
Ex Vitro Simultaneous Acclimatization and Rooting of In Vitro Propagated Tamarillo Plants (Solanum betaceum Cav.): Effect of the Substrate and Mineral Nutrition



Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Culture
2.2. Rooting, Acclimatization and Treatment with Fertilizers
2.2.1. Dry Biomass
2.2.2. Stomatal Conductance
2.2.3. Chlorophyll Content Index
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boris, D. Les Plantes Alimentaires chez tous les peoples et à travers les âges. In Phanérogames légumières; Lechevalier, P., Ed.; Lechevalier: Paris, Franc, 1927; Volume I, pp. 365–366. [Google Scholar]
- Bohs, L. Crossing studies in Cyphomandra (Solanaceae) and their systematic and evolutionary significance. Am. J. Bot. 1991, 78, 1683–1693. [Google Scholar] [CrossRef]
- Prohens, J.; Nuez, F. The tamarillo (Cyphomandra betacea): A review of a promising small fruit crop. Small Fruits Rev. 2000, 1, 43–68. [Google Scholar] [CrossRef]
- Bakshi, P.; Kour, G.; Ahmed, R. Tamarillo (Cyphomandra betacea). In Underutilized Fruit Crops: Importance and Cultivation, 1st ed.; Ghosh, S.N., Ed.; Jaya Publications: Delhi, India, 2016; pp. 1271–1294. [Google Scholar]
- McCane, J.; Widdowson, D.A. Fruit and nut. In Suppl. to the Composition of Foods, 5th ed.; Unwin and Buss: London, UK, 1992; pp. 74–87. [Google Scholar]
- Kou, M.-C.; Yen, J.-H.; Hong, J.-T.; Wang, C.-L.; Lin, C.-W.; Wu, M.-J. Cyphomandra betacea Sendt. phenolics protect LDL from oxidation and PC12 cells from oxidative stress. LWT-Food Sci. Technol. 2009, 42, 458–463. [Google Scholar] [CrossRef]
- Villegas-Ruiz, X.; Harris, G.K.; Bárcenas-Pozos, M.E.; Jordan, C. Antioxidant and anti-inflammatory properties of tamarillo fruit (Cyphomandra betacea Sendt.) extracts on LPS-activated RAW 264.7 macrophages. FASEB J. 2012, 26, 823–824. [Google Scholar] [CrossRef]
- Noor Atiqah, A.A.K.; Maisarah, A.M.; Asmah, R. Comparison of antioxidant properties of tamarillo (Cyphomandra betacea), cherry tomato (Solanum lycopersicum var. cerasiforme) and tomato (Lycopersicon esculentum). Int. Food Res. J. 2014, 21, 2355–2362. [Google Scholar]
- El-Far, M.M.M. Optimization of growth conditions during sweetpotato micropropagation. In Proceedings of the African Potato Association Conference Proceedings, Alexandria, Egypt, 6–11 July 2007; Volume 7, pp. 204–211. [Google Scholar]
- Dewir, Y.H.; Aldubai, A.A.; Kher, M.M.; Alsadon, A.A.; El-Hendawy, S.; Al-Suhaibani, N.A. Optimization of media formulation for axillary shoot multiplication of the red-peeled sweet potato (Ipomoea batatas [L.] Lam.) “Abees”. Chil. J. Agric. Res. 2020, 80, 3–10. [Google Scholar] [CrossRef]
- Valasevich, N.; Kukharchyk, N.; Krasinskaya, T. Influence of adaptation substrates on morphological development of raspberry plantlets during acclimatizatiom ex vitro. Acta Hortic. 2009, 812, 409–414. [Google Scholar] [CrossRef]
- Peréz, L.P.; Montesinos, Y.P.; Olmedo, J.C.; Rodriguez, R.B.; Sánchez, R.R.; Montenegro, O.N.; Escriba, R.C.R.; Daniels, D.; Gómez-Kosky, R. Effetc oh phloroglucinol on rooting and in vitro acclimatization of papaya (Carica papaya L. var. Maradol Roja). Vitr. Cell. Biol.-Plant 2016, 52, 169–203. [Google Scholar] [CrossRef]
- Hiti Bandaralage, J.C.A.; Hayward, A.; O’Brien, C.; Beveridge, C.; Mitter, N. Acclimatization of micropropagated mature avocado. Acta Hortic. 2018, 1224, 13–20. [Google Scholar] [CrossRef]
- Kozai, T. Acclimatization of micropropagated plants. Biotechnology in Agriculture and Foresty 1991, 17, 127–141. [Google Scholar]
- Pospíšilová, J.; Tichá, I.; Kadlecek, P.; Haisel, D.; Plzáková, Š. Acclimatization of micropropagated plants to ex vitro conditions. Biologia Plantarum 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Pospíšilová, J.; Synková, H.; Haisel, D.; Semorádová, Š. Acclimation of plantlets to ex vitro conditions: Effects of air humidity, irradiance, CO2 concentration and abscisic acid (a Review). Acta Hortic. 2007, 748, 29–38. [Google Scholar] [CrossRef]
- Gonçalves, J.C.; Coelho, M.T.; Diogo, G. Avaliação de parâmetros fotossintéticos em microplantas de Castanheiro durante a fase de aclimatação. Agroforum 2014, 32, 27–40. [Google Scholar]
- Conover, C.A.; Poole, R.T. Acclimatization of indoor foliage plants. Hortic. Rev. 1984, 6, 120–154. [Google Scholar]
- Moreira, M.A.; Carvalho, J.G.; Pasqual, M.; Fráguas, C.B.; Silva, A.B. Acclimatization of micropropagated pineapple plants cv. “Pérola”: Substrate effect. Ciênc. Agrotec. 2005, 30, 875–879. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Rose, R.; Haase, D.L.; Morgan, P.D. Influence of nursery soil amendments on water relations, root architectural development, and field performance of Douglas-fir transplants. New For. 2003, 26, 263–277. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Y.; Shang, Y.; Liu, D.; Liesack, W.; Cui, Z.; Peng, J.; Zhang, F. Peat-vermiculite alters microbiota composition towards increase soil fertility and crop productivity. Plant Soil 2022, 470, 21–34. [Google Scholar] [CrossRef]
- Santos, C.B.; Longhi, S.J.; Hoppe, J.M.; Moscovich, F.A. Effect of container content and kinds of substrate on the quality of Cryptomeria japonica (L.F.) D. Don seedlings. Ciência Florest. 2000, 10, 1–15. [Google Scholar] [CrossRef]
- Betancourt-Osorio, J.; Sánches-Canro, D.; Restrepo-Diáz, H. Effect of nitrogen nutritional statuses and waterlogging conditions on growth parameters, nitrogen use efficiency and chlorophyll fluorescence in Tamarillo seedlings. Not. Bot. Horti Agrobot. 2016, 44, 375–381. [Google Scholar] [CrossRef]
- Oliet, J.; Planelles, R.; Segura, M.L.; Artero, F.; Jacobs, D.F. Mineral nutrition and growth of containerized Pinus halepensis seedlings under controlled-release fertilizer. Sci. Hortic. 2004, 103, 113–129. [Google Scholar] [CrossRef]
- Liu, G.; Zotarelli, L.; Li, Y.; Dinkins, D.; Wang, Q.; Ozores-Hampton, M. 2014 Controlled-Release and Slow-Release Fertilizers as Nutrient Management Tools. UF/IFAS Extention. Available online: https://edis.ifas.ufl.edu/pdffiles/HS/HS125500.pdf (accessed on 22 February 2021).
- Smiderle, O.J.; Souza, A.G.; Menegatti, R.D. Controlled-release fertilizer in the production of seedlings of Anonna cacans Warm. J. Agric. Studies 2020, 8, 125–133. [Google Scholar] [CrossRef][Green Version]
- Fan, Z.; Moore, J.A.; Wenny, D.L. Growth and nutrition of container-grown ponderosa pine seedlings with controlled-release fertilizer incorporated in the root plug. Ann. For. Sci. 2004, 61, 117–124. [Google Scholar] [CrossRef]
- Santos, A.R.; Gonçalves, E.O.; Gibson, E.L.; Araújo, E.F.; Caldeira, M.V.W. Controlled-release fertilizer in the growth of Dalbergia nigra seedlings. Floresta 2020, 50, 1203–1212. [Google Scholar] [CrossRef]
- Haase, D.L.; Rose, R.; Trobaugh, J. Field performance of three stock size of Douglas-fir container seedlings grown with slow-release fertilizer in the nursery growing medium. New For. 2006, 31, 1–24. [Google Scholar] [CrossRef]
- Almeida, L.V.B.; Marinho, C.S.; Muniz, R.A.; Carvalho, A.J.C. Disponibilidade de nutrientes e crescimento de porta-enxertos de citros fertilizados com fertilizantes convencionais e de liberação lenta. Rev. Bras. Frutic. 2012, 34, 289–296. [Google Scholar] [CrossRef][Green Version]
- Rossa, U.B.; Angelo, A.C.; Bognola, I.A.; Westphalen, D.J.; Milani, J.E.F. Fertilização de libertação lenta no crescimento de mudas de Eucalyptus grandis. Floresta 2015, 45, 85–96. [Google Scholar] [CrossRef]
- Obando, M.; Goreux, A.; Jordan, M. Regeneration in vitro de Cyphomandra betacea (tamarillo), una especie frutal andina. Cienc. Investig. Agrar. 1992, 19, 125–130. [Google Scholar] [CrossRef]
- Guimarães, M.L.; Tomé, M.C.; Cruz, G.S. Cyphomandra betacea (Cav.) Sendt. (tamarillo). In Biotechnology in Agriculture and Forestry; Trees, I.V., Bajaj, Y.P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 35, pp. 120–137. [Google Scholar]
- Obando, M.; Jordan, M. Regenerative response of Cyphomandra betacea (Cav.) Sendt (tamarillo) cultivated in vitro. Acta Hortic. 2001, 560, 429–432. [Google Scholar] [CrossRef]
- Lopes, M.L.; Ferreira, M.R.; Carloto, J.M.; Cruz, G.S.; Canhoto, J.M. Somatic embryogenesis induction in tamarillo (Cyphomandra betacea). In Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Newton, R.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Volume 6, pp. 433–455. [Google Scholar]
- Correia, S.; Canhoto, J.M. Somatic embryogenesis of Tamarillo (Solanum betaceum Cav.). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S., Gupta, P., Eds.; Forestry Sciences; Springer: New York, NY, USA, 2018; Volume 85. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lopes, T.M.R. Avaliação, Micropropagação e Seleção de Plantas de Tamarilho (Solanum betaceum Cav.). Master’s Thesis, Biodiversity and Plant Biotechnology, Department of Life Sciences of the Faculty of Sciences and Technology of the University of Coimbra, Coimbra, Portugal, 2019; 80p. [Google Scholar]
- Puga, A.P.N. Desenvolvimento de Biorreatores Para a Propagação de Solanum betaceum Cav. Master’s Thesis, Biodiversity and Plant Biotechnology, Department of Life Sciences of the Faculty of Sciences and Technology of the University of Coimbra, Coimbra, Portugal, 2019; 90p. [Google Scholar]
- Mielke, M.S.; Schaffer, B. Photosynthetic and growth responses of Eugenia uniflora L. seedlings to soil flooding and light intensity. Environ. Exp. Bot. 2010, 68, 113–121. [Google Scholar] [CrossRef]
- Chen, U.C.; Hsia, C.N.; Yeh, M.S.; Agrawal, D.C.; Tsay, H.S. In Vitro micropropagation and ex vitro acclimation of Bupleurum kaoi—An endangered medicinal plant native to Taiwan. Vitr. Cell. Dev. Biol.-Plant 2006, 42, 128–133. [Google Scholar] [CrossRef]
- Hazarika, B.M. Morpho-physiological disorders in in vitro culture of plants. Sci. Hortic. 2006, 108, 105–120. [Google Scholar] [CrossRef]
- Salgado Pirata, M. Estudo do stress hídrico da vinha—Castas Aragonês e Trincadeira. Ph.D. Thesis, Agricultural and Environmental Sciences, Institute of Research and Advanced Training, University of Évora, Évora, Portugal, 2018; 154p. Available online: http://hdl.handle.net/10174/23792 (accessed on 20 January 2021).
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2001. [Google Scholar]
- Argenta, G.; Silva, P.R.F.; Bortolini, C.G.; Forsthofer, E.L.; Strieder, M.L. Relação da leitura do clorofilômetro com os teores de clorofila extraível e de nitrogênio na folha de milho. R. Bras. Fisiol. Veg. 2001, 13, 158–167. [Google Scholar] [CrossRef]
- Esposti, M.D.D.; Siqueira, D.L.; Pereira, P.R.G.; Venegas, V.H.A.; Salomão, L.C.C.; Machado Filho, J.A. Assessment of nitrogenized nutrition of citrus rootstocks using chlorophyll concentrations in the leaf. J. Plant Nutr. 2003, 26, 1287–1299. [Google Scholar] [CrossRef]
- Cole, J.C.; Smith, M.W.; Penn, C.J.; Cheary, B.S.; Conaghan, K.J. Nitrogen, phosphorus, calcium, and magnesium applied individually or as a slow release or controlled release fertilizer increase growth and yield and affect macronutriente and micronutriente concentration and content of field-growth tomato plants. Sci. Hortic. 2016, 211, 420–430. [Google Scholar] [CrossRef]
- Neto, A.A.; Mendes, A.N.G.; Guimarães, P.T.G. Avaliação de substratos alternativos e tipos de adubação para a produção de mudas de cafeeiro (Coffea arabica L.) em tubetes. Ciência Agrotecnologia 1999, 23, 270–280. [Google Scholar]
- Afonso, M.V.; Martinazzo, E.G.; Aumonde, T.Z.; Villela, F.A. Parâmetros fisiológicos de mudas de Albizia niopoides produzidas em diferentes composições de substrato. Ciência Florest. 2017, 27, 1395–1402. [Google Scholar] [CrossRef][Green Version]
- Costa, A.R. Nutrição Mineral em Plantas Vasculares; Notes on plant physiology; School of Science and Technology of the University of Évora: Évora, Portugal, 2014; Available online: http://hdl.handle.net/10174/12007 (accessed on 3 June 2020).
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer Science&Business Media, LLC: Chicago, IL, USA, 2008. [Google Scholar] [CrossRef]
- Ramírez-Soler, C.H.; Magnitskiy, S.; Melo, S.E.; Melgarejo, L.M. Efecto de dosis de nitrógeno, fósforo y potássio sobre el crecimiento del tomate de arból (Solanum betaceum Cav.) en etapa vegetativa. Rev. Columbiana Cienc. Hortícolas 2018, 12, 31–40. [Google Scholar] [CrossRef]
- Clark, C.; Richardson, A. Biomass and mineral nutriente partitioning in a developing tamarillo (Cyphomandra betacea) crop. Sci. Hortic. 2002, 94, 41–51. [Google Scholar] [CrossRef]
- Almeida, L.S.; Maia, N.; Ortega, A.R.; Angelo, A.C. Crescimento de mudas de Jacaranda puberula Cham. em viveiro submetidas a diferentes níveis de luminosidade. Ciência Florest. 2005, 15, 323–329. [Google Scholar] [CrossRef]
- Behling, M.; Neves, J.C.L.; Barros, N.F.; Kishimoto, C.B.; Smit, L. Eficiencia da utilização de nutrientes para formação de raízes finas e médias em povoamento de teca. Rev. Árvore 2014, 38, 837–846. [Google Scholar] [CrossRef]
- Silva, L.D.D.; Lima, A.P.L.; Lima, S.F.; Silva, R.C.; Paniago, G.F. Controlled-release fertilizer in the production and quality of Acacia mangium Seedling. Floresta E Ambiente 2019, 26, e02092017. [Google Scholar] [CrossRef]
- Halloran, S.M.; Adelberg, J. A macronutrient optimization platform for micropropagation and acclimatization: Using turmeric (Curcuma longa L.) as a model plant. Vitr. Cell. Dev. Biol.-Plant 2011, 47, 257–273. [Google Scholar] [CrossRef]
- Monja-Mio, K.M.; Olvera-Casanova, D.; Herrera-Herrera, G.; Herrera-Alamillo, M.A.; Sánchez-Teyer, F.L.; Robert, M.L. Improving of rooting and ex vitro acclimatization phase of Agave tequilana by temporary immersion system (BioMINT™). Vitr. Cell. Dev. Biol.-Plant 2020, 56, 662–669. [Google Scholar] [CrossRef]
- Khai, H.D.; Mai, N.T.N.; Tung, H.T.; Luan, V.Q.; Cuong, D.M.; Ngan, H.T.M.; Chau, N.H.; Buu, N.Q.; Vinh, N.Q.; Dung, D.M.; et al. Selenium nanoparticles as in vitro rooting agent, regulates stomata closure and antioxidant activity of gerbera to tolerance acclimatization. Plant Cell Tiss. Organ Cult. 2022, 1–6. [Google Scholar] [CrossRef]
- Peréz-Jiménez, M.; López-Pérez, A.J.; Otálora-Alcón, G.; Márin-Nicolás, D.; Piñero, M.C.; del Amor, F.M. A regime of high CO2 concentration improves the acclimatization process and increases plant quality and survival. Plant Cell Tiss. Organ Cult. 2015, 121, 547–557. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Substrates | Fertilizer | |||||
|---|---|---|---|---|---|---|
| P | PP | PV | PPV | CRF | QRF | |
| gs | 74.2 ± 28.8 a | 72.5 ± 38.4 a | 61.9 ± 29.4 a | 88.1 ± 72.9 a | 91.6 ± 54.1 a | 56.7 ± 27.0 b |
| CCI | 6.4 ± 2.1 b,c | 5.8 ± 2.1 c | 8.3 ± 2.8 a,b | 9.5 ± 3.1 a | 9.7 ± 2.3 a | 5.3 ± 1.5 b |
| Fertilizer | Dry Biomass | ||
|---|---|---|---|
| Root | Shoot | Total | |
| C | 80.6 ± 30.4 c | 159.2 ± 51.1 b | 119.9 ± 57.5 b |
| QRF | 210.7 ± 57.6 b | 820.3 ± 287.2 a | 515.0 ± 370.7 a |
| CRF | 361.8 ± 111.3 a | 910.0± 319.2 a | 635.9 ± 364.5 a |
| Substrate | Dry Biomass | ||
|---|---|---|---|
| Root | Shoot | Total | |
| P | 196.3 ± 82.3 a,b | 591.6 ± 312.8 a,b | 393.9 ± 301.3 a |
| PP | 176.9 ± 82.7 b | 485.4 ± 370.4 b | 331.1 ± 306.1 a |
| PV | 251.3 ± 191.0 a | 702.8 ± 500.4 a,b | 477.1 ± 436.4 a |
| PPV | 246.2 ± 160.3 a | 739.6 ± 464.0 a | 492.9 ± 422.9 a |
| Sum of | df | Mean | F | p-Value | ||
|---|---|---|---|---|---|---|
| Squares | Square | |||||
| gs | substrate | 5578.9 | 3 | 1859.6 | 0.975 | 0.411 |
| fertilizer | 19,496.3 | 1 | 19,496.3 | 10.219 | 0.002 | |
| substrate × fertilizer | 964.8 | 3 | 321.6 | 0.169 | 0.917 | |
| CCI | substrate | 138.9 | 3 | 46.3 | 31.706 | 0 |
| fertilizer | 311.8 | 1 | 311.8 | 213.455 | 0 | |
| substrate × fertilizer | 7.1 | 3 | 2.4 | 1.615 | 0.196 | |
| Roots | substrate | 48,799.8 | 3 | 16,266.6 | 5.794 | 0.002 |
| dry | fertilizer | 633,841.6 | 2 | 316,920.8 | 112.878 | 0 |
| biomass | substrate × fertilizer | 99,655.7 | 6 | 16,609.3 | 5.916 | 0 |
| Shoots | substrate | 476,560.2 | 3 | 158,853.4 | 3.169 | 0.036 |
| dry | fertilizer | 5,380,192 | 2 | 2,690,096 | 53.664 | 0 |
| biomass | substrate × fertilizer | 524,065.8 | 6 | 87,344.3 | 1.742 | 0.139 |
| Total | substrate | 410,249.5 | 3 | 136,764.8 | 1.505 | 0.219 |
| dry | fertilizer | 4,664,187 | 2 | 2,332,094 | 25.671 | 0 |
| biomass | substrate × fertilizer | 440,497.2 | 6 | 73,416.2 | 0.808 | 0.566 |
| CRF | Dry Biomass | ||
|---|---|---|---|
| Amount | Roots | Shoots | Total |
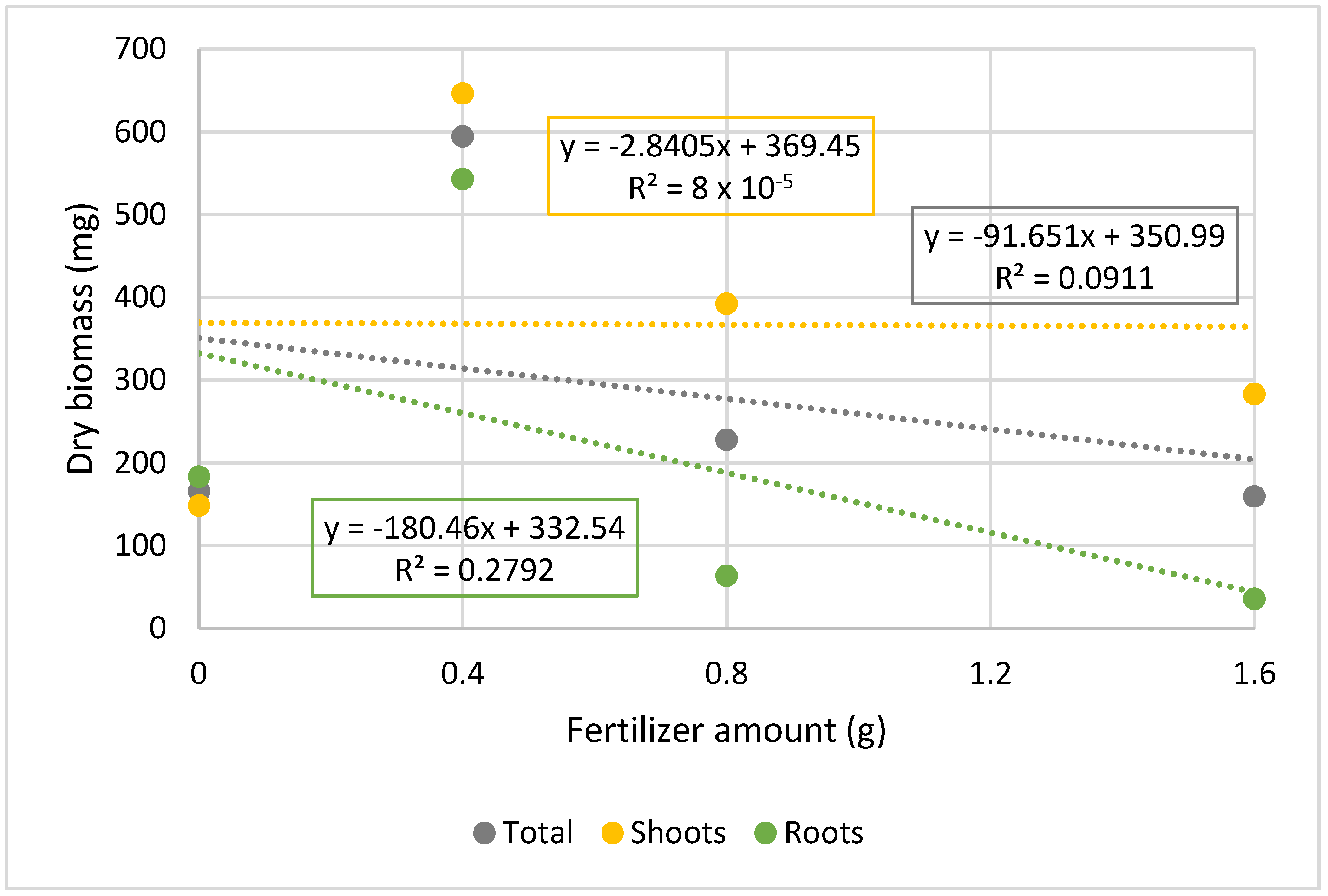
| 0 g | 183.2 ± 52.0 b | 148.5 ± 55.3 a | 165.9 ± 51.7 b |
| 0.4 g | 542.7 ± 138.7 a | 646.3 ± 168.6 a | 594.5 ± 149.3 a |
| 0.8 g | 63.3 ± 44.2 b | 392.2 ± 265.7 a | 227.7 ± 248.0 b |
| 1.6 g | 35.7 ± 36.6 b | 282.9 ± 258.3 a | 159.3 ± 213.4 b |
| CRF | gs | CCI |
|---|---|---|
| Amount | ||
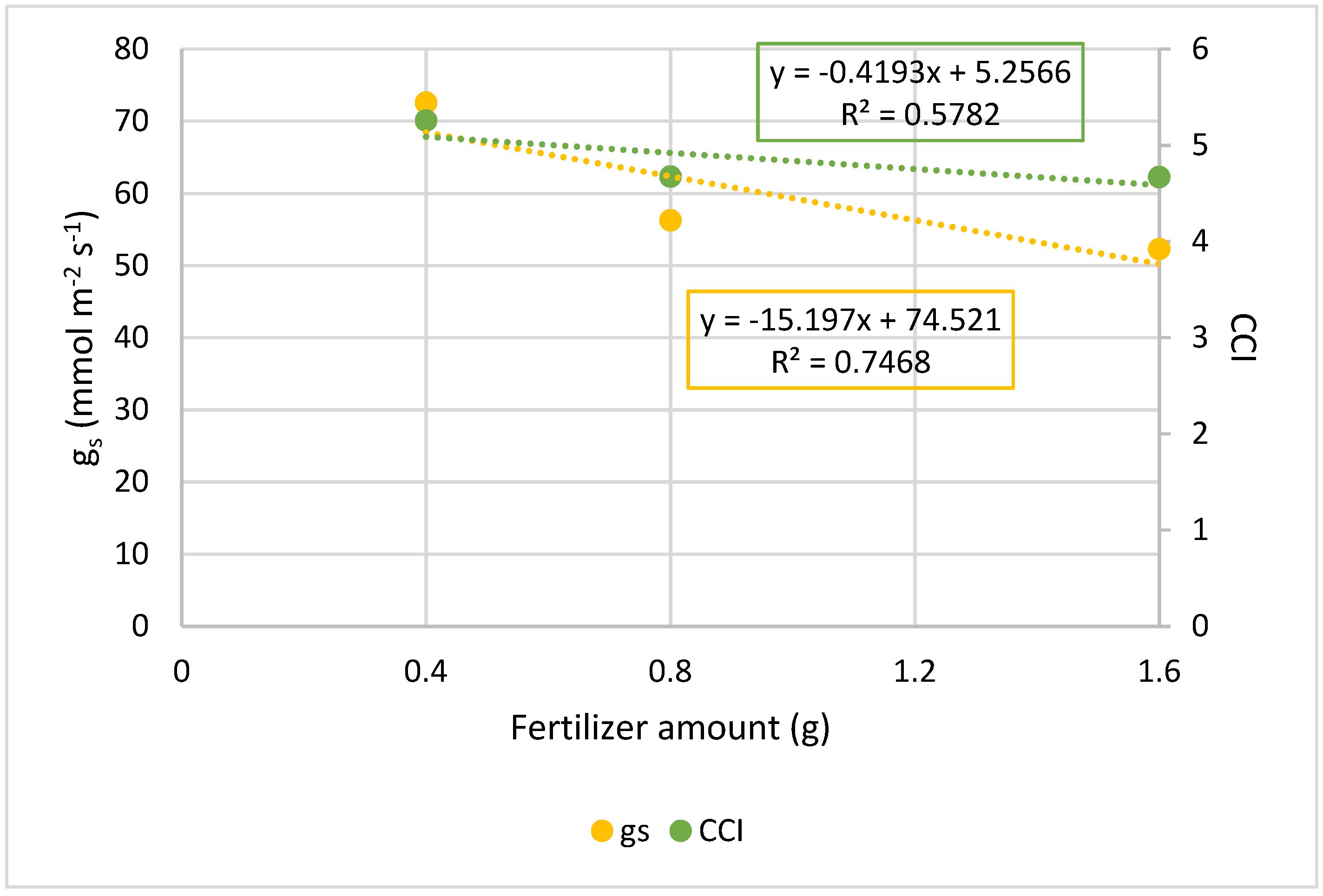
| 0 g * | --- | --- |
| 0.4 g | 72.5 ± 12.9 a | 5.3 ± 0.5 a |
| 0.8 g | 56.2 ± 8.2 b | 4.7 ± 0.3 b |
| 1.6 g | 52.2 ± 12.7 b | 4.7 ± 0.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgado Pirata, M.; Correia, S.; Canhoto, J. Ex Vitro Simultaneous Acclimatization and Rooting of In Vitro Propagated Tamarillo Plants (Solanum betaceum Cav.): Effect of the Substrate and Mineral Nutrition. Agronomy 2022, 12, 1082. https://doi.org/10.3390/agronomy12051082
Salgado Pirata M, Correia S, Canhoto J. Ex Vitro Simultaneous Acclimatization and Rooting of In Vitro Propagated Tamarillo Plants (Solanum betaceum Cav.): Effect of the Substrate and Mineral Nutrition. Agronomy. 2022; 12(5):1082. https://doi.org/10.3390/agronomy12051082
Chicago/Turabian StyleSalgado Pirata, Madalena, Sandra Correia, and Jorge Canhoto. 2022. "Ex Vitro Simultaneous Acclimatization and Rooting of In Vitro Propagated Tamarillo Plants (Solanum betaceum Cav.): Effect of the Substrate and Mineral Nutrition" Agronomy 12, no. 5: 1082. https://doi.org/10.3390/agronomy12051082
APA StyleSalgado Pirata, M., Correia, S., & Canhoto, J. (2022). Ex Vitro Simultaneous Acclimatization and Rooting of In Vitro Propagated Tamarillo Plants (Solanum betaceum Cav.): Effect of the Substrate and Mineral Nutrition. Agronomy, 12(5), 1082. https://doi.org/10.3390/agronomy12051082