Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress


Abstract
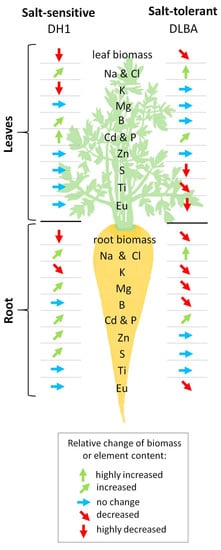
1. Introduction
2. Material and Methods
2.1. Plant Material and Cultivation
2.2. Analysis of Elements in Plants
2.3. Analysis of Elements in Soil
2.4. Gene Expression
2.5. Statistical Analysis
3. Results
3.1. Plant Growth
3.2. Uptake and Accumulation of Na and Cl in Plants
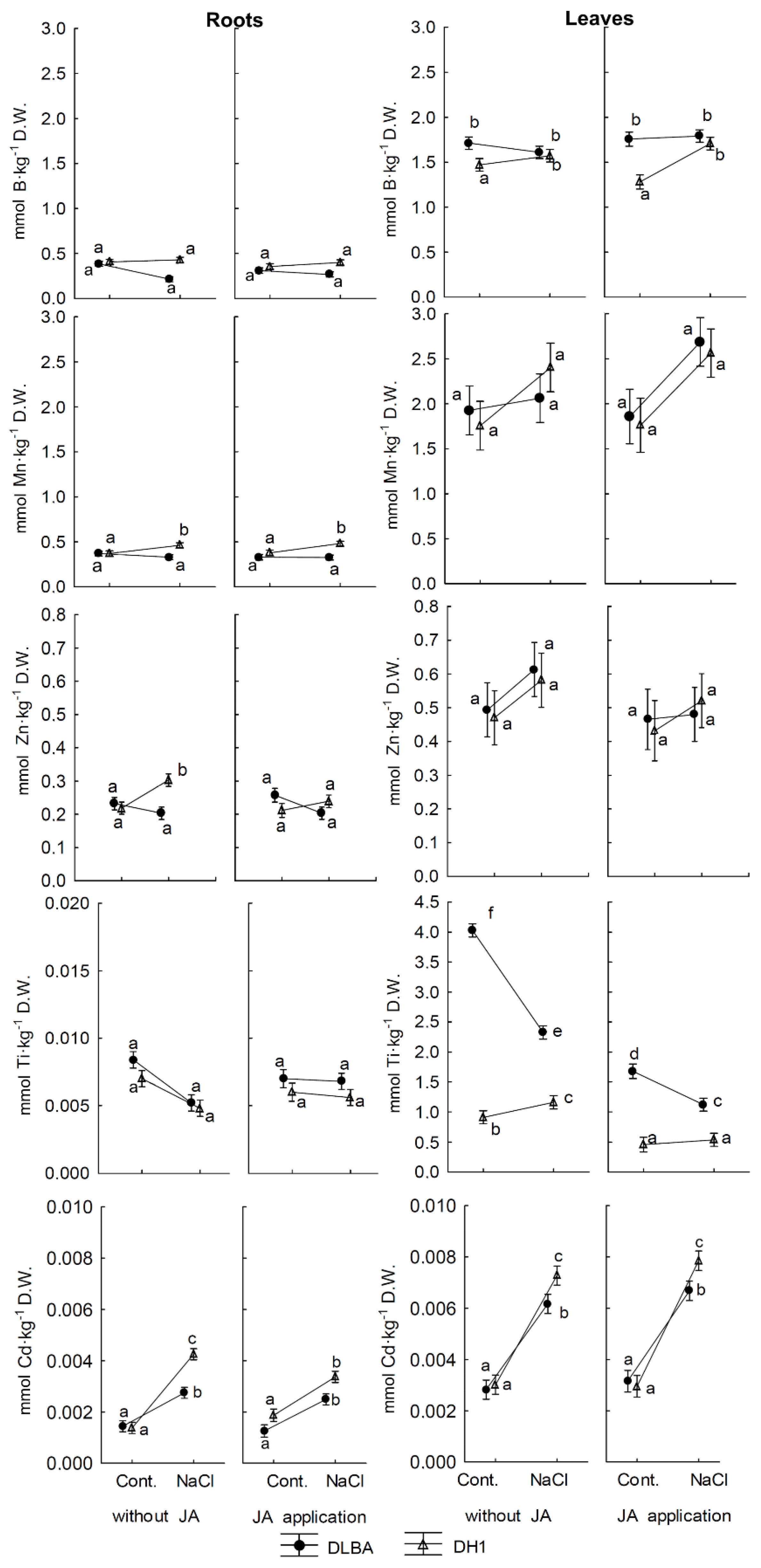
3.3. Influence of Salinity on the Uptake and Accumulation of Other Elements in Plants
3.4. Interaction of Salinity and Foliar Application of JA on the Accumulation of other Elements in Plants
3.5. Concentrations of Elements in the Soil after Carrot Cultivation
3.6. Expression of the DcNHX4 Cation-Proton Exchange Transporter Gene
4. Discussion
4.1. Response of Carrot Genotypes to Salinity Stress
4.2. Ability to Na+ and Cl− Uptake and Their Transport in the Plant in the ‘Storage Root-Leaves’ System
4.3. Possibility of Selective Uptake of K+ Ions under Conditions of High Na Concentration
4.4. Expression of the DcNHX4 Cation-Proton Exchange Transporter Gene
4.5. Relationship between Cd and Zn in the ‘Soil-Plant’ System
4.6. Uptake by Plants and Transport of P, Mg, S, B, Ti in the ‘Roots-Leaves’ System
4.7. Influence of JA on the Mineral Balance in Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Extent of Salt-Affected Soils. Available online: http://www.fao.org/soils-portal/soil-management/management-of-some-problem-soils/salt-affected-soils/more-information-on-salt-affected-soils/en/ (accessed on 23 January 2020).
- Tanveer, M.; Shabala, S. Targeting redox regulatory mechanisms for salinity stress tolerance in crops. In Salinity Responses and Tolerance in Plants, 1st ed.; Kumar, V., Wani, S.H., Suprasanna, P., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 213–234. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, D.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2014, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667–685. [Google Scholar]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Turkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Tajima, H.; Liang, Y.-C.; Ohto, M.-A.; Ushijima, K.; Nakano, R.; Esumi, T.; Coku, A.; Belmonte, M.; Blumwald, E. The arabidopsis Na+/H+ antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell 2011, 23, 3482–3497. [Google Scholar] [CrossRef]
- Sze, H.; Chanroj, S. Plant endomembrane dynamics: Studies of K+/H+ antiporters provide insights on the effects of pH and ion homeostasis. Plant Physiol. 2018, 177, 875–895. [Google Scholar] [CrossRef]
- Takahashi, H.; Kanayama, Y.; Zheng, M.S.; Kusano, T.; Hase, S.; Ikegami, M.; Shah, J. Antagonistic interactions between the SA and JA signaling pathways in Arabidopsis modulate expression of defense genes and gene-for-gene resistance to cucumber mosaic virus. Plant Cell Physiol. 2004, 45, 803–809. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates and octadecanoids: Signals in plant stress responses and development. Prog. Nucleic Acid Res. Mol. Biol. 2002, 72, 165–221. [Google Scholar] [CrossRef]
- Avanci, N.; Luche, D.; Goldman, G.H.; Goldman, M.H. Review Jasmonates are phytohormones with multiple functions, including plant defense and reproduction. Genet. Mol. Res. 2010, 9, 484–505. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.; Qi, M.; Sheng, G.; Yang, Y. Inducible overexpression of a rice allene oxide synthase gene increases the endogenous jasmonic acid level, prgene expression, and host resistance to fungal infection. Mol. Plant-Microbe Interact. 2006, 19, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Del Amor, F.M.; Cuadra-Crespo, P. Alleviation of salinity stress in broccoli using foliar urea or methyl-jasmonate: Analysis of growth, gas exchange, and isotope composition. Plant Growth Regul. 2010, 63, 55–62. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Hamayun, M.; Lee, S.-K.; Lee, I.-J. Methyl jasmonate alleviated salinity stress in soybean. J. Crop. Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Grzebelus, D.; Iorizzo, M.; Senalik, D.; Ellison, S.; Cavagnaro, P.; Macko-Podgorni, A.; Heller-Uszynska, K.; Kilian, A.; Nothnagel, T.; Allender, C.; et al. Diversity, genetic mapping, and signatures of domestication in the carrot (Daucus carota L.) genome, as revealed by Diversity Arrays Technology (DArT) markers. Mol. Breed. 2013, 33, 625–637. [Google Scholar] [CrossRef]
- Iorizzo, M.; Senalik, D.; Ellison, S.L.; Grzebelus, D.; Cavagnaro, P.; Allender, C.; Brunet, J.; Spooner, D.M.; Van Deynze, A.; Simon, P.W. Genetic structure and domestication of carrot (Daucus carota subsp. sativus) (Apiaceae). Am. J. Bot. 2013, 100, 930–938. [Google Scholar] [CrossRef]
- Rubatzky, V.E.; Quiros, C.F.; Simon, P.W. Carrot and Related Vegetable Umbelliferae; CABI Publishing: Wallingford, UK, 1999. [Google Scholar]
- Shannon, M.; Grieve, C. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Mangal, J.L.; Lal, S.; Hooda, P.S. Salt tolerance in carrot seed crop. Haryana Agric. Univ. J. Res. 1989, 19, 256–259. [Google Scholar]
- Gibberd, M.R.; Turner, N.C.; Storey, R. Influence of saline irrigation on growth, ion accumulation and partitioning, and leaf gas exchange of carrot (Daucus carota L.). Ann. Bot. 2002, 90, 715–724. [Google Scholar] [CrossRef]
- Kumar, S.; Dhingra, A.; Daniell, H. Plastid-expressed betaine aldehyde dehydrogenase gene in carrot cultured cells, roots, and leaves confers enhanced salt tolerance1. Plant Physiol. 2004, 136, 2843–2854. [Google Scholar] [CrossRef] [PubMed]
- Kasiri, M.R.; Hassandokht, M.R.; Kashi, A.; Shahi-Gharahlar, A. Evaluation of genetic diversity in Iranian yellow carrot accessions (Daucus carota var. sativus), an exposed to extinction rooty vegetable, using morphological characters. Int. J. Agric. Crop Sci. 2013, 6, 151–156. [Google Scholar]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef]
- Persson, J.Å.; Wennerholm, M. The Guide of the Mineralization of Kjeldahl Method; Labconsult: Warsaw, Poland, 1999. [Google Scholar]
- Pasławski, P.; Migaszewski, Z.M. The quality of element determinations in plant materials by instrumental methods. Pol. J. Environ. Stud. 2006, 15, 154–164. [Google Scholar]
- Nowosielski, O. The Rules in Development of Fertilizing Strategies in Horticulture; PWRiL Publisher: Warsaw, Poland, 1988. (In Polish) [Google Scholar]
- PN-EN ISO 13395:2001. Water Quality—Determination of Nitrite Nitrogen and Nitrate and the Sum of both by Flow Analysis (CFA and FIA) and Spectrometric Detection; Polish Committee of Standardization: Warsaw, Poland, 2001. (In Polish)
- PN-EN ISO 11732:2005. Water Quality. Determination of Ammonium Nitrogen. Method by flow analysis (CFA and FIA) and Spectrometric Detection; Polish Committee of Standardization: Warsaw, Poland, 2005. (In Polish)
- Gorlach, E.; Curyło, T.; Gambuś, F.; Grzywnowicz, I.; Jasiewicz, C.; Kopeć, M.; Mazur, B.; Olkuśnik, S.; Rogóż, A.; Wiśniowska-Kielian, B. Przewodnik do ćwiczeń z chemii rolnej [A Guidebook to Experimentation in Agricultural Chemistry]; Wyd. AR w: Krakowie, Poland, 1999. (In Polish) [Google Scholar]
- Pfaffl, M.W. Relative expression software tool (REST(C)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 36. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, F.; Inal, A.; Gunes, A.; Alpaslan, M. Impact of exogenous salicylic acid on the growth, antioxidant activity and physiology of carrot plants subjected to combined salinity and boron toxicity. Sci. Hortic. 2007, 113, 120–128. [Google Scholar] [CrossRef]
- De Pascale, S.; Barbieri, G. Yield and quality of carrot as affected by soil salinity from long-term irrigation with saline water. Acta Hortic. 2000, 537, 621–628. [Google Scholar] [CrossRef]
- Nagaz, K.; Masmoudi, M.M.; Ben Mechlia, N. Impacts of irrigation regimes with saline water on carrot productivity and soil salinity. J. Saudi Soc. Agric. Sci. 2012, 11, 19–27. [Google Scholar] [CrossRef]
- Ünlükara, A.; Cemek, B.; Kesmez, D.; Öztürk, A. Carrot (Daucus carota L.): Yield and quality under saline conditions. Anadolu Tarım Bilim. Derg. 2011, 26, 51–56. [Google Scholar] [CrossRef]
- Rasouli, F.; Pouya, A.K.; Karimian, N. Wheat yield and physico-chemical properties of a sodic soil from semi-arid area of Iran as affected by applied gypsum. Geoderma 2013, 193, 246–255. [Google Scholar] [CrossRef]
- Pshenichnikova, T.A.; Doroshkov, A.V.; Osipova, S.V.; Permyakov, A.V.; Permyakova, M.D.; Efimov, V.M.; Afonnikov, D.A. Quantitative characteristics of pubescence in wheat (Triticum aestivum L.) are associated with photosynthetic parameters under conditions of normal and limited water supply. Planta 2018, 249, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.H.; Marschner, H. The Mineral Nutrition of Higher Plants. J. Ecol. 1988, 76, 1250. [Google Scholar] [CrossRef]
- Ashraf, M.; Zafar, R. Time-course changes in organic metabolites and mineral nutrients in germinating maize seeds under salt (NaCl) stress. Seed Sci. Technol. 2000, 28, 641–656. [Google Scholar] [CrossRef]
- Schachtman, D.; Liu, W. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends Plant Sci. 1999, 4, 281–287. [Google Scholar] [CrossRef]
- Golldack, R.; Quigley, F.; Michalowski, C.B.; Kamasani, U.R.; Bohnert, H.J. Salinity stress-tolerant and -sensitive rice (Oryza sativa L.) regulate AKT1-type potassium channel transcripts differently. Plant Mol. Biol. 2003, 51, 71–81. [Google Scholar] [CrossRef]
- Kader, A.; Seidel, T.; Golldack, D.; Lindberg, S. Expressions of OsHKT1, OsHKT2, and OsVHA are differentially regulated under NaCl stress in salt-sensitive and salt-tolerant rice (Oryza sativa L.) cultivars. J. Exp. Bot. 2006, 57, 4257–4268. [Google Scholar] [CrossRef]
- Liu, J.; Shabala, S.; Shabala, L.; Zhou, M.; Meinke, H.; Venkataraman, G.; Chen, Z.; Zeng, F.; Zhao, Q. Tissue-specific regulation of Na+ and K+ transporters explains genotypic differences in salinity stress tolerance in rice. Front. Plant Sci. 2019, 10, 1361. [Google Scholar] [CrossRef]
- Su, H.; Golldack, R.; Zhao, C.; Bohnert, H.J. The expression of HAK-type K+ transporters is regulated in response to salinity stress in Common Ice Plant1. Plant Physiol. 2002, 129, 1482–1493. [Google Scholar] [CrossRef]
- Stassart, J.M.; Neirinckx, L.; De Jaegere, R. The interactions between monovalent cations and calcium during their adsorption on isolated cell walls and absorption by intact barley roots. Ann. Bot. 1981, 47, 647–652. [Google Scholar] [CrossRef]
- Zhu, J. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Kreuzer, I.; Stölzle, S.; Ivashikina, N.; Hedrich, R. Rice K+ uptake channel OsAKT1 is sensitive to salt stress. Planta 2004, 221, 212–221. [Google Scholar] [CrossRef]
- Reuveni, M.; Lerner, H.R.; Poljakoff-Mayber, A. Osmotic adjustment and dynamic changes in distribution of low molecular weight solutes between cellular compartments of carrot and beet root cells exposed to salinity. Am. J. Bot. 1991, 78, 601. [Google Scholar] [CrossRef]
- Bassil, E.; Zhang, S.; Gong, H.; Tajima, H.; Blumwald, E. Cation specificity of vacuolar NHX-type cation/H+ antiporters. Plant Physiol. 2018, 179, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Charfeddine, S.; Charfeddine, M.; Hanana, M.; Gargouri-Bouzid, R. Ectopic expression of a grape vine vacuolar NHX antiporter enhances transgenic potato plant tolerance to salinity. J. Plant Biochem. Biotechnol. 2019, 28, 50–62. [Google Scholar] [CrossRef]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef]
- Li, H.-T.; Liu, H.; Gao, X.-S.; Zhang, H. Knock-out of Arabidopsis AtNHX4 gene enhances tolerance to salt stress. Biochem. Biophys. Res. Commun. 2009, 382, 637–641. [Google Scholar] [CrossRef]
- Sharma, H.; Taneja, M.; Upadhyay, S.K. Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics 2019, 112, 356–370. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; Tiller, K.G.; Smart, M.K. Speciation of cadmium in soil solutions of saline/sodic soils and relationship with cadmium concentrations in potato tubers (Solanum tuberosum L.). Soil Res. 1997, 35, 183–198. [Google Scholar] [CrossRef]
- López-Chuken, U.J.; Young, S.D. Plant screening of halophyte species for cadmium phytoremediation. Zeitschrift für Naturforschung C 2005, 60, 236–243. [Google Scholar]
- Weggler, K.; McLaughlin, M.J.; Graham, R.D. Effect of chloride in soil solution on the plant availability of biosolid-borne cadmium. J. Environ. Qual. 2004, 33, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Smolders, E.; McLaughlin, M.J. Chloride increases cadmium uptake in swiss chard in a resin-buffered nutrient solution. Soil Sci. Soc. Am. J. 1996, 60, 1443–1447. [Google Scholar] [CrossRef]
- Norvell, W.A.; Wu, J.; Hopkins, D.G.; Welch, R.M. Association of cadmium in durum wheat grain with soil chloride and chelate-extractable soil cadmium. Soil Sci. Soc. Am. J. 2000, 64, 2162–2168. [Google Scholar] [CrossRef]
- Ghallab, A.; Usman, A.R.A. Effect of sodium chloride-induced salinity on phyto-availability and speciation of cd in soil solution. Water Air Soil Pollut. 2007, 185, 43–51. [Google Scholar] [CrossRef]
- Lefevre, I.; Marchal, G.; Meerts, P.; Correal, E.; Lutts, S. Chloride salinity reduces cadmium accumulation by the Mediterranean halophyte species Atriplex halimus L. Environ. Exp. Bot. 2009, 65, 142–152. [Google Scholar] [CrossRef]
- Khoshgoftarmanesh, A.H.; Shariatmadari, H.; Karimian, N.; Kalbasi, M.; Van Der Zee, S.E.A.T.M.; Parker, D.R. Salinity and zinc application effects on phytoavailability of cadmium and zinc. Soil Sci. Soc. Am. J. 2004, 68, 1885–1889. [Google Scholar] [CrossRef]
- Khoshgoftar, A.H.; Jaafari, B.; Shariatmadari, H. Effect of salinity on Cd and Zn availability. In Proceedings of the 17th World Congress of Soil Science, Bangkok, Thailand, 12–14 August 2002; Available online: https://pdfs.semanticscholar.org/b23d/6b86bd26ce54ae2a889f3512c732e2cabfbc.pdf (accessed on 25 March 2020).
- Grotz, N.; Fox, T.; Connolly, E.; Park, W.; Guerinot, M.L.; Eide, D. Identification of a family of zinc transporter genes from Arabidopsis that respond to zinc deficiency. Proc. Natl. Acad. Sci. USA 1998, 95, 7220–7224. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Aarts, M.G.M. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef]
- Ma, J.F.; Ueno, D.; Zhao, F.-J.; McGrath, S.P. Subcellular localisation of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Planta 2004, 220, 731–736. [Google Scholar] [CrossRef]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.-F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef]
- Lombi, E.; Tearall, K.L.; Howarth, J.R.; Zhao, F.-J.; Hawkesford, M.J.; McGrath, S.P. Influence of iron status on cadmium and zinc uptake by different ecotypes of the hyperaccumulator thlaspi caerulescens1. Plant Physiol. 2002, 128, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 Is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Inal, A.; Gunes, A.; Pilbeam, D.J.; Kadioglu, Y.K.; Eraslan, F. Concentrations of essential and nonessential elements in shoots and storage roots of carrot grown in NaCl and Na2SO4salinity. X-Ray Spectrom. 2009, 38, 45–51. [Google Scholar] [CrossRef]
- Kleiber, T. Effect of titanium application on lettuce growth under Mn stress. J. Elem. 2016, 22, 329–337. [Google Scholar] [CrossRef]
- Soylemez, S.; Kaya, C.; Dikilitas, S.K. Promotive effects of epibrassinolide on plant growth, fruit yield, antioxidant, and mineral nutrition of saline stressed tomato plants. Pak. J. Bot. 2017, 49, 1655–1661. [Google Scholar]
- Salachna, P.; Piechocki, R. Effects of sodium chloride on growth and mineral nutrition of purpletop vervain. J. Ecol. Eng. 2016, 17, 148–152. [Google Scholar] [CrossRef]
- Cantabella, D.; Piqueras, A.; Acosta-Motos, J.R.; Bernal-Vicente, A.; Hernández, J.A.; Diaz-Vivancos, P. Salt-tolerance mechanisms induced in Stevia rebaudiana Bertoni: Effects on mineral nutrition, antioxidative metabolism and steviol glycoside content. Plant Physiol. Biochem. 2017, 115, 484–496. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2014, 75, 391–404. [Google Scholar] [CrossRef]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates counter plant stress: A Review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Enteshari, S.H.; Jafari, T. The effects of methyl jasmonate and salinity on germination and seedling growth in Ocimum basilicum L. stress. Iran. J. Plant Physiol. 2013, 3, 749–756. [Google Scholar] [CrossRef]
- Velitchkova, M.; Fedina, I.S. Response of photosynthesis of Pisum sativum to salt stress as affected by methyl jasmonate. Photosynthetica 1998, 35, 89–97. [Google Scholar] [CrossRef]
- Kang, D.-J.; Seo, Y.-J.; Lee, J.-D.; Ishii, R.; Kim, K.U.; Shin, D.H.; Park, S.K.; Jang, S.W.; Lee, I.-J. Jasmonic acid differentially affects growth, ion uptake and abscisic acid concentration in salt-tolerant and salt-sensitive rice cultivars. J. Agron. Crop. Sci. 2005, 191, 273–282. [Google Scholar] [CrossRef]
- Pedranzani, H.; Racagni, G.; Alemano, S.; Miersch, O.; Ramírez, I.; Peña-Cortés, H.; Taleisnik, E.; Machado-Domenech, E.; Abdala, G. Salt tolerant tomato plants show increased levels of jasmonic acid. Plant Growth Regul. 2003, 41, 149–158. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| JA Application | Variety | Treatment | Storage Root | Leaves | Total Biomass | Total Dry Weight |
|---|---|---|---|---|---|---|
| No JA | – | – | 36.3 ± 4.11a | 21.6 ± 2.21a | 57.9 ± 6.05a | 9.0 ± 0.91a |
| JA | – | – | 35.9 ± 4.46a | 20.1 ± 2.01a | 56.0 ± 6.26a | 8.4 ± 0.92a |
| – | DLBA | – | 41.1 ± 4.25b | 26.8 ± 1.83b | 68.0 ± 5.88b | 10.7 ± 0.86b |
| – | DH1 | – | 31.1 ± 3.95a | 15.0 ± 1.34a | 46.1 ± 5.28a | 6.7 ± 0.73a |
| – | – | Control | 54.2 ± 1.62b | 27.5 ± 1.74b | 81.7 ± 3.11b | 12.1 ± 0.60b |
| – | – | NaCl | 19.8 ± 1.11a | 15.0 ± 1.37a | 34.8 ± 2.40a | 5.6 ± 0.41a |
| No JA | DLBA | Control | 57.8 ± 1.33d | 35.9 ± 1.32c | 93.7 ± 1.92d | 14.4 ± 0.40d |
| No JA | DLBA | NaCl | 23.8 ± 1.33b | 20.1 ± 1.32b | 43.9 ± 1.92b | 7.3 ± 0.40b |
| No JA | DH1 | Control | 48.8 ± 1.33c | 20.8 ± 1.32b | 69.6 ± 1.92c | 10.3 ± 0.40c |
| No JA | DH1 | NaCl | 14.9 ± 1.33a | 9.6 ± 1.32a | 24.5 ± 1.92a | 3.9 ± 0.40a |
| JA | DLBA | Control | 62.1 ± 1.48d | 31.6 ± 1.47c | 93.7 ± 2.15d | 14.3 ± 0.45d |
| JA | DLBA | NaCl | 25.1 ± 1.33b | 20.6 ± 1.32b | 45.7 ± 1.92b | 7.3 ± 0.40b |
| JA | DH1 | Control | 48.6 ± 1.48c | 21.1 ± 1.47b | 69.7 ± 1.92c | 9.5 ± 0.45c |
| JA | DH1 | NaCl | 15.6 ± 1.33a | 9.7 ± 1.32a | 25.3 ± 2.15a | 3.8 ± 0.40a |
| Element | JA Application | Variety | Treatment | Uptake (mmol) by Single Plant | |||
|---|---|---|---|---|---|---|---|
| Storage Root | Leaves | Whole Plant | Balance in Soil–Plant † (%) | ||||
| Na | No JA | – | – | 1.73 ± 0.278a | 2.33 ± 0.526a | 4.12 ± 0.209a | 8.27 ± 0.791b |
| Na | JA | – | – | 1.50 ± 0.287a | 2.39 ± 0.416a | 3.57 ± 0.222a | 5.89 ± 0.839a |
| Na | – | DLBA | – | 1.90 ± 0.306b | 3.06 ± 0.563a | 4.77 ± 0.226b | 5.63 ± 0.816a |
| Na | – | DH1 | – | 1.35 ± 0.223a | 1.66 ± 0.264a | 2.93 ± 0.226a | 8.52 ± 0.816b |
| Na | – | – | Control | 0.62 ± 0.082a | 0.70 ± 0.065a | 1.30 ± 0.226a | 13.07 ± 0.839b |
| Na | – | – | NaCl | 2.53 ± 0.196b | 3.86 ± 0.378b | 6.39 ± 0.209b | 1.09 ± 0.791a |
| Na | No JA | DLBA | Control | 0.74 ± 0.196a | 0.81 ± 0.273a | 1.55 ± 0.355a | 12.31 ± 1.390bc |
| Na | No JA | DLBA | NaCl | 2.90 ± 0.196c | 4.79 ± 0.273c | 7.69 ± 0.355d | 1.28 ± 0.390a |
| Na | No JA | DH1 | Control | 0.61 ± 0.196a | 0.70 ± 0.273a | 1.31 ± 0.355a | 18.14 ± 1.390c |
| Na | No JA | DH1 | NaCl | 2.69 ± 0.196c | 3.25 ± 0.273c | 5.94 ± 0.355c | 1.35 ± 0.390a |
| Na | JA | DLBA | Control | 0.56 ± 0.219a | 0.61 ± 0.305a | 1.17 ± 0.397a | 7.86 ± 1.554b |
| Na | JA | DLBA | NaCl | 3.12 ± 0.196c | 5.54 ± 0.273c | 8.66 ± 0.355d | 1.08 ± 0.390a |
| Na | JA | DH1 | Control | 0.53 ± 0.219a | 0.63 ± 0.305a | 1.16 ± 0.397a | 13.96 ± 1.554c |
| Na | JA | DH1 | NaCl | 1.42 ± 0.196b | 1.87 ± 0.273b | 3.29 ± 0.355b | 0.65 ± 0.390a |
| Cl | No JA | – | – | 1.42 ± 0.290a | 3.21 ± 0.481a | 4.73 ± 0.169a | 40.00 ± 2.667a |
| Cl | JA | – | – | 1.34 ± 0.317a | 3.31 ± 0.528a | 4.29 ± 0.174a | 39.66 ± 2.828a |
| Cl | – | DLBA | – | 1.70 ± 0.343b | 4.39 ± 0.557b | 5.91 ± 0.169b | 36.03 ± 2.749a |
| Cl | – | DH1 | – | 1.06 ± 0.233a | 2.13 ± 0.232a | 3.11 ± 0169a | 43.63 ± 2.749a |
| Cl | – | – | Control | 0.21 ± 0.021a | 1.68 ± 0.132a | 1.90 ± 0.174a | 78.59 ± 2.828b |
| Cl | – | – | NaCl | 2.44 ± 0.204b | 4.68 ± 0.467b | 7.12 ± 0.164b | 1.07 ± 2.667a |
| Cl | No JA | DLBA | Control | 0.24 ± 0.169a | 1.84 ± 0.233a | 2.08 ± 0.274a | 68.49 ± 4.737b |
| Cl | No JA | DLBA | NaCl | 2.86 ± 0.169c | 6.49 ± 0.233c | 9.35 ± 0.274d | 1.39 ± 0.737a |
| Cl | No JA | DH1 | Control | 0.21 ± 0.169a | 1.36 ± 0.233a | 1.57 ± 0.274a | 88.92 ± 4.737c |
| Cl | No JA | DH1 | NaCl | 2.38 ± 0.169c | 3.56 ± 0.233b | 5.94 ± 0.274c | 1.19 ± 0.737a |
| Cl | JA | DLBA | Control | 0.22 ± 0.189a | 2.22 ± 0.260ab | 2.44 ± 0.306a | 73.15 ± 5.296b |
| Cl | JA | DLBA | NaCl | 3.18 ± 0.169c | 6.59 ± 0.233c | 9.77 ± 0.274d | 1.08 ± 0.437a |
| Cl | JA | DH1 | Control | 0.17 ± 0.189a | 1.35 ± 0.260a | 1.52 ± 0.306a | 83.80 ± 5.296c |
| Cl | JA | DH1 | NaCl | 1.32 ± 0.169b | 2.08 ± 0.233a | 3.40 ± 0.274b | 0.60 ± 0.237a |
| JA Application | Variety | Treatment | Content | Uptake | |
|---|---|---|---|---|---|
| Storage Root | Leaves | Whole Plant | |||
| No JA | – | – | 2 218.6 ± 54.5a | 4 146.6 ± 272.9a | 26.4 ± 0.76b |
| JA | – | – | 2 191.0 ± 57.8a | 4 305.7 ± 297.4a | 23.0 ± 0.81a |
| – | DLBA | – | 2 160.8 ± 56.1a | 4 350.3 ± 342.1b | 29.2 ± 0.78b |
| – | DH1 | – | 2 248.8 ± 56.1a | 4 102.0 ± 212.1a | 20.2 ± 0.78a |
| – | – | Control | 1 430.5 ± 57.8a | 3 074.5 ± 60.5a | 23.8 ± 0.81a |
| – | – | NaCl | 2 979.1 ± 54.5b | 5 377.7 ± 108.9b | 25.6 ± 0.76a |
| No JA | DLBA | Control | 1 321.5 ± 46.2a | 2 900.1 ± 128.5a | 27.6 ± 2.2cd |
| No JA | DLBA | NaCl | 2 954.2 ± 115.8b | 5 662.3 ± 159.3c | 30.7 ± 1.2d |
| No JA | DH1 | Control | 1 594.8 ± 65.8a | 3 133.4 ± 83.4a | 21.9 ± 1.6bc |
| No JA | DH1 | NaCl | 3 003.9 ± 69.7b | 4 890.6 ± 91.7b | 25.5 ± 1.8bcd |
| JA | DLBA | Control | 1 288.5 ± 126.1a | 2 943.1 ± 108.6a | 26.4 ± 1.7bcd |
| JA | DLBA | NaCl | 3 079.2 ± 213.5b | 5 895.8 ± 92.2c | 32.0 ± 1.8d |
| JA | DH1 | Control | 1 517.5 ± 79.5a | 3 321.5 ± 47.4a | 19.4 ± 0.5ab |
| JA | DH1 | NaCl | 2 879.2 ± 73.8b | 5 062.4 ± 108.1b | 14.2 ± 0.4a |
| JA Application | Variety | Treatment | Na | Cl |
|---|---|---|---|---|
| No JA | – | – | 29.98 ± 6.652a | 26.25 ± 6.007a |
| JA | – | – | 30.40 ± 6.545a | 30.45 ± 6.865a |
| – | DLBA | – | 31.87 ± 6.986b | 31.30 ± 7.077b |
| – | DH1 | – | 28.18 ± 6.191a | 25.19 ± 5.624a |
| – | – | Control | 0.95 ± 0.061a | 0.32 ± 0.011a |
| – | – | NaCl | 56.18 ± 1.644b | 53.37 ± 2.078b |
| No JA | DLBA | Control | 0.92 ± 0.154a | 0.33 ± 0.066a |
| No JA | DLBA | NaCl | 59.92 ± 2.848c | 54.87 ± 2.664c |
| No JA | DH1 | Control | 0.99 ± 0.176a | 0.30 ± 0.065a |
| No JA | DH1 | NaCl | 56.88 ± 2.572bc | 49.52 ± 2.664b |
| JA | DLBA | Control | 1.06 ± 0.099a | 0.33 ± 0.097a |
| JA | DLBA | NaCl | 59.41 ± 3.294c | 63.49 ± 2.664c |
| JA | DH1 | Control | 0.83 ± 0.051a | 0.34 ± 0.097a |
| JA | DH1 | NaCl | 48.53 ± 2.226b | 45.62 ± 2.664b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smoleń, S.; Lukasiewicz, A.; Klimek-Chodacka, M.; Baranski, R. Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress. Agronomy 2020, 10, 659. https://doi.org/10.3390/agronomy10050659
Smoleń S, Lukasiewicz A, Klimek-Chodacka M, Baranski R. Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress. Agronomy. 2020; 10(5):659. https://doi.org/10.3390/agronomy10050659
Chicago/Turabian StyleSmoleń, Sylwester, Aneta Lukasiewicz, Magdalena Klimek-Chodacka, and Rafal Baranski. 2020. "Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress" Agronomy 10, no. 5: 659. https://doi.org/10.3390/agronomy10050659
APA StyleSmoleń, S., Lukasiewicz, A., Klimek-Chodacka, M., & Baranski, R. (2020). Effect of Soil Salinity and Foliar Application of Jasmonic Acid on Mineral Balance of Carrot Plants Tolerant and Sensitive to Salt Stress. Agronomy, 10(5), 659. https://doi.org/10.3390/agronomy10050659