
Phytic Acid Content of Faba Beans (Vicia faba)—Annual and Varietal Effects, and Influence of Organic Cultivation Practices



Abstract
1. Introduction
2. Materials and Methods
2.1. Bean Samples
2.2. Chemicals
2.3. Determination of the PA Content
2.3.1. Sample Extraction
2.3.2. Photometric Determination of Total and Free Phosphorus
2.4. Statistical Analysis and Processing
3. Results
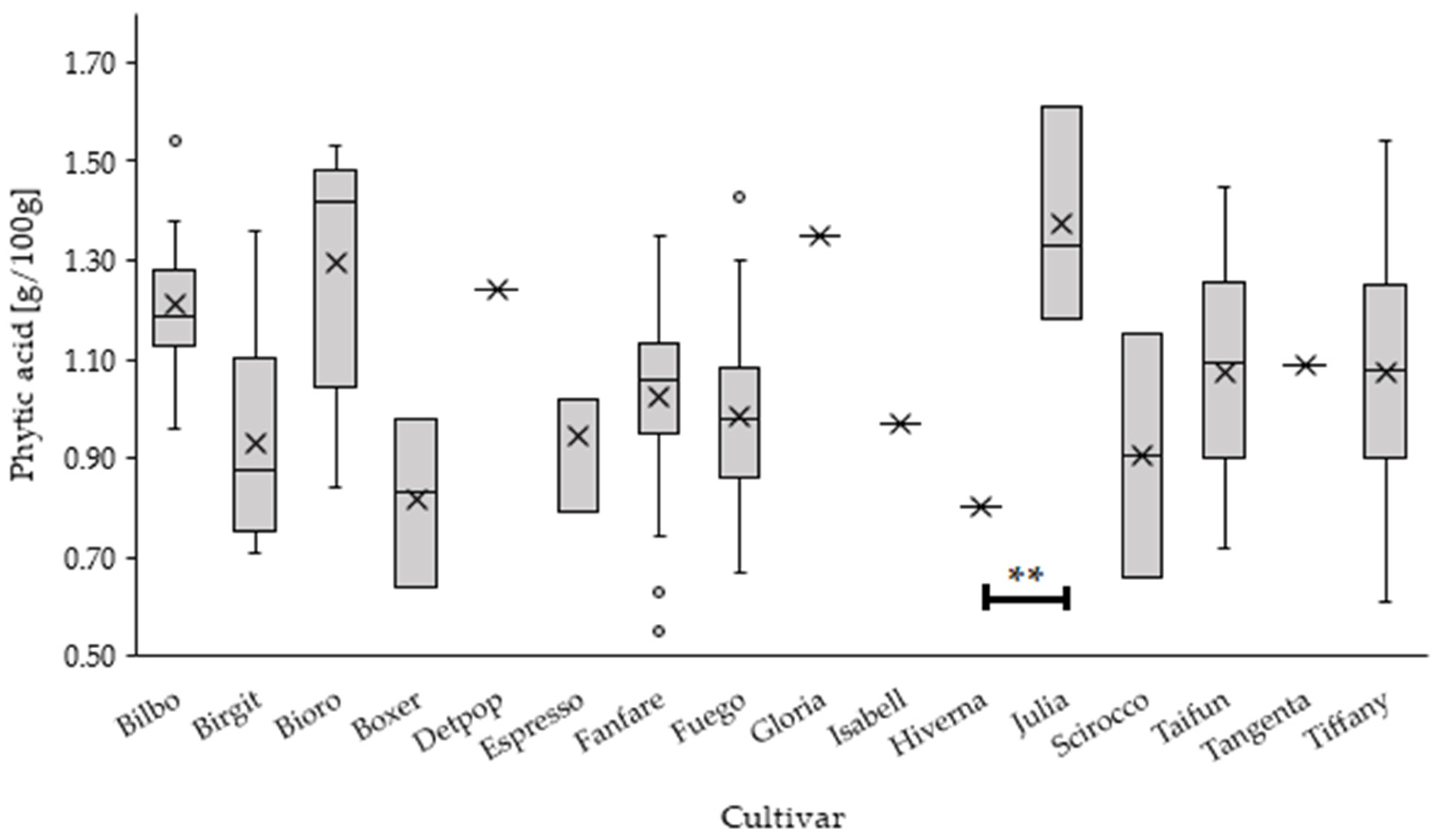
3.1. Influence of the Cultivar on the PA Content
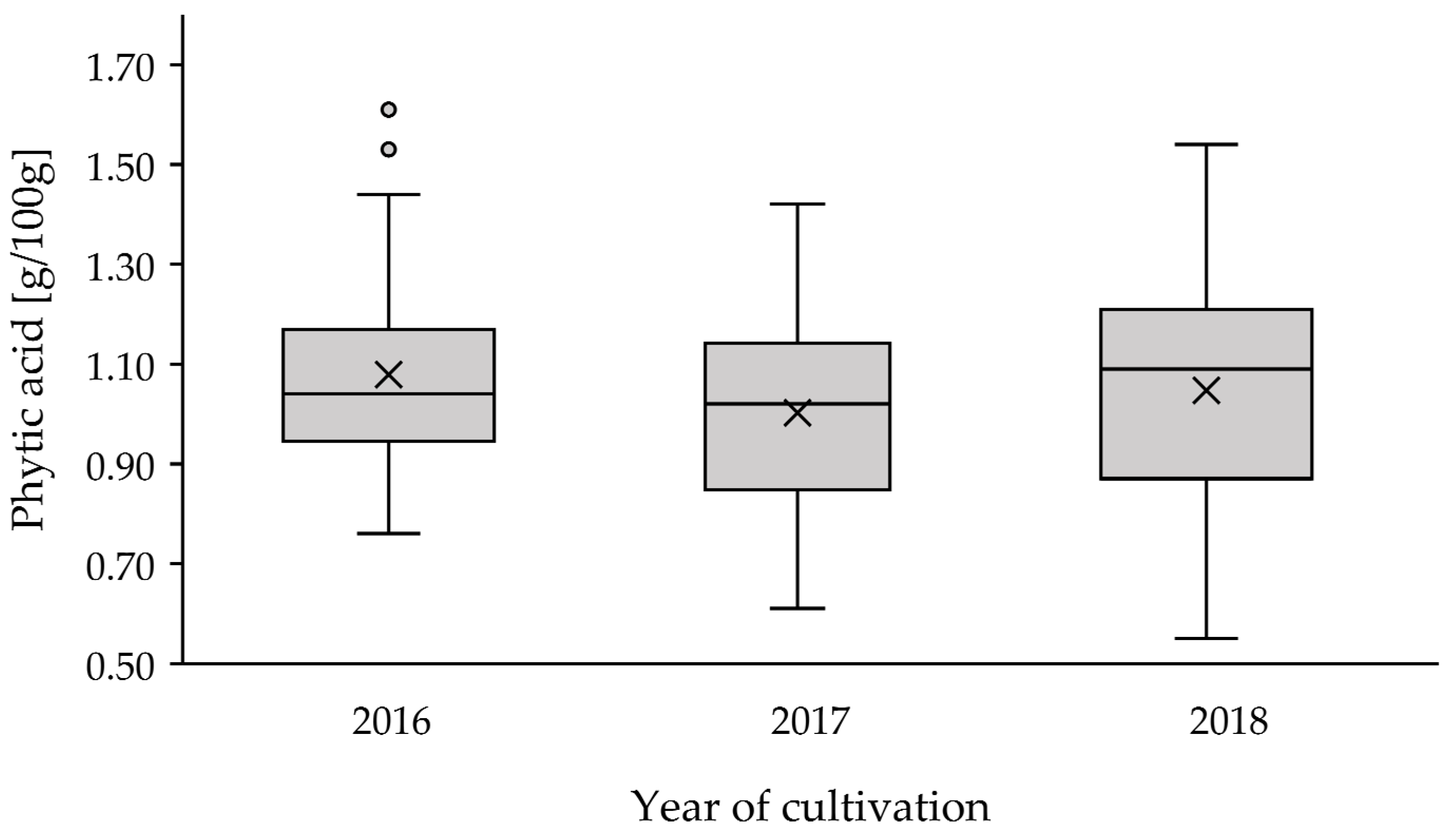
3.2. Annual Effects on PA Content
3.3. Influence of the Cultivation System (Conventional vs. Organic) on the PA Content
3.4. Influence of Fungicide Treatment on Phytic Acid Content
3.5. Influence of Insecticide Treatment on Phytic Acid Content
3.6. Cross-Correlations of Farming System and Fungicide-/Insecticide-Usage
4. Discussion
4.1. Influence of Cultivation Year and Cultivar
4.2. Influence of Farming System (Conventional vs. Organic Cultivation)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in Foods and Significance for Humans: Food Sources, Intake, Processing, Bioavailability, Protective Role and Analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.W.; Thomas, T.A. Phytate and Total Phosphorus Content of Field Beans (Vicia faba L.). J. Sci. Food Agric. 1981, 32, 187–192. [Google Scholar] [CrossRef]
- Steiner, T.; Mosenthin, R.; Zimmermann, B.; Greiner, R.; Roth, S. Distribution of Phytase Activity, Total Phosphorus and Phytate Phosphorus in Legume Seeds, Cereals and Cereal By-Products as Influenced by Harvest Year and Cultivar. Anim. Feed Sci. Technol. 2007, 133, 320–334. [Google Scholar] [CrossRef]
- Reddy, N.R.P.; Sathe, S.K.; Salunkhe, D.K. Occurrence, Distribution, Content and Dietary Intake of Phytate; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Rickard, S.E.; Thompson, L.U. Interactions and Biological Effects of Phytic Acid. In Antinutrients and Phytochemicals in Food; American Chemical Society: Chicago, IL, USA, 1997; Volume 662, pp. 294–312. [Google Scholar]
- Selle, P.H.; Ravindran, V.; Caldwell, A.; Bryden, W.L. Phytate and Phytase: Consequences for Protein Utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in Pig and Poultry Nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 605–625. [Google Scholar] [CrossRef] [PubMed]
- Sheard, L.B.; Tan, X.; Mao, H.; Withers, J.; Ben-Nissan, G.; Hinds, T.R.; Kobayashi, Y.; Hsu, F.F.; Sharon, M.; Browse, J.; et al. Jasmonate Perception by Inositol-Phosphate-Potentiated COI1-JAZ Co-Receptor. Nature 2010, 468, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Calderon-Villalobos, L.I.; Sharon, M.; Zheng, C.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of Auxin Perception by the TIR1 Ubiquitin Ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tang, R.; Zhu, J.; Liu, H.; Mueller-Roeber, B.; Xia, H.; Zhang, H. Enhancement of Stress Tolerance in Transgenic Tobacco Plants Constitutively Expressing AtIpk2beta, an Inositol Polyphosphate 6-/3-kinase from Arabidopsis thaliana. Plant Mol. Biol. 2008, 66, 329–343. [Google Scholar] [CrossRef]
- Winter, C.K.; Davis, S.F. Organic Foods. J. Food Sci. 2006, 71, R117–R124. [Google Scholar] [CrossRef]
- Asami, D.K.; Hong, Y.-J.; Barrett, D.M.; Mitchell, A.E. Comparison of the Total Phenolic and Ascorbic Acid Content of Freeze-Dried and Air-Dried Marionberry, Strawberry, and Corn Grown Using Conventional, Organic, and Sustainable Agricultural Practices. J. Agric. Food Chem. 2003, 51, 1237–1241. [Google Scholar] [CrossRef] [PubMed]
- Caris-Veyrat, C.; Amiot, M.-J.; Tyssandier, V.; Grasselly, D.; Buret, M.; Mikolajczak, M.; Guilland, J.-C.; Bouteloup-Demange, C.; Borel, P. Influence of Organic versus Conventional Agricultural Practice on the Antioxidant Microconstituent Content of Tomatoes and Derived Purees; Consequences on Antioxidant Plasma Status in Humans. J. Agric. Food Chem. 2004, 52, 6503–6509. [Google Scholar] [CrossRef]
- Ren, H.; Endo, H.; Hayashi, T. Antioxidative and Antimutagenic Activities and Polyphenol Content of Pesticide-Free and Organically Cultivated Green Vegetables Using Water-Soluble Chitosan as a Soil Modifier and Leaf Surface Spray. J. Sci. Food Agric. 2001, 81, 1426–1432. [Google Scholar] [CrossRef]
- McKie, V.A.M.; Barry, V. A Novel and Rapid Colorimetric Method for Measuring Total Phosphorus and Phytic Acid in Foods and Animal Feeds. J. AOAC Int. 2016, 99, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.S.; Bewley, J.D. Subcellular Distribution of Phytin in the Endosperm of Developing Castor Bean: A Possibility for Its Synthesis in the Cytoplasm Prior to Deposition within Protein Bodies. Planta 1984, 160, 113–120. [Google Scholar] [CrossRef]
- Cominelli, E.; Pilu, R.; Sparvoli, F. Phytic Acid and Transporters: What Can We Learn from low phytic acid Mutants? Plants 2020, 9, 69. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, B.; Raigond, P.; Sahu, C.; Mishra, U.N.; Sharma, S.; Lal, M.K. Phytic acid: Blessing in Disguise, a Prime Compound Required for Both Plant and Human Nutrition. Food Res. Int. 2021, 142, 110193. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, N.; Ohnishi, M.; Sekiguchi, Y.; Kwon, Y.-U.; Chang, Y.-T.; Chung, S.-K.; Inoue, Y.; Reid, R.J.; Yagisawa, H.; Mimura, T. Phytic Acid Synthesis and Vacuolar Accumulation in Suspension-Cultured Cells of Catharanthus roseus Induced by High Concentration of Inorganic Phosphate and Cations. Plant Physiol. 2005, 138, 1607–1614. [Google Scholar] [CrossRef]
- Raboy, V.; Dickinson, D.B. The Timing and Rate of Phytic Acid Accumulation in Developing Soybean Seeds 1. Plant Physiol. 1987, 85, 841–844. [Google Scholar] [CrossRef]
- Iwai, T.; Takahashi, M.; Oda, K.; Terada, Y.; Yoshida, K.T. Dynamic Changes in the Distribution of Minerals in Relation to Phytic Acid Accumulation during Rice Seed Development. Plant Physiol. 2012, 160, 2007–2014. [Google Scholar] [CrossRef]
- Bhati, K.K.; Aggarwal, S.; Sharma, S.; Mantri, S.; Singh, S.P.; Bhalla, S.; Kaur, J.; Tiwari, S.; Roy, J.K.; Tuli, R.; et al. Differential Expression of Structural Genes for the Late Phase of Phytic Acid Biosynthesis in Developing Seeds of Wheat (Triticum aestivum L.). Plant Sci. 2014, 224, 74–85. [Google Scholar] [CrossRef]
- Thitisaksakul, M.; Jiménez, R.C.; Arias, M.C.; Beckles, D.M. Effects of Environmental Factors on Cereal Starch Biosynthesis and Composition. J. Cereal Sci. 2012, 56, 67–80. [Google Scholar] [CrossRef]
- Bloot, A.P.M.; Kalschne, D.L.; Amaral, J.A.S.; Baraldi, I.J.; Canan, C. A Review of Phytic Acid Sources, Obtention, and Applications. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Becker, K. Dietary Roles of Phytate and Phytase in Human Nutrition: A Review. Food Chem. 2010, 120, 945–959. [Google Scholar] [CrossRef]
- Peksen, E.; Bozoglu, H.; Peksen, A.; Gülümser, A. Determination of the Effects of Different Row Spacings on Yield and Some Other Properties of Pea (Pisum sativum L.) Cultivars Sown in Spring and Autumn. In Proceedings of the II Balkan Symposium on Vegetables and Potatoes, Thessaloniki, Greece, 11–15 October 2000; Volume 579, pp. 313–318. [Google Scholar]
- Walter, S.; Zehring, J.; Mink, K.; Quendt, U.; Zocher, K.; Rohn, S. Protein Content of Peas (Pisum sativum) and Beans (Vicia faba)—Influence of Cultivation Conditions. J. Food Compos. Anal. 2022, 105, 104257. [Google Scholar] [CrossRef]
- Neugschwandtner, R.W.; Böhm, K.; Hall, R.M.; Kaul, H.-P. Development, Growth, and Nitrogen Use of Autumn-and Spring-Sown Facultative Wheat. Acta Agric. Scand. 2015, 65, 6–13. [Google Scholar] [CrossRef]
- Gselman, A.; Kramberger, B. Benefits of Winter Legume Cover Crops Require Early Sowing. Aust. J. Agric. Res. 2008, 59, 1156–1163. [Google Scholar] [CrossRef]
- McCauley, A.M.; Jones, C.A.; Miller, P.R.; Burgess, M.H.; Zabinski, C.A. Nitrogen Fixation by Pea and Lentil Green Manures in a Semi-arid Agroecoregion: Effect of Planting and Termination Timing. Nutr. Cycl. Agroecosyst. 2012, 92, 305–314. [Google Scholar] [CrossRef]
- Mitchell, A.E.; Hong, Y.-J.; Koh, E.; Barrett, D.M.; Bryant, D.E.; Denison, R.F.; Kaffka, S. Ten-Year Comparison of the Influence of Organic and Conventional Crop Management Practices on the Content of Flavonoids in Tomatoes. J. Agric. Food Chem. 2007, 55, 6154–6159. [Google Scholar] [CrossRef]
- Reeve, J.R.; Hoagland, L.A.; Villalba, J.J.; Carr, P.M.; Atucha, A.; Cambardella, C.; Davis, D.R.; Delate, K. Organic Farming, Soil Health, and Food Quality: Considering Possible Links. Adv. Agron. 2016, 137, 319–367. [Google Scholar] [CrossRef]
- Huber, M.; van de Vijver, L.; Parmentier, H.; Savelkoul, H.; Coulier, L.; Wopereis, S.; Verheij, E.; van der Greef, J.; Nierop, D.; Hoogenboom, R.A.P. Effects of Organically and Conventionally Produced Feed on Biomarkers of Health in a Chicken Model. Br. J. Nutr. 2010, 103, 663–676. [Google Scholar] [CrossRef]
- Baranski, M.; Srednicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotic, C.; et al. Higher Antioxidant and Lower Cadmium Concentrations and Lower Incidence of Pesticide Residues in Organically Grown Crops: A Systematic Literature Review and Meta-Analyses. Br. J. Nutr. 2014, 112, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Gershenzon, J. Constitutive Plant Toxins and Their Role in Defense against Herbivores and Pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Mohammadi, M. Effect of Phosphate and Zinc Biofertilizers on Seed Yield and Molar Ratio of Phytic Acid to Zinc in Two Cultivars of Bean (Phaseolus vulgaris L.). Int. J. Agric. Biosyst. Eng. 2017, 11, 66–71. [Google Scholar]
- Chinthapalli, B. A Comparative Study on the Effect of Organic and Inorganic Fertilizers on Agronomic Performance of Faba Bean (Vicia faba L.) and Pea (Pisum sativum L.). Agric. For. Fish. 2015, 4, 263–268. [Google Scholar] [CrossRef]
- Cucci, G.; Lacolla, G.; Summo, C.; Pasqualone, A. Effect of Organic and Mineral Fertilization on Faba Bean (Vicia faba L.). Sci. Hortic. 2019, 243, 338–343. [Google Scholar] [CrossRef]
- Herencia, J.F.; García-Galavís, P.A.; Dorado, J.A.R.; Maqueda, C. Comparison of Nutritional Quality of the Crops Grown in an Organic and Conventional Fertilized Soil. Sci. Hortic. 2011, 129, 882–888. [Google Scholar] [CrossRef]
- Janati, W.; Benmrid, B.; Elhaissoufi, W.; Zeroual, Y.; Nasielski, J.; Bargaz, A. Will Phosphate Bio-Solubilization Stimulate Biological Nitrogen Fixation in Grain Legumes? Front. Agron. 2021, 3, 637196. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S., III; Klein, D.R. Carbon/Nutrient Balance of Boreal Plants in Relation to Vertebrate Herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Berger, J.; Siddique, K.; Loss, S. Cool Season Grain Legumes for Mediterranean Environments: Species× Environment Interaction in Seed Quality Traits and Anti-nutritional Factors in the Genus Vicia. Aust. J. Agric. Res. 1999, 50, 389–402. [Google Scholar] [CrossRef]
- Langenkamper, G.; Zorb, C.; Seifert, M.; Mader, P.; Fretzdorff, B.; Betsche, T. Nutritional Quality of Organic and Conventional Wheat. J. Appl. Bot. Food Qual. 2006, 80, 150. [Google Scholar]
- Madsen, C.K.; Brinch-Pedersen, H. Globoids and Phytase: The Mineral Storage and Release System in Seeds. Int. J. Mol. Sci. 2020, 21, 7519. [Google Scholar] [CrossRef] [PubMed]
- Bohn, L.; Meyer, A.S.; Rasmussen, S.K. Phytate: Impact on Environment and Human Nutrition. A Challenge for Molecular Breeding. J. Zhejiang Univ. Sci. B 2008, 9, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Ito, K.; Yonekura, K.; Tsuda, T.; Ichiyanagi, T.; Hirayama, M.; Konishi, T. Enhanced Absorption of Anthocyanins after Oral Administration of Phytic Acid in Rats and Humans. J. Agric. Food Chem. 2007, 55, 2489–2496. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Luo, H.; Duan, J.; Hong, C.; Ma, P.; Li, G.; Zhang, T.; Wu, T.; Ji, G. Phytic Acid Enhances the Oral Absorption of Isorhamnetin, Quercetin, and Kaempferol in Total Flavones of Hippophae rhamnoides L. Fitoterapia 2014, 93, 216–225. [Google Scholar] [CrossRef]
- Zhai, H.; Wang, F.; Si, Z.; Huo, J.; Xing, L.; An, Y.; He, S.; Liu, Q. A myo-inositol-1-phosphate Synthase Gene, IbMIPS1, Enhances Salt and Drought Tolerance and Stem Nematode Resistance in Transgenic Sweet Potato. Plant Biotechnol. J. 2016, 14, 592–602. [Google Scholar] [CrossRef]
- Murphy, A.M.; Otto, B.; Brearley, C.A.; Carr, J.P.; Hanke, D.E. A Role for Inositol Hexakisphosphate in the Maintenance of Basal Resistance to Plant Pathogens. Plant J. 2008, 56, 638–652. [Google Scholar] [CrossRef]
- Sharma, S.; AnandKumar, L.H.D.; Tyagi, A.; Muthumilarasan, M.; Kumar, K.; Gaikwad, K. An Insight into Phytic Acid Biosynthesis and Its Reduction Strategies to Improve Mineral Bioavailability. Nucleus 2021. [Google Scholar] [CrossRef]
- Zhang, Z.; Liao, H.; Lucas, W.J. Molecular Mechanisms Underlying Phosphate Sensing, Signaling, and Adaptation in Plants. J. Integr. Plant Biol. 2014, 56, 192–220. [Google Scholar] [CrossRef]
- Luo, Y.; Qin, G.; Zhang, J.; Liang, Y.; Song, Y.; Zhao, M.; Tsuge, T.; Aoyama, T.; Liu, J.; Gu, H.; et al. D-myo-inositol-3-Phosphate Affects Phosphatidylinositol-Mediated Endomembrane Function in Arabidopsis and Is Essential for Auxin-Regulated Embryogenesis. Plant Cell 2011, 23, 1352–1372. [Google Scholar] [CrossRef]
- Silva, V.M.; Putti, F.F.; White, P.J.; Reis, A.R.D. Phytic Acid Accumulation in Plants: Biosynthesis Pathway Regulation and Role in Human Diet. Plant Physiol. Biochem. 2021, 164, 132–146. [Google Scholar] [CrossRef]
- Raboy, V. The ABCs of Low-Phytate Crops. Nat. Biotechnol. 2007, 25, 874–875. [Google Scholar] [CrossRef] [PubMed]
- Sparvoli, F.; Cominelli, E. Seed Biofortification and Phytic Acid Reduction: A Conflict of Interest for the Plant? Plants 2015, 4, 728–755. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, D.H.; Cho, H.K.; Kim, S.H.; Auh, J.H.; Pai, H.S. InsP6-Sensitive Variants of the Gle1 mRNA Export Factor Rescue Growth and Fertility Defects of the ipk1 Low-Phytic-Acid Mutation in AR-Abidopsis. Plant Cell 2015, 27, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Redekar, N.R.; Biyashev, R.M.; Jensen, R.V.; Helm, R.F.; Grabau, E.A.; Maroof, M.A. Genome-Wide Transcriptome Analyses of Developing Seeds from Low and Normal Phytic Acid Soybean Lines. BMC Genomics 2015, 16, 1074. [Google Scholar] [CrossRef]
- Raboy, V. Approaches and Challenges to Engineering Seed Phytate and Total Phosphorus. Plant Sci. 2009, 177, 281–296. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cultivar | n | Mean ± SD | Min | Max |
|---|---|---|---|---|
| Bilbo | 10 | 1.21 ± 0.16 | 0.96 | 1.54 |
| Birgit | 8 | 0.93 ± 0.23 | 0.71 | 1.36 |
| Bioro | 5 | 1.30 ± 0.27 | 0.84 | 1.53 |
| Boxer | 3 | 0.82 ± 0.17 | 0.64 | 0.98 |
| Detpop | 1 | 1.24 | - | - |
| Espresso | 3 | 0.94 ± 0.13 | 0.79 | 1.02 |
| Fanfare | 43 | 1.02 ± 0.17 | 0.55 | 1.35 |
| Fuego | 45 | 0.99 ± 0.18 | 0.67 | 1.43 |
| Gloria | 1 | 1.35 | - | - |
| Isabell | 1 | 0.97 | - | - |
| Hiverna | 1 | 0.80 | - | - |
| Julia | 3 | 1.37 ± 0.22 | 1.18 | 1.61 |
| Scirocco | 2 | 0.91 ± 0.35 | 0.66 | 1.15 |
| Taifun | 12 | 1.07 ± 0.23 | 0.72 | 1.45 |
| Tangenta | 1 | 1.09 | - | - |
| Tiffany | 23 | 1.07 ± 0.25 | 0.61 | 1.54 |
| total | 162 | 1.04 ± 0.21 | 0.55 | 1.61 |
| Year | n (Samples) | n (Cultivars) | Mean ± SD | Min | Max |
|---|---|---|---|---|---|
| 2016 | 57 | 11 | 1.08 ± 0.20 | 0.76 | 1.61 |
| 2017 | 66 | 12 | 1.00 ± 0.21 | 0.61 | 1.42 |
| 2018 | 39 | 8 | 1.05 ± 0.24 | 0.55 | 1.54 |
| total | 162 | 16 | 1.04 ± 0.21 | 0.55 | 1.61 |
| Cultivation System | n | Mean ± SD | Min | Max |
|---|---|---|---|---|
| Conventional * | 115 | 0.99 ± 0.19 | 0.55 | 1.45 |
| Organic * | 47 | 1.16 ± 0.23 | 0.66 | 1.61 |
| total | 162 | 1.04 ± 0.21 | 0.55 | 1.61 |
| Fungicide Usage | n | Mean ± SD | Min | Max |
|---|---|---|---|---|
| yes * | 75 | 0.96 ± 0.18 | 0.55 | 1.45 |
| no * | 77 | 1.11 ± 0.22 | 0.63 | 1.61 |
| total | 152 | 1.04 ± 0.22 | 0.55 | 1.61 |
| Insecticide Usage | n | Mean ± SD | Min | Max |
|---|---|---|---|---|
| yes * | 75 | 0.97 ± 0.19 | 0.55 | 1.45 |
| no * | 83 | 1.11 ± 0.21 | 0.66 | 1.61 |
| total | 158 | 1.04 ± 0.21 | 0.55 | 1.61 |
| No. | Factor/Criteria | n | df | PA (Mean Squares) | Significance (p) |
|---|---|---|---|---|---|
| 1 | no fungicide × both farming systems (conv vs. org) | 77 | 1 | 0.273 | 0.018 * |
| 2 | no insecticide x both farming systems (conv vs. org) | 83 | 1 | 0.303 | 0.009 ** |
| 3 | organic × fungicide | 0 | / | / | / |
| 4 | organic × insecticide | 1 | 1 | (0.005) | (0.754 ns) |
| 5 | conventional × fungicide | 105 | 1 | 0.085 | 0.087 ns |
| 6 | conventional × insecticide | 111 | 1 | 0.076 | 0.114 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zehring, J.; Walter, S.; Quendt, U.; Zocher, K.; Rohn, S. Phytic Acid Content of Faba Beans (Vicia faba)—Annual and Varietal Effects, and Influence of Organic Cultivation Practices. Agronomy 2022, 12, 889. https://doi.org/10.3390/agronomy12040889
Zehring J, Walter S, Quendt U, Zocher K, Rohn S. Phytic Acid Content of Faba Beans (Vicia faba)—Annual and Varietal Effects, and Influence of Organic Cultivation Practices. Agronomy. 2022; 12(4):889. https://doi.org/10.3390/agronomy12040889
Chicago/Turabian StyleZehring, Jenny, Sinja Walter, Ulrich Quendt, Kathleen Zocher, and Sascha Rohn. 2022. "Phytic Acid Content of Faba Beans (Vicia faba)—Annual and Varietal Effects, and Influence of Organic Cultivation Practices" Agronomy 12, no. 4: 889. https://doi.org/10.3390/agronomy12040889
APA StyleZehring, J., Walter, S., Quendt, U., Zocher, K., & Rohn, S. (2022). Phytic Acid Content of Faba Beans (Vicia faba)—Annual and Varietal Effects, and Influence of Organic Cultivation Practices. Agronomy, 12(4), 889. https://doi.org/10.3390/agronomy12040889