Phytochemical Composition and Nutritional Value of Pot-Grown Turnip-Rooted and Plain and Curly-Leafed Parsley Cultivars

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
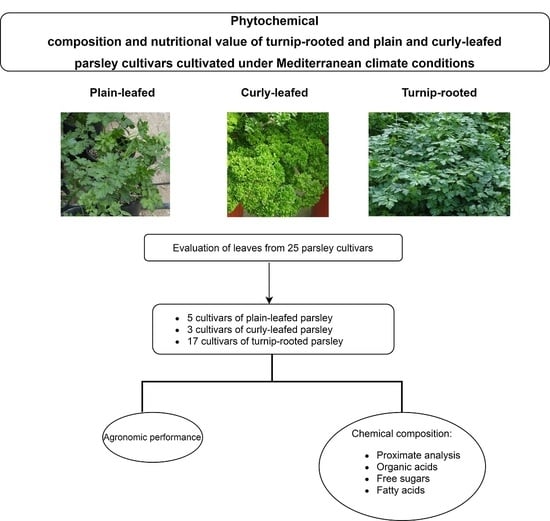
1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Plant Material and Growing Conditions
2.3. Nutritional and Energetic Value Determination
2.4. Chemical Composition Analysis
2.4.1. Organic Acids
2.4.2. Sugars Composition
2.4.3. Tocopherols
2.4.4. Fatty Acids
2.5. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agyare, C.; Appiah, T.; Boakye, Y.D.; Antwi, J. Petroselinum crispum: A review. In Medicinal Spices and Vegetables from Africa; Victor, K., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 18–31. [Google Scholar]
- Alan, O.; Avci, A.B.; Akcali Giachino, R.R. Harvest number and growing season effects on quality and health related compounds in parsley. Indian J. Pharm. Educ. Res. 2017, 51, S276–S280. [Google Scholar] [CrossRef]
- Akıncı, A.; Eşrefoğlu, M.; Taşlıdere, E.; Ateş, B. Petroselinum crispum is effective in reducing stress-induced gastric oxidative damage. Balk. Med. J. 2017, 34, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Abbasabadi, Z.; Ardekani, M.R.S.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef]
- Dadan, M.; Rybak, K.; Wiktor, A.; Nowacka, M.; Zubernik, J.; Witrowa-Rajchert, D. Selected chemical composition changes in microwave-convective dried parsley leaves affected by ultrasound and steaming pre-treatments–An optimization approach. Food Chem. 2018, 239, 242–251. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.M.; Metwally, N.H.; Ibrahim, I.A.; Abdel-Hady, H.; Abdel-Wahab, B.S.A. Antioxidant Activity, total phenolic and flavonoid contents of Petroselinum crispum Mill. J. Appl. Life Sci. Int. 2018, 19, 1–7. [Google Scholar] [CrossRef]
- Chauhan, E.S.; Aishwaya, J. Nutraceuticals potential of Petroselinum crispum: A Review. J. Complement. Med. Altern. Med. 2018, 7, 1–6. [Google Scholar]
- Dobričević, N.; Šic Žlabur, J.; Voća, S.; Pliestić, S.; Galić, A.; Delić, A.; Fabek Uher, S. Bioactive compounds content and nutritional potential of different parsley parts (Petroselinum crispum Mill.). J. Cent. Eur. Agric. 2019, 20, 900–910. [Google Scholar]
- Petropoulos, S.A.; Daferera, D.; Polissiou, M.G.; Passam, H.C. Effect of freezing, drying and the duration of storage on the composition of essential oils of plain-leafed parsley [Petroselinum crispum (Mill.) Nym. ssp. neapolitanum Danert] and turnip-rooted parsley [Petroselinum crispum (Mill.) Nym. ssp. tuberosum (Bernh.) Crov]. Flavour Fragr. J. 2010, 25, 28–34. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Daferera, D.; Polissiou, M.G.; Passam, H.C. The effect of water deficit stress on the growth, yield and composition of essential oils of parsley. Sci. Hortic. (Amsterdam) 2008, 115, 393–397. [Google Scholar] [CrossRef]
- Calderón-Montaño, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini-Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Dražić, S.; Živanović, T.; Prodanović, S. Stability of productive traits of genotypes of cultivated medicinal plants of the family apiaceae. Biotechnol. Biotechnol. Equip. 2007, 21, 100–106. [Google Scholar] [CrossRef][Green Version]
- Bekhradi, F.; Delshad, M.; Marín, A.; Luna, M.; Garrido, Y.; Kashi, A.; Babalar, M.; Gil, M. Effects of salt stress on physiological and postharvest quality characteristics of different Iranian genotypes of basil. Hortic. Environ. Biotechnol. 2015, 56, 777–785. [Google Scholar] [CrossRef]
- Elia, A.; Santamaria, P. Biodiversity in vegetable crops, a heritage to save: The case of Puglia region. Ital. J. Agron. 2013, 8, 21–34. [Google Scholar] [CrossRef]
- Riggi, E.; Siracusa, L.; Lanza, C.M.; Mazzaglia, A.; Di Silvestro, I.; La Rosa, S.; Pellegrino, A.; Ippolito, C.; Testa, G.; Scordia, D.; et al. Valorisation of typical products through characterising and promoting actions: Morpho-biometric traits, sensory analysis and flavonol content in Cipolla di Giarratana. Ital. J. Agron. 2013, 8, 79–84. [Google Scholar] [CrossRef]
- Petropoulos, S.A.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef]
- Maxim, A.; Andor, M.Ş.; Jidavu, M.; Sima, R.; Lucian, C.; Opincariu, A.; Maxim, O.; Lucaci, A.; Bolboaca, V. Research concerning the conservation of genetic diversity of parsley (Petroselinum crispum MILL.). Bull. UASVM Agric. 2009, 66, 144–148. [Google Scholar]
- El-Zaeddi, H.; Calín-Sánchez, Á.; Martínez-Tomé, J.; Noguera-Artiaga, L.; Burló, F.; Carbonell-Barrachina, Á.A. Irrigation dose and plant density affect the essential oil content and sensory quality of parsley (Petroselinum sativum). Sci. Hortic. 2016, 206, 1–6. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Daferera, D.; Polissiou, M.G.; Passam, H.C. Effect of nitrogen-application rate on the biomass, concentration, and composition of essential oils in the leaves and roots of three types of parsley. J. Plant Nutr. Soil Sci. 2009, 172, 210–215. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Daferera, D.; Polissiou, M.G.; Passam, H.C. The effect of salinity on the growth, yield and essential oils of turnip-rooted and leaf parsley cultivated within the Mediterranean region. J. Sci. Food Agric. 2009, 89, 1534–1542. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Constantopoulou, E.; Karapanos, I.; Akoumianakis, C.A.; Passam, H.C. Diurnal variation in the nitrate content of parsley foliage. Int. J. Plant Prod. 2011, 5, 431–438. [Google Scholar]
- Petropoulos, S.A.; Daferera, D.; Al, E. The effect of sowing date and growth stage on the essential oil composition of three types of parsley (Petroselinum crispum). J. Sci. Food Agric. 2004, 1610, 1606–1610. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Akoumianakis, C.A.; Passam, H.C. Effect of sowing date and cultivar on yield and quality of turnip-rooted parsley (Petroselinum crispum ssp. tuberosum). J. Food Agric. Environ. 2005, 3, 205–207. [Google Scholar]
- Petropoulos, S.A.; Akoumianakis, C.A.; Passam, H.C. Evaluation of turnip-rooted parsley (Petroselinum crispum ssp. tuberosum) for root and foliage production under a warm, Mediterranean climate. Sci. Hortic. 2006, 109, 282–287. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â; Dias, M.I.; Pereira, C.; Calhelha, R.; Gioia, F.D.; Tzortzakis, N.; Ivanov, M.; Sokovic, M.; Barros, L.; et al. Wild and cultivated Centaurea raphanina subsp. mixta: A valuable source of bioactive compounds. Antioxidants 2020, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- AOAC Official Methods of analysis of association of official analytical chemists international. In Official Methods of Analysis of AOAC International; Horwitz, W., Latimer, G., Eds.; AOAC Int.: Gaithersburg, MD, USA, 2016; ISBN 0935584773. [Google Scholar]
- Silva, A.R.; Fernandes, Â.; García, P.A.; Barros, L.; Ferreira, I.C.F.R. Cytinus hypocistis (L.) L. subsp. macranthus wettst.: Nutritional characterization. Molecules 2019, 24, 1111. [Google Scholar] [CrossRef]
- Pereira, C.; Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Use of UFLC-PDA for the analysis of organic acids in thirty-five species of food and medicinal plants. Food Anal. Methods 2013, 6, 1337–1344. [Google Scholar] [CrossRef]
- Golubkina, N.A.; Kharchenko, V.A.; Moldovan, A.I.; Koshevarov, A.A.; Zamana, S.; Nadezhkin, S.; Soldatenko, A.; Sekara, A.; Tallarita, A.; Caruso, G. Yield, growth, quality, biochemical characteristics and elemental composition of plant parts of celery leafy, stalk and root types grown in the northern hemisphere. Plants 2020, 9, 484. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Olympios, C.M.; Passam, H.C. The effect of nitrogen fertilization on plant growth and the nitrate content of leaves and roots of parsley in the Mediterranean region. Sci. Hortic. 2008, 118, 255–259. [Google Scholar] [CrossRef]
- Saleh, H.A.R.; El-Nashar, Y.I.; Serag-El-Din, M.F.; Dewir, Y.H. Plant growth, yield and bioactive compounds of two culinary herbs as affected by substrate type. Sci. Hortic. 2019, 243, 464–471. [Google Scholar] [CrossRef]
- Khalil, E.; Esoh, R.; Rababah, T.; Almajwal, A.M.; Alu, M.H. Minerals, proximate composition and their correlations of medicinal plants from Jordan. J. Med. Plants Res. 2012, 6, 5757–5762. [Google Scholar]
- El Gindy, A.A.; Youssef, M.E.; Youssif, M.R.G. Effect of adding drying parsley leaves on processing and quality of “crackers.”. Middle East J. Appl. Sci. 2017, 7, 421–429. [Google Scholar]
- Saleh, A.M.; Selim, S.; Al Jaouni, S.; AbdElgawad, H. CO2 enrichment can enhance the nutritional and health benefits of parsley (Petroselinum crispum L.) and dill (Anethum graveolens L.). Food Chem. 2018, 269, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Gîrd, C.E.; Nencu, I.; Duţu, L.E.; Popescu, M.L.; Costea, T.; Neagu, A.F. HPLC evaluation of the ascorbic acid content of romanian fruits and vegetables from retail markets. Farmacia 2018, 66, 894–898. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Serio, F.; Todaro, E. A survey of nitrate and oxalate content in fresh vegetables. J. Sci. Food Agric. 1999, 79, 1882–1888. [Google Scholar] [CrossRef]
- Boldizsár, I.; Füzfai, Z.; Molnár-Perl, I. Characterization of the endogenous enzymatic hydrolyses of Petroselinum crispum glycosides: Determined by chromatography upon their sugar and flavonoid products. J. Chromatogr. A 2013, 1293, 100–106. [Google Scholar] [CrossRef]
- Gómez-Coronado, D.J.M.; Ibañez, E.; Rupérez, F.J.; Barbas, C. Tocopherol measurement in edible products of vegetable origin. J. Chromatogr. A 2004, 1054, 227–233. [Google Scholar] [CrossRef]
- Zeweil, H.S.; El-Gindy, Y.M. Effects of parsley supplementation on the seminal quality, blood lipid profile and oxidant status of young and old male rabbits. World Rabbit Sci. 2017, 25, 215–223. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Viršilė, A.; Jankauskienė, J.; Sakalauskienė, S.; Duchovskis, P. Red Light-dose or wavelength-dependent photoresponse of antioxidants in herb microgreens. PLoS ONE 2016, e0163405. [Google Scholar] [CrossRef]
- Mert, A.; Timur, M. Essential oil and fatty acid composition and antioxidant capacity and total phenolic content of parsley seeds (Petroselinum crispum) grown in Hatay Region. Indian J. Pharm. Educ. Res. 2017, 51, S437–S440. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Cultivar Type | Cultivar Name | Seed Company | Leaf Fresh Weight (g/pot) |
|---|---|---|---|
| Plain-leafed | 97 ± 8 A,* | ||
| Astra | Polan | 50 ± 9 e | |
| Festival 68 | W. Legutko | 81 ± 8 d | |
| Fest | Polan | 87 ± 8 c | |
| Gigante Di Italia | W. Legutko | 122 ± 7 b | |
| Rialto Bejo | Bejo Zaden | 192 ± 11 a | |
| Curly leafed | 87 ± 8 B | ||
| Depuis 1743 | Vilmorin Garden | 91 ± 7 b | |
| Mooskrause | Semenarna Ljubljana | 107 ± 9 a | |
| Moss Curled 2 | W. Legutko | 66 ± 9 c | |
| Turnip-rooted | 86 ± 8 B | ||
| Alba | Vilmorin Garden | 71 ± 9 g | |
| Arat | Bejo Zaden | 153 ± 12 a | |
| Berlinski Halblange Springer | Springer semena | 91 ± 8 d | |
| Cukrowa | W. Legutko | 66 ± 6 ij | |
| Halblange Berlinska | W. Legutko | 81 ± 9 f | |
| Halblange Eagle | W. Legutko | 56 ± 6 k | |
| Hanacka | Vilmorin Garden | 56 ± 3 k | |
| Kaśka | PNOS | 83 ± 5 e,f | |
| Konika | Toraf | 69 ± 9 g,h | |
| Lenka | W. Legutko | 68 ± 8 h,i | |
| Linga | Polan | 102 ± 10 b | |
| Olomuńcka | W. Legutko | 88 ± 6 d | |
| Osborne | PNOS | 92 ± 9 c | |
| Pólna | Toraf | 63 ± 8 j | |
| Root parsley (Common variety) | - | 156 ± 10 a | |
| Sonata | PNOS | 95 ± 4 c | |
| Vistula | Polan | 85 ± 10 e |
| Cultivar Type | Cultivar Name | Fat | Proteins | Ash | Carbohydrates | Energy |
|---|---|---|---|---|---|---|
| Plain-leafed | 2.7 ± 0.4 A,B,* | 11.4 ± 0.8 B | 11.6 ± 0.7 A,B | 74 ± 2 A | 367 ± 3 A | |
| Astra | 2.99 ± 0.03 a | 12.28 ± 0.08 a | 11.73 ± 0.09 c | 73.00 ± 0.01 d | 368.0 ± 0.4 b | |
| Fest | 2.01 ± 0.06 c | 10.74 ± 0.06 d | 10.52 ± 0.08 e | 76.7 ± 0.1 a | 368.0 ± 0.4 b | |
| Festival 68 | 2.91 ± 0.06 a | 10.16 ± 0.09 e | 11.23 ± 0.07 d | 75.7 ± 0.2 b | 369.6 ± 0.1 a | |
| Gigante Di Italia | 2.48 ± 0.04 b | 11.52 ± 0.06 c | 12.58 ± 0.08 a | 73.4 ± 0.1 c | 362.1 ± 0.1 c | |
| Rialto Bejo | 3.00 ± 0.09 a | 12.15 ± 0.02 b | 11.95 ± 0.01 b | 72.90 ± 0.07 d | 367.2 ± 0.3 b | |
| Curly leafed | 2.9 ± 0.1 A | 12.4 ± 0.8 A | 11.3 ± 0.7 B | 73 ± 2 A | 369 ± 3 A | |
| Depuis 1743 | 2.89 ± 0.03 b | 13.43 ± 0.08 a | 12.19 ± 0.01 a | 71.5 ± 0.1 c | 365.7 ± 0.2 b | |
| Mooskrause | 2.97 ± 0.06 a | 11.53 ± 0.02 c | 10.84 ± 0.09 b | 74.7 ± 0.1 a | 371.5 ± 0.5 a | |
| Moss Curled 2 | 2.70 ± 0.01 c | 12.22 ± 0.04 b | 10.85 ± 0.07 b | 74.2 ± 0.1 b | 370.1 ± 0.2 a,b | |
| Turnip-rooted | 2.5 ± 0.2 B | 12 ± 1 A | 12.1 ± 0.7 A | 73 ± 2 A | 364 ± 3 B | |
| Alba | 2.59 ± 0.02 c,d | 11.56 ± 0.08 k | 11.98 ± 0.05 f | 73.9 ± 0.1 d,e | 365.1 ± 0.1 f | |
| Arat | 2.59 ± 0.01 d | 10.62 ± 0.05 m | 10.96 ± 0.03 k | 75.8 ± 0.1 a | 369.1 ± 0.1 a | |
| Berlinski Halblange Springer | 2.86 ± 0.03 a | 11.80 ± 0.04 i | 11.45 ± 0.09 i | 73.9 ± 0.1 d,e | 368.5 ± 0.2 b | |
| Cukrowa | 2.69 ± 0.06 b | 11.10 ± 0.07 l | 11.63 ± 0.01 g,h | 74.6 ± 0.1 b | 366.9 ± 0.2 d | |
| Halblange Berlinska | 2.30 ± 0.03 f | 12.92 ± 0.04 e | 12.19 ± 0.02 d | 72.6 ± 0.1 g | 362.8 ± 0.2 h | |
| Halblange Eagle | 2.56 ± 0.01 d | 11.70 ± 0.04 i,j | 11.41 ± 0.08 i,j | 74.3 ± 0.1 c | 367.2 ± 0.2 d | |
| Hanacka | 2.60 ± 0.09 c,d | 12.07 ± 0.03 h | 11.59 ± 0.02 g,h | 73.7 ± 0.1 e,f | 366.6 ± 0.3 d | |
| Kaśka | 2.47 ± 0.02 e | 13.79 ± 0.06 a | 11.58 ± 0.04 h | 72.2 ± 0.1 h | 366.1 ± 0.2 e | |
| Konika | 2.44 ± 0.01 e | 13.55 ± 0.06 c,d | 12.89 ± 0.05 b | 71.12 ± 0.08 i | 360.6 ± 0.1 i | |
| Lenka | 2.30 ± 0.01 f | 10.71 ± 0.03 m | 12.97 ± 0.01 b | 74.0 ± 0.1 d | 359.6 ± 0.1 j | |
| Linga | 2.85 ± 0.02 a | 13.63 ± 0.01 b,c,d | 12.33 ± 0.01 c | 71.20 ± 0.02 i | 364.9 ± 0.1 f | |
| Olomuńcka | 2.20 ± 0.02 g | 13.53 ± ± 0.08 d | 12.12 ± 0.05 d,e | 72.2 ± 0.1 h | 362.5 ± 0.1 h | |
| Osborne | 2.67 ± 0.03 b,c | 13.74 ± 0.06 a,b | 13.38 ± 0.02 a | 70.2 ± 0.1 k | 359.8 ± 0.1 j | |
| Pólna | 2.15 ± 0.05 g | 11.61 ± 0.06 j,k | 12.03 ± 0.02 e,f | 74.2 ± 0.1 c | 362.6 ± 0.1 h | |
| Root parsley (Common variety) | 2.62 ± 0.06 b,c,d | 12.33 ± 0.05 g | 11.32 ± 0.08 j | 73.7 ± 0.1 e,f | 367.8 ± 0.4 c | |
| Sonata | 2.56 ± 0.02 d | 13.67 ± 0.09 a,b,c | 13.36 ± 0.03 a | 70.4 ± 0.1 j | 359.4 ± 0.1 j | |
| Vistula | 2.15 ± 0.01 g | 12.50 ± 0.05 f | 11.68 ± 0.04 g | 73.7 ± 0.1 f | 364.0 ± 0.1 g |
| Cultivar Type | Cultivar Name | Oxalic acid | Malic Acid | Ascorbic Acid | Shikimic Acid | Citric Acid | Total Organic Acids |
|---|---|---|---|---|---|---|---|
| Plain-leafed | 1.79 ± 0.07 B * | 5.4 ± 0.1 C | 0.024 ± 0.005 A | 0.044 ± 0.009 A | 1.35 ± 0.05 B | 8.6 ± 0.2 C | |
| Astra | 1.71 ± 0.01 e | 5.25 ± 0.01 e | 0.023 ± 0.001 c | 0.050 ± 0.001 b | 1.28 ± 0.01 e | 8.32 ± 0.01 e | |
| Fest | 1.73 ± 0.01 d | 5.28 ± 0.01 d | 0.020 ± 0.001 d | 0.030 ± 0.001 c | 1.42 ± 0.01 a | 8.50 ± 0.01 d | |
| Festival 68 | 1.82 ± 0.01 c | 5.48 ± 0.01 b | 0.020 ± 0.001 d | 0.040 ± 0.001 c | 1.33 ± 0.01 d | 8.69 ± 0.01 b | |
| Gigante Di Italia | 1.83 ± 0.01 b | 5.50 ± 0.01 a | 0.030 ± 0.001 b | 0.050 ± 0.001 a | 1.35 ± 0.01 c | 8.76 ± 0.01 a | |
| Rialto Bejo | 1.87 ± 0.01 a | 5.37 ± 0.01 c | 0.030 ± 0.001 a | 0.050 ± 0.001 b | 1.36 ± 0.01 b | 8.68 ± 0.01 c | |
| Curly-leafed | 2.80 ± 0.05 A | 6.7 ± 0.1 A | 0.017 ± 0.006 B | 0.030 ± 0.001 B | 1.65 ± 0.05 A | 11.2 ± 0.2 A | |
| Depuis 1743 | 2.81 ± 0.01 b | 6.88 ± 0.01 a | 0.010 ± 0.001 c | 0.030 ± 0.001 c | 1.70 ± 0.06 a | 11.45 ± 0.06 a | |
| Mooskrause | 2.85 ± 0.01 a | 6.69 ± 0.01 b | 0.020 ± 0.001 b | 0.030 ± 0.001 a | 1.63 ± 0.01 b | 11.23 ± 0.01 b | |
| Moss Curled 2 | 2.75 ± 0.01 c | 6.59 ± 0.01 c | 0.020 ± 0.001 a | 0.030 ± 0.001 b | 1.61 ± 0.01 b | 11.02 ± 0.01 c | |
| Turnip-rooted | 2.8 ± 0.4 A | 5.9 ± 0.4 B | 0.021 ± 0.006 B | 0.039 ± 0.007 A | 1.4 ± 0.2 B | 10.1 ± 0.8 B | |
| Alba | 3.13 ± 0.01 d | 5.59 ± 0.01 i | 0.010 ± 0.001 a | 0.030 ± 0.001 h | 1.25 ± 0.01 k | 10.01 ± 0.01 h | |
| Arat | 3.03 ± 0.01 e | 6.15 ± 0.01 e | 0.030 ± 0.001 b | 0.040 ± 0.001 g | 1.35 ± 0.01 i | 10.60 ± 0.01 d | |
| Berlinski Halblange Springer | 3.13 ± 0.01 d | 6.63 ± 0.01 a | 0.030 ± 0.001 a | 0.060 ± 0.001 a | 1.31 ± 0.01 j | 11.15 ± 0.01 a | |
| Cukrowa | 2.85 ± 0.01 h | 6.61 ± 0.01 b | 0.020 ± 0.001 e | 0.040 ± 0.001 g | 1.23 ± 0.01 l | 10.75 ± 0.01 c | |
| Halblange Berlinska | 3.01 ± 0.01 f | 6.50 ± 0.01 c | 0.020 ± 0.001 | 0.040 ± 0.001 f | 1.39 ± 0.01 h | 10.95 ± 0.03 b | |
| Halblange Eagle | 3.33 ± 0.01 a | 5.84 ± 0.01 f | 0.020 ± 0.001 d | 0.040 ± 0.001 e | 1.74 ± 0.01 a | 10.97 ± 0.01 b | |
| Hanacka | 2.96 ± 0.01 g | 5.58 ± 0.01 i | 0.020 ± 0.001 d | 0.030 ± 0.001 h | 1.57 ± 0.01 d | 10.17 ± 0.01 f | |
| Lenka | 3.18 ± 0.01 b | 5.66 ± 0.01 h | 0.010 ± 0.001 b | 0.030 ± 0.001 h | 1.71 ± 0.01 b | 10.60 ± 0.01 d | |
| Linga | 3.16 ± 0.01 c | 5.75 ± 0.01 g | 0.020 ± 0.001 h | 0.040 ± 0.001 e | 1.64 ± 0.01 c | 10.61 ± 0.01 d | |
| Kaśka | 2.67 ± 0.01 i | 6.19 ± 0.02 d | 0.020 ± 0.001 f | 0.040 ± 0.001 c | 1.55 ± 0.01 e | 10.47 ± 0.01 e | |
| Konika | 2.48 ± 0.01 l | 6.17 ± 0.01 d | 0.020 ± 0.001 e | 0.040 ± 0.001 e | 1.41 ± 0.01 f | 10.12 ± 0.01 g | |
| Olomuńcka | 2.20 ± 0.01 m | 5.46 ± 0.01 k | 0.020 ± 0.001 h | 0.040 ± 0.001 e | 1.40 ± 0.01 g | 9.13 ± 0.02 k | |
| Osborne | 2.10 ± 0.01o | 5.52 ± 0.01 j | 0.020 ± 0.001 g | 0.040 ± 0.001 e | 1.21 ± 0.01 m | 8.89 ± 0.01 m | |
| Pólna | 2.52 ± 0.01 k | 5.82 ± 0.01 f | 0.020 ± 0.001 h | 0.050 ± 0.001 b | 1.13 ± 0.01 o | 9.54 ± 0.01 i | |
| Root parsley (Common variety) | 2.58 ± 0.01 j | 5.51 ± 0.01 j | 0.020 ± 0.001 f | 0.030 ± 0.001 h | 1.16 ± 0.01 n | 9.31 ± 0.01 j | |
| Sonata | 2.67 ± 0.01 i | 5.22 ± 0.01 l | 0.030 ± 0.001 c | 0.040 ± 0.001 d | 1.06 ± 0.01 p | 9.02 ± 0.02 l | |
| Vistula | 2.18 ± 0.01 n | 5.75 ± 0.01 g | 0.020 ± 0.001 e | 0.040 ± 0.001 f | 1.03 ± 0.01 q | 9.02 ± 0.01 l |
| Cultivar Type | Cultivar Name | Apiose | Fructose | Glucose | Sucrose | Total Sugars | α-Tocopherol | γ-Tocopherol | Total Tocopherols |
|---|---|---|---|---|---|---|---|---|---|
| Plain-leafed | 0.8 ± 0.2 B * | 1.4 ± 0.5 A | 2.2 ± 0.5 B | 3.4 ± 0.7 A | 7.7 ± 0.7 A | 26.1 ± 0.8 A | 5 ± 1 A | 31 ± 2 A | |
| Astra | 0.98 ± 0.01 b | 1.24 ± 0.04 b | 1.46 ± 0.02 e | 3.61 ± 0.01 b | 7.29 ± 0.08 d | 26.04 ± 0.01 d | 3.95 ± 0.01 e | 29.99 ± 0.01 c | |
| Fest | 0.59 ± 0.09 c | 0.98 ± 0.01 c | 2.77 ± 0.06 a | 4.42 ± 0.01 a | 8.76 ± 0.04 a | 24.88 ± 0.02 e | 4.23 ± 0.04 d | 29.11 ± 0.07 d | |
| Festival 68 | 0.58 ± 0.01 c | 1.01 ± 0.03 c | 2.02 ± 0.04 d | 3.01 ± 0.03 d | 6.6 ± 0.1 e | 27.18 ± 0.05 a | 5.56 ± 0.04 b | 32.74 ± 0.08 a | |
| Gigante Di Italia | 0.76 ± 0.02 b | 1.26 ± 0.03 b | 2.52 ± 0.05 b | 3.52 ± 0.03 c | 8.06 ± 0.09 b | 26.15 ± 0.06 c | 6.67 ± 0.01 a | 32.83 ± 0.05 a | |
| Rialto Bejo | 0.88 ± 0.01 a | 2.33 ± 0.02 a | 2.11 ± 0.01 c | 2.49 ± 0.01 e | 7.81 ± 0.01 c | 26.31 ± 0.06 b | 4.51 ± 0.01 c | 30.83 ± 0.07 b | |
| Curly-leafed | 0.87 ± 0.03 A,B | 1.22 ± 0.08 A | 2.53 ± 0.08 A,B | 3.2 ± 0.5 A | 7.9 ± 0.4 A | 15.1 ± 0.5 B | 4.71 ± 0.05 B | 19.8 ± 0.6 B | |
| Depuis 1743 | 0.89 ± 0.02 a | 1.32 ± 0.02 a | 2.45 ± 0.05 b | 2.68 ± 0.03 c | 7.3 ± 0.1 c | 15.83 ± 0.06 a | 4.76 ± 0.04 a | 20.6 ± 0.1 a | |
| Mooskrause | 0.83 ± 0.01 b | 1.15 ± 0.01 c | 2.63 ± 0.02 a | 3.77 ± 0.01 a | 8.37 ± 0.02 a | 14.72 ± 0.02 b | 4.71 ± 0.01 a,b | 19.43 ± 0.01 b | |
| Moss Curled 2 | 0.89 ± 0.04 a | 1.20 ± 0.01 b | 2.51 ± 0.05 b | 3.30 ± 0.04 b | 7.90 ± 0.04 b | 14.76 ± 0.05 b | 4.67 ± 0.06 b | 19.42 ± 0.01 b | |
| Turnip-rooted | 1.0 ± 0.3 A | 0.7 ± 0.1 B | 2.6 ± 0.5 A | 3.2 ± 0.5 A | 7.6 ± 0.9 A | 26 ± 2 A | 4 ± 1 B | 30 ± 3 A | |
| Alba | 0.92 ± 0.04 i | 0.79 ± 0.04 d,e,f | 3.43 ± 0.03 a | 3.27 ± 0.05 e | 8.40 ± 0.07 d | 27.05 ± 0.04 e,f | 2.64 ± 0.01 h | 29.69 ± 0.03 h | |
| Arat | 0.95 ± 0.01 g,h,i | 0.56 ± 0.01 h,i | 3.16 ± 0.01 c | 3.11 ± 0.01 f | 7.78 ± 0.01 f,g | 27.01 ± 0.04 f | 2.37 ± 0.01 i | 29.38 ± 0.04 i | |
| Berlinski Halblange Springer | 1.00 ± 0.02 f | 0.56 ± 0.02 h,i | 2.55 ± 0.01 g,h | 3.49 ± 0.07 d | 7.60 ± 0.02 h | 25.65 ± 0.07 h | 4.68 ± 0.02 d | 30.33 ± 0.09 g | |
| Cukrowa | 0.97 ± 0.01 f,g,h | 0.88 ± 0.05 b,c | 3.01 ± 0.02 d | 3.03 ± 0.01 h | 7.89 ± 0.09 e,f | 27.12 ± 0.07 e | 2.30 ± 0.02 j | 29.42 ± 0.05 i | |
| Halblange Berlinska | 0.80 ± 0.01 j | 0.55 ± 0.01 i | 2.72 ± 0.01 f | 3.07 ± 0.01 g,h | 7.14 ± 0.01 i | 24.20 ± 0.08 k | 3.14 ± 0.02 g | 27.34 ± 0.06 l | |
| Halblange Eagle | 0.93 ± 0.01 h,i | 0.79 ± 0.04 d,e | 3.02 ± 0.05 d | 3.98 ± 0.01 b | 8.72 ± 0.09 c | 30.32 ± 0.01 a | 4.70 ± 0.02 c,d | 35.02 ± 0.02 a | |
| Hanacka | 0.92 ± 0.01 i | 0.77 ± 0.01 d,e,f | 2.57 ± 0.01 g | 2.79 ± 0.07 j | 7.06 ± 0.08 i | 25.52 ± 0.04 i | 3.50 ± 0.03 f | 29.02 ± 0.01 j | |
| Kaśka | 1.72 ± 0.02 a | 0.73 ± 0.03 f | 2.50 ± 0.01 h,i | 3.92 ± 0.03 b | 8.87 ± 0.09 b | 27.26 ± 0.01 d | 4.76 ± 0.03 c | 32.03 ± 0.02 e | |
| Konika | 0.99 ± 0.02 f,g | 0.76 ± 0.01 e,f | 1.99 ± 0.01 j | 2.18 ± 0.02 l | 5.92 ± 0.01 k | 24.90 ± 0.07 j | 4.73 ± 0.03 c,d | 29.63 ± 0.04 h | |
| Lenka | 1.48 ± 0.01 b | 0.96 ± 0.01 a | 2.00 ± 0.06 j | 3.52 ± 0.04 d | 7.95 ± 0.02 e | 26.44 ± 0.01 g | 5.30 ± 0.06 a | 31.74 ± 0.06 f | |
| Linga | 1.01 ± 0.05 f | 0.68 ± 0.01 g | 2.82 ± 0.01 e | 3.16 ± 0.04 f | 7.66 ± 0.01 g,h | 28.92 ± 0.01 b | 5.28 ± 0.04 a | 34.21 ± 0.04 b | |
| Olomuńcka | 1.16 ± 0.03 d | 0.83 ± 0.01 c,d | 2.50 ± 0.01 h | 3.50 ± 0.02 d | 7.99 ± 0.07 e | 23.91 ± 0.02 l | 3.15 ± 0.04 g | 27.07 ± 0.02 m | |
| Osborne | 1.10 ± 0.01 e | 0.66 ± 0.02 g | 2.43 ± 0.02 i | 2.93 ± 0.03 i | 7.11 ± 0.04 i | 23.93 ± 0.03 l | 4.34 ± 0.06 e | 28.27 ± 0.03 k | |
| Pólna | 0.80 ± 0.01 j | 0.60 ± 0.03 h | 2.73 ± 0.01 f | 2.52 ± 0.04 k | 6.66 ± 0.08 j | 27.58 ± 0.07 c | 5.08 ± 0.02 b | 32.66 ± 0.08 c | |
| Root parsley (Common variety) | 0.48 ± 0.02 k | 0.47 ± 0.01 j | 1.72 ± 0.09 k | 2.82 ± 0.05 j | 5.50 ± 0.05 l | 22.46 ± 0.06 m | 2.14 ± 0.01 k | 24.61 ± 0.06 n | |
| Vistula | 1.25 ± 0.03 c | 0.90 ± 0.02 a,b | 2.51 ± 0.02 h | 3.78 ± 0.03 c | 8.45 ± 0.09 d | 30.24 ± 0.01 a | 4.75 ± 0.05 c,d | 34.99 ± 0.04 a |
| Cultivar Type | Cultivar Name | C16:0 | C18:2n6c | C18:3n3 | SFA | MUFA | PUFA | PUFA/SFA | n6/n3 |
|---|---|---|---|---|---|---|---|---|---|
| Plain-leafed | 21 ± 1 A * | 31 ± 2 A,B | 32 ± 3 A | 32 ± 1 B | 5.8 ± 0.6 A | 63 ± 2 B | 2.0 ± 0.1 B | 1.0 ± 0.1 A | |
| Astra | 20.12 ± 0.02 d | 30.08 ± 0.06 c | 33.2 ± 0.2 b | 31.2 ± 0.4 b | 5.3 ± 0.1 c | 63.5 ± 0.3 b | 2.03 ± 0.03 b | 0.91 ± 0.01 c | |
| Fest | 21.9 ± 0.2 a,b | 32.2 ± 0.1 a | 28.9 ± 0.1 d | 32.9 ± 0.1 a | 5.9 ± 0.1 b | 61.2 ± 0.2 d | 1.86 ± 0.01 c | 1.11 ± 0.01 a | |
| Festival 68 | 22.7 ± 0.5 a | 32.07 ± 0.01 a | 28.6 ± 0.2 e | 32.5 ± 0.2 a | 6.77 ± 0.05 a | 60.7 ± 0.1 d | 1.87 ± 0.02 c | 1.12 ± 0.01 a | |
| Gigante Di Italia | 21.5 ± 0.8 b,c | 30.6 ± 0.5 b | 31.96 ± 0.07 c | 31.5 ± 0.7 b | 5.8 ± 0.2 b | 62.7 ± 0.4 c | 1.99 ± 0.06 b | 0.96 ± 0.02 b | |
| Rialto Bejo | 20.90 ± 0.04 c,d | 28.13 ± 0.01 d | 36.18 ± 0.06 a | 30.07 ± 0.07 c | 5.33 ± 0.01 c | 64.57 ± 0.03 a | 2.15 ± 0.01 a | 0.78 ± 0.01 d | |
| Curly-leafed | 21.6 ± 0.5 A | 29 ± 1 B | 31.6 ± 0.3 A | 34.6 ± 0.6 A | 4.4 ± 1 B | 61 ± 1 C | 1.76 ± 0.05 C | 0.92 ± 0.05 A | |
| Depuis 1743 | 21.8 ± 0.2 a | 29.16 ± 0.07 b | 31.69 ± 0.01 a | 33.86 ± 0.01 c | 5.14 ± 0.04 a | 61.01 ± 0.05 b | 1.80 ± 0.01 a | 0.92 ± 0.01 b | |
| Mooskrause | 21.11 ± 0.01 a | 30.6 ± 0.6 a | 31.3 ± 0.3 b | 34.8 ± 0.1 b | 2.88 ± 0.01 b | 62.3 ± 0.2 a | 1.79 ± 0.01 a | 0.98 ± 0.03 a | |
| Moss Curled 2 | 21.9 ± 0.8 a | 27.6 ± 0.1 c | 31.8 ± 0.1 a | 35.2 ± 0.1 a | 5.17 ± 0.08 a | 59.6 ± 0.2 c | 1.69 ± 0.01 b | 0.87 ± 0.01 c | |
| Turnip-rooted | 21 ± 1 A | 33 ± 3 A | 31 ± 3 A | 30 ± 2 C | 6 ± 1 A | 64 ± 2 A | 2.1 ± 0.2 A | 1.1 ± 0.2 A | |
| Alba | 19.20 ± 0.02 e | 31.4 ± 0.5 g | 30.6 ± 0.4 f,g | 30.62 ± 0.04 e,f,g | 7.19 ± 0.01 c | 62.20 ± 0.02 j | 2.03 ± 0.01 f,g,h | 1.03 ± 0.03 g | |
| Arat | 20.03 ± 0.01 d | 33.02 ± 0.01 e | 30.67 ± 0.02 f,g | 30.47 ± 0.04 f,g | 5.62 ± 0.01 f | 63.91 ± 0.03 e,f | 2.10 ± 0.01 d,e,f | 1.08 ± 0.01 e,f | |
| Berlinski Halblange Springer | 22.58 ± 0.03 a | 34.9 ± 0.5 c | 28.5 ± 0.4 i | 28.43 ± 0.01 h,i | 7.94 ± 0.08 b | 63.63 ± 0.07 f,g,h | 2.24 ± 0.01 c | 1.22 ± 0.03 c | |
| Cukrowa | 20.24 ± 0.01 d | 30.50 ± 0.04 h | 32.3 ± 0.1 e | 31.57 ± 0.08 b,c,d | 5.35 ± 0.03 g | 63.1 ± 0.1 g,h | 2.00 ± 0.01 h | 0.94 ± 0.01 h | |
| Halblange Berlinska | 20.0 ± 0.7 d | 30.1 ± 0.3 h,i | 32.7 ± 0.4 d,e | 30.96 ± 0.7 d,e,f | 6.04 ± 0.04 e | 63.00 ± 0.7 h,i | 2.04 ± 0.07 f,g,h | 0.92 ± 0.01 h,i | |
| Halblange Eagle | 19.12 ± 0.01 e | 29.93 ± 0.08 i,j | 33.6 ± 0.1 c | 30.75 ± 0.03 e,f,g | 5.55 ± 0.01 f | 63.71 ± 0.04 f,g | 2.07 ± 0.01 e,f | 0.89 ± 0.01 i,j | |
| Hanacka | 21.64 ± 0.07 b,c | 39.32 ± 0.07 b | 26.42 ± 0.08 j | 27.8 ± 0.1 i | 6.2 ± 0.3 d,e | 66.0 ± 0.2 b | 2.37 ± 0.01 b | 1.49 ± 0.01 a | |
| Kaśka | 20.2 ± 0.3 d | 31.4 ± 0.2 g | 33.03 ± 0.08 c,d | 30.26 ± 0.07 g | 4.96 ± 0.07 h | 64.8 ± 0.1 c,d | 2.14 ± 0.01 d,e | 0.95 ± 0.01 h | |
| Konika | 21.22 ± 0.01 c | 40.1 ± 0.2 a | 26.5 ± 0.4 j | 27.1 ± 0.1 j | 6.25 ± 0.05 d | 66.7 ± 0.2 a | 2.46 ± 0.02 a | 1.51 ± 0.03 a | |
| Lenka | 21.50 ± 0.01 b,c | 32.27 ± 0.07 f | 29.7 ± 0.3 h | 33.23 ± 0.2 a | 4.38 ± 0.08 j | 62.39 ± 0.3 i,j | 1.88 ± 0.02 i | 1.09 ± 0.01 e | |
| Linga | 21.38 ± 0.03 b,c | 34.42 ± 0.06 d | 30.30 ± 0.03 g,h | 28.66 ± 0.07 h | 6.38 ± 0.03 d | 64.96 ± 0.1 c,d | 2.27 ± 0.01 c | 1.14 ± 0.01 d | |
| Olomuńcka | 21.98 ± 0.16 a,b | 32.1 ± 0.1 f | 31.07 ± 0.04 f | 31.7 ± 0.1 b,c | 4.98 ± 0.05 h | 63.35 ± 0.06 f,g,h | 2.00 ± 0.01 g,h | 1.03 ± 0.01 g | |
| Osborne | 20.1 ± 0.6 d | 30.4 ± 0.3 h,i | 36.2 ± 0.4 a | 28.5 ± 0.8 h,i | 4.66 ± 0.04 i | 66.9 ± 0.7 a | 2.35 ± 0.09 b | 0.84 ± 0.01 l | |
| Pólna | 21.26 ± 0.04 c | 32.3 ± 0.2 f | 30.8 ± 0.3 f,g | 32.0 ± 0.2 b | 4.74 ± 0.08 i | 63.2 ± 0.1 g,h | 1.97 ± 0.01 h | 1.05 ± 0.02 f,g | |
| Root parsley (Common variety) | 22.58 ± 0.18 a | 34.3 ± 0.2 d | 26.41 ± 0.04 j | 30.6 ± 0.2 e,f,g | 8.49 ± 0.02 a | 60.9 ± 0.2 k | 1.99 ± 0.02 h | 1.30 ± 0.01 b | |
| Sonata | 20.15 ± 0.28 d | 29.5 ± 0.2 j | 34.7 ± 0.4 b | 31.2 ± 0.1 c,d,e | 4.31 ± 0.06 j | 64.5 ± 0.2 d,e | 2.07 ± 0.02 f,g | 0.85 ± 0.02 k,l | |
| Vistula | 19.2 ± 0.8 e | 30.44 ± 0.06 h | 34.6 ± 0.7 b | 30.2 ± 0.6 g | 4.43 ± 0.01 j | 65.4 ± 0.6 b,c | 2.16 ± 0.06 d | 0.88 ± 0.02 j,k |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, Â.; Polyzos, N.; Petropoulos, S.A.; Pinela, J.; Ardohain, E.; Moreira, G.; Ferreira, I.C.F.R.; Barros, L. Phytochemical Composition and Nutritional Value of Pot-Grown Turnip-Rooted and Plain and Curly-Leafed Parsley Cultivars. Agronomy 2020, 10, 1416. https://doi.org/10.3390/agronomy10091416
Fernandes Â, Polyzos N, Petropoulos SA, Pinela J, Ardohain E, Moreira G, Ferreira ICFR, Barros L. Phytochemical Composition and Nutritional Value of Pot-Grown Turnip-Rooted and Plain and Curly-Leafed Parsley Cultivars. Agronomy. 2020; 10(9):1416. https://doi.org/10.3390/agronomy10091416
Chicago/Turabian StyleFernandes, Ângela, Nikolaos Polyzos, Spyridon A. Petropoulos, José Pinela, Elizandra Ardohain, Glaucia Moreira, Isabel C. F. R. Ferreira, and Lillian Barros. 2020. "Phytochemical Composition and Nutritional Value of Pot-Grown Turnip-Rooted and Plain and Curly-Leafed Parsley Cultivars" Agronomy 10, no. 9: 1416. https://doi.org/10.3390/agronomy10091416
APA StyleFernandes, Â., Polyzos, N., Petropoulos, S. A., Pinela, J., Ardohain, E., Moreira, G., Ferreira, I. C. F. R., & Barros, L. (2020). Phytochemical Composition and Nutritional Value of Pot-Grown Turnip-Rooted and Plain and Curly-Leafed Parsley Cultivars. Agronomy, 10(9), 1416. https://doi.org/10.3390/agronomy10091416