable environment. In Adaptation of Plants to Water and High Temperature Stress, 1st ed.; Turner, N.C., Kramer, P.J., Eds.; Wiley: Hoboken, NJ, USA, 1980; Volume 1, pp. 353–365. [Google Scholar]
- Tardieu, F.; Simonneau, T. Variability among Species of Stomatal Control Under Fluctuating Soil Water Status and Evaporative Demand: Modelling Isohydric and Anysohydric Behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef]
- Williams, L.E.; Araujo, F.J. Correlations among Predawn Leaf, Midday Leaf, and Midday Stem Water Potential and their Correlations with other Measures of Soil and Plant Water Status in Vitis vinifera. J. Am. Soc. Hortic. Sci. 2002, 127, 448–454. [Google Scholar] [CrossRef]
- Bota, J.; Tomás, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among Grapevine Cultivars in their Stomatal Behaviour and Water Use Efficiency under Progressive Water Stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Toro, G.; Douthe, C.; Medrano, H. Clonal Behavior in Response to Soil Water Availability in Tempranillo Grapevine cv: From Plant Growth to Water Use Efficiency. Agronomy 2020, 10, 862. [Google Scholar] [CrossRef]
- Romero, P.; Fernández-Fernández, J.I.; Martínez-Cutillas, A. Physiological Thresholds for Efficient Regulated Deficit-Irrigation Management in Winegrapes Grown under Semiarid Conditions. Am. J. Enol. Vitic 2010, 61, 300–312. [Google Scholar]
- Galdon-Armero, J.; Fullana-Pericas, M.; Mulet, P.A.; Conesa, M.A.; Martin, C.; Galmes, J. The Ratio of Trichomes to Stomata is Associated with Water Use Efficiency in Solanum lycopersicum (Tomato). Plant J. 2018, 96, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-González, L.; Muñoz-Araya, M.; Franck, N.; Pastenes, C. Controversies in Midday Water Potential Regulation and Stomatal Behaviour Might Result from the Environment, Genotype, and/or Rootstock: Evidence from Carménère and Syrah Grapevine Varieties. Front. Plant Sci. 2019, 10, 1522. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.; Sutherland, R. Stomatal Control of Xylem Embolism. Plant Cell Environ. 1991, 14, 607–612. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-Induced Changes in Development and Function of Grapevine (Vitis spp.) Organs and in their Hydraulic and Non-Hydraulic Interactions at the Whole-Plant Level: A Physiological and Molecular Update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Keller, M. Managing Grapevines to Optimise Fruit Development in a Challenging Environment: A Climate Change Primer for Viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–60. [Google Scholar] [CrossRef]
- Teixeira, G.; Monteiro, A.; Santos, C.; Lopes, C.M. Leaf Morphoanatomy Traits in White Grapevine Cultivars with Distinct Geographical Origin. Ciência Técnica Vitivinícola 2018, 33, 90–101. [Google Scholar] [CrossRef]
- Nassuth, A.; Rahman, M.A.; Nguyen, T.; Ebadi, A.; Lee, C. Leaves of more Cold Hardy Grapes Have a Higher Density of Small, Sunken Stomata. Vitis 2021, 60, 63–67. [Google Scholar]
- Costa, J.M.; Ortuño, M.F.; Lopes, C.M.; Chavesria, M.M. Grapevine Varieties Exhibiting Differences in Stomatal Response to Water Deficit. Funct. Plant Biol. 2012, 39, 179–189. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Greer, D.H.; Hutton, R.J.; Landsberg, J.J. Does Night Time Transpiration Contribute to Anisohydric Behaviour in a Vitis vinifera Cultivar? J. Exp. Bot. 2009, 60, 3751–3763. [Google Scholar] [CrossRef]
- Hetherington, A.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–907. [Google Scholar] [CrossRef]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, Faster Stomata: Scaling of Stomatal Size, Rate of Response, and Stomatal Conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Arcos | Bobal | Forcallat | Garnacha | Monastrell | Veremeta Clara | |
|---|---|---|---|---|---|---|
| Young shoot | ||||||
| OIV 1 003 | 3 | 1 | 1 | 5 | 3 | 3 |
| OIV 004 | 7 | 7 | 7 | 1 | 9 | 9 |
| Young leaf | ||||||
| OIV 051 | 3 | 2 | 2 | 3 | 3 | 3 |
| OIV 053 | 5 | 7 | 7 | 1 | 7 | 7 |
| Mature leaf | ||||||
| OIV 067 | 3 | 4 | 4 | 3 | 3 | 3 |
| OIV 068 | 3 | 3 | 4 | 3 | 3 | 3 |
| OIV 076 | 5 | 3 | 2 | 2 | 2 | 2 |
| OIV 079 | 5 | 5 | 3 | 3 | 5 | 3 |
| OIV 080 | 3 | 3 | 1 | 1 | 5 | 1 |
| OIV 081-2 | 1 | 1 | 1 | 1 | 1 | 1 |
| OIV 084 | 5 | 7 | 7 | 1 | 7 | 7 |
| OIV 087 | 1 | 3 | 1 | 1 | 1 | 1 |
| OIV 094 | 5 | 5 | 7 | 3 | 1 | 3 |
| Bunch | ||||||
| OIV 202 | 7 [234] (210–260) | 5 [182] (170–190) | 7 [226] (220–230) | 3 [130] 120–150 | 5 [166] (150–180) | 7 [196] (180–230) |
| OIV 203 | 5 [128] (110–140) | 5 [134] (130–140) | 5 [144] (130–160) | 3 [96] (80–110) | 5 [128] (120–140) | 5 [120] (110–140) |
| OIV 204 | 5 | 7 | 5 | 7 | 7 | 5 |
| OIV 206 | 3 [40] (40–10) | 1 [16] (10–20) | 3 [36] (30–40) | 1 [18] (10–20) | 1 [18] (10–30) | 1 [24] (20–30) |
| OIV 208 | 2 | 2 | 2 | 3 | 2 | 2 |
| OIV 502 | 5 [390] (300–450) | 5 [417] (355–483) | 5 [394] (320–450) | 3 [177] (133–266) | 3 [258] (230–287) | 3 [218] (180–288) |
| Berry | ||||||
| OIV 220 | 3 [13.4] (13.0–14.0) | 3 [16.6] (16.0–17.0) | 3 [16.3] (16.0–17.0) | 3 [14.7] (14.0–15.0) | 3 [14.4] (13.0–15.0) | 3 [11.8] (11.0–13.0) |
| OIV 221 | 3 [13.2] (13.0–14.0) | 3 [15.8] (15.0–16.0) | 3 [13.2] (13.0–14.0) | 3 [13.3] (13.0–14.0) | 3 [12.6] (12.0–13.0) | 3 [12.6] (12.0–13.0) |
| OIV 223 | 2 | 2 | 3 | 2 | 2 | 2 |
| OIV 225 | 6 | 6 | 6 | 5 | 6 | 6 |
| OIV 236 | 4 | 4 | 4 | 1 | 1 | 4 |
| OIV 242 | 5 [5.3] (5.0–5.5) | 7 [7.1] (7.0–7.5) | 5 [5.3] (5.0–5.5) | 5 [5.1] (5.0–5.5) | 5 [5.2] (5.0–5.5) | 5 [5.8] (5.5–6.0) |
| OIV 503 | 1 [1.89] (1.80–1.90) | 3 [3.07] (3.00–3.20) | 1 [1.89] (1.80–1.90) | 1 [1.73] (1.70–1.80) | 1 [1.97] (1.90–2.00) | 1 [1.65] (1.60–1.70) |
| Variety | VI | VH | VW | WB | TSS | pH | TA | TPI | A | A/TSS |
|---|---|---|---|---|---|---|---|---|---|---|
| Arcos | 2.4 bc | 71 a | 110 bc | 144 a | 16.4 a | 3.5 | 4.74 b | 3.3 b | 1.09 b | 0.066 |
| Bobal | 3.6 de | 85 b | 199 d | 264 b | 18.5 b | 4.2 d | 3.14 a | 3.0 b | 1.16 c | 0.063 |
| Forcallat | 2.0 b | 68 a | 76 a | 142 a | 15.4 a | 3.9 bc | 3.48 a | 2.5 a | 0.70 a | 0.046 |
| Garnacha | 4.0 e | 153 d | 207 d | 153 a | 21.3 c | 4.2 d | 3.56 a | 2.2 a | 0.69 a | 0.032 |
| Monastrell | 1.2 a | 63 a | 124 c | 147 a | 19.2 b | 3.8 b | 3.23 a | 3.1 b | 1.20 d | 0.063 |
| Veremeta | 3 cd | 133 c | 98 b | 158 a | 23.1 c | 3.9 c | 4.88 b | 3.2 b | 1.59 e | 0.069 |
| 2020 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variety | 16 July (am) | 27 July (am) | 10 August (am) | 10 August (pm) | % R | |||||
| Arcos | 277 a 1 | 279 a 1 | 282 ab 1 A | 172 a A | 38.5 | |||||
| Bobal | 377 ab 1 | 430 bc 1 | 438 bc 1 A | 424 b A | 0.0 | |||||
| Forcallat | 369 ab 2 | 366 ab 2 | 247 a 1 A | 146 a A | 55.5 | |||||
| Garnacha | 405 ab 1 | 413 bc 1 | 388 abc 1 A | 380 ab A | 5.5 | |||||
| Monastrell | 408 b 1 | 391 b 1 | 348 abc 1 A | 282 ab A | 26.4 | |||||
| Veremeta | 587 c 1 | 504 c 1 | 530 c 1 B | 411 b A | 23.9 | |||||
| Mean | 404 | 397 | 372 A | 302 A | 28.8 | |||||
| 2021 | ||||||||||
| Variety | 6 July | 11.00 am | 15.00 pm | % R | 28 July | 11.00 am | 15.00 pm | % R | ||
| Arcos | 150 a | 152 a A | 149 a A | 2.5 | 168 b | 188 b B | 148 ab A | 21.3 | ||
| Bobal | 255 c | 272 c A | 238 c A | 12.8 | 206 d | 222 c B | 190 c A | 14.2 | ||
| Forcallat | 158 a | 176 a B | 139 a A | 21.3 | 141 a | 150 a A | 132 a A | 12.3 | ||
| Garnacha | 225 b | 237 bc A | 212 bc A | 10.9 | 207 d | 204 bc A | 211 c A | 0.0 | ||
| Monastrell | 222 b | 242 bc B | 202 b A | 16.7 | 184 c | 205 bc B | 164 b A | 20.4 | ||
| Veremeta | 212 b | 223 b A | 200 b A | 10.3 | 229 e | 249 d B | 209 c A | 16.3 | ||
| Mean | 204 | 217 B | 190 A | 12.7 | 189 | 203 B | 176 A | 14.1 | ||
| Varieties | ΨStem | An | GS | Ci | E | WUEi |
|---|---|---|---|---|---|---|
| 20 August | ||||||
| Arcos | −1.53 a | 6.67 a | 70.3 a | 209.67 a | 1.97 a | 102.27 d |
| Bobal | −1.35 b | 10.09 b | 153.4 b | 256.10 b | 3.56 b | 70.37 bc |
| Forcallat | −1.61 a | 6.98 a | 75.2 a | 217.33 a | 2.02 a | 97.37 d |
| Garnacha | −1.14 c | 13.73 c | 269.7 c | 276.20 c | 5.55 c | 53.53 a |
| Monastrell | −1.40 b | 9.94 b | 126.3 ab | 239.78 b | 2.93 b | 80.84 c |
| Veremeta | −1.13 c | 15.59 c | 251.8 c | 257.00 b | 5.40 c | 64.04 ab |
| Mean | −1.36 | 10.5 | 157.78 | 242.68 | 3.58 | 78.07 |
| 10 September | ||||||
| Arcos | −1.48 a | 6.09 a | 47.0 a | 165.00 a | 1.10 a | 133.65 c |
| Bobal | −1.12 bc | 10.42 ab | 100.7 ab | 189.67 ab | 1.85 a | 114.80 bc |
| Forcallat | −1.49 a | 8.68 a | 68.4 a | 163.50 a | 1.62 a | 131.33 c |
| Garnacha | −0.99 cd | 14.58 c | 167.1 c | 213.40 bc | 3.01 b | 94.77 ab |
| Monastrell | −1.18 b | 10.47 ab | 104.7 ab | 193.14 ab | 1.80 a | 112.72 bc |
| Veremeta | −0.98 d | 13.32 bc | 161.1 bc | 229.00 c | 2.28 ab | 88.00 a |
| Mean | −1.21 | 10.6 | 108.17 | 192.29 | 1.94 | 112.55 |
| Variety | SD | SL | ED | ECA | SI |
|---|---|---|---|---|---|
| Arcos | 209 a | 22.7 a | 3785 c | 265 b | 6.5 abc |
| Bobal | 205 a | 25.0 c | 2695 a | 372 d | 7.5 cd |
| Forcallat | 280 d | 23.2 a | 3434 b | 295 c | 7.3 bc |
| Garnacha | 236 bc | 26.0 d | 2626 a | 382 d | 8.5 d |
| Monastrell | 257 cd | 24.6 bc | 4280 d | 234 a | 6.2 ab |
| Veremeta | 226 ab | 24.2 b | 4119 cd | 244 ab | 6.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisbert, C.; Soler, J.X.; Fos, M.; Intrigliolo, D.S.; Yuste, A.; Picó, B.; Torrent, D.; Peiró, R. Characterization of Local Mediterranean Grapevine Varieties for Their Resilience to Semi-Arid Conditions under a Rain-Fed Regime. Agronomy 2022, 12, 2234. https://doi.org/10.3390/agronomy12092234
Gisbert C, Soler JX, Fos M, Intrigliolo DS, Yuste A, Picó B, Torrent D, Peiró R. Characterization of Local Mediterranean Grapevine Varieties for Their Resilience to Semi-Arid Conditions under a Rain-Fed Regime. Agronomy. 2022; 12(9):2234. https://doi.org/10.3390/agronomy12092234
Chicago/Turabian StyleGisbert, Carmina, Jaume X. Soler, Mariano Fos, Diego S. Intrigliolo, Alberto Yuste, Belén Picó, Daniel Torrent, and Rosa Peiró. 2022. "Characterization of Local Mediterranean Grapevine Varieties for Their Resilience to Semi-Arid Conditions under a Rain-Fed Regime" Agronomy 12, no. 9: 2234. https://doi.org/10.3390/agronomy12092234
APA StyleGisbert, C., Soler, J. X., Fos, M., Intrigliolo, D. S., Yuste, A., Picó, B., Torrent, D., & Peiró, R. (2022). Characterization of Local Mediterranean Grapevine Varieties for Their Resilience to Semi-Arid Conditions under a Rain-Fed Regime. Agronomy, 12(9), 2234. https://doi.org/10.3390/agronomy12092234