
Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Experimental Design and Irrigation Treatments
2.3. Measurements
2.3.1. Plant Water Status
2.3.2. Plant Growth, Leaf Mass Area and Biomass
2.3.3. Pressure–Volume Curves
2.3.4. Leaf Gas Exchange Measurements
2.4. Statistical Analysis
3. Results
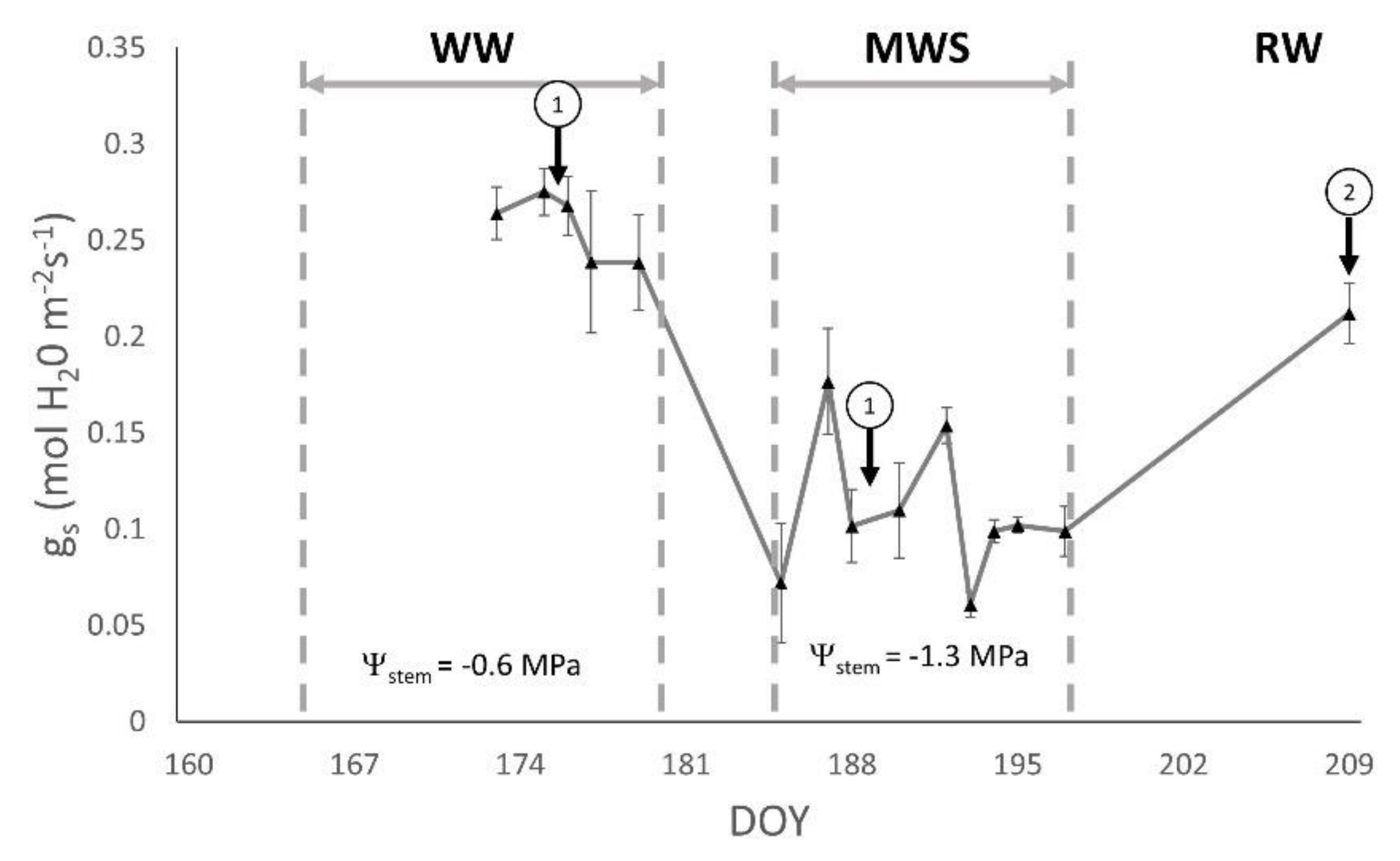
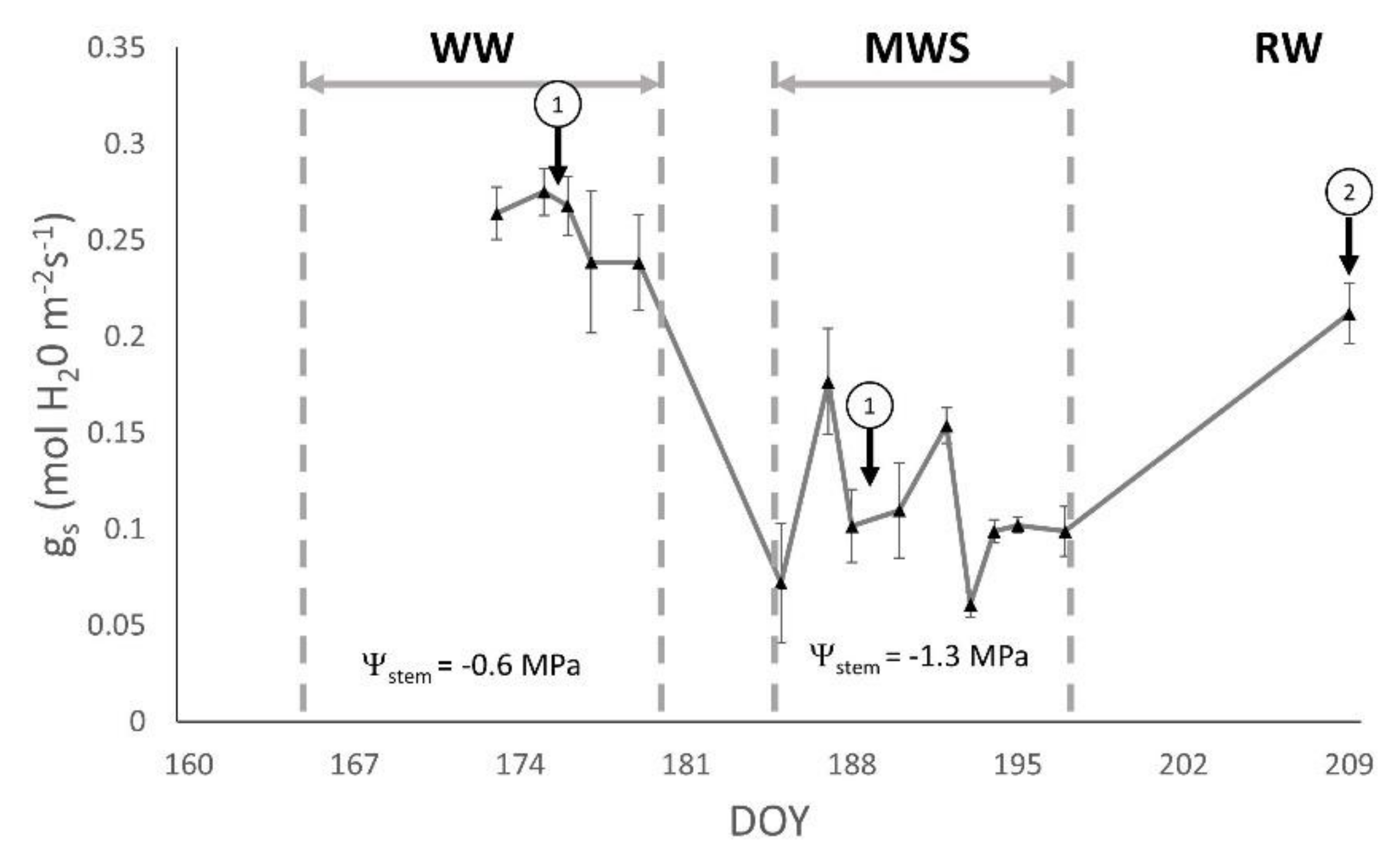
3.1. Plant Water Status
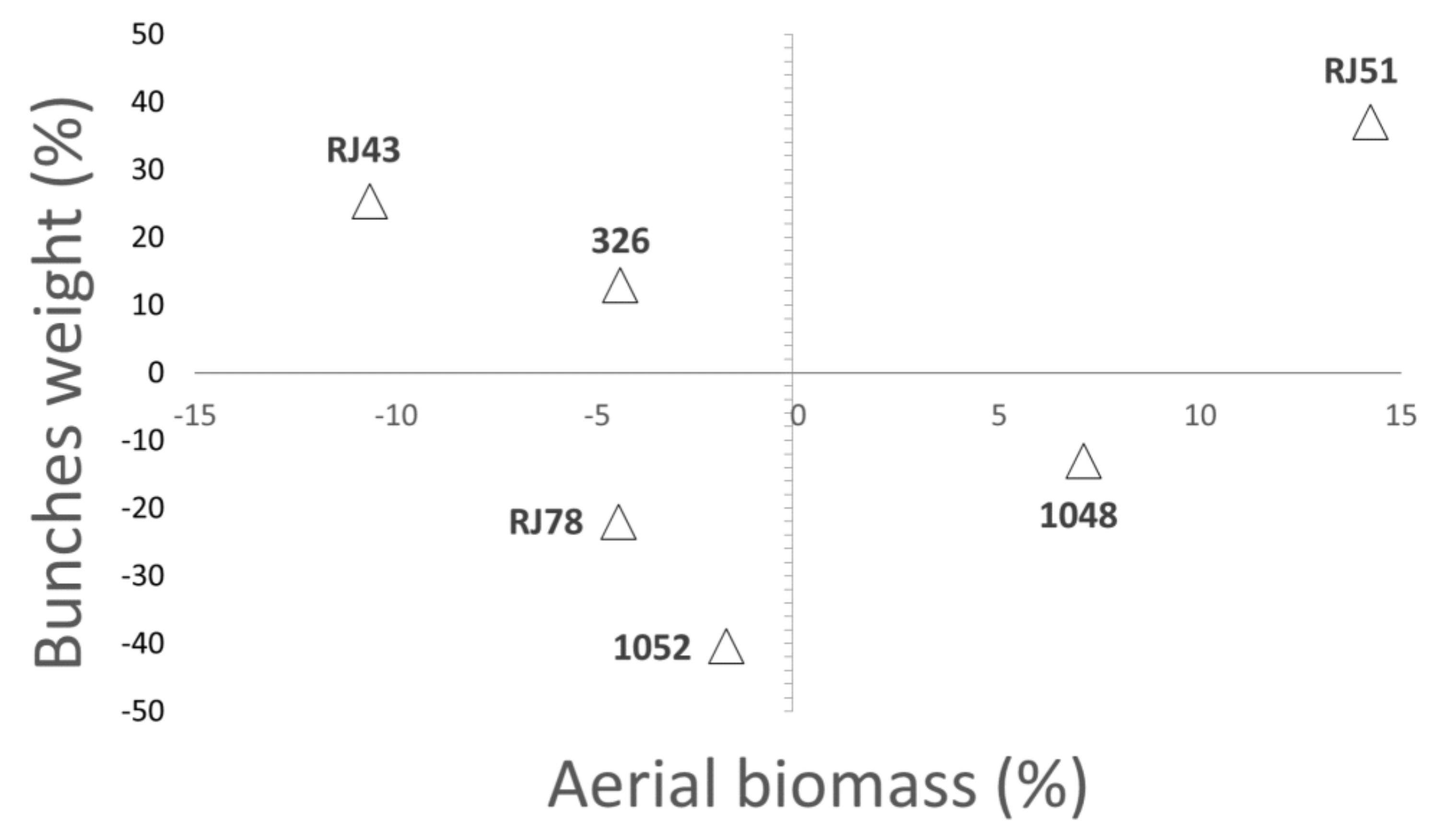
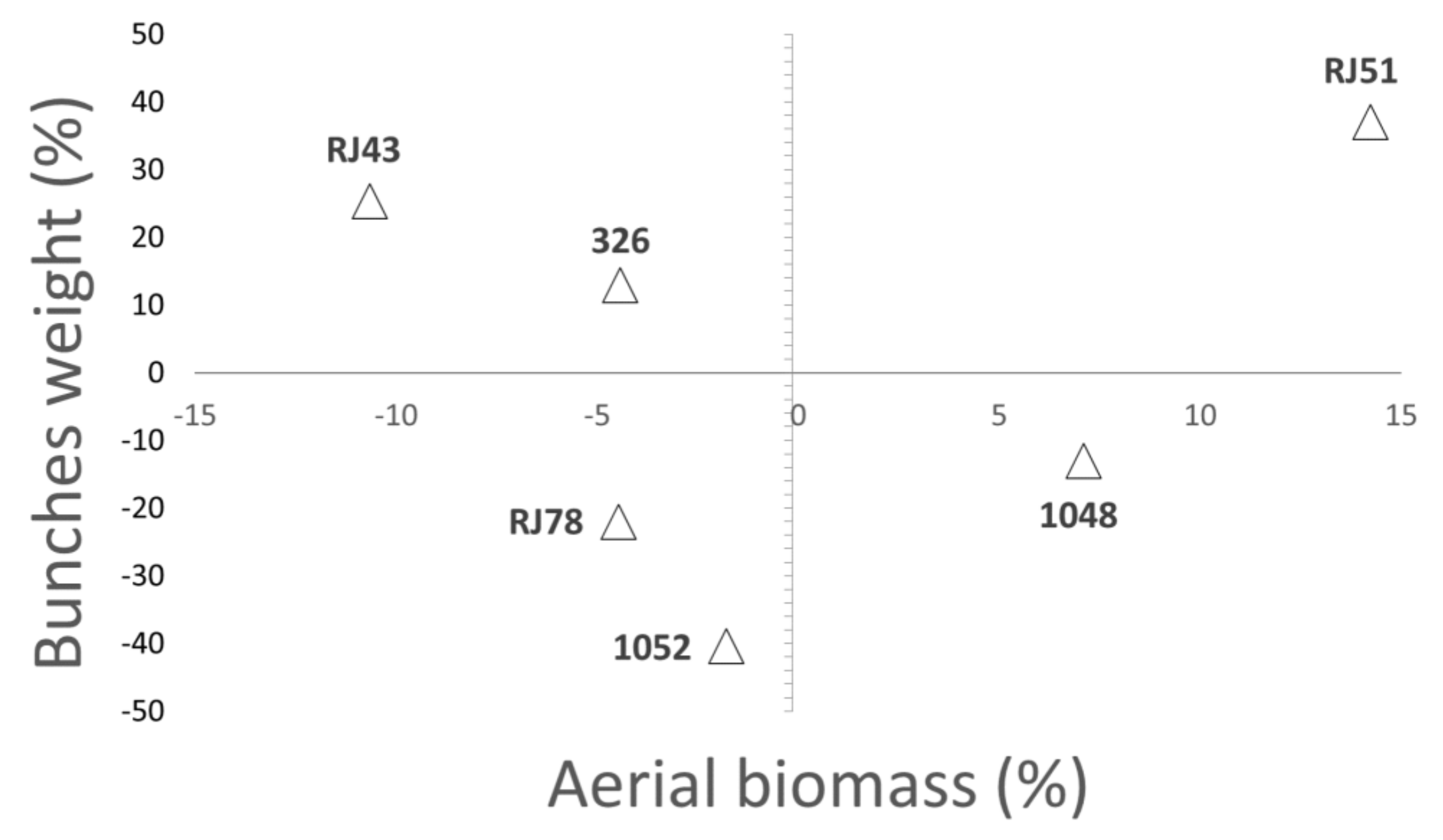
3.2. Plant Growth and Final Biomass
3.3. Hydraulic Traits
3.4. Photosynthetic Traits
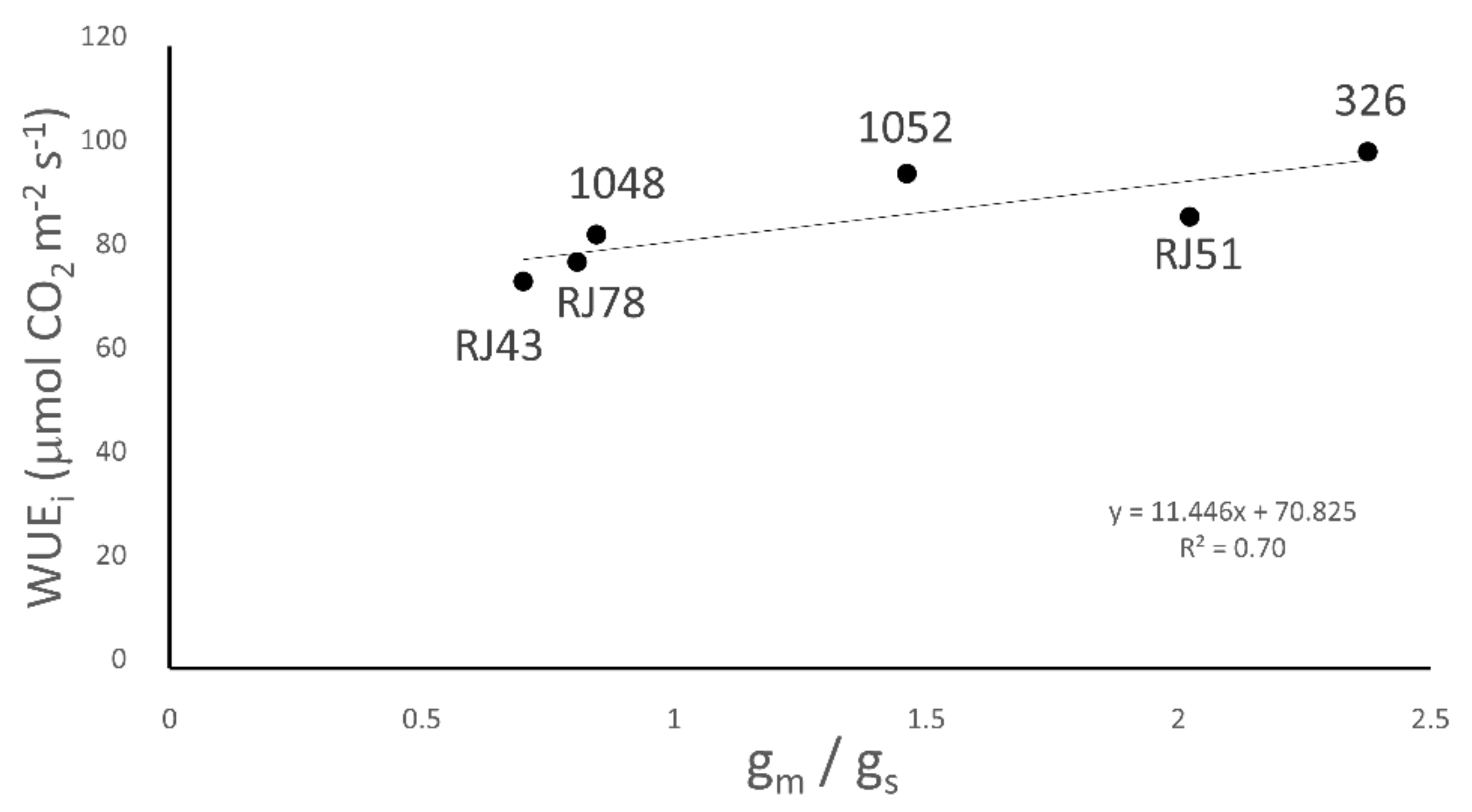
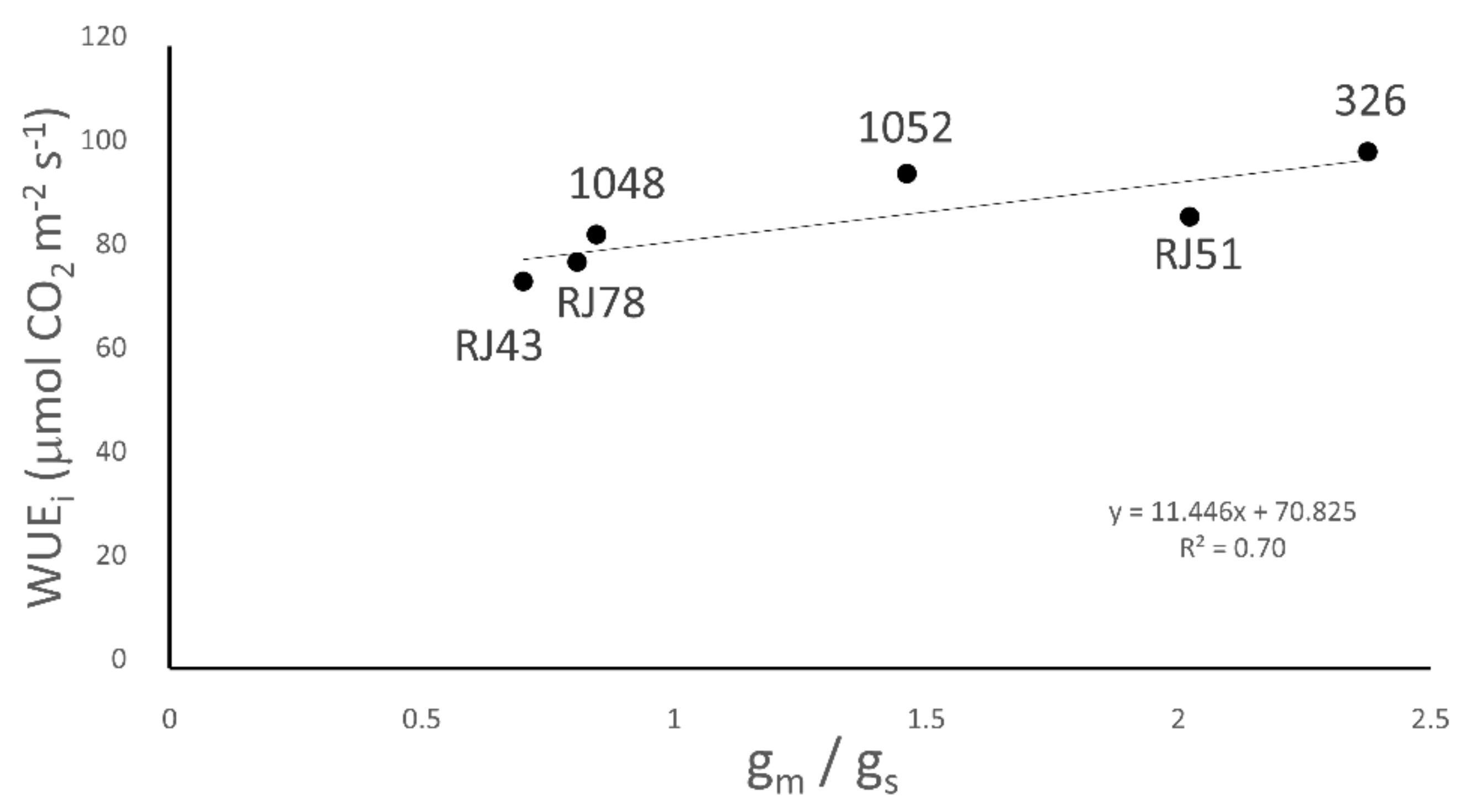
3.5. Water Use Efficiency
4. Discussion
4.1. Plant Water Status, Biomass and Growth Traits
4.2. Hydraulic Traits
4.3. Genotype Variability in Photosynthetic Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Piere, P.; Parker, A.; de Rességuier, L.; Ollat, N. An update on the impact of climate change in viticulture and potential adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Escalona, J.M.; Pou, A.; Fuentes, S.; Flexas, J.; Bota, J. Improving water use efficiency of vineyards in semi-arid regions. A review. Agron. Sustain. Dev. 2015, 35, 499–517. [Google Scholar] [CrossRef]
- Duchêne, E. How can grapevine genetics contribute to the adaptation to climate change? Oeno One 2016, 50, 113–124. [Google Scholar] [CrossRef]
- Delrot, S.; Grimplet, J.; Carbonell-Bejerano, P.; Schwandner, A.; Bert, P.F.; Bavaresco, L.; Dalla, L.; Di Gaspero, G.; Duchêne, E.; Hausman, L.; et al. Genetic and Genomic Approaches for Adaptation of Grapevine to Climate Change. In Genomic Designing of Climate-Smart Fruit Crops; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 157–270. [Google Scholar]
- Galet, P. Dictionnaire Encyclopédique des Cépages (Encyclopaedic Dictionary of Grape Varieties); Hachette: Paris, France, 2000. [Google Scholar]
- Carbonell-Bejerano, P.; Royo, C.; Mauri, N.; Ibáñez, J.; Martínez-Zapater, J.M. Somatic Variation and Cultivar Innovaion in Grapevine. In Advances in Grape and Wine Biotechnology, 1st ed.; Morata, A., Loira, I., Eds.; Intechopen: London, UK, 2016; p. 8. [Google Scholar]
- Ibáñez, J.; Carreño, J.; Yuste, J.; Martínez-Zapater, J.M. Grapevine breeding and clonal selection programmes in Spain. In AG Reynolds; Grapevine Breeding Programs for the Wine Industry; Woodhead Publishing: Oxford, UK, 2015; pp. 183–209. [Google Scholar]
- Martin-Benito, D.; Anchukaitis, K.; Evans, M.; del Río, M.; Beeckman, H.; Cañellas, I. Effects of Drought on Xylem Anatomy and Water-Use Efficiency of Two Co-Occurring Pine Species. Forests 2017, 8, 332. [Google Scholar] [CrossRef]
- Gonzalez de Andres, E.; Camarero, J.J.; Blanco, J.A.; Imbert, J.B.; Lo, Y.H.; Sangüesa-Barreda, G.; Castillo, F.J. Tree-to-tree competition in mixed European beech–Scots pine forests has different impacts on growth and water-use efficiency depending on site conditions. J. Ecol. 2018, 106, 59–75. [Google Scholar] [CrossRef]
- Bota, J.; Tomas, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Pazzagli, P.T.; Weiner, J.; Liu, F. Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agric. Water Manag. 2016, 169, 26–33. [Google Scholar] [CrossRef]
- Meena, R.P.; Karnam, V.; Tripathi, S.C.; Jha, A.; Sharma, R.K.; Singh, G.P. Irrigation management strategies in wheat for efficient water use in the regions of depleting water resources. Agric. Water Manag. 2019, 214, 38–46. [Google Scholar] [CrossRef]
- Fracasso, A.; Trindade, L.M.; Amaducci, S. Drought stress tolerance strategies revealed by RNA-seq in two sorghum genotypes with contrasting WUE. BMC Plant Biol. 2016, 16, 115. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I. Water use efficiency and biomass partitioning of three different Miscanthus genotypes with limited and unlimited water supply. Ann. Bot. 2000, 86, 191–200. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Carneiro, R.L.; Campoe, O.; Alvares, C.A.; Figura, M.A.; Moreira, G.G. Contrasting water use of two Eucalyptus clones across a precipitation and temperature gradient in Brazil. For. Ecol. Manag. 2020, 475, 118407. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.; Bota, J.; Tomas, M.; Hernandez, E.; Escudero, E.; Medrano, H. Exploring the genetic variability in water use efficiency: Evaluation of inter and intra cultivar genetic diversity in grapevines. Plant Sci. 2016, 251, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Tortosa, I.; Douthe, C.; Pou, A.; Balda, P.; Hernandez-Montes, E.; Toro, G.; Escalona, J.M.; Medrano, H. Variability in water use efficiency of grapevine Tempranillo clones and stability over years at field conditions. Agronomy 2019, 9, 701. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Douthe, C.; Pou, A.; Garcia-Escudero, E.; Toro, G.; Medrano, H. The intra-cultivar variability on water use efficiency at different water status as a target selection in grapevine: Influence of ambient and genotype. Agric. Water Manag. 2019, 223, 105648. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Toro, G.; Douthe, C.; Medrano, H. Clonal behaviour in response to soil water availability in Tempranillo Grapevine cv: From plant growth to water use efficiency. Agronomy 2020, 10, 862. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.-M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop. J. 2015, 3, 220–228. [Google Scholar] [CrossRef]
- Yi, K.; Maxwell, J.T.; Wenzel, M.K.; Roman, D.T.; Sauer, P.E.; Phillips, R.P.; Novick, K.A. Linking variation in intrinsic water-use efficiency to isohydricity: A comparison at multiple spatiotemporal scales. New Phytol. 2019, 221, 195–208. [Google Scholar] [CrossRef]
- Kretzler, B.; Rodrigues, C.; Karady, M.; Carmo-Silva, E.; Dodd, I.C. Maintenance of Photosynthesis as Leaves Age Improves Whole Plant Water Use Efficiency in an Australian Wheat Cultivar. Agronomy 2020, 10, 1102. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403.24. [Google Scholar] [CrossRef]
- Hernández-Montes, E.; Escalona, J.M.; Tomás, M.; Martorell, S.; Bota, J.; Tortosa, I.; Medrano, H. Carbon balance in grapevines (Vitis vinifera L.): Effect of environment, cultivar and phenology on carbon gain, losses and allocation. Aust. J. Grape Wine Res. 2022. Online Version of Record before inclusion in an issue. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Brugnoli, E.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbo, M.; Flexas, J. Variability of mesophyll conductance in grapevine cultivars under water stress conditions in relation to leaf anatomy and water use efficiency. Aust. J. Grape Wine Res. 2014, 20, 272–280. [Google Scholar] [CrossRef]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulias, J.; Pou, A.; Ribas, M.; Tomàs, M.; Medrano, H. Improving water use efficiency in grapevines: Potential physiological targets for biotechnological improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Martorell, S.; Diaz-Espejo, A.; Tomas, M.; Pou, A.; El Aou-Ouad, H.; Escalona, J.M.; Vadell, J.; Ribas-Carbo, M.; Flexas, J.; Medrano, H. Differences in water-use-efficiency between two Vitis vinifera cultivars (Grenache and Tempranillo) explained by the combined response of stomata to hydraulic and chemical signals during water stress. Agric. Water Manag. 2015, 156, 1–9. [Google Scholar] [CrossRef]
- Buckley, T.N. How do stomata respond to water status? New Phytol. 2019, 224, 21–36. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Díaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Medrano, H.; Ribas-Carbo, M.; et al. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef]
- Nadal, M.; Flexas, J. Mesophyll conductance to CO2 diffusion: Effects of drought and opportunities for improvement. In Water Scarcity and Sustainable Agriculture in Semiarid Environment; García-Tejero, I.F., Durán-Zuazo, V.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 404–438. [Google Scholar]
- Sack, L.; Pasquet-Kok, J. “Leaf Pressure-Volume Curve Parameters.” Prometheus Wiki Website. Available online: http://prometheuswiki.publish.csiro.au/tikiindex.php (accessed on 20 August 2019).
- Sack, L.; Cowanm, P.D.; Jaikumar, N.; Holbrook, N.M. The ‘hydrology’ of leaves: Co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef]
- Flexas, J.; Díaz-Espejo, A.; Berry, J.A.; Cifre, J.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbó, M. Analysis of leakage in IRGA’s leaf chambers of open gas exchange systems: Quantification and its effects in photosynthesis parameterization. J. Exp. Bot. 2007, 58, 1533–1543. [Google Scholar] [CrossRef]
- Bellasio, C.; Beerling, D.J.; Griffiths, H. Deriving C4 photosynthetic parameters from combined gas exchange and chlorophyll fluorescence using an Excel tool: Theory and practice. Plant Cell Environ. 2016, 39, 1164–1179. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 20 August 2019)ISBN 3-900051-07-0.
- Fox, J.; Weisberg, S. Multivariate linear models. In R. An R Companion to Applied Regression; Sage Publications: Los Angeles, CA, USA,; Thousand Oaks, CA, USA, 2011. [Google Scholar]
- El Aou-ouad, H.; Pou, A.; Tomas, M.; Montero, R.; Ribas-Carbo, M.; Medrano, H.; Bota, J. Combined effect of virus infection and water stress on water flow and water economy in grapevines. Physiol. Plant. 2017, 160, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Buesa, I.; Mirás-Avalos, J.M.; Intrigliolo, D.S. Row orientation effects on potted-vines performance and water-use efficiency. Agric. For. Meteorol. 2020, 294, 108–148. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef] [PubMed]
- McNickle, G.G. Interpreting plant root responses to nutrients, neighbours and pot volume depends on researchers’ assumptions. Funct. Ecol. 2020, 34, 2199–2209. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant–environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef]
- Villalobos-González, L.; Muñoz-Araya, M.; Franck, N.; Pastenes, C. Controversies in the midday water potential regulation and stomatal behauvior might result by the environment, genotype and/or roostock: Evidence from Carménère and Syrah grapevine varieties. Front. Plant Sci. 2019, 10, 1522. [Google Scholar] [CrossRef]
- Dayer, S.; Herrera, J.C.; Zhanwu, D.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G. The sequence and thresholds of leaf hydraulic traits underlying grapevine varietal differences in drought tolerance. J. Exp. Bot. 2020, 71, 4333–4344. [Google Scholar] [CrossRef]
- Alsina, M.M.; Herralde, F.D.; Aranda, X.; Savé, R.; Biel, C. Water relations and vulnerability to embolism are not related: Experiments with eight grapevine cultivars. Vitis 2007, 46, 1–6. [Google Scholar]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The Determinants of Leaf Turgor Loss Point and Prediction of Drought Tolerance of Species and Biomes: A Global Meta-Analysis: Drivers of Plant Drought Tolerance. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Gago, J.; Daloso, D.M.; Carriquí, M.; Nadal, M.; Morales, M.; Araújo, W.L.; Flexas, J. The photosynthesis game is in the” inter-play”: Mechanisms underlying CO2 diffusion in leaves. Environ. Exp. Bot 2020, 178, 104174. [Google Scholar] [CrossRef]
- Flexas, J.; Miquel, R.C.; Antonio, D.E.; Jeroni, G.; Hipolito, M. Mesophyll conductance to CO2: Current knowledge and future prospects. Plant Cell Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Barón, M.; Bota, J.; Ducruet, J.M.; Gallé, A.; Galmés, J.; Jiménez, M.; Pou, A.; Ribas-Carbó, M.; Sajnani, C.; et al. Photosynthesis limitations during water stress acclimation and recovery in the drought–adapted Vitis hybrid Richter–110 (V. berlandierixV. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef]
- Evans, J.R.; Kaldenhoff, R.; Genty, B.; Terashima, I. Resistances along the CO2 diffusion pathway inside leaves. J. Exp. Bot. 2009, 60, 2235–2248. [Google Scholar] [CrossRef]
- Han, J.; Lei, Z.; Flexas, J.; Zhang, Y.; Carriqui, M.; Zhang, W.; Zhang, Y. Mesophyll conductance in cotton bracts: Anatomically determined internal CO2 diffusion constraints on photosynthesis. J. Exp. Bot. 2018, 69, 5433–5443. [Google Scholar] [CrossRef]
- Clemente-Moreno, M.J.; Gago, J.; Daz-Vivancos, P.; Bernal, A.; Miedes, E.; Bresta, P.; Liakopoulos, G.; Fernie, A.R.; Hernández, J.A.; Flexas, J. The apoplastic antioxidant system and altered cell wall dynamics influence mesophyll conductance and the rate of photosynthesis. Plant J. 2019, 99, 1031–1046. [Google Scholar] [CrossRef]
- Roig-Oliver, M.; Nadal, M.; Clemente-Moreno, M.J.; Bota, J.; Flexas, J. Cell wall components regulate photosynthesis and leaf water relations of Vitis vinifera cv. Grenache acclimated to contrasting environmental conditions. J. Plant Physiol. 2020, 244, 153084. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Sisó, S.; Flexas, J.; Galmés, J.; Niinemets, Ü.; Sancho-Knapik, D.; Gil-Pelegrín, E. Coordinated modifications in mesophyll conductance, photosynthetic potentials and leaf nitrogen contribute to explain the large variation in foliage net assimilation rates across Quercus ilex provenances. Tree Physiol. 2017, 37, 1084–1094. [Google Scholar] [CrossRef]
- Mediavilla, S.; Santiago, H.; Escudero, A. Stomatal and mesophyll limitations to photosynthesis in one evergreen and one deciduous Mediterranean oak species. Photosynthetica 2002, 40, 553–559. [Google Scholar] [CrossRef]
- Walker, A.P.; Beckerman, A.P.; Gu, L.; Kattge, J.; Cernusak, L.A.; Domingues, T.F.; Scales, J.C.; Wohlfahrt, G.; Wullschleger, S.D.; Woodward, F.I. The Relationship of Leaf Photosynthetic Traits—Vcmax and Jmax—to Leaf Nitrogen, Leaf Phosphorus, and Specific Leaf Area: A Meta-Analysis and Modeling Study. Ecol. Evol. 2014, 4, 3218–3235. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.G.R.; Thornley, J.H.M. Modeling the components of plant respiration: Some guiding principles. Ann. Bot. 2000, 85, 45–54. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Romero-Munar, A.; Alonso, D.; Aroca, R.; Baraza, E.; Flexas, J.; Ribas-Carbo, M. Respiratory ATP cost and benefit of arbuscular mycorrhizal symbiosis with Nicotiana tabacum at different growth stages and under salinity. J. Plant Physiol. 2017, 218, 243–248. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| gs (mol H2O m−2 s−1) | Ψstem (MPa) | |||
|---|---|---|---|---|
| Genotype | WW | MWS | WW | MWS |
| 326 | 0.292 ± 0.040 | 0.111 ± 0.013 a | −0.60 ± 0.21 | −1.39 ± 0.15 |
| 1048 | 0.258 ± 0.017 | 0.071 ± 0.005 b | −0.74 ± 0.06 | −1.37 ± 0.24 |
| 1052 | 0.241 ± 0.034 | 0.116 ± 0.013 ab | −0.59 ± 0.19 | −1.15 ± 0.20 |
| RJ43 | 0.232 ± 0.024 | 0.092 ± 0.009 ab | −0.62 ± 0.23 | −1.12 ± 0.24 |
| RJ51 | 0.262 ± 0.029 | 0.081 ± 0.009 b | −0.72 ± 0.15 | −1.36 ± 0.18 |
| RJ78 | 0.307 ± 0.043 | 0.101 ± 0.011 ab | −0.61 ± 0.19 | −1.39 ± 0.15 |
| p-value | 0.50 | 0.05 | 0.64 | 0.12 |
| Leaf Expansion Rate (LER) | Leaf Mass Area (LMA) | |||||
|---|---|---|---|---|---|---|
| Genotype | WW (cm2 day−1) | MWS (cm2 day−1) | Reduction % | WW (g m−2) | MWS (g m−2) | Increment (%) |
| 326 | 24.0 ± 6.1 | 11.2 ± 3.6 | 53 ± 20 | 88.9 ± 3.6 | 100.1 ± 2.2 | 13 ± 4 |
| 1048 | 30.0 ± 6.2 | 17.3 ± 5.2 | 42 ± 11 | 80.2 ± 3.4 | 92.6 ± 5.9 | 16 ± 8 |
| 1052 | 36.6 ± 8.0 | 15.1 ± 1.8 | 59 ± 20 | 81.5 ± 4.0 | 104.2 ± 8.6 | 34 ± 14 |
| RJ43 | 27.1 ± 2.8 | 19.5 ± 3.7 | 28 ± 7 | 76.8 ± 2.2 | 85.6 ± 3.2 | 15 ± 5 |
| RJ51 | 31.7 ± 5.4 | 12.5 ± 3.2 | 60 ± 15 | 83.1 ± 5.7 | 90.6 ± 5.8 | 11 ± 9 |
| RJ78 | 26.2 ± 3.8 | 13.7 ± 2.4 | 48 ± 10 | 83.8 ± 5.2 | 92.1 ± 6.1 | 10 ± 5 |
| p-value | 0.66 | 0.6 | 0.42 | 0.35 | 0.46 | 0.28 |
| Genotype | TW/DW | Πo (MPa) | ΨTLP (MPa) | ε (MPa) | sf |
|---|---|---|---|---|---|
| 326 | 2.78 ± 0.18 | −1.39 ± 0.13 | −2.04 ± 0.19 | 14.87 ± 1.22 | 0.31 ± 0.02 |
| 1048 | 2.89 ± 0.10 | −1.44 ± 0.09 | −1.99 ± 0.11 | 13.57 ± 2.41 | 0.41 ± 0.04 |
| 1052 | 2.85 ± 0.02 | −1.44 ± 0.18 | −1.94 ± 0.16 | 15.04 ± 4.48 | 0.41 ± 0.03 |
| RJ43 | 2.86 ± 0.05 | −1.19 ± 0.14 | −1.76 ± 0.11 | 10.93 ± 1.10 | 0.34 ± 0.05 |
| RJ51 | 2.71 ± 0.12 | −1.30 ± 0.16 | −1.76 ± 0.12 | 15.11 ± 3.02 | 0.37 ± 0.05 |
| RJ78 | 2.99 ± 0.09 | −1.14 ± 0.03 | −1.64 ± 0.01 | 11.91 ± 1.52 | 0.30 ± 0.02 |
| p-value | 0.51 | 0.45 | 0.20 | 0.70 | 0.22 |
| Genotype | Amax | Vcmax | Jmax | gm | R |
|---|---|---|---|---|---|
| (µmol CO2 m−2 s−1) | (µmol CO2 m−2 s−1) | (µmol CO2 m−2 s−1) | (mol H2O m−2 s−1) | (µmol CO2 m−2 s−1) | |
| 326 | 34.2 ± 3.0 | 114.5 ± 5.9 a | 120.0 ± 10.4 | 0.180 ± 0.064 ab | 1.00 ± 0.08 bc |
| 1048 | 38.2 ± 4.2 | 76.7 ± 7.8 b | 92.0 ± 11.1 | 0.127 ± 0.013 b | 0.56 ± 0.04 c |
| 1052 | 41.7 ± 2.2 | 80.1 ± 10.0 ab | 90.9 ± 13.2 | 0.187 ± 0.051 ab | 1.00 ± 0.07 bc |
| RJ43 | 33.1 ± 3.8 | 79.6 ± 5.8 b | 108.4 ± 8.9 | 0.095 ± 0.016 b | 1.10 ± 0.05 ab |
| RJ51 | 34.5 ± 4.6 | 72.4 ± 1.6 b | 85.7 ± 1.3 | 0.332 ± 0.082 a | 0.92 ± 0.06 bc |
| RJ78 | 35.5 ± 4.2 | 101.2 ± 10.7 ab | 111.1 ± 6.6 | 0.104 ± 0.011 b | 1.36 ± 0.07 a |
| p-value | 0.57 | 0.046 | 0.088 | 0.013 | 0.031 |
| WUEi (µmol CO2 mol H2O−1) | Whole Plant (g Dry Weight L−1) | ||
|---|---|---|---|
| Genotype | WW | MWS | |
| 326 | 51.4 ± 5.4 | 80.2 ± 4.1 bc | 0.65 ± 0.10 |
| 1048 | 55.0 ± 2.3 | 98.4 ± 3.1 a | 0.65 ± 0.07 |
| 1052 | 52.4 ± 5.2 | 80.5 ± 4.7c | 0.55 ± 0.06 |
| RJ43 | 59.1 ± 3.6 | 81.7 ± 3.8 bc | 0.65 ± 0.08 |
| RJ51 | 54.7 ± 3.8 | 94.9 ± 3.4 ab | 0.79 ± 0.09 |
| RJ78 | 50.6 ± 6.5 | 84.3 ± 4.9 bc | 0.57 ± 0.10 |
| p-value | 0.56 | 0.003 | 0.44 |
| Range | 8 (15%) | 20 (23%) | 0.24 (37%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortosa, I.; Escalona, J.M.; Opazo, I.; Douthe, C.; Medrano, H. Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones. Agronomy 2022, 12, 1874. https://doi.org/10.3390/agronomy12081874
Tortosa I, Escalona JM, Opazo I, Douthe C, Medrano H. Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones. Agronomy. 2022; 12(8):1874. https://doi.org/10.3390/agronomy12081874
Chicago/Turabian StyleTortosa, Ignacio, José Mariano Escalona, Ismael Opazo, Cyril Douthe, and Hipólito Medrano. 2022. "Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones" Agronomy 12, no. 8: 1874. https://doi.org/10.3390/agronomy12081874
APA StyleTortosa, I., Escalona, J. M., Opazo, I., Douthe, C., & Medrano, H. (2022). Genotype Variations in Water Use Efficiency Correspond with Photosynthetic Traits in Tempranillo Grapevine Clones. Agronomy, 12(8), 1874. https://doi.org/10.3390/agronomy12081874