
Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions

 ,
,  ,
, 
 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Rhizobacteria Selection
2.3. Treatments and Growth Analysis
2.4. Delayed Fluorescence Measurements
2.5. Statistical Analysis
3. Results
3.1. Crop Growth and Delayed Fluorescence Measurements
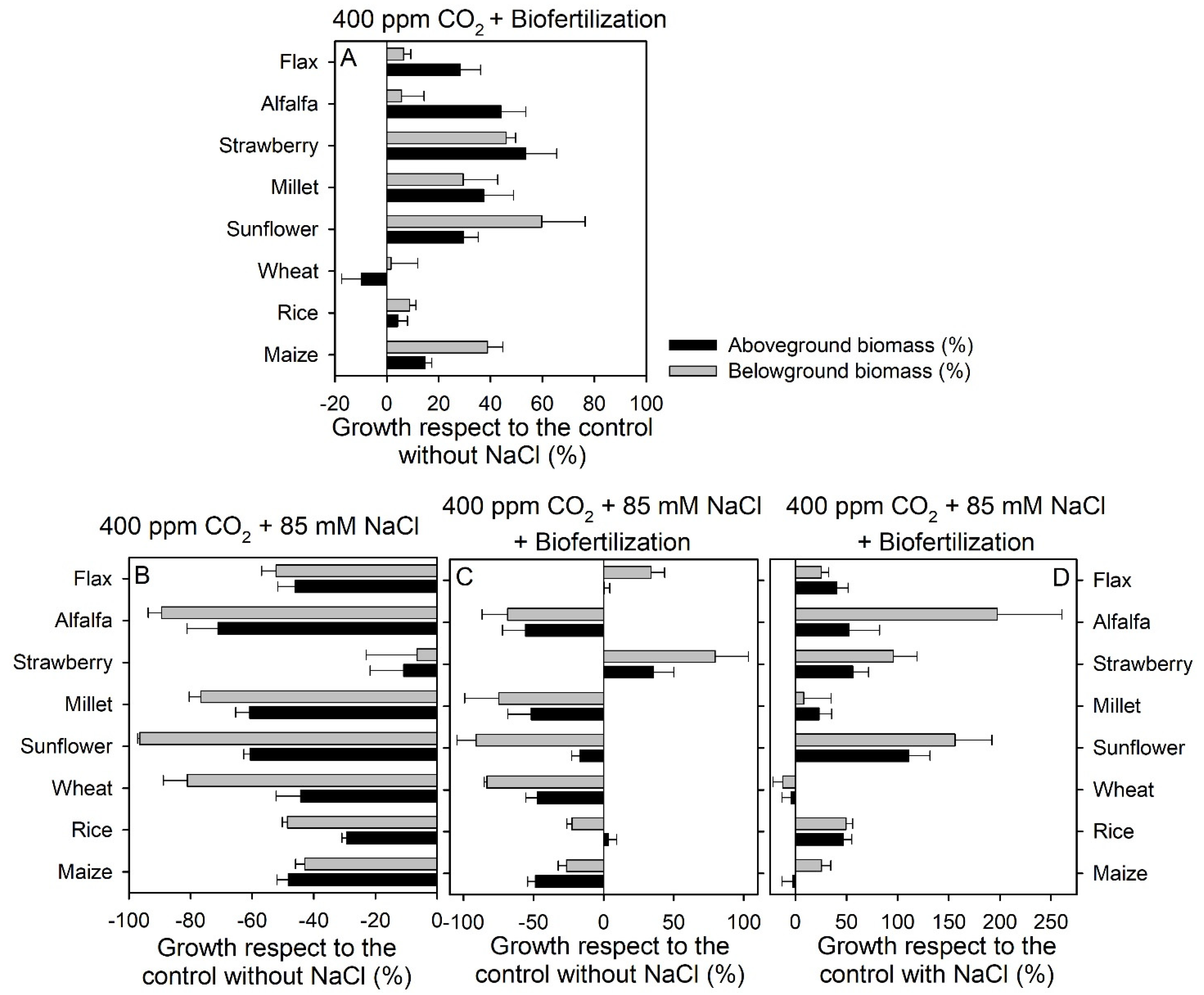
3.1.1. Biofertilizers from Halophytes in the Current Atmosphere
3.1.2. Biofertilizers from Halophytes under Salinity in the Present Climate
3.1.3. Biofertilizers from Halophytes in the Atmosphere of the Future
3.1.4. Biofertilizers from Halophytes under Salinity in the Future Climate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The Future of Food and Agriculture—Trends and Challenges; FAO: Rome, Italy, 2017. [Google Scholar]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.C.; Rosegrant, M.W.; Koo, J.; Robertson, R.; Sulser, T.; Zhu, T.; Ringler, C.; Msangi, S.; Palazzo, A.; Batka, M.; et al. Climate Change: Impact on Agriculture and Costs of Adaptation; IFPRI Food Policy Report: Washington, DC, USA, 2009. [Google Scholar]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Kafi, M.; Khan, M.A. Crop and Forage Production using Saline Waters; Daya Publishing House: New Delhi, India, 2008. [Google Scholar]
- Etesami, H.; Beattie, G.A. Mining halophytes for plant growth-promoting halotolerant. Bacteria to enhance the salinity tolerance of non-halophytic crops. Front. Microbiol. 2018, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.C.; Colletti, J.P.; Faltonson, R.R. Agroforestry opportunities for the United States of America. Agrofor. Syst. 1995, 31, 117–142. [Google Scholar] [CrossRef]
- Socolow, R.H. Nitrogen management and the future of food: Lessons from the management of energy and carbon. Proc. Natl. Acad. Sci. USA 1999, 96, 6001–6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, A.; Ahmad, E.; Khan, M.S.; Saif, S.; Rizvi, A. Role of plant growth promoting rhizobacteria in sustainable production of vegetables: Current perspective. Sci. Hortic. Amst. 2015, 193, 231–239. [Google Scholar] [CrossRef]
- Thomas, L.; Singh, I. Microbial Biofertilizers: Types and Applications. In Biofertilizers for Sustainable Agriculture and Environment; Giri, B., Prasad, R., Wu, Q.S., Varma, A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 1–19. [Google Scholar]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. Amst. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.C. Perspectives and challenges of microbial application for crop improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Mesa-Marín, J.; Mateos-Naranjo, E.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Redondo-Gómez, S. Synergic Effect Rhizobacteria—Halophytes as a Rising Strategy to Face a Changing World. In Halophytes and Climatic Change: Adaptive mechanisms and Potential Uses; Hasanuzzaman, M., Shabala, S., Fujita, M., Eds.; CABI: Wallingford, UK, 2019; pp. 240–254. [Google Scholar]
- Giongo, A.; Ambrosini, A.; Vargas, L.K.; Freire, J.R.J.; Bodanese-Zanettini, M.H.; Passaglia, L.M.P. Evaluation of genetic diversity of bradyrhizobia strains nodulating soybean [Glycine max (L.) Merrill] isolated from South Brazilian fields. Appl. Soil Ecol. 2008, 38, 261–269. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P.; Saikia, R. Genetic diversity of plant growth promoting rhizobacteria isolated from rhizospheric soil of wheat under saline condition. Curr. Microbiol. 2009, 59, 489–496. [Google Scholar] [CrossRef]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Bernabeu-Meana, M.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Redondo-Gómez, S. Effect of plant growth promoting rhizobacteria on Salicornia ramosissima seed germination under salinity, CO2 and temperature stress. Agronomy 2019, 9, 655. [Google Scholar] [CrossRef] [Green Version]
- Olivares, F.L.; Busato, J.G.; de Paula, A.M.; Da Silva Lima, L.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef]
- FAO. FAOstat. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 22 June 2020).
- Álvarez, C.; Navarro, J.A.; Molina-Heredia, F.P.; Mariscal, V. Endophytic colonization of rice (Oryza sativa L.) by the symbiotic strain Nostoc punctiforme PCC 73102. Mol. Plant Microbe Interact. 2020, 33, 1040–1045. [Google Scholar] [CrossRef]
- Ariza, M.T.; Soria, C.; Medina-Mínguez, J.J.; Martínez-Ferri, E. Incidence of Misshapen fruits in strawberry plants grown under tunnels is affected by cultivar, planting date, pollination, and low temperatures. Hort. Sci. 2012, 47, 1569–1573. [Google Scholar] [CrossRef] [Green Version]
- Andrades-Moreno, L.; Del Castillo, I.; Parra, R.; Doukkali, B.; Redondo-Gómez, S.; Pérez-Palacios, P.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Prospecting metal-resistant plant-growth promoting rhizobacteria for rhizoremediation of metal contaminated estuaries using Spartina densiflora. Environ. Sci. Pollut. Res. Int. 2014, 21, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Bortolus, A.; Adam, P.; Adams, J.; Ainouche, M.; Ayres, D.; Bertness, M.; Bouma, T.; Bruno, J.; Caçador, I.; Carlton, J.; et al. Supporting Spartina: Interdisciplinary perspective shows Spartina as a distinct solid genus. Ecology 2019, 100, e02863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of plant-growth-promoting and metal-resistant cultivable bacteria from Arthrocnemum macrostachyum in the Odiel marshes with potential use in phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef]
- Mesa, J.; Mateos-Naranjo, E.; Caviedes, M.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D. Scouting contaminated estuaries: Heavy metal resistant and plant growth promoting rhizobacteria in the native metal rhizoaccumulator Spartina maritima. Mar. Pollut. Bull. 2015, 90, 150–159. [Google Scholar] [CrossRef] [PubMed]
- López-Jurado, J.; Balao, F.; Mateos-Naranjo, E. Polyploidy-mediated divergent light-harvesting and photoprotection strategies under temperature stress in a Maditerranean carnation complex. Env. Exp. Bot. 2020, 171, 103956. [Google Scholar] [CrossRef]
- Dimkpa, C.; Weinamd, T.; Asch, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef]
- Van Overbeek, L.; van Elsas, J.D. Effects of plant genotype and growth stage on the structure of bacterial communities associated with potato (Solanumtuberosum L.). FEMS Microbiol. Ecol. 2008, 64, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Pillay, V.K.; Nowak, J. Inoculum density, temperature, and genotype effects on in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L) seedlings inoculated with a pseudomonad bacterium. Can. J. Microbiol. 1997, 43, 354–361. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophyticcolonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Env. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Liu, J.; Tang, L.; Gao, H.; Zhang, M.; Guo, C. Enhancement of alfalfa yield and quality by plant growth-promoting rhizobacteria under saline-alkali conditions. J. Sci. Food Agric. 2019, 99, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Rana, A.; Saharan, B.; Joshi, M.; Prasanna, R.; Kumar, K.; Nain, L. Identification of multi-trait PGPR isolates and evaluating their potential as inoculants for wheat. Ann. Microbiol. 2011, 61, 893–900. [Google Scholar] [CrossRef]
- Ambrosini, A.; Beneduzi, A.; Stefanski, T.; Pinheiro, F.G.; Vargas, L.K.; Passaglia, L.M.P. Screening of plant growth promoting Rhizobacteria isolated from sunflower (Helianthus annuus L.). Plant Soil 2012, 356, 245–264. [Google Scholar] [CrossRef]
- Braga, L.F.; De Oliveira, F.A.; Prado do Couto, E.A.; D’ECA Nogueira Santos, K.F.; De Brito Ferreira, E.P.; Garcia Martin-Didonet, C.C. Polyphasic characterization of bacteria obtained from upland rice cultivated in Cerrado soil. Braz. J. Microbiol. 2018, 49, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Vestberg, M.; Kukkonen, S.; Saari, K.; Parikka, P.; Huttunen, J.; Tainio, L.; Devos, N.; Weekers, F.; Kevers, C.; Thonart, P.; et al. Microbial inoculation for improving the growth and health ofmicropropagated strawberry. Appl. Soil Ecol. 2004, 27, 243–258. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B.; Patel, S. Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol. Plant. 2011, 33, 797–802. [Google Scholar] [CrossRef]
- Ansari, M.; Shekari, F.; Mohammadi, M.H.; Juhos, K.; Végvári, G.; Biró, B. Salt-tolerant plant growth-promoting bacteria enhanced salinity tolerance of salt-tolerant alfalfa (Medicago sativa L.) cultivars at high salinity. Acta Physiol. Plant. 2019, 41, 195. [Google Scholar] [CrossRef] [Green Version]
- Kiani, M.Z.; Sultan, T.; Ali, A.; Qadir, G.; Mahmood, I.A.; Tabassam, T.; Ullah, M.A.; Abbas, N. Effect of PGPR strains on sunflower growth and nutrient contents under salinity stress. Pak. J. Agric. Res. 2016, 29, 2. [Google Scholar]
- Karlidag, H.; Yildirim, E.; Turan, M.; Pehluvan, M.; Donmez, F. Plant Growth-promoting Rhizobacteria Mitigate Deleterious Effects of Salt Stress on Strawberry Plants (Fragaria × ananassa). Hortscience 2013, 48, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Singh, P.; Tiwari, R.; Meena, K.K.; Yandigeri, M.; Singh, D.P.; Arora, D.K. Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biol. Fertil. Soils 2011, 47, 907. [Google Scholar] [CrossRef]
- Cao, Y.R.; Chen, S.Y.; Zhang, J.S. Ethylene signaling regulates salt stress response. Plant Signal. Behav. 2008, 3, 761–763. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Glick, B.R. Bacterial Modulation of Plant Ethylene Levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Gitz, D.C.; Sicher, R.C.; Baker, J.T.; Timlin, D.J.; Reddy, V.R. Temperature dependence of growth, development, and photosynthesis in maize under elevated CO2. Environ. Exp. Bot. 2007, 61, 224–236. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lim, S.S.; Kwak, J.H.; Lee, D.S.; Ro, H.M.; Choi, W.J. Dry matter and nitrogen accumulation and partitioning in rice (Oryza sativa L.) exposed to experimental warming with elevated CO2. Plant Soil 2011, 342, 59–71. [Google Scholar] [CrossRef]
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agr. For. Meteorol. 2009, 149, 51–58. [Google Scholar] [CrossRef]
- De Costa, W.A.J.M.; Weerakoon, W.M.W.; Herath, H.M.L.K.; Amaratunga, K.S.P.; Abeywardena, R.M.I. Physiology of yield determination of rice under elevated carbon dioxide at high temperatures in a subhumid tropical climate. Field Crop Res. 2006, 96, 336–347. [Google Scholar] [CrossRef]
- Keutgen, N.; Chen, K.; Lenz, F. Responses of strawberry leaf photosynthesis, chlorophyll fluorescence and macronutrient contents to elevated CO2. J. Plant Physiol. 1997, 150, 395–400. [Google Scholar] [CrossRef]
- Balasooriya, H.N.; Dassanayake, K.B.; Seneweera, S.; Ajlouni, S. Interaction of Elevated Carbon Dioxide and Temperature on Strawberry (Fragaria × ananassa) Growth and Fruit Yield. Int. J. Arg. Biosyst. Eng. 2018, 12, 279–287. [Google Scholar]
- Marilley, L.; Hartwig, U.A.; Aragno, M. Influence of an Elevated Atmospheric CO2 Content on Soil and Rhizosphere Bacterial Communities Beneath Lolium perenne and Trifolium repens under Field Conditions. Microb. Ecol. 1999, 38, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.; Pétriacq, P.; Beerling, D.J.; Cotton, T.E.A.; Ton, J. Impacts of atmospheric CO2 and soil nutritional value on plant responses to rhizosphere colonization by soil bacteria. Front. Plant Sci. 2018, 9, 1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montealegre, C.M.; Van Kessel, C.; Russelle, M.P.; Sadowsky, M.J. Changes in microbial activity and composition in a pasture ecosystem exposed to elevated atmospheric carbon dioxide. Plant Soil 2002, 243, 197–207. [Google Scholar] [CrossRef]
- Van Veen, J.A.; Liljeroth, E.; Lekkerkerk, L.J.A.; Van de Geijn, S.C. Carbon fluxes in plant–soil systems at elevated atmospheric CO2 levels. Ecol. Appl. 1991, 1, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Neumann, G.; Kania, A.; Kuzyakov, Y.; Romheld, V.; Kandeler, E. Elevation of atmospheric CO2 and N-nutritional status modify nodulation, nodule-carbon supply, and root exudation of Phaseolus vulgaris L. Soil Biol. Biochem. 2007, 39, 2208–2221. [Google Scholar] [CrossRef]
- Smith, P.; Gregory, P.J. Climate change and sustainable food production. Proc. Nutr. Soc. 2013, 72, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Naranjo, E.; Redondo-Gómez, S. Interpopulation differences in salinity tolerance of the invasive cordgrass Spartina densiflora: Implications for invasion process. Estuaries Coast 2016, 39, 98–107. [Google Scholar] [CrossRef]
- Mesa-Marín, J.; Pérez-Romero, J.A.; Redondo-Gómez, S.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Impact of plant growth promoting bacteria on Salicornia ramosissima ecophysiology and heavy metal phytoremediation capacity in estuarine soils. Front. Microbiol. 2020, 11, 553018. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| PGPR Properties | ||||||||
|---|---|---|---|---|---|---|---|---|
| Consortium Number | Bacterial Strains | N Fixation | P Solubilization | Siderophores Production | IAA Production (mg/mL) | Biofilm Production | ACC Deaminase | Reference |
| 1 | Pseudomonas composti SDT3 | - | + | + | - | - | - | |
| Aeromonas aquariorum SDT13 | - | + | + | 3.40 | - | - | [22] | |
| Bacillus thuringiensis SDT14 | + | - | + | - | - | - | ||
| 2 | Vibrio kanaloae RA1 | + | - | + | 0.63 | - | - | |
| Pseudoalteromonas prydzensis RA15 | + | - | + | 1.58 | - | - | [24] | |
| Staphylococcus warneri RA18 | - | - | - | 2.84 | - | - | ||
| 3 | Bacillus methylotrophicus SMT38 | + | - | + | - | + | - | |
| Bacillus aryabhattai SMT48 | + | + | + | 3.25 | - | - | [25] | |
| Bacillus licheniformis SMT51 | + | + | + | 1.06 | + | - | ||
| 4 | Vibrio spartinae HPJ2 | + | + | + | 4.12 | + | + | |
| Marinobacter sediminum HPJ15 | - | - | + | 15.41 | - | - | [16] | |
| Vibrio parahaemolyticus HPJ50 | + | + | + | 7.48 | + | - | ||
| 5 | Vibrio neocaledonicus SRT1 | + | + | + | 5.65 | + | - | |
| Thalassospira australica SRT8 | - | - | - | - | + | + | [16] | |
| Pseudarthrobacter oxydans SRT15 | + | + | - | 20.99 | - | - | ||
| 400 ppm CO2 | +4 °C and 700 ppm CO2 | |||
|---|---|---|---|---|
| Crop | 0 mM NaCl | 85 mM NaCl | 0 mM NaCl | 85 mM NaCl |
| Maize | 3 (2, 5) *** | 4 ** | 1 *** | 4 ns |
| Rice | 1 (3) *** | 2 (1) *** | 1 (3) ** | 3 (1) *** |
| Wheat | 1 ns | 1 ns | 1 (5) ** | 3 (1, 4, 5) * |
| Sunflower | 4 (1, 2) ** | 1 *** | 2ns | 2*** |
| Millet | 2 (3, 5) * | 2 (5) ** | 5 (1, 2, 3) * | 2 ns |
| Strawberry | 1 (2, 3, 4) * | 3 *** | 3 (5) *** | 3 *** |
| Alfalfa | 3 *** | 5 *** | 2 ns | 2 (1) * |
| Flax | 3 (1) *** | 5 *** | 1 (3, 5) *** | 3 (5) * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redondo-Gómez, S.; Mesa-Marín, J.; Pérez-Romero, J.A.; López-Jurado, J.; García-López, J.V.; Mariscal, V.; Molina-Heredia, F.P.; Pajuelo, E.; Rodríguez-Llorente, I.D.; Flowers, T.J.; et al. Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy 2021, 11, 1609. https://doi.org/10.3390/agronomy11081609
Redondo-Gómez S, Mesa-Marín J, Pérez-Romero JA, López-Jurado J, García-López JV, Mariscal V, Molina-Heredia FP, Pajuelo E, Rodríguez-Llorente ID, Flowers TJ, et al. Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy. 2021; 11(8):1609. https://doi.org/10.3390/agronomy11081609
Chicago/Turabian StyleRedondo-Gómez, Susana, Jennifer Mesa-Marín, Jesús A. Pérez-Romero, Javier López-Jurado, Jesús V. García-López, Vicente Mariscal, Fernando P. Molina-Heredia, Eloisa Pajuelo, Ignacio D. Rodríguez-Llorente, Timothy J. Flowers, and et al. 2021. "Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions" Agronomy 11, no. 8: 1609. https://doi.org/10.3390/agronomy11081609
APA StyleRedondo-Gómez, S., Mesa-Marín, J., Pérez-Romero, J. A., López-Jurado, J., García-López, J. V., Mariscal, V., Molina-Heredia, F. P., Pajuelo, E., Rodríguez-Llorente, I. D., Flowers, T. J., & Mateos-Naranjo, E. (2021). Consortia of Plant-Growth-Promoting Rhizobacteria Isolated from Halophytes Improve Response of Eight Crops to Soil Salinization and Climate Change Conditions. Agronomy, 11(8), 1609. https://doi.org/10.3390/agronomy11081609