Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century

 , , , ,
, , , , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Agro-Morphological Traits
2.3. Grain Morphometrics
2.4. DNA Isolation
2.5. ISSR Analysis
2.6. Scoring and Data Analysis
3. Results
3.1. Summary of Passport Data
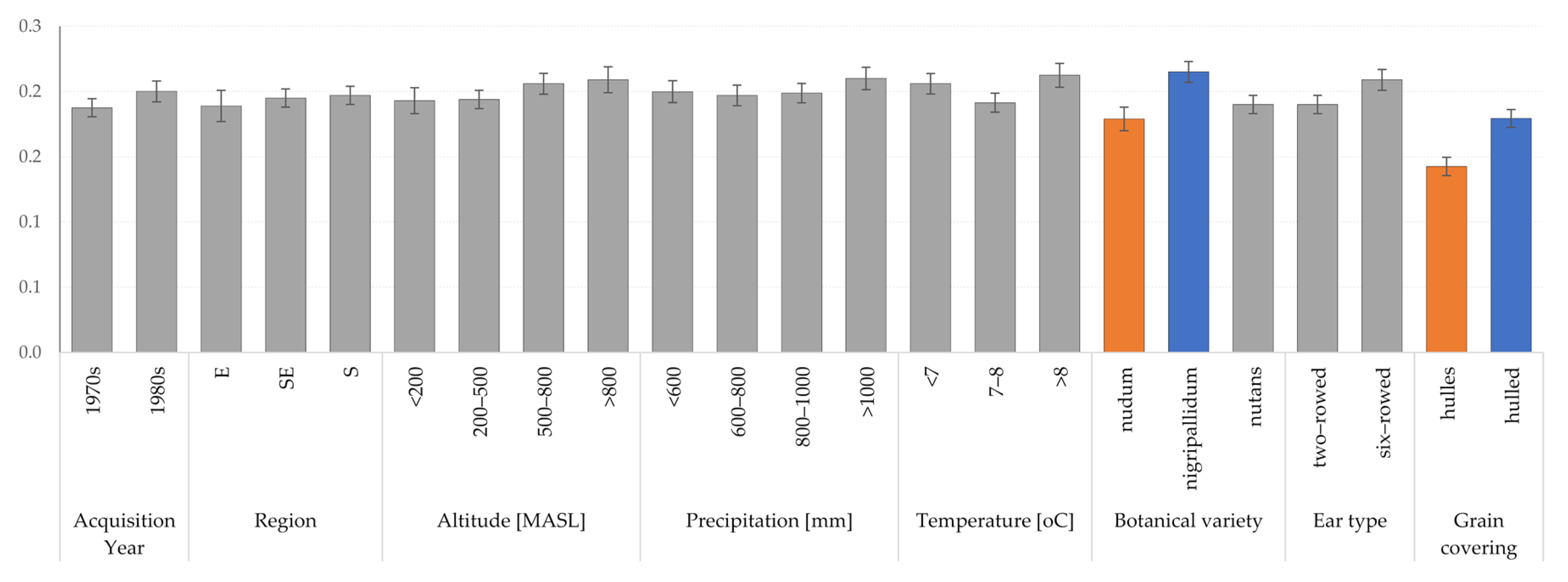
3.2. Summary of Eco-Geographic Conditions
3.3. Agro-Morphological Diversity
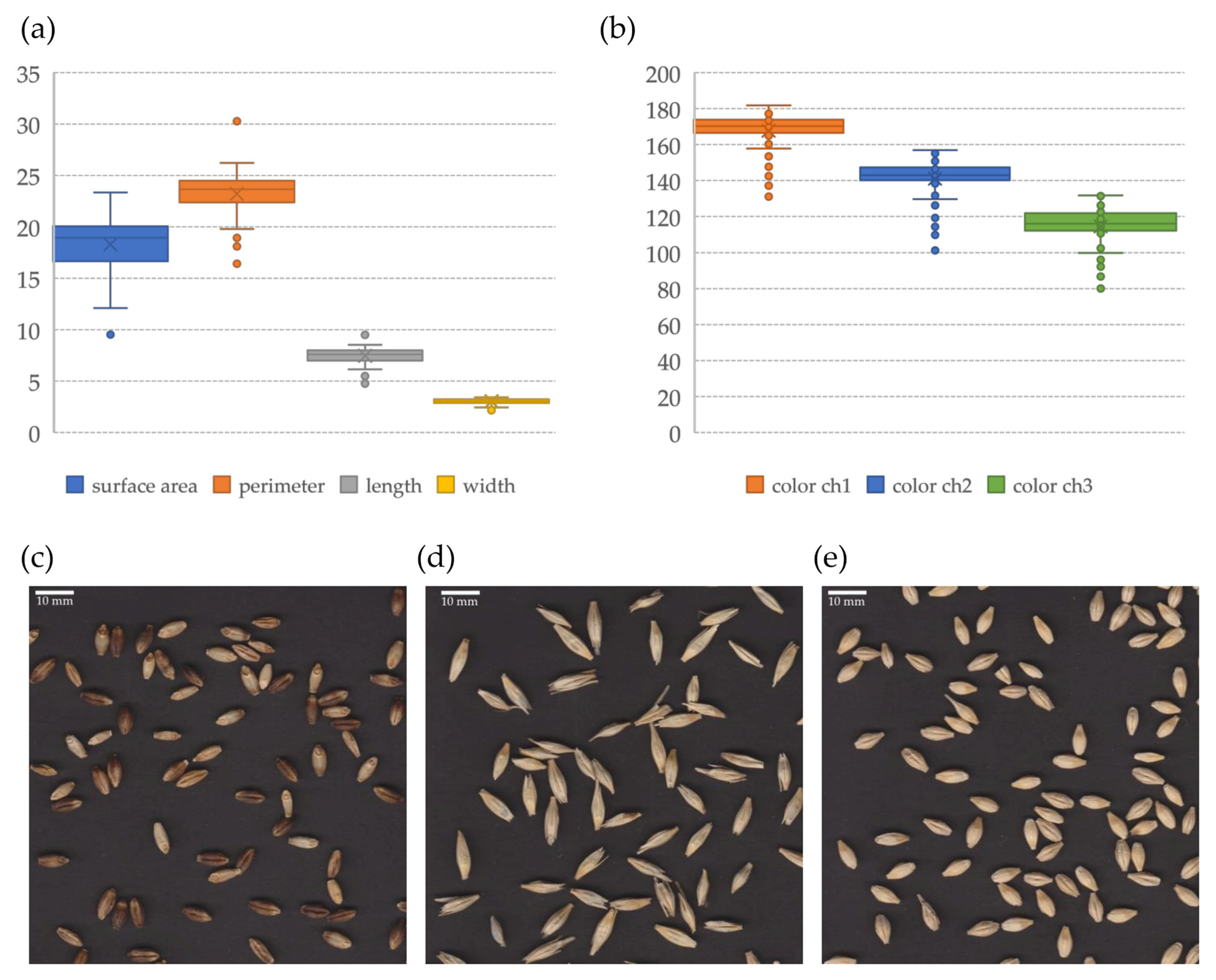
3.4. Grain Morphology
3.4.1. Variation within the Collection
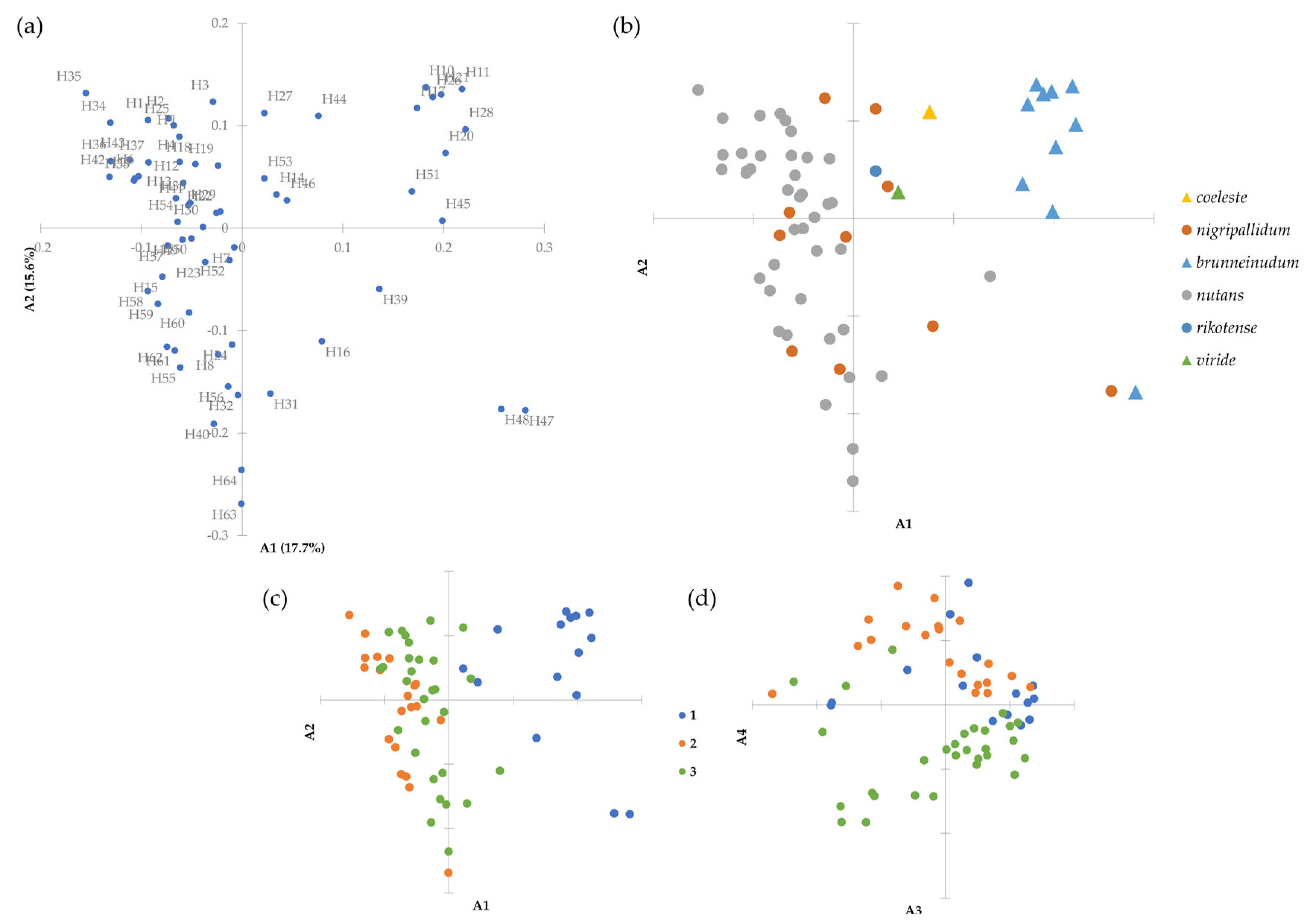
3.4.2. Variation within Groups
3.4.3. Clustering Analysis
3.5. Genetic Analysis
3.5.1. Marker Efficiency
3.5.2. Genetic Diversity
3.5.3. Genetic Distance
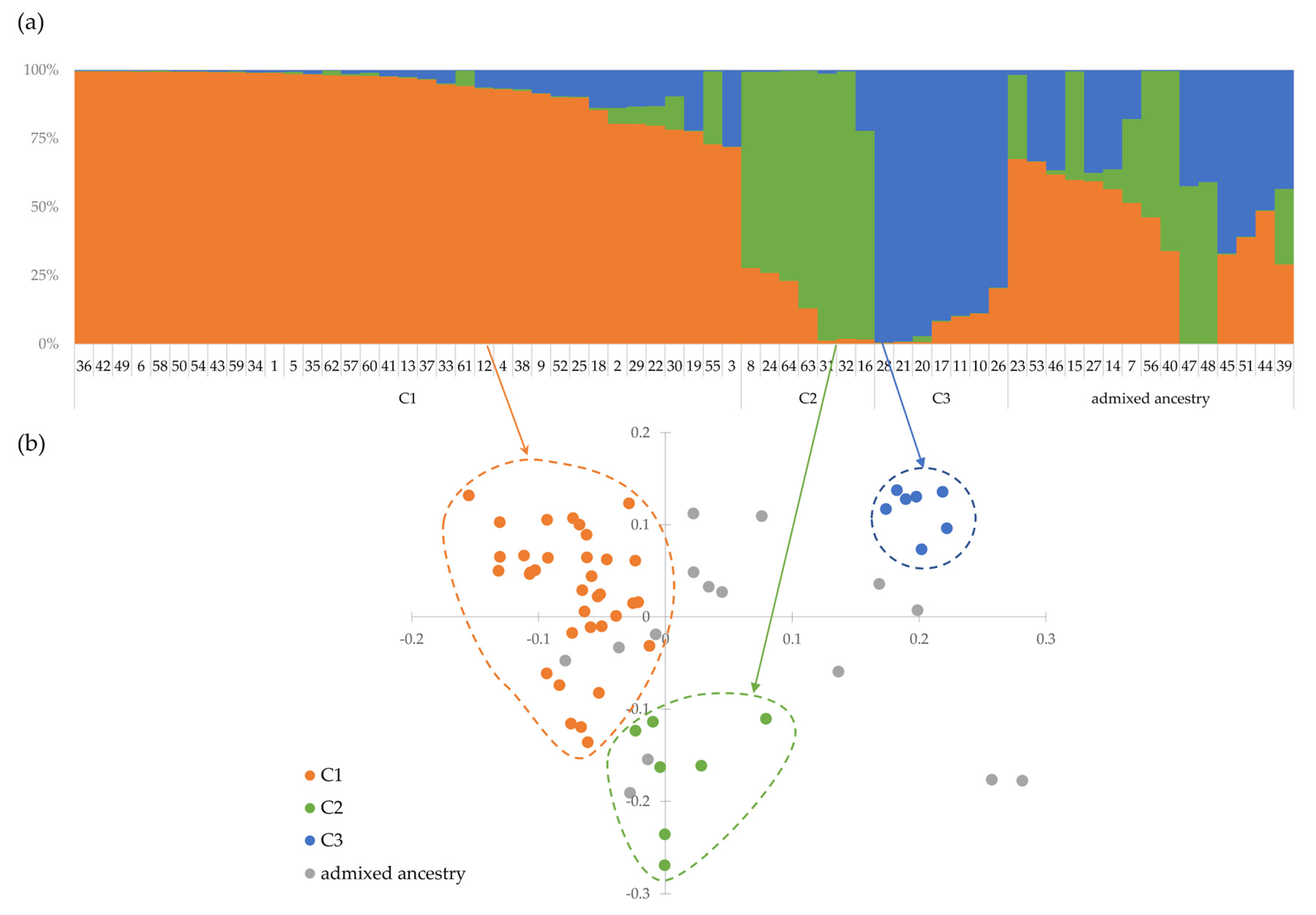
3.5.4. Clustering Analysis
3.5.5. Population Structure
3.6. Joint Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Faostat, Statistical Databases; Food and Agriculture Organization of the United Nations: Roma, Italy, 2018. [Google Scholar]
- Friedt, W.; Horsley, R.D.; Harvey, B.L.; Poulsen, D.M.; Lance, R.C.; Ceccarelli, S.; Grando, S.; Capettini, F. Barley breeding history, progress, objectives, and technology. In Barley: Production, Improvement, and Uses; Ullrich, S.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 160–220. [Google Scholar]
- European Commission. EU Plant Variety Database. 2020. Available online: https://ec.europa.eu/food/plant/plant_propagation_material/plant_variety_catalogues_databases/search/public/index.cfm (accessed on 10 November 2020).
- Caldwell, K.S.; Russell, J.; Langridge, P.; Powell, W. Extreme population-dependent linkage disequilibrium detected in an inbreeding plant species, Hordeum vulgare. Genetics 2006, 172, 557–567. [Google Scholar] [CrossRef] [PubMed]
- FAO. WIEWS—World Information and Early Warning System on Plant Genetic Resources for Food and Agriculture; FAO: Roma, Italy, 2018. [Google Scholar]
- NCPGR. EGISET. 2015. Available online: https://wyszukiwarka.ihar.edu.pl/pl (accessed on 10 November 2020).
- Azeez, M.A.; Adubi, A.O.; Durodola, F.A. Landraces and crop genetic improvement. In Rediscovery of Landraces as a Resource for the Future; IntechOpen: London, UK, 2018. [Google Scholar]
- Schmalenbach, I.; Körber, N.; Pillen, K. Selecting a set of wild barley introgression lines and verification of QTL effects for resistance to powdery mildew and leaf rust. Theor. Appl. Genet. 2008, 117, 1093–1106. [Google Scholar] [CrossRef] [PubMed]
- Kalladan, R.; Worch, S.; Rolletschek, H.; Harshavardhan, V.T.; Kuntze, L.; Seiler, C.; Sreenivasulu, N.; Röder, M.S. Identification of quantitative trait loci contributing to yield and seed quality parameters under terminal drought in barley advanced backcross lines. Mol. Breed. 2013, 32, 71–90. [Google Scholar] [CrossRef]
- Fan, Y.; Shabala, S.; Ma, Y.; Xu, R.; Zhou, M. Using QTL mapping to investigate the relationships between abiotic stress tolerance (drought and salinity) and agronomic and physiological traits. BMC Genom. 2015, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mora, F.; Quitral, Y.A.; Matus, I.; Russell, J.; Waugh, R.; Del Pozo, A. SNP-based QTL mapping of 15 complex traits in barley under rain-fed and well-watered conditions by a mixed modeling approach. Front. Plant Sci. 2016, 7, 909. [Google Scholar] [CrossRef] [PubMed]
- Sayed, M.A.; Hamada, A.; Lèon, J.; Naz, A.A. Genetic mapping reveals novel exotic QTL alleles for seminal root architecture in barley advanced backcross double haploid population. Euphytica 2017, 213, 2. [Google Scholar] [CrossRef]
- Kloc, G.; Dostatny, D.F.; Sekutowski, T.; Podyma, W. The role of collection missions in gathering plant genetic resources material. Ann. UMCS Sectio E Agric. 2018, 73, 4. [Google Scholar] [CrossRef]
- Hammer, K.; Górski, M.; Hanelt, P.; Kühn, F.; Kulpa, W.; Schultze-Motel, J. Variability of wheat land-races from Czechoslovakia and Poland. Die Kult. 1981, 29, 91–97. [Google Scholar] [CrossRef]
- Hanelt, P.; Hammer, K. Bericht über eine Reise nach der VR Polen 1976 zur Sammlung autochthoner Sippen von Kulturpflanzen. Die Kult. 1977, 25, 33–44. [Google Scholar] [CrossRef]
- Kulpa, W.; Górski, M. Zasoby miejscowych form roślin uprawnych Cz. I. Wyniki eksploracji Płaskowyżu Kolbuszowskiego, Pogórza Karpackiego i Beskidów w latach 1976 i 1978. [Resources of landraces of cultivated plants Part I. Results of the exploration of the Kolbuszowski Plateau, Carpathian Foothills and Beskids in 1976 and 1978]. Biul. IHAR 1986, 160, 27–45. [Google Scholar]
- Mansfeld, R. Das morphologische System der Saatgerste, Hordeum vulgare L. sl. Der Züchter 1950, 20, 8–24. [Google Scholar] [CrossRef]
- Whan, A.P.; Smith, A.B.; Cavanagh, C.R.; Ral, J.-P.F.; Shaw, L.M.; Howitt, C.A.; Bischof, L. GrainScan: A low cost, fast method for grain size and colour measurements. Plant Methods 2014, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.; Cameron, S.; Parra, J. WorldClim. In Version 1.4 (Release 3). A Square Kilometer Resolution Database of Global Terrestrial Surface Climate; 2004; Available online: https://worldclim.org/data/worldclim21.html (accessed on 1 November 2020).
- Gower, J.C. Generalized procrustes analysis. Psychometrika 1975, 40, 33–51. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- QGIS.org QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2020. Available online: http://www.gdal.org/ogr/ogr_formats.html (accessed on 10 November 2020).
- South, A. Rworldxtra: Country Boundaries at High Resolution, R Package Version 1.01; 2012. Available online: https://rdrr.io/cran/rworldxtra/ (accessed on 1 November 2020).
- GIMP. 2019. Available online: https://www.gimp.org (accessed on 10 November 2020).
- Amezrou, R.; Gyawali, S.; Belqadi, L.; Chao, S.; Arbaoui, M.; Mamidi, S.; Rehman, S.; Sreedasyam, A.; Verma, R.P.S. Molecular and phenotypic diversity of ICARDA spring barley (Hordeum vulgare L.) collection. Genet. Resour. Crop Evol. 2018, 65, 255–269. [Google Scholar] [CrossRef]
- Mohammadi, S.A.; Sisi, N.A.; Sadeghzadeh, B. The influence of breeding history, origin and growth type on population structure of barley as revealed by SSR markers. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Asmamaw, M.; Keneni, G.; Tesfaye, K. Genetic diversity of Ethiopian durum wheat (Triticum durum Desf.) landrace collections as reveled by SSR markers. Adv. Crop Sci. Technol. 2019, 7, 1–9. [Google Scholar] [CrossRef]
- Singh, N.; Choudhury, D.R.; Tiwari, G.; Singh, A.K.; Kumar, S.; Srinivasan, K.; Tyagi, R.; Sharma, A.; Singh, N.; Singh, R. Genetic diversity trend in Indian rice varieties: An analysis using SSR markers. BMC Genet. 2016, 17, 127. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, E. Conserving the Wild Relatives of Crops; Nternational Board for Plant Genetic Resources: Rome, Italy, 1988; pp. 1–45. [Google Scholar]
- Hawkes, J.G. The Diversity of Crop Plants; Harvard Univerity Press: Cambridge, MA, USA, 1983; pp. 1–184. [Google Scholar]
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Plant Genetic Resources for Food and Agriculture; FAO: Roma, Italy, 1998. [Google Scholar]
- Brown, A.H. Genes in the Field, Chapter the Genetic Structure of Crop Landraces and the Challenge to Conserve Them In Situ on Farms; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 29–48. [Google Scholar]
- Boczkowska, M.; Łapiński, B.; Kordulasińska, I.; Dostatny, D.F.; Czembor, J.H. Promoting the use of common oat genetic resources through diversity analysis and core collection construction. PLoS ONE 2016, 11, e0167855. [Google Scholar] [CrossRef] [PubMed]
- Boczkowska, M.; Tarczyk, E. Genetic diversity among Polish landraces of common oat (Avena sativa L.). Genet. Resour. Crop Evol. 2013, 60, 2157–2169. [Google Scholar] [CrossRef]
- Boczkowska, M.; Onyśk, A. Unused genetic resources: A case study of Polish common oat germplasm. Ann. Appl. Biol. 2016, 169, 155–165. [Google Scholar] [CrossRef]
- Arseniuk, E.; Krzymuski, J.; Martyniak, J.; Oleksiak, T. Historia Hodowli i Nasiennictwa na Ziemiach Polskich w XX Wieku (The History of Breeding and Seed Production on Polish Territory in the XX Century); IHAR Radzików: Poznań, Poland, 2003. [Google Scholar]
- COBORU. Krajowy Rejestr Odmian Gatunków Roślin Uprawnych [National Register of Cultivated Plant Species Cultivars]; Centralny Ośrodek Badania Odmian Roślin Uprawnych: Słupia Wielka, Poland, 2020. [Google Scholar]
- Von Bothmer, R.; van Hintum, T.; Knüpffer, H.; Sato, K. Diversity in barley (Hordeum vulgare); Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Frankel, O.H.; Brown, A.H.; Burdon, J.J. The Conservation of Plant Biodiversity; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Arca, M.; Gouesnard, B.; Mary-Huard, T.; Le Paslier, M.-C.; Bauland, C.; Madur, D.; Combes, V.; Charcosset, A.D.; Nicolas, S.D. Genome-wide SNP genotyping of DNA pools identifies untapped landraces and genomic regions that could enrich the maize breeding pool. bioRxiv 2020. [Google Scholar] [CrossRef]
- Riaz, A.; Hathorn, A.; Dinglasan, E.; Ziems, L.; Richard, C.; Singh, D.; Mitrofanova, O.; Afanasenko, O.; Aitken, E.; Godwin, I. Into the vault of the Vavilov wheats: Old diversity for new alleles. Genet. Resour. Crop Evol. 2017, 64, 531–544. [Google Scholar] [CrossRef]
- Riaz, A.; Athiyannan, N.; Periyannan, S.K.; Afanasenko, O.; Mitrofanova, O.P.; Platz, G.J.; Aitken, E.A.; Snowdon, R.J.; Lagudah, E.S.; Hickey, L.T. Unlocking new alleles for leaf rust resistance in the Vavilov wheat collection. Theor. Appl. Genet. 2018, 131, 127–144. [Google Scholar] [CrossRef]
- Boczkowska, M.; Zebrowski, J.; Nowosielski, J.; Kordulasińska, I.; Nowosielska, D.; Podyma, W. Environmentally-related genotypic, phenotypic and metabolic diversity of oat (Avena sativa L.) landraces based on 67 Polish accessions. Genet. Resour. Crop Evol. 2017, 64, 1829–1840. [Google Scholar] [CrossRef]
- Chen, F.; Chen, D.; Vallés, M.-P.; Gao, Z.; Chen, X. Analysis of diversity in Chinese cultivated barley with simple sequence repeats: Differences between eco-geographic populations. Biochem. Genet. 2010, 48, 44–56. [Google Scholar] [CrossRef]
- Lasa, J.; Igartua, E.; Ciudad, F.; Codesal, P.; Garcíaa, E.; Gracia, M.; Medina, B.; Romagosa, I.; Molina-Cano, J.; Montoya, J. Morphological and agronomical diversity patterns in the Spanish barley core collection. Hereditas 2001, 135, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Usubaliev, B.; Brantestam, A.K.; Salomon, B.; Garkava-Gustavson, L.; von Bothmer, R. Genetic diversity in farmer grown spring barley material from Kyrgyzstan. Genet. Resour. Crop Evol. 2013, 60, 1843–1858. [Google Scholar] [CrossRef]
- Komatsuda, T.; Pourkheirandish, M.; He, C.; Azhaguvel, P.; Kanamori, H.; Perovic, D.; Stein, N.; Graner, A.; Wicker, T.; Tagiri, A. Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene. Proc. Natl. Acad. Sci. USA 2007, 104, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Youssef, H.M.; Eggert, K.; Koppolu, R.; Alqudah, A.M.; Poursarebani, N.; Fazeli, A.; Sakuma, S.; Tagiri, A.; Rutten, T.; Govind, G. VRS2 regulates hormone-mediated inflorescence patterning in barley. Nat. Genet. 2017, 49, 157–161. [Google Scholar] [CrossRef]
- Bull, H.; Casao, M.C.; Zwirek, M.; Flavell, A.J.; Thomas, W.T.; Guo, W.; Zhang, R.; Rapazote-Flores, P.; Kyriakidis, S.; Russell, J. Barley SIX-ROWED SPIKE3 encodes a putative Jumonji C-type H3K9me2/me3 demethylase that represses lateral spikelet fertility. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- van Esse, G.W.; Walla, A.; Finke, A.; Koornneef, M.; Pecinka, A.; Von Korff, M. Six-Rowed Spike3 (VRS3) is a histone demethylase that controls lateral spikelet development in barley. Plant Physiol. 2017, 174, 2397–2408. [Google Scholar] [CrossRef]
- Koppolu, R.; Anwar, N.; Sakuma, S.; Tagiri, A.; Lundqvist, U.; Pourkheirandish, M.; Rutten, T.; Seiler, C.; Himmelbach, A.; Ariyadasa, R. Six-rowed spike4 (Vrs4) controls spikelet determinacy and row-type in barley. Proc. Natl. Acad. Sci. USA 2013, 110, 13198–13203. [Google Scholar] [CrossRef]
- Ramsay, L.; Comadran, J.; Druka, A.; Marshall, D.F.; Thomas, W.T.; Macaulay, M.; MacKenzie, K.; Simpson, C.; Fuller, J.; Bonar, N. INTERMEDIUM-C, a modifier of lateral spikelet fertility in barley, is an ortholog of the maize domestication gene TEOSINTE BRANCHED 1. Nat. Genet. 2011, 43, 169–172. [Google Scholar] [CrossRef]
- Saisho, D.; Pourkheirandish, M.; Kanamori, H.; Matsumoto, T.; Komatsuda, T. Allelic variation of row type gene Vrs1 in barley and implication of the functional divergence. Breed. Sci. 2009, 59, 621–628. [Google Scholar] [CrossRef][Green Version]
- Youssef, H.M.; Mascher, M.; Ayoub, M.A.; Stein, N.; Kilian, B.; Schnurbusch, T. Natural diversity of inflorescence architecture traces cryptic domestication genes in barley (Hordeum vulgare L.). Genet. Resour. Crop Evol. 2017, 64, 843–853. [Google Scholar] [CrossRef]
- Li, Y.; Long, C.; Kato, K.; Yang, C.; Sato, K. Indigenous knowledge and traditional conservation of hulless barley (Hordeum vulgare) germplasm resources in the Tibetan communities of Shangri-la, Yunnan, SW China. Genet. Resour. Crop Evol. 2011, 58, 645–655. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Taketa, S.; Kikuchi, S.; Awayama, T.; Yamamoto, S.; Ichii, M.; Kawasaki, S. Monophyletic origin of naked barley inferred from molecular analyses of a marker closely linked to the naked caryopsis gene (nud). Theor. Appl. Genet. 2004, 108, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Taketa, S.; Amano, S.; Tsujino, Y.; Sato, T.; Saisho, D.; Kakeda, K.; Nomura, M.; Suzuki, T.; Matsumoto, T.; Sato, K. Barley grain with adhering hulls is controlled by an ERF family transcription factor gene regulating a lipid biosynthesis pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 4062–4067. [Google Scholar] [CrossRef] [PubMed]
- Assefa, A.; Labuschagne, M.T. Phenotypic variation in barley (Hordeum vulgare L.) landraces from north Shewa in Ethiopia. Biodivers. Conserv. 2004, 13, 1441–1451. [Google Scholar] [CrossRef]
- Berglung, P.; Holm, E.; Fastnaught, C. Hulless barley: Alternative uses. Barley Newsl. 1993, 36, 130–131. [Google Scholar]
- Dziamba, S.; Rachon, L. Differentiation of yield component in naked-grained and husked spring barley varieties grown in pure and mixed. Biul. Inst. Hod. Aklim. Rosl. 1988, 167, 79–85. [Google Scholar]
- Gol’denberg, Z.; Kvachadze, M. Content of protein and of the amino acid tryptophan in the grain of some forms of barley. Soobshcheniya Akad. Nauk Gruz. SSR 1990, 139, 397–400. [Google Scholar]
- Oscarsson, M.; Andersson, R.; Salomonsson, A.-C.; Åman, P. Chemical composition of barley samples focusing on dietary fibre components. J. Cereal Sci. 1996, 24, 161–170. [Google Scholar] [CrossRef]
- Bhatty, R. The potential of hull-less barley, a review. Cereal Chem. 1986, 63, 97–103. [Google Scholar]
- Peterson, C.; Shelton, D.; Martin, T.; Sears, R.; Williams, E.; Graybosch, R. Grain color stability and classification of hard white wheat in the US. Euphytica 2001, 119, 101–107. [Google Scholar] [CrossRef]
- Kehel, Z.; Sanchez-Garcia, M.; El Baouchi, A.; Aberkane, H.; Tsivelikas, A.; Chen, C.; Amri, A. Predictive characterization for seed morphometric traits for genebank accessions using genomic selection. Front. Ecol. Evol. 2020, 8, 32. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Accession Number with Country Prefix | Botanical Variety | Acquisition Year | Latitude | Longitude | Altitude (Meters above Sea Level) | Region | Resistance |
|---|---|---|---|---|---|---|---|---|
| H1 | PL 40004 | 4 | 1978 | 49 36 N | 21 44 E | 400 | SE | sr, nb |
| H2 | PL 40068 | 4 | 1978 | 49 37 N | 22 06 E | 400 | SE | sr, nb, s |
| H3 | PL 40070 | 3 | 1978 | 49 33 N | 21 56 E | 400 | SE | sr, nb, s |
| H4 | PL 40073 | 4 | 1978 | 49 36 N | 20 29 E | 860 | S | sr, nb, s |
| H5 | PL 40215 | 4 | 1978 | 49 30 N | 22 05 E | 900 | SE | sr, nb, s |
| H6 | PL 40552 | 4 | 1977 | 52 05 N | 22 07 E | 160 | E | pm, nb |
| H7 | PL 40553 | 3 | 1977 | 50 49 N | 22 19 E | 260 | SE | nb |
| H8 | PL 40554 | 4 | 1977 | 50 22 N | 21 50 E | 200 | SE | nb |
| H9 | PL 41265 | 4 | 1978 | 50 18 N | 21 45 E | 220 | SE | nb, s |
| H10 | PL 41266 | 1 | 1978 | 49 32 N | 22 06 E | 400 | SE | sr, nb, s |
| H11 | PL 41267 | 1 | 1978 | 49 33 N | 20 33 E | 340 | S | sr, nb, s |
| H12 | PL 41268 | 4 | 1978 | 49 36 N | 21 03 E | 400 | SE | sr, nb, s |
| H13 | PL 41269 | 4 | 1978 | 49 32 N | 21 02 E | 440 | SE | sr, nb, s |
| H14 | PL 41270 | 3 | 1978 | 49 33 N | 20 59 E | 400 | S | nb, s |
| H15 | PL 41271 | 4 | 1978 | 49 33 N | 20 59 E | 400 | S | sr, nb, s |
| H16 | PL 41272 | 3 | 1978 | 49 27 N | 20 59 E | 680 | S | sr, nb, s |
| H17 | PL 41273 | 1 | 1978 | 49 39 N | 20 36 E | 500 | S | sr, nb, s |
| H18 | PL 41274 | 4 | 1978 | 49 41 N | 20 21 E | 640 | S | sr, nb, s |
| H19 | PL 41279 | 4 | 1978 | 49 41N | 20 21 E | 640 | S | sr, nb, s |
| H20 | PL 41280 | 1 | 1978 | 49 41 N | 20 21 E | 640 | S | sr, nb, s |
| H21 | PL 41281 | 1 | 1978 | 49 36 N | 21 03 E | 360 | SE | sr, nb, s |
| H22 | PL 41282 | 4 | 1978 | 49 22 N | 20 11 E | 900 | S | sr, nb, s |
| H23 | PL 41284 | 4 | 1978 | 49 24 N | 20 18 E | 500 | S | nb, s |
| H24 | PL 41365 | 4 | 1979 | 53 16 N | 23 47 E | 180 | NE | sr, nb, s |
| H25 | PL 41430 | 4 | 1978 | 49 33 N | 20 33 E | 380 | S | sr, nb, s |
| H26 | PL 41431 | 1 | 1981 | 49 32 N | 21 02 E | 400 | SE | sr, s |
| H27 | PL 41432 | 3 | 1981 | 49 33 N | 20 33 E | 400 | S | sr, s |
| H28 | PL 41433 | 1 | 1981 | 49 36 N | 21 03 E | 360 | SE | s |
| H29 | PL 41434 | 4 | 1981 | 49 26 N | 20 19 E | 400 | S | sr, s |
| H30 | PL 41435 | 4 | 1981 | 49 24 N | 20 21 E | 500 | S | s |
| H31 | PL 41525 | 4 | 1977 | 49 24 N | 20 21 E | 500 | S | pm, sr, nb |
| H32 | PL 41526 | 4 | 1977 | 49 22 N | 20 11 E | 660 | S | l, pm, sr, nb |
| H33 | PL 41527 | 4 | 1977 | 49 35 N | 20 28 E | 500 | S | pm, sr, nb |
| H34 | PL 41528 | 4 | 1977 | 49 29 N | 22 04 E | 440 | SE | l, pm, sr, nb |
| H35 | PL 41529 | 4 | 1977 | 49 19 N | 22 41 E | 500 | SE | pm, sr, nb |
| H36 | PL 41530 | 4 | 1977 | 49 36 N | 21 48 E | 540 | SE | pm, sr, nb |
| H37 | PL 41531 | 4 | 1977 | 49 27 N | 22 20 E | 500 | SE | pm, sr, nb |
| H38 | PL 41532 | 4 | 1977 | 49 21 N | 22 17 E | 500 | SE | l, pm, sr, nb |
| H39 | PL 41533 | 4 | 1978 | 49 32 N | 22 06 E | 400 | SE | sr, nb, s |
| H40 | PL 41730 | 4 | 1977 | 49 20 N | 20 06 E | 900 | S | pm, sr, nb |
| H41 | PL 41731 | 4 | 1977 | 49 23 N | 20 08 E | 800 | S | pm, sr, nb |
| H42 | PL 41732 | 4 | 1977 | 49 30 N | 20 14 E | 900 | S | sr, nb, s |
| H43 | PL 41768 | 4 | 1977 | 49 24 N | 20 21 E | 500 | S | l, pm, sr, nb |
| H44 | PL 41867 | 2 | 1978 | 49 33 N | 20 33 E | 400 | S | sr, nb, s |
| H45 | PL 41868 | 1 | 1978 | 49 39 N | 20 36 E | 500 | S | sr, nb, s |
| H46 | PL 41869 | 6 | 1977 | 49 29 N | 22 04 E | 440 | SE | pm, sr, nb |
| H47 | PL 41871 | 1 | 1978 | 49 32 N | 22 06 E | 400 | SE | sr, nb, s |
| H48 | PL 42017 | 3 | 1977 | 49 26 N | 20 44 E | 505 | S | l, pm, sr, nb |
| H49 | PL 42120 | 4 | 1977 | 49 33 N | 20 33 E | 360 | S | l, pm, sr, nb |
| H50 | PL 42121 | 4 | 1977 | 49 29 N | 22 04 E | 440 | SE | pm, sr, nb |
| H51 | PL 42122 | 1 | 1977 | 49 31 N | 22 09 E | 360 | SE | sr, nb |
| H52 | PL 42388 | 4 | 1984 | 51 50 N | 22 29 E | 155 | E | |
| H53 | PL 42389 | 5 | 1984 | 51 50 N | 22 29 E | 155 | E | |
| H54 | PL 42768 | 3 | 1986 | 50 39 N | 22 18 E | 195 | SE | |
| H55 | PL 42769 | 3 | 1986 | 50 39 N | 23 22 E | 220 | SE | |
| H56 | PL 42770 | 3 | 1986 | 50 52 N | 23 04 E | 210 | SE | |
| H57 | PL 42771 | 3 | 1986 | 50 51 N | 23 27 E | 280 | SE | |
| H58 | PL 43051 | 4 | 1988 | na | na | na | na | sr |
| H59 | PL 43052 | 4 | 1988 | na | na | na | na | sr |
| H60 | PL 43053 | 4 | 1988 | na | na | na | na | sr |
| H61 | PL 43054 | 4 | 1988 | na | na | na | na | |
| H62 | PL 43055 | 4 | 1988 | na | na | na | na | |
| H63 | PL 43075 | 4 | 1988 | na | na | na | na | |
| H64 | PL 43357 | 4 | 1988 | na | na | na | na | sr |
| Trait | Unit | % of Evaluated Accessions | Mean | Min. | Max. | Variation | |
|---|---|---|---|---|---|---|---|
| Phenological | days to heading | days | 98.4% | 71.6 | 52.0 (PL 42 389) | 82.0 (PL 41267) | 9% |
| grain filling duration | days | 98.4% | 36.1 | 26.3 (PL 42769) | 44.0 (PL 40554) | 11% | |
| days to maturity | days | 98.4% | 108.1 | 81.7 (PL 42769) | 117.0 (PL 41532) | 6% | |
| Metrical | plant height | cm | 98.4% | 85.4 | 69.0 (PL 42017) | 109.0 (PL 42389) | 10% |
| ear length | cm | 60.9% | 9.2 | 7.0 (PL 42389) | 11.0 (PL 41365) | 11% | |
| number of grains per ear | no. | 46.9% | 11.6 | 7.7 (PL 40553) | 19.3 (PL 41279) | 21% | |
| thousand grain weight | g | 98.4% | 46.2 | 36.0 (PL 41270) | 55.2 (PL 41730) | 10% | |
| Bonitations | lodging resistance | scale 1 | 98.4% | 4.3 | 1.0 (17 access.) | 9.0 (6 access.) | 65% |
| powdery mildew resistance | scale 1 | 98.4% | 6.1 | 2.0 (2 access.) | 9.0 (16 access.) | 35% | |
| net blotch resistance | scale 1 | 90.6% | 8.7 | 6.0 (2 access.) | 9.0 (47 access.) | 8% | |
| stem rust resistance | scale 1 | 79.7% | 8.5 | 3.0 (1 access.) | 9.0 (46 access.) | 18% | |
| skald resistance | scale 1 | 50.0% | 8.9 | 7.0 (1 access.) | 9.0 (31 access.) | 4% |
| Primer | Sequence | Dye | No. Fragments | % Polymorphic Fragments | PIC |
|---|---|---|---|---|---|
| UBC 807 | (AG)8T | 6-FAM | 81 | 43% | 0.145 |
| UBC 825 | (AC)8T | 86 | 62% | 0.231 | |
| UBC 834 | (AG)8YT | VIC | 81 | 36% | 0.106 |
| UBC 841 | (GA)8YC | 107 | 60% | 0.191 | |
| UBC 856 | (AC)8YA | NED | 61 | 62% | 0.219 |
| UBC 857 | (AC)8YG | 102 | 76% | 0.247 | |
| UBC 884 | HBH(AG)7 | PET | 85 | 55% | 0.192 |
| UBC 885 | BHB(GA)7 | 90 | 51% | 0.189 |
| Parameter | Unit | Mean | Min. | Max. | Variation |
|---|---|---|---|---|---|
| Surface area | mm2 | 18.28 | 9.53 (PL 42122) | 23.33 (PL 41533) | 14% |
| Perimeter | mm | 23.22 | 16.42 (PL 42122) | 30.28 (PL 41533) | 9% |
| Length | mm | 7.45 | 4.75 (PL 42122) | 9.48 (PL 41533) | 10% |
| Width | mm | 3.03 | 2.17 (PL 42122) | 3.42 (PL 41279) | 7% |
| Color (Red) | 8-bit/channel | 167.71 | 131.05 (PL 42122) | 181.75 (PL 41279) | 6% |
| Color (Green) | 8-bit/channel | 141.04 | 101.14 (PL 42122) | 156.9 (PL 41270) | 8% |
| Color (Blue) | 8-bit/channel | 114.55 | 80.04 (PL 42122) | 131.81 (PL 41270) | 9% |
| ΦPT | p | |
|---|---|---|
| Acquisition Year | 0.06 | 0.001 |
| Region | 0.00 | ns |
| Altitude | 0.00 | ns |
| Precipitation | 0.01 | ns |
| Temperature | 0.03 | 0.015 |
| Botanical variety | 0.17 | 0.001 |
| Ear type | 0.02 | ns |
| Grain covering | 0.204 | 0.001 |
| Source | DF | Sum of Squares | Mean Squares | F | Pr > F |
|---|---|---|---|---|---|
| Residuals after scaling | 419 | 125.989 | 0.301 | ||
| Scaling | 1 | 83.931 | 83.931 | 279.130 | <0.0001 |
| Residuals after rotation | 420 | 209.920 | 0.500 | ||
| Rotation | 21 | 23.701 | 1.129 | 3.753 | <0.0001 |
| Residuals after translation | 441 | 233.621 | 0.530 | ||
| Translation | 7 | 0.000 | 0.000 | 0.000 | 1.000 |
| Corrected Total | 448 | 233.621 | 0.521 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziurdziak, J.; Bolc, P.; Wlodarczyk, S.; Puchta, M.; Gryziak, G.; Podyma, W.; Boczkowska, M. Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century. Agronomy 2020, 10, 1958. https://doi.org/10.3390/agronomy10121958
Dziurdziak J, Bolc P, Wlodarczyk S, Puchta M, Gryziak G, Podyma W, Boczkowska M. Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century. Agronomy. 2020; 10(12):1958. https://doi.org/10.3390/agronomy10121958
Chicago/Turabian StyleDziurdziak, Joanna, Paulina Bolc, Sylwia Wlodarczyk, Marta Puchta, Grzegorz Gryziak, Wiesław Podyma, and Maja Boczkowska. 2020. "Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century" Agronomy 10, no. 12: 1958. https://doi.org/10.3390/agronomy10121958
APA StyleDziurdziak, J., Bolc, P., Wlodarczyk, S., Puchta, M., Gryziak, G., Podyma, W., & Boczkowska, M. (2020). Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century. Agronomy, 10(12), 1958. https://doi.org/10.3390/agronomy10121958