
Resting Subtropical Grasslands from Grazing in the Wet Season Boosts Biocrust Hotspots to Improve Soil Health

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
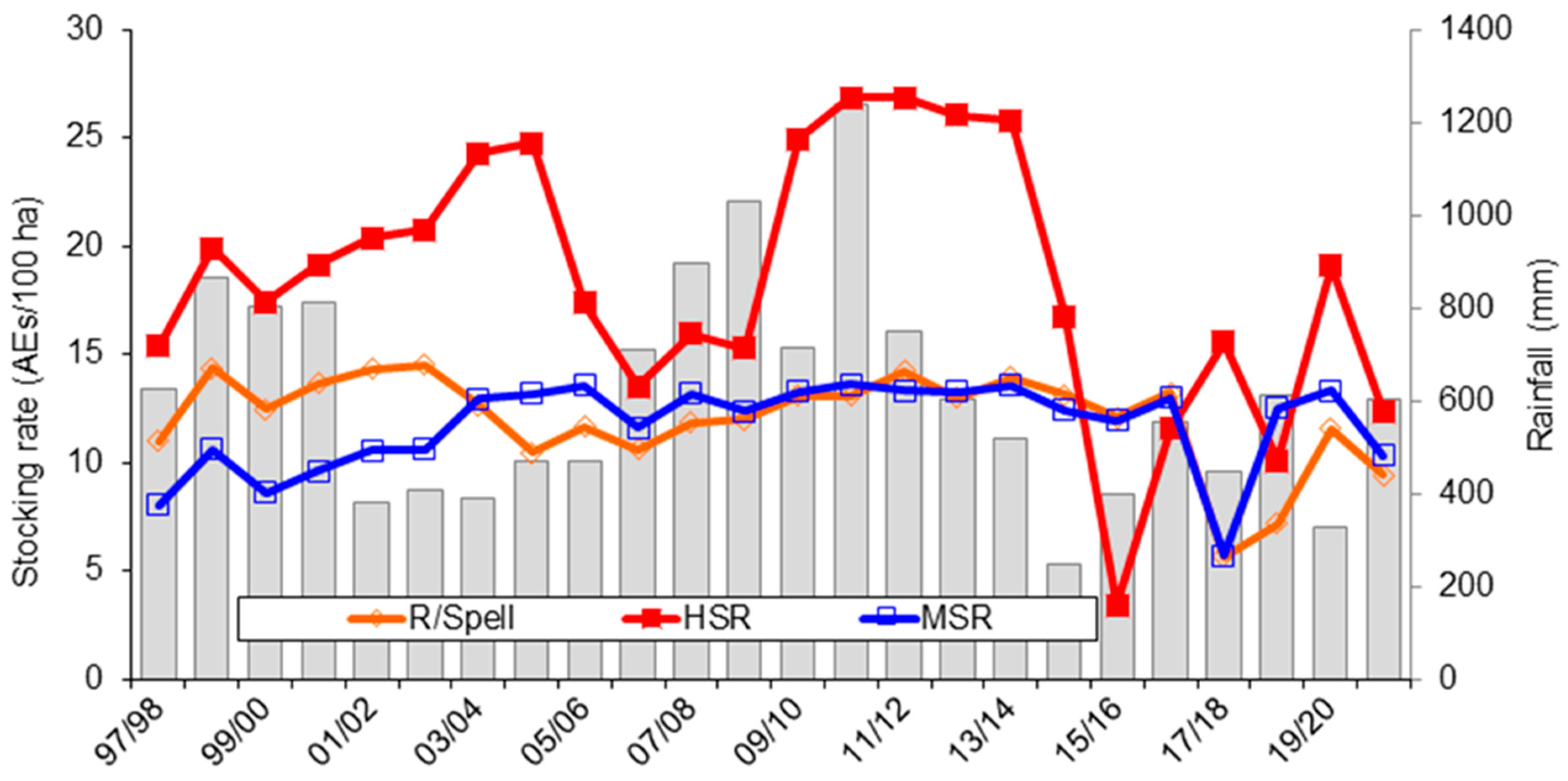
2.1. Site Background
2.2. Field Methods
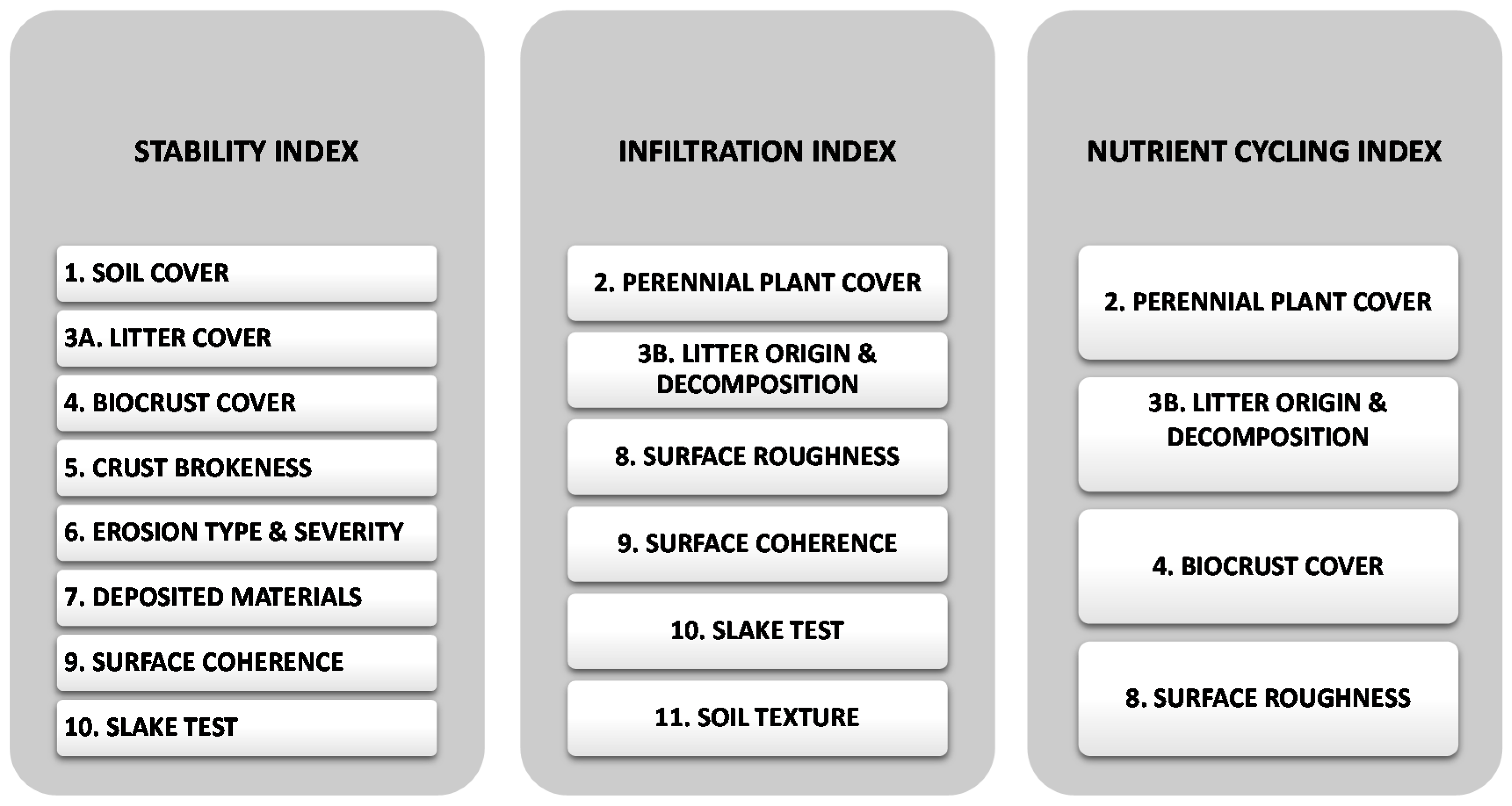
2.2.1. Landscape Function
2.2.2. Ground Cover
2.3. Biocrust Structure
Scanning Electron Microscope (SEM) Imaging
2.4. Statistical Analysis
3. Results
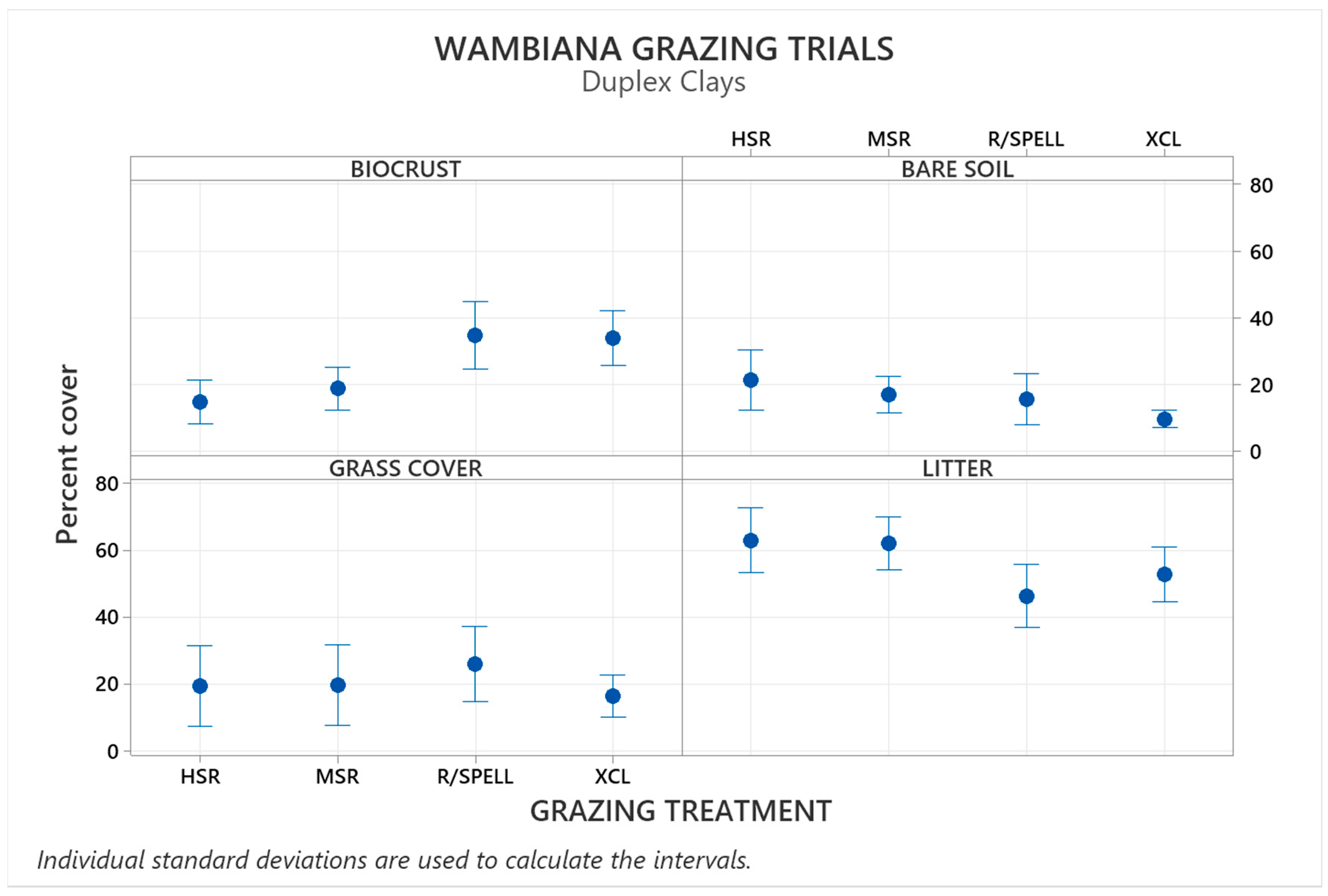
3.1. Biocrust Hotspots in Duplex Soils
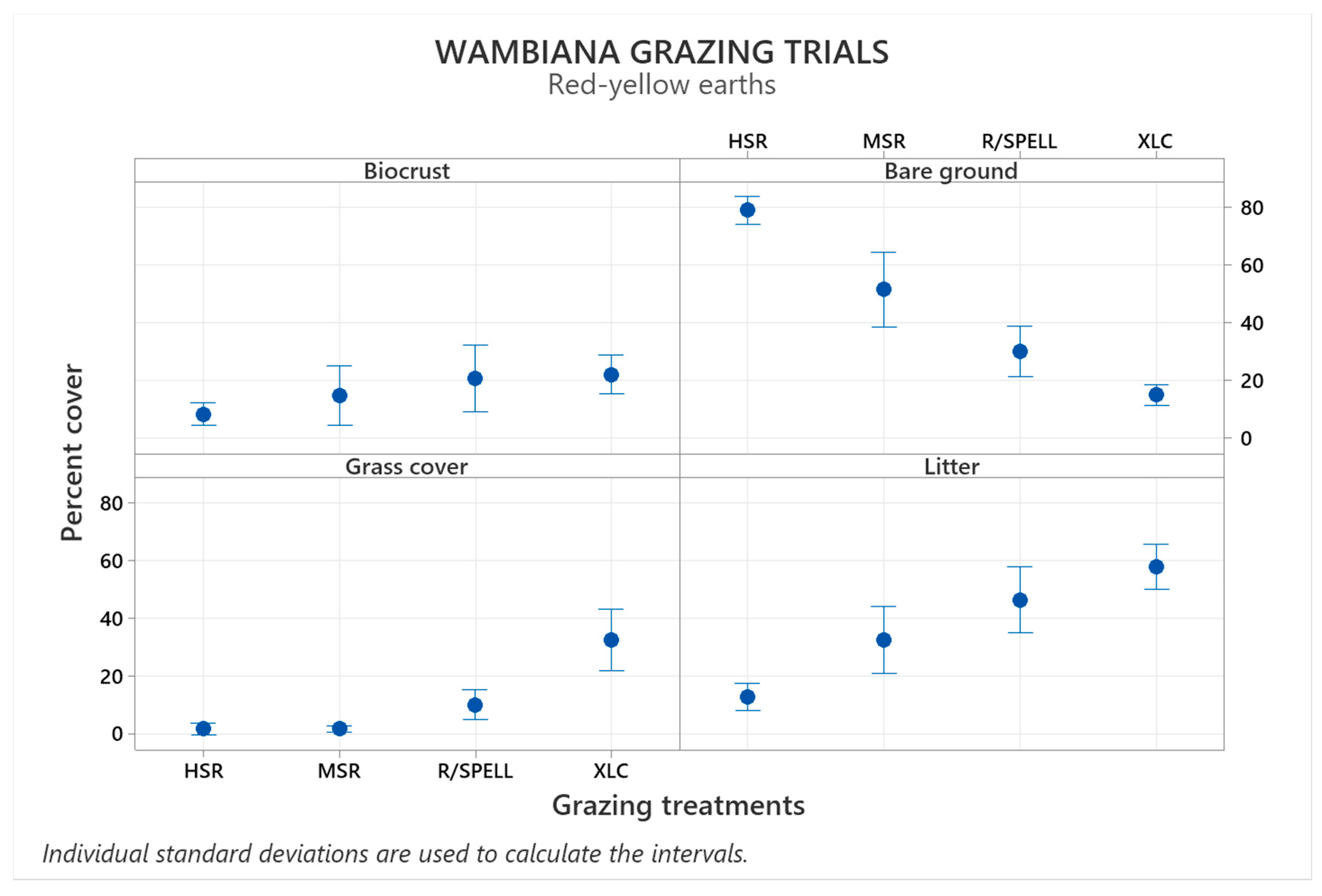
3.2. Biocrusts in Red-Yellow Earths
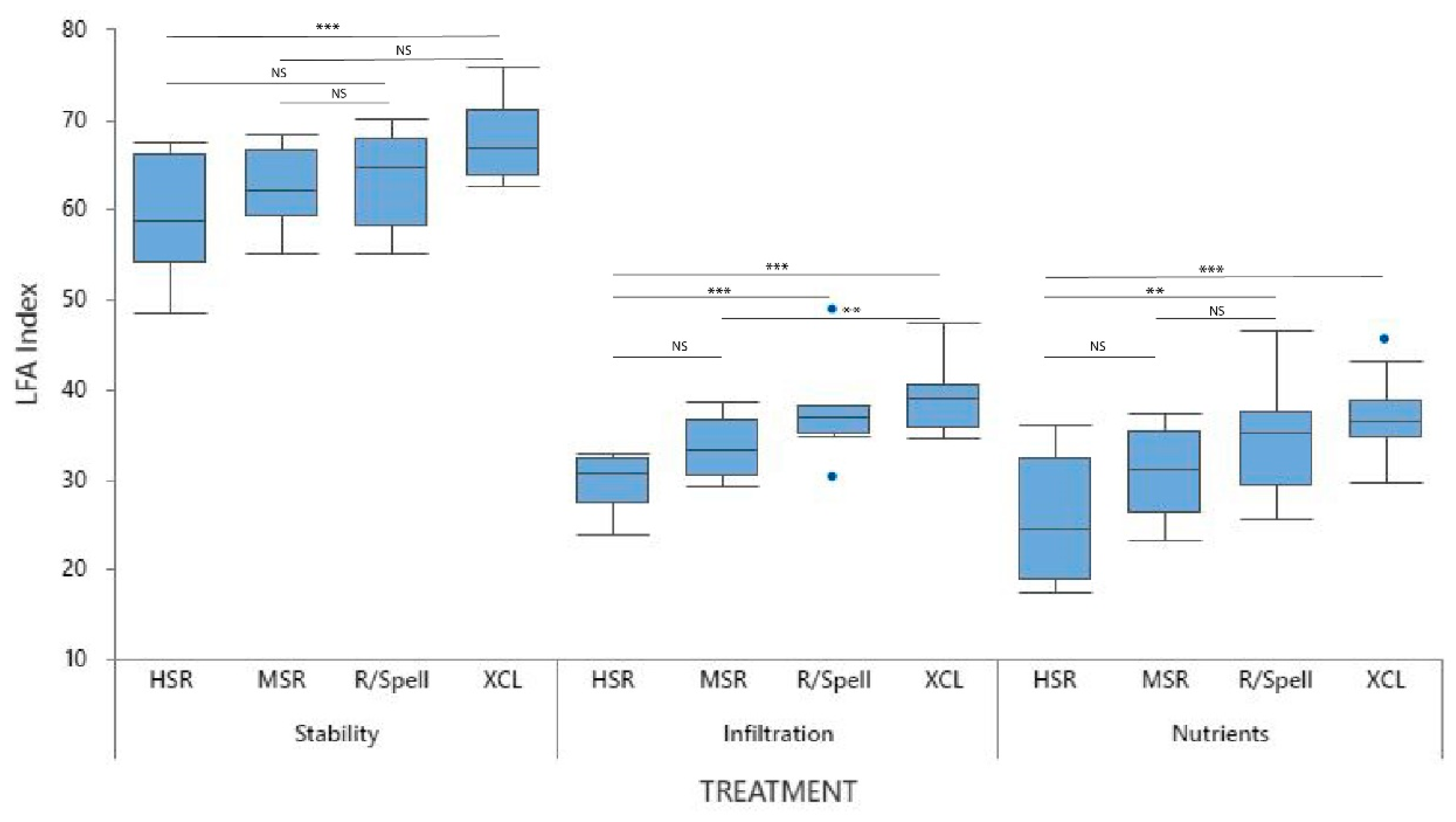
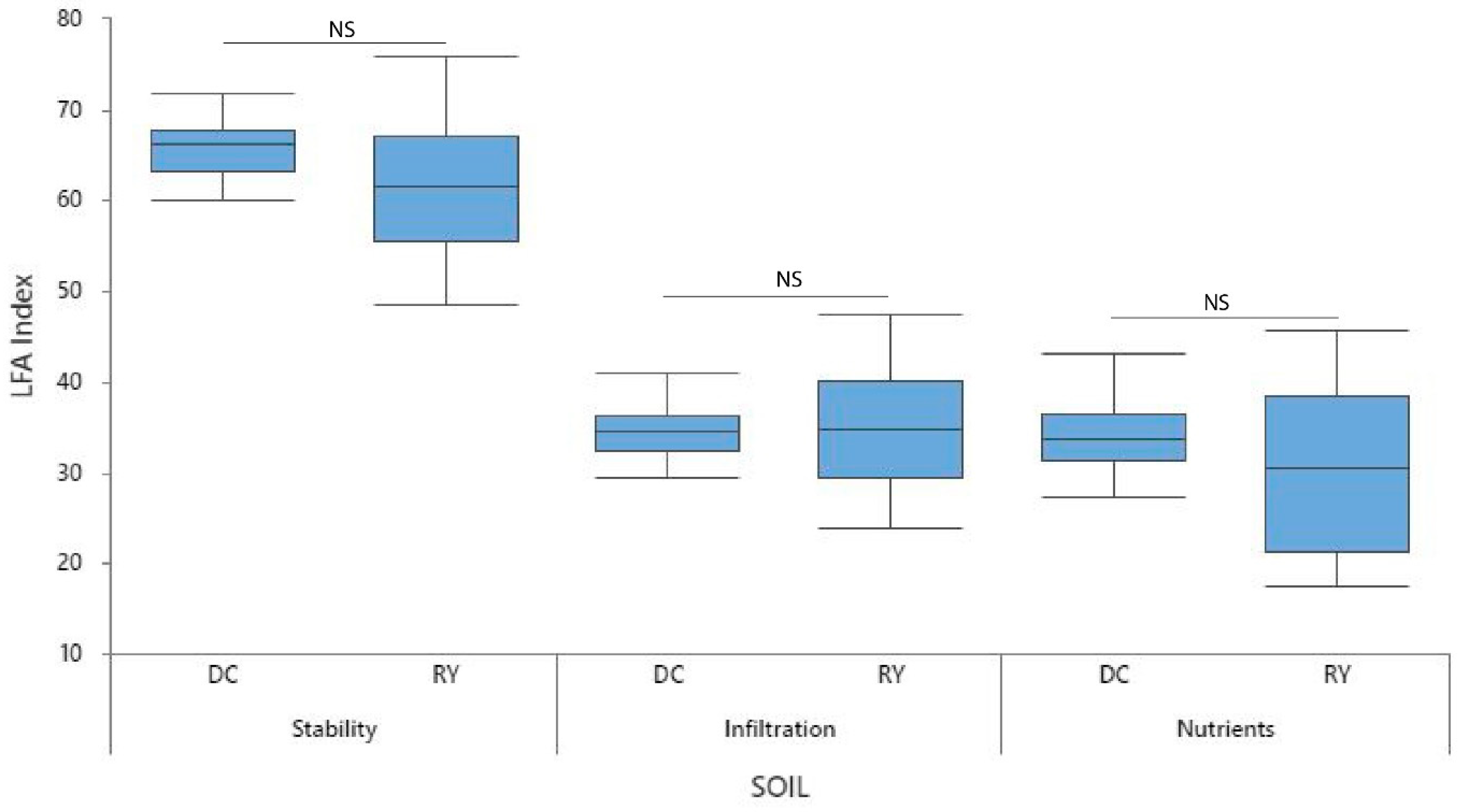
3.3. Landscape Function across Interspaces
3.3.1. Stability
3.3.2. Infiltration
3.3.3. Nutrient Cycling
4. Discussion
4.1. Sustaining Landscape Function during Drought
4.2. Biocrusts Protect the Soil during Drought
4.3. Biocrust Hotspots—The Engine Room of the Landscape
4.4. Managing and Monitoring the Interspaces
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Reagain, P.J.; Scanlan, J.C. Sustainable Management for Rangelands in a Variable Climate: Evidence and Insights from Northern Australia. Animal 2013, 7, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, J.C.; McIvor, J.G.; Bray, S.G.; Cowley, R.A.; Hunt, L.P.; Pahl, L.I.; MacLeod, N.D.; Whish, G.L. Resting Pastures to Improve Land Condition in Northern Australia: Guidelines Based on the Literature and Simulation Modelling. Rangel. J. 2014, 36, 429–443. [Google Scholar] [CrossRef]
- Tongway, D.J. Soil and Landscape Processes in the Restoration of Rangelands. Rangel. J. 1990, 12, 54–57. [Google Scholar] [CrossRef]
- Ash, A.J.; Corfield, J.P.; McIvor, J.G.; Ksiksi, T.S. Grazing Management in Tropical Savannas: Utilization and Rest Strategies to Manipulate Rangeland Condition. Rangel. Ecol. Manag. 2011, 64, 223–239. [Google Scholar] [CrossRef]
- Ash, A.; Thornton, P.; Stokes, C.r.s.; Togtohyn, C. Is Proactive Adaptation to Climate Change Necessary in Grazed Rangelands? Rangel. Ecol. Manag. 2012, 65, 563–568. [Google Scholar] [CrossRef][Green Version]
- Godde, C.M.; Garnett, T.; Thornton, P.K.; Ash, A.J.; Herrero, M. Grazing Systems Expansion and Intensification: Drivers, Dynamics, and Trade-Offs. Glob. Food Secur. 2018, 16, 93–105. [Google Scholar] [CrossRef]
- Holt, J.A.; Coventry, R.J. Nutrient Cycling in Australian Savannas. J. Biogeogr. 1990, 17, 427–432. [Google Scholar] [CrossRef]
- Housman, D.C.; Yeager, C.M.; Darby, B.J.; Sanford, R.L.; Kuske, C.R.; Neher, D.A.; Belnap, J. Heterogeneity of Soil Nutrients and Subsurface Biota in a Dryland Ecosystem. Soil Biol. Biochem. 2007, 39, 2138–2149. [Google Scholar] [CrossRef]
- Mott, J.J.; Williams, J.; Andrew, M.H.; Gillison, A.N. Australian Savanna Ecosystems. In Ecology and Management of the World’s Savannas; Australian Academy of Sciences: Canberra, Australia, 1985; pp. 56–82. [Google Scholar]
- McKeon, G.M.; Day, K.A.; Howden, S.M.; Mott, J.J.; Orr, D.M.; Scattini, W.; Weston, E.J. Northern Australian Savannas: Management for Pastoral Production. J. Biogeogr. 1990, 355–372. [Google Scholar] [CrossRef]
- Hill, J.O.; Coates, D.B.; Whitbread, A.M.; Clem, R.L.; Robertson, M.J.; Pengelly, B.C.; Hill, J.O.; Coates, D.B.; Whitbread, A.M.; Clem, R.L.; et al. Seasonal Changes in Pasture Quality and Diet Selection and Their Relationship with Liveweight Gain of Steers Grazing Tropical Grass and Grass–Legume Pastures in Northern Australia. Anim. Prod. Sci. 2009, 49, 983–993. [Google Scholar] [CrossRef]
- Tongway, D.; Hindley, N. Landscape Function Analysis: A System for Monitoring Rangeland Function. Afr. J. Range Forage Sci. 2004, 21, 109–113. [Google Scholar] [CrossRef]
- Pointing, S.B.; Belnap, J. Microbial Colonization and Controls in Dryland Systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Poore, A.G.B.; Ruiz-Colmenero, M.; Letnic, M.; Soliveres, S. Ecosystem Structure, Function, and Composition in Rangelands Are Negatively Affected by Livestock Grazing. Ecol. Appl. 2016, 26, 1273–1283. [Google Scholar] [CrossRef]
- Williams, W.J.; Büdel, B.; Reichenberger, H.; Rose, N. Cyanobacteria in the Australian Northern Savannah Detect the Difference between Intermittent Dry Season and Wet Season Rain. Biodivers. Conserv. 2014, 23, 1827–1844. [Google Scholar] [CrossRef]
- Williams, W.; Büdel, B.; Williams, S. Wet Season Cyanobacterial N Enrichment Highly Correlated with Species Richness and Nostoc in the Northern Australian Savannah. Biogeosciences 2018, 15, 2149–2159. [Google Scholar] [CrossRef]
- Barger, N.N.; Weber, B.; Garcia-Pichel, F.; Zaady, E.; Belnap, J. Patterns and Controls on Nitrogen Cycling of Biological Soil Crusts. In Biological Soil Crusts: An Organizing Principle in Drylands; Ecological Studies; Springer: Cham, Switzerland, 2016; pp. 257–285. ISBN 978-3-319-30212-6. [Google Scholar]
- Hunt, L.P.; McIvor, J.G.; Grice, A.C.; Bray, S.G. Principles and Guidelines for Managing Cattle Grazing in the Grazing Lands of Northern Australia: Stocking Rates, Pasture Resting, Prescribed Fire, Paddock Size and Water Points—A Review. Rangel. J. 2014, 36, 105–119. [Google Scholar] [CrossRef]
- Williams, W.J.; Eldridge, D.J.; Alchin, B.M. Grazing and Drought Reduce Cyanobacterial Soil Crusts in an Australian Acacia Woodland. J. Arid Environ. 2008, 72, 1064–1075. [Google Scholar] [CrossRef]
- O’Reagain, P.; Bushell, J.; Holloway, C.; Reid, A.; O’Reagain, P.; Bushell, J.; Holloway, C.; Reid, A. Managing for Rainfall Variability: Effect of Grazing Strategy on Cattle Production in a Dry Tropical Savanna. Anim. Prod. Sci. 2009, 49, 85–99. [Google Scholar] [CrossRef]
- Aspandiar, M.F.; Taylor, G.; Eggleton, R.A. Charters Towers Region, Queensland. In Regolith Landscape Evolution Across Australia: A Compilation of Regolith Landscape Case Studies with Regolith Landscape Evolution Models; Cooperative Research Centre for Landscape Environments and Mineral Exploration: Perth, Australia, 2003. [Google Scholar]
- Tongway, D.J.; Ludwig, J.A. Restoring Disturbed Landscapes: Putting Principles into Practice; Island Press: Washington, DC, USA, 2011; ISBN 1-59726-580-2. [Google Scholar]
- Tongway, D.; Hindley, N. Landscape Function Analysis: Procedures for Monitoring and Assessing Landscapes—with Special Reference to Minesites and Rangelands; CSIRO Sustainable Ecosystems: Canberra, Australia, 2004. [Google Scholar]
- Data Analysis Software | Statistical Software Package. Available online: https://www.minitab.com/content/www/websites/en-us/products/minitab.html (accessed on 3 November 2021).
- Williams, W.J.; Büdel, B. Species Diversity, Biomass and Long-Term Patterns of Biological Soil Crusts with Special Focus on Cyanobacteria of the Acacia Aneura Mulga Lands of Queensland, Australia. Algol. Stud. 2012, 140, 23–50. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Zaady, E.; Shachak, M. Microphytic Crusts, Shrub Patches and Water Harvesting in the Negev Desert: The Shikim System. Landsc. Ecol. 2002, 17, 587–597. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Leys, J.F. Exploring Some Relationships between Biological Soil Crusts, Soil Aggregation and Wind Erosion. J. Arid Environ. 2003, 53, 457–466. [Google Scholar] [CrossRef]
- Bowker, M.A.; Belnap, J.; Bala Chaudhary, V.; Johnson, N.C. Revisiting Classic Water Erosion Models in Drylands: The Strong Impact of Biological Soil Crusts. Soil Biol. Biochem. 2008, 40, 2309–2316. [Google Scholar] [CrossRef]
- Bowker, M.A.; Maestre, F.T.; Eldridge, D.; Belnap, J.; Castillo-Monroy, A.; Escolar, C.; Soliveres, S. Biological Soil Crusts (Biocrusts) as a Model System in Community, Landscape and Ecosystem Ecology. Biodivers. Conserv. 2014, 23, 1619–1637. [Google Scholar] [CrossRef]
- Büdel, B.; Williams, W.J.; Reichenberger, H. Annual Net Primary Productivity of a Cyanobacteria-Dominated Biological Soil Crust in the Gulf Savannah, Queensland, Australia. Biogeosciences 2018, 15, 491. [Google Scholar] [CrossRef]
- Slate, M.L.; Callaway, R.M.; Pearson, D.E. Life in Interstitial Space: Biocrusts Inhibit Exotic but Not Native Plant Establishment in Semi-Arid Grasslands. J. Ecol. 2019, 107, 1317–1327. [Google Scholar] [CrossRef]
- Chamizo, S.; Mugnai, G.; Rossi, F.; Certini, G.; De Philippis, R. Cyanobacteria Inoculation Improves Soil Stability and Fertility on Different Textured Soils: Gaining Insights for Applicability in Soil Restoration. Front. Environ. Sci. 2018, 6, 49. [Google Scholar] [CrossRef]
- Williams, W.; Chilton, A.M.; Schneemilch, M.; Williams, S.; Neilan, B.A.; Driscoll, C. Microbial Biobanking–Cyanobacteria-Richtopsoil Facilitates Mine Rehabilitation. Biogeosciences 2019, 16, 1–16. [Google Scholar] [CrossRef]
- Bowker, M.A. Biological Soil Crust Rehabilitation in Theory and Practice: An Underexploited Opportunity. Restor. Ecol. 2007, 15, 13–23. [Google Scholar] [CrossRef]
- Román, J.R.; Roncero-Ramos, B.; Chamizo, S.; Rodríguez-Caballero, E.; Cantón, Y. Restoring Soil Functions by Means of Cyanobacteria Inoculation: Importance of Soil Conditions and Species Selection. Land Degrad. Dev. 2018, 29, 3184–3193. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Soil | HSR | MSR | R/SPELL | XCL |
|---|---|---|---|---|---|
| Stability | DC | 65.0 ± 3.39 | 65.1 ± 1.64 | 65.1 ± 1.21 | 66.9 ± 1.24 |
| RY | 53.8 ± 3.88 | 59.8 ± 1.94 | 62.1 ± 3.66 | 68.5 ± 2.24 | |
| Infiltration | DC | 31.6 ± 0.76 | 34.8 ± 0.99 | 36.8 ± 0.82 | 37.1 ± 1.17 |
| RY | 28.2 ± 1.93 | 32.3 ± 4.25 | 38.3 ± 3.89 | 40.6 ± 1.49 | |
| Nutrients | DC | 31.8 ± 1.8 | 33.9 ± 3.38 | 35.6 ± 0.97 | 35.5 ± 1.86 |
| RY | 19.4 ± 0.94 | 27.5 ± 4.11 | 33.8 ± 4.67 | 36.7 ± 1.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, W.J.; Schmidt, S.; Zaady, E.; Alchin, B.; Myint Swe, T.; Williams, S.; Dooley, M.; Penfold, G.; O’Reagain, P.; Bushell, J.; et al. Resting Subtropical Grasslands from Grazing in the Wet Season Boosts Biocrust Hotspots to Improve Soil Health. Agronomy 2022, 12, 62. https://doi.org/10.3390/agronomy12010062
Williams WJ, Schmidt S, Zaady E, Alchin B, Myint Swe T, Williams S, Dooley M, Penfold G, O’Reagain P, Bushell J, et al. Resting Subtropical Grasslands from Grazing in the Wet Season Boosts Biocrust Hotspots to Improve Soil Health. Agronomy. 2022; 12(1):62. https://doi.org/10.3390/agronomy12010062
Chicago/Turabian StyleWilliams, Wendy J., Susanne Schmidt, Eli Zaady, Bruce Alchin, Than Myint Swe, Stephen Williams, Madeline Dooley, Grace Penfold, Peter O’Reagain, John Bushell, and et al. 2022. "Resting Subtropical Grasslands from Grazing in the Wet Season Boosts Biocrust Hotspots to Improve Soil Health" Agronomy 12, no. 1: 62. https://doi.org/10.3390/agronomy12010062
APA StyleWilliams, W. J., Schmidt, S., Zaady, E., Alchin, B., Myint Swe, T., Williams, S., Dooley, M., Penfold, G., O’Reagain, P., Bushell, J., Cowley, R., Driscoll, C., & Robinson, N. (2022). Resting Subtropical Grasslands from Grazing in the Wet Season Boosts Biocrust Hotspots to Improve Soil Health. Agronomy, 12(1), 62. https://doi.org/10.3390/agronomy12010062