
Rendered-Protein Hydrolysates as a Low-Cost Nitrogen Source for the Fungal Biotransformation of 5-Hydroxymethylfurfural


 , ,
, ,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Pre-Treatment and Partially Defatted Fines (PDF) Characterization
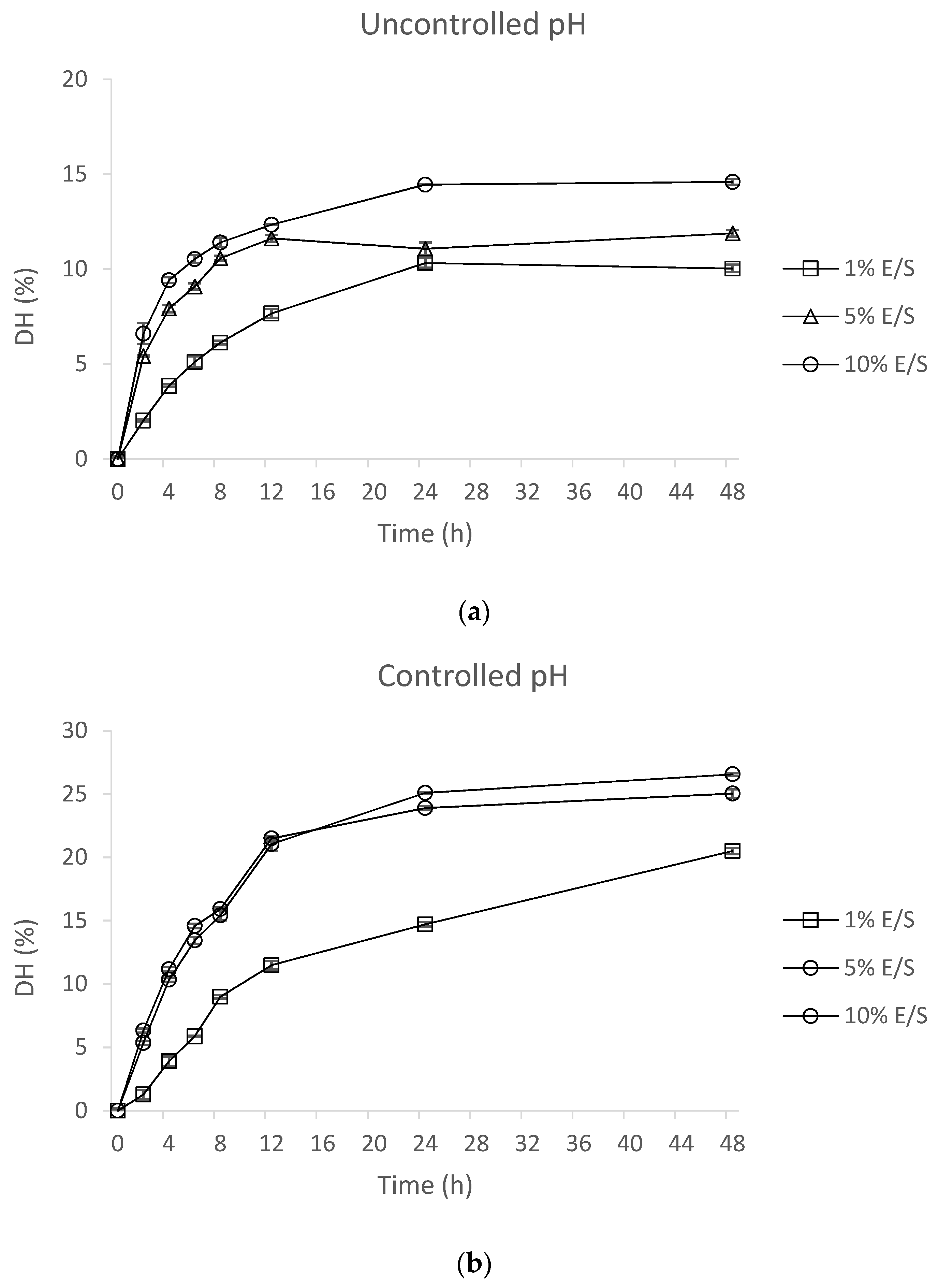
2.2. Optimization of Hydrolysis with Alcalase 2.4 L
2.3. Optimization of Hydrolysis with Neutrase 0.8 L
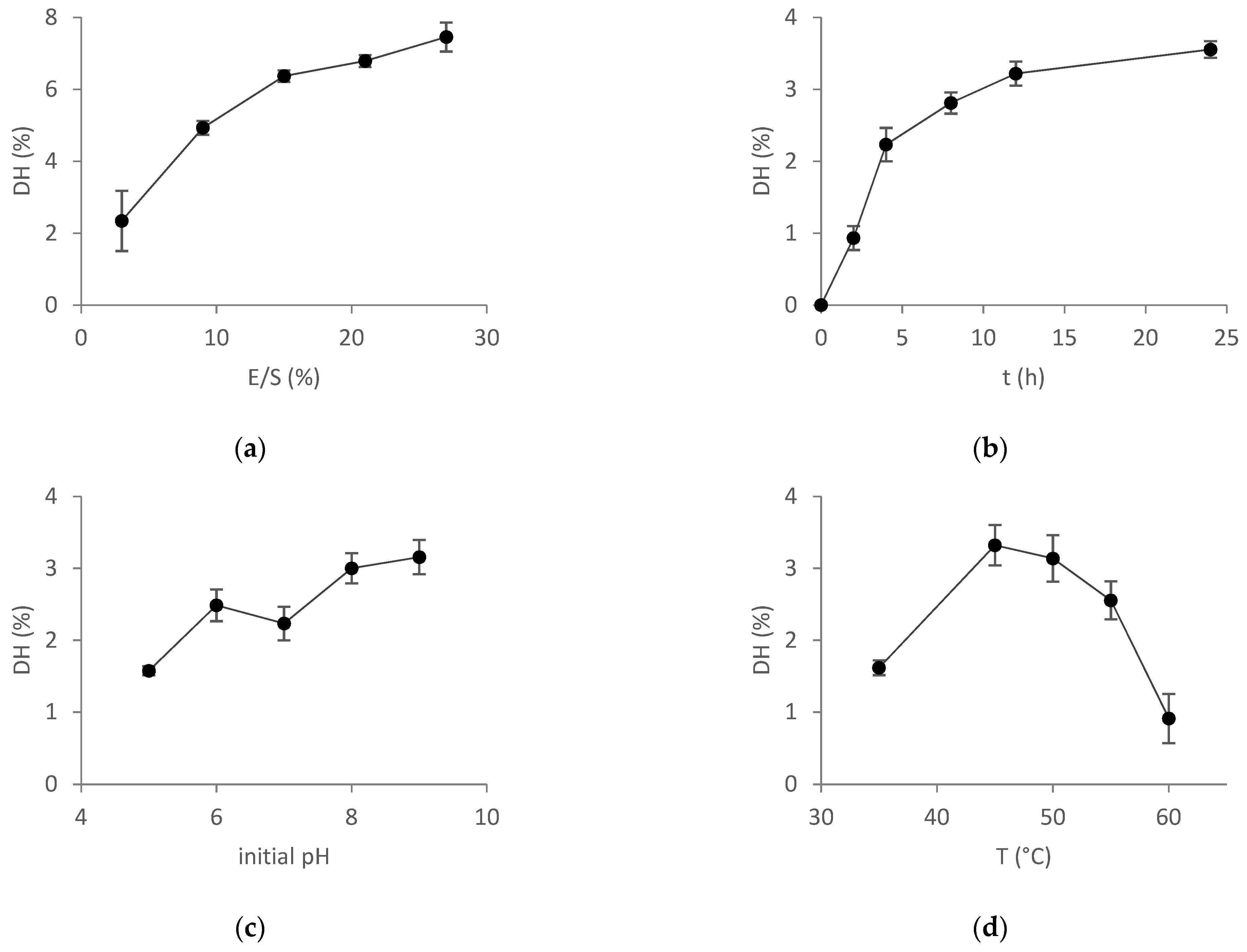
2.3.1. OFAT Experiments
2.3.2. RSM Optimization of Hydrolysis Parameters
2.3.3. Effect of Parameters
2.3.4. Optimum Conditions
2.3.5. Validation of the Predictive Model
2.4. Characterization of the Protein Hydrolysates
2.4.1. Macrocomponents and Elementary Analysis
2.4.2. Amino Acid Composition
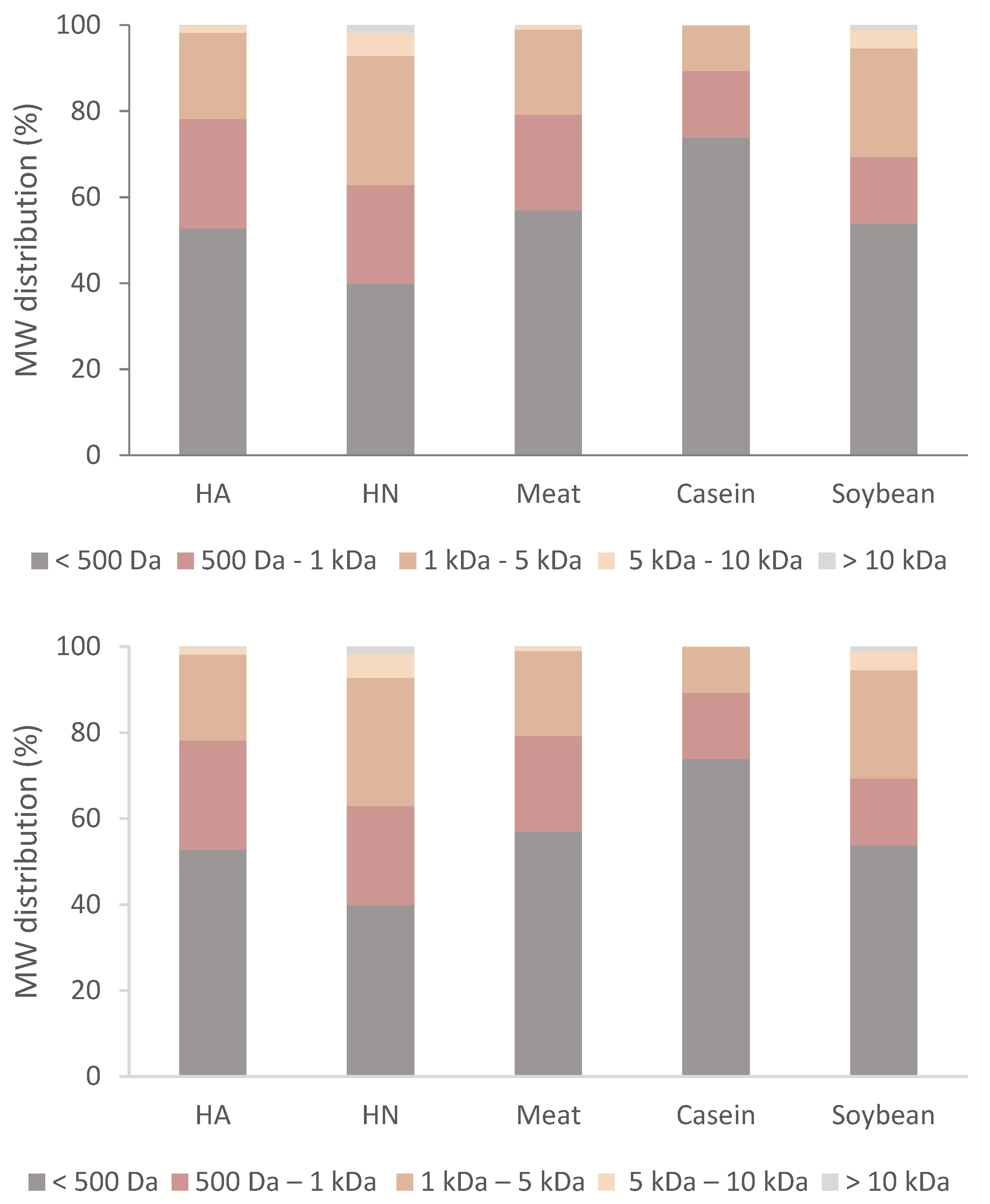
2.5. Size Exclusion Chromatography (SEC)
2.6. Biotransformations with F. striatum
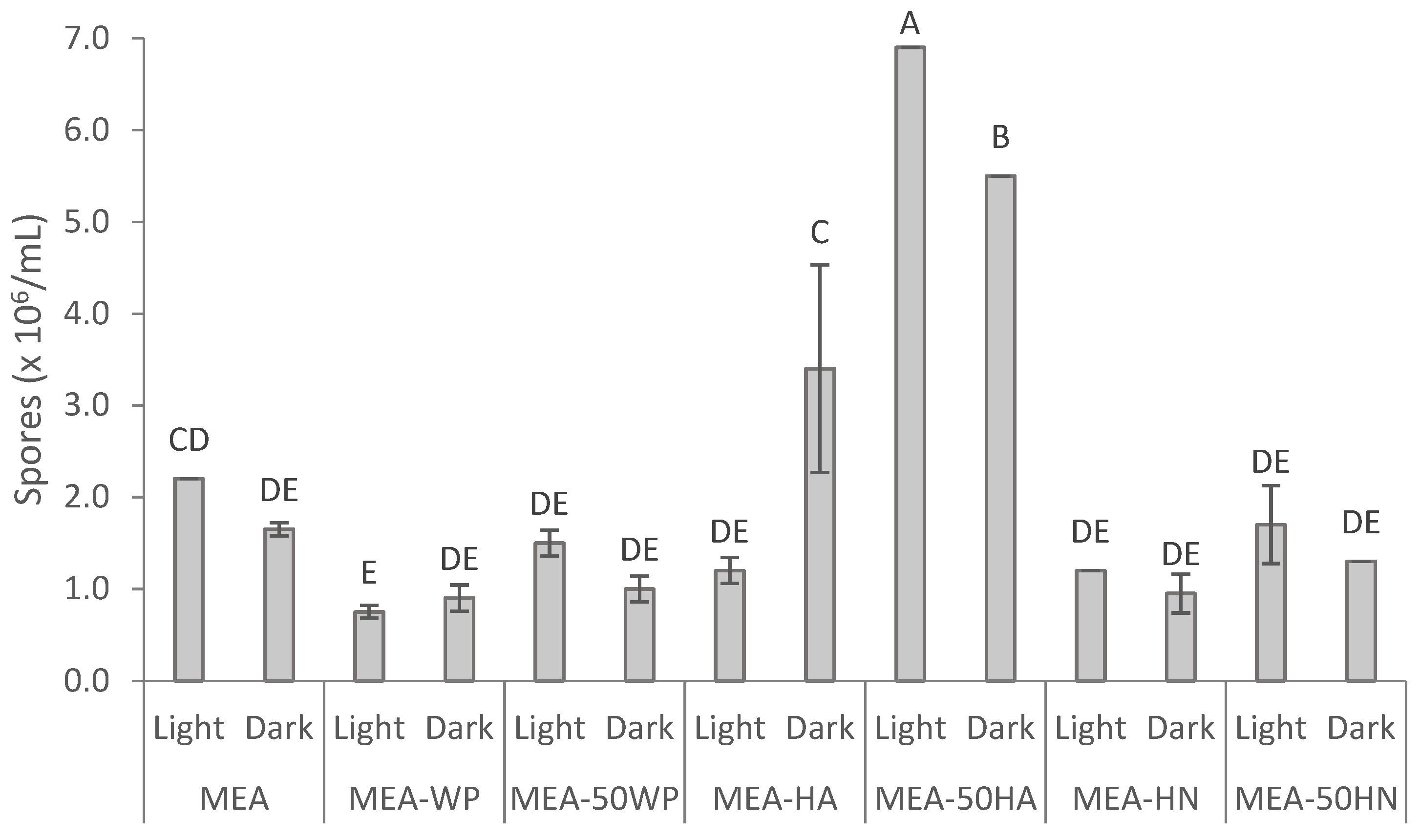
2.6.1. Effect of Nitrogen Source on F. striatum Sporulation
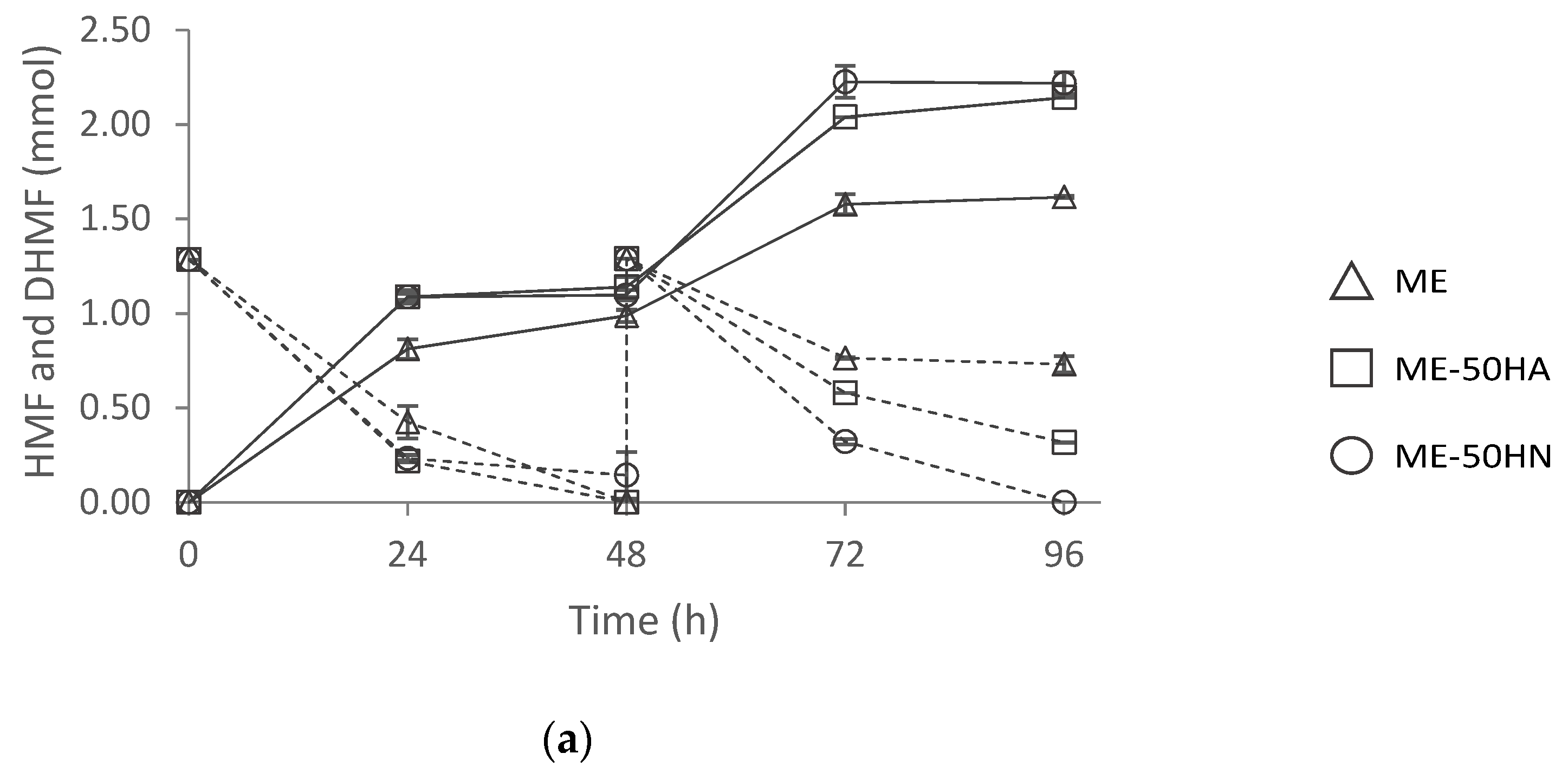
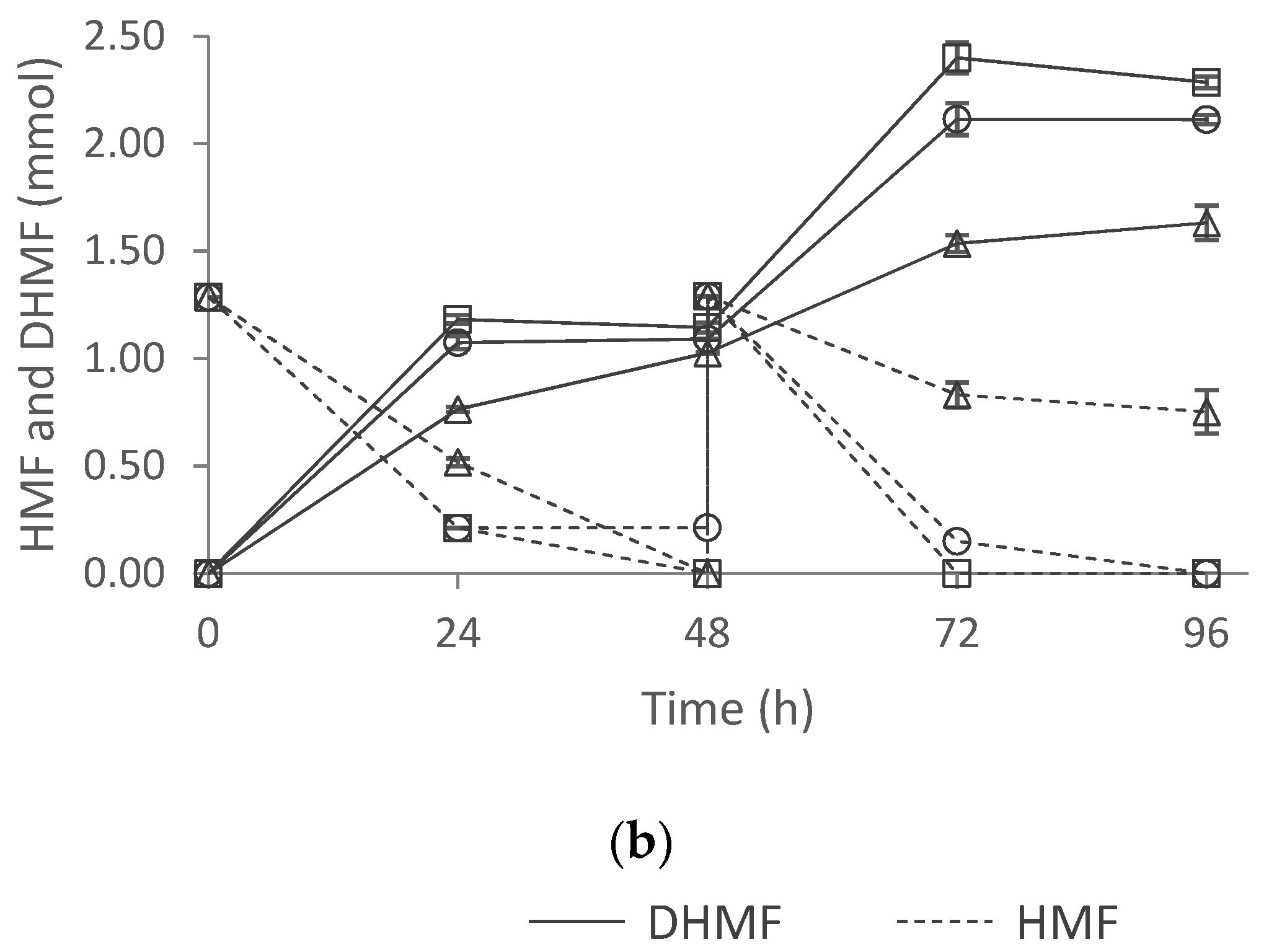
2.6.2. Effect of Inoculum Type in HMF Biotransformation
2.6.3. Effect of Nitrogen Source in HMF Biotransformation
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Raw Material and Protein Hydrolysates Preparation. Sample Pre-Treatment
3.2.2. Enzymatic Hydrolysis Optimization for Alcalase 2.4 L
3.2.3. Enzymatic Hydrolysis Optimization for Neutrase 0.8 L
3.2.4. Protein Hydrolysates Production in Shake Flasks
3.2.5. Microorganisms, Media, and Inoculum Preparation
3.2.6. Nitrogen Dose and Source Effect on F. striatum Sporulation and HMF Biotransformation
- (a)
- Substrate feeding with 75 mM HMF
- (b)
- Substrate feeding with 45 mM HMF.
3.2.7. Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, L.; Lin, L.; Wu, Z.; Zhou, S.; Liu, S. Recent Advances in Catalytic Transformation of Biomass-Derived 5-Hydroxymethylfurfural into the Innovative Fuels and Chemicals. Renew. Sustain. Energy Rev. 2017, 74, 230–257. [Google Scholar] [CrossRef]
- Lalanne, L.; Nyanhongo, G.S.; Guebitz, G.M.; Pellis, A. Biotechnological Production and High Potential of Furan-Based Renewable Monomers and Polymers. Biotechnol. Adv. 2021, 107707. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Xu, J.; Zhou, S.; He, A.; Tang, X.; Lin, L.; Xu, J.; Zhao, Y. Catalytic Advances in the Production and Application of Biomass-Derived 2,5-Dihydroxymethylfuran. ACS Catal. 2018, 8, 2959–2980. [Google Scholar] [CrossRef]
- Millán, A.; Sala, N.; Torres, M.; Canela-Garayoa, R. Biocatalytic Transformation of 5-Hydroxymethylfurfural into 2,5-Di(Hydroxymethyl)Furan by a Newly Isolated Fusarium striatum Strain. Catalysts 2021, 11, 216. [Google Scholar] [CrossRef]
- Domínguez de María, P.; Guajardo, N. Biocatalytic Valorization of Furans: Opportunities for Inherently Unstable Substrates. ChemSusChem 2017, 10, 4123–4134. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; He, X.; Li, B.; Pan, X. Improved Bio-Synthesis of 2,5-Bis(Hydroxymethyl)Furan by Burkholderia Contaminans NJPI-15 with Co-Substrate. Front. Chem. 2021, 9, 20. [Google Scholar] [CrossRef]
- Chen, D.; Cang, R.; Zhang, Z.D.; Huang, H.; Zhang, Z.G.; Ji, X.J. Efficient Reduction of 5-Hydroxymethylfurfural to 2, 5-Bis (Hydroxymethyl) Furan by a Fungal Whole-Cell Biocatalyst. Mol. Catal. 2021, 500, 111341. [Google Scholar] [CrossRef]
- He, Y.C.; Jiang, C.X.; Chong, G.G.; Di, J.H.; Ma, C.L. Biological Synthesis of 2,5-Bis(Hydroxymethyl)Furan from Biomass-Derived 5-Hydroxymethylfurfural by E. Coli CCZU-K14 Whole Cells. Bioresour. Technol. 2018, 247, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wu, S.; Yao, D.; Liu, L.; Zhang, L.; Yao, Z.; Pan, Y.; Chang, S.; Li, B. Efficient Biotransformation of 5-Hydroxymethylfurfural to 5-Hydroxymethyl-2-Furancarboxylic Acid by a New Whole-Cell Biocatalyst Pseudomonas aeruginosa PC-1. React. Chem. Eng. 2020, 5, 1397. [Google Scholar] [CrossRef]
- Paukner, R.; Staudigl, P.; Choosri, W.; Haltrich, D.; Leitner, C. Expression, Purification, and Characterization of Galactose Oxidase of Fusarium sambucinum in E. coli. Protein Expr. Purif. 2015, 108, 73–79. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, Z.H.; Cheng, A.D.; Xing, X.P.; Zong, M.H.; Bai, Y.P.; Li, N. Improved Synthesis of 2,5-Bis(Hydroxymethyl)Furan from 5-Hydroxymethylfurfural Using Acclimatized Whole Cells Entrapped in Calcium Alginate. Bioresour. Technol. 2018, 262, 177–183. [Google Scholar] [CrossRef]
- Yuan, H.; Li, J.; Shin, H.-D.; Du, G.; Chen, J.; Shi, Z.; Liu, L. Improved Production of 2,5-Furandicarboxylic Acid by Overexpression of 5-Hydroxymethylfurfural Oxidase and 5-Hydroxymethylfurfural/Furfural Oxidoreductase in Raoultella ornithinolytica BF60. Bioresour. Technol. 2018, 247, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Zhang, X.Y.; Li, N.; Xu, P.; Lou, W.Y.; Zong, M.H. Biocatalytic Reduction of HMF to 2,5-Bis(Hydroxymethyl)Furan by HMF-Tolerant Whole Cells. ChemSusChem 2017, 10, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Slininger, P.J.; Dien, B.S.; Berhow, M.A.; Kurtzman, C.P.; Gorsich, S.W. Adaptive Response of Yeasts to Furfural and 5-Hydroxymethylfurfural and New Chemical Evidence for HMF Conversion to 2,5-Bis-Hydroxymethylfuran. J. Ind. Microbiol. Biotechnol. 2004, 31, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Ra, C.H.; Jeong, G.T.; Shin, M.K.; Kim, S.K. Biotransformation of 5-Hydroxymethylfurfural (HMF) by Scheffersomyces stipitis during Ethanol Fermentation of Hydrolysate of the Seaweed Gelidium amansii. Bioresour. Technol. 2013, 140, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Kowbel, D.J.; Glass, N.L.; Yarden, O.; Hadar, Y. Detoxification of 5-Hydroxymethylfurfural by the Pleurotus Ostreatus Lignolytic Enzymes Aryl Alcohol Oxidase and Dehydrogenase. Biotechnol. Biofuels 2015, 8, 63. [Google Scholar] [CrossRef]
- Ran, H.; Zhang, J.; Gao, Q.; Lin, Z.; Bao, J. Analysis of Biodegradation Performance of Furfural and 5-Hydroxymethylfurfural by Amorphotheca resinae ZN1. Biotechnol. Biofuels 2014, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Millán Acosta, A.; Cosovanu, D.; Cabañeros López, P.; Tjalfe Thomsen, S.; Gernaey, K.V.; Canela-Garayoa, R. Co-Cultivation of a Novel Fusarium striatum Strain and a Xylose Consuming Saccharomyces cerevisiae Yields an Efficient Process for Simultaneous Detoxification and Fermentation of Lignocellulosic Hydrolysates. Chem. Eng. J. 2021, 426, 131575. [Google Scholar] [CrossRef]
- Parrado, J.; Rodriguez-Morgado, B.; Tejada, M.; Hernandez, T.; Garcia, C. Proteomic Analysis of Enzyme Production by Bacillus Licheniformis Using Different Feather Wastes as the Sole Fermentation Media. Enzyme Microb. Technol. 2014, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kampen, W.H. Nutritional Requirements in Fermentation Processes. In Fermentation and Biochemical Engineering Handbook: Principles, Process Design, and Equipment; Vogel, H.C., Todaro, C.M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 37–57. ISBN 9781455730469. [Google Scholar]
- Godan, T.K.; Rajesh, R.O.; Loreni, P.C.; Kumar Rai, A.; Sahoo, D.; Pandey, A.; Binod, P. Biotransformation of 5-Hydroxymethylfurfural by Acinetobacter Oleivorans S27 for the Synthesis of Furan Derivatives. Bioresour. Technol. 2019, 282, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Hossain, G.S.; Yuan, H.; Li, J.; Shin, H.-D.; Wang, M.; Du, G.; Chen, J.; Liu, L. Metabolic Engineering of Raoultella ornithinolytica BF60 for Production of 2,5-Furandicarboxylic Acid from 5-Hydroxymethylfurfural. Appl. Environ. Microbiol. 2017, 83, e02312-16. [Google Scholar] [CrossRef] [PubMed]
- Koopman, F.; Wierckx, N.; de Winde, J.H.; Ruijssenaars, H.J. Efficient Whole-Cell Biotransformation of 5-(Hydroxymethyl)Furfural into FDCA, 2,5-Furandicarboxylic Acid. Bioresour. Technol. 2010, 101, 6291–6296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Zong, M.H.; Li, N. Whole-Cell Biocatalytic Selective Oxidation of 5-Hydroxymethylfurfural to 5-Hydroxymethyl-2-Furancarboxylic Acid. Green Chem. 2017, 19, 4544–4551. [Google Scholar] [CrossRef]
- Bhaskar, N.; Modi, V.K.; Govindaraju, K.; Radha, C.; Lalitha, R.G. Utilization of Meat Industry by Products: Protein Hydrolysate from Sheep Visceral Mass. Bioresour. Technol. 2007, 98, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Šližyte, R.; Daukšas, E.; Falch, E.; Storrø, I.; Rustad, T. Characteristics of Protein Fractions Generated from Hydrolysed Cod (Gadus morhua) by-Products. Process Biochem. 2005, 40, 2021–2033. [Google Scholar] [CrossRef]
- Šližyte, R.; Daukšas, E.; Falch, E.; Storrø, I.; Rustad, T. Yield and Composition of Different Fractions Obtained after Enzymatic Hydrolysis of Cod (Gadus morhua) by-Products. Process Biochem. 2005, 40, 1415–1424. [Google Scholar] [CrossRef]
- Nilsang, S.; Lertsiri, S.; Suphantharika, M.; Assavanig, A. Optimization of Enzymatic Hydrolysis of Fish Soluble Concentrate by Commercial Proteases. J. Food Eng. 2005, 70, 571–578. [Google Scholar] [CrossRef]
- Benjakul, S.; Morrissey, M.T. Protein Hydrolysates from Pacific Whiting Solid Wastes. J. Agric. Food Chem. 1997, 45, 3423–3430. [Google Scholar] [CrossRef]
- Ovissipour, M.; Rasco, B.; Shiroodi, S.G.; Modanlow, M.; Gholami, S.; Nemati, M. Antioxidant Activity of Protein Hydrolysates from Whole Anchovy Sprat (Clupeonella engrauliformis) Prepared Using Endogenous Enzymes and Commercial Proteases. J. Sci. Food Agric. 2013, 93, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, N.C.; Pessato, T.B.; Fernandes, L.G.R.; de Lima Zollner, R.; Netto, F.M. Physicochemical Characteristics and Antigenicity of Whey Protein Hydrolysates Obtained with and without PH Control. Int. Dairy J. 2017, 71, 24–34. [Google Scholar] [CrossRef]
- Sousa, R.; Lopes, G.P.; Pinto, G.A.; Almeida, P.I.F.; Giordano, R.C. GMC-Fuzzy Control of PH during Enzymatic Hydrolysis of Cheese Whey Proteins. Comput. Chem. Eng. 2004, 28, 1661–1672. [Google Scholar] [CrossRef]
- Guérard, F.; Dufossé, L.; De La Broise, D.; Binet, A. Enzymatic Hydrolysis of Proteins from Yellowfin Tuna (Thunnus albacares) Wastes Using Alcalase. J. Mol. Catal. B Enzym. 2001, 11, 1051–1059. [Google Scholar] [CrossRef]
- Qi, W.; He, Z. Enzymatic Hydrolysis of Protein: Mechanism and Kinetic Model. Front. Chem. China 2006, 1, 308–314. [Google Scholar] [CrossRef]
- Valencia, P.; Pinto, M.; Almonacid, S. Identification of the Key Mechanisms Involved in the Hydrolysis of Fish Protein by Alcalase. Process Biochem. 2014, 49, 258–264. [Google Scholar] [CrossRef]
- Lapeña, D.; Vuoristo, K.S.; Kosa, G.; Horn, S.J.; Eijsink, V.G.H. Comparative Assessment of Enzymatic Hydrolysis for Valorization of Different Protein-Rich Industrial Byproducts. J. Agric. Food Chem. 2018, 66, 9738–9749. [Google Scholar] [CrossRef]
- Noman, A.; Xu, Y.; AL-Bukhaiti, W.Q.; Abed, S.M.; Ali, A.H.; Ramadhan, A.H.; Xia, W. Influence of Enzymatic Hydrolysis Conditions on the Degree of Hydrolysis and Functional Properties of Protein Hydrolysate Obtained from Chinese Sturgeon (Acipenser sinensis) by Using Papain Enzyme. Process Biochem. 2018, 67, 19–28. [Google Scholar] [CrossRef]
- Liaset, B.; Lied, E.; Espe, M. Enzymatic Hydrolysis of By-Products from the Fish-Filleting Industry; Chemical Characterisation and Nutritional Evaluation. J. Sci. Food Agric. 2000, 80, 581–589. [Google Scholar] [CrossRef]
- Piazza, G.J.; Garcia, R.A. Proteolysis of Meat and Bone Meal to Increase Utilisation. Anim. Prod. Sci. 2014, 54, 200. [Google Scholar] [CrossRef]
- Webster, J.D.; Ledward, D.A.; Lawrie, R.A. Protein Hydrolysates from Meat Industry By-Products. Meat Sci. 1982, 7, 147–157. [Google Scholar] [CrossRef]
- Kurozawa, L.E.; Park, K.J.; Hubinger, M.D. Optimization of the Enzymatic Hydrolysis of Chicken Meat Using Response Surface Methodology. J. Food Sci. 2008, 73, C405–C412. [Google Scholar] [CrossRef]
- Bhaskar, N.; Benila, T.; Radha, C.; Lalitha, R.G. Optimization of Enzymatic Hydrolysis of Visceral Waste Proteins of Catla (Catla catla) for Preparing Protein Hydrolysate Using a Commercial Protease. Bioresour. Technol. 2008, 99, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, N.; Mahendrakar, N.S. Protein Hydrolysate from Visceral Waste Proteins of Catla (Catla catla): Optimization of Hydrolysis Conditions for a Commercial Neutral Protease. Bioresour. Technol. 2008, 99, 4105–4111. [Google Scholar] [CrossRef] [PubMed]
- de Queiroz, A.L.M.; Bezerra, T.K.A.; de Freitas Pereira, S.; da Silva, M.E.C.; de Almeida Gadelha, C.A.; Gadelha, T.S.; Pacheco, M.T.B.; Madruga, M.S. Functional Protein Hydrolysate from Goat By-Products: Optimization and Characterization Studies. Food Biosci. 2017, 20, 19–27. [Google Scholar] [CrossRef]
- Dey, S.S.; Dora, K.C. Optimization of the Production of Shrimp Waste Protein Hydrolysate Using Microbial Proteases Adopting Response Surface Methodology. J. Food Sci. Technol. 2014, 51, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Li, Z.G.; Gai, Q.Y.; Li, X.J.; Wei, F.Y.; Fu, Y.J.; Ma, W. Microwave-Assisted Aqueous Enzymatic Extraction of Oil from Pumpkin Seeds and Evaluation of Its Physicochemical Properties, Fatty Acid Compositions and Antioxidant Activities. Food Chem. 2014, 147, 17–24. [Google Scholar] [CrossRef]
- Yusoff, M.M.; Gordon, M.H.; Ezeh, O.; Niranjan, K. Aqueous Enzymatic Extraction of Moringa Oleifera Oil. Food Chem. 2016, 211, 400–408. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, B.; Chen, B.; Jing, L.; Zhu, Z.; Kazemi, K. Modeling and Optimization of Newfoundland Shrimp Waste Hydrolysis for Microbial Growth Using Response Surface Methodology and Artificial Neural Networks. Mar. Pollut. Bull. 2016, 109, 245–252. [Google Scholar] [CrossRef]
- Puri, S.; Beg, Q.K.; Gupta, R. Optimization of Alkaline Protease Production from Bacillus Sp. by Response Surface Methodology. Curr. Microbiol. 2002, 44, 286–290. [Google Scholar] [CrossRef]
- Montgomery, D.C. Design and Analysis of Experiments, 8th ed.; Montgomery, D.C., Ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Cheong, C.W.; Lee, Y.S.; Ahmad, S.A.; Ooi, P.T.; Phang, L.Y. Chicken Feather Valorization by Thermal Alkaline Pretreatment Followed by Enzymatic Hydrolysis for Protein-Rich Hydrolysate Production. Waste Manag. 2018, 79, 658–666. [Google Scholar] [CrossRef]
- Elmalimadi, M.B.; Jovanović, J.R.; Stefanović, A.B.; Tanasković, S.J.; Djurović, S.B.; Bugarski, B.M.; Knežević-Jugović, Z.D. Controlled Enzymatic Hydrolysis for Improved Exploitation of the Antioxidant Potential of Wheat Gluten. Ind. Crops Prod. 2017, 109, 548–557. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Rao, G.N.; Rao, D.G.; Jyothirmayi, T. Protein Hydrolysates from Meriga (Cirrhinus mrigala) Egg and Evaluation of Their Functional Properties. Food Chem. 2010, 120, 652–657. [Google Scholar] [CrossRef]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmenter, M. Influence of Hydrolysis Degree on the Functional Properties of Salmon Byproducts Hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional Properties of Tropical Banded Cricket (Gryllodes sigillatus) Protein Hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Kristinsson, H.G.; Rasco, B.A. Fish Protein Hydrolysates: Production, Biochemical, and Functional Properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.; Pyle, D.J.; Piazza, G.J.; Wen, Z. Hydrolysis of Animal Protein Meals for Improved Utility in Non-Feed Applications. Appl. Eng. Agric. 2011, 27, 269–276. [Google Scholar] [CrossRef]
- Deydier, E.; Guilet, R.; Sarda, S.; Sharrock, P. Physical and Chemical Characterisation of Crude Meat and Bone Meal Combustion Residue: “Waste or Raw Material?”. J. Hazard. Mater. 2005, 121, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Characteristics and Use of Yellow Stripe Trevally Hydrolysate as Culture Media. J. Food Sci. 2009, 74, S219–S225. [Google Scholar] [CrossRef]
- Lobo-Alfonso, J.; Price, P.; Jayme, D. Benefits and Limitations of Protein Hydrolysates as Components of Serum-Free Media for Animal Cell Culture Applications Protein Hydrolysates in Serum Free Media. In Protein Hydrolysates in Biotechnology; Springer: Dordrecht, The Netherlands, 2010; pp. 55–78. ISBN 9781402066733. [Google Scholar]
- Taskin, M. A New Strategy for Improved Glutathione Production from Saccharomyces Cerevisiae: Use of Cysteine- and Glycine-Rich Chicken Feather Protein Hydrolysate as a New Cheap Substrate. J. Sci. Food Agric. 2013, 93, 535–541. [Google Scholar] [CrossRef]
- Stanbury, P.F.; Whitaker, A.; Hall, S.J. Media for Industrial Fermentations. In Principles of Fermentation Technology; Stanbury, P.F., Whitaker, A., Hall, S.J., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; pp. 213–272. [Google Scholar]
- Colgrave, M.L.; Allingham, P.G.; Jones, A. Hydroxyproline Quantification for the Estimation of Collagen in Tissue Using Multiple Reaction Monitoring Mass Spectrometry. J. Chromatogr. A 2008, 1212, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wu, Z.; Jia, S.; Wu, G. Analysis of Amino Acid Composition in Proteins of Animal Tissues and Foods as Pre-Column o-Phthaldialdehyde Derivatives by HPLC with Fluorescence Detection. J. Chromatogr. B 2014, 964, 116–127. [Google Scholar] [CrossRef]
- Guo, S.; Duan, J.A.; Qian, D.; Tang, Y.; Qian, Y.; Wu, D.; Su, S.; Shang, E. Rapid Determination of Amino Acids in Fruits of Ziziphus Jujuba by Hydrophilic Interaction Ultra-High-Performance Liquid Chromatography Coupled with Triple-Quadrupole Mass Spectrometry. J. Agric. Food Chem. 2013, 61, 2709–2719. [Google Scholar] [CrossRef] [PubMed]
- Fountoulakis, M.; Lahm, H.W. Hydrolysis and Amino Acid Composition Analysis of Proteins. J. Chromatogr. A 1998, 826, 109–134. [Google Scholar] [CrossRef]
- Anzani, C.; Prandi, B.; Tedeschi, T.; Baldinelli, C.; Sorlini, G.; Wierenga, P.A.; Dossena, A.; Sforza, S. Degradation of Collagen Increases Nitrogen Solubilisation during Enzymatic Hydrolysis of Fleshing Meat. Waste Biomass Valorization 2018, 9, 1113–1119. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Li-Chan, E.C.Y. Enzymatic Production of Protein Hydrolysates from Steelhead (Oncorhynchus Mykiss) Skin Gelatin as Inhibitors of Dipeptidyl-Peptidase IV and Angiotensin-I Converting Enzyme. J. Funct. Foods 2017, 28, 254–264. [Google Scholar] [CrossRef]
- Chi, C.F.; Cao, Z.H.; Wang, B.; Hu, F.Y.; Li, Z.R.; Zhang, B. Antioxidant and Functional Properties of Collagen Hydrolysates from Spanish Mackerel Skin as Influenced by Average Molecular Weight. Molecules 2014, 19, 11211–11230. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.A.; You, S.G.; Kim, S.M. Molecular and Physical Characteristics of Squid (Todarodes Pacificus) Skin Collagens and Biological Properties of Their Enzymatic Hydrolysates. J. Food Sci. 2008, 73, C249–C255. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, C.G.; Comesaña, M.B.; Ariza, P.R.; Pérez-Martín, R.I. Characterization of Collagen from Different Discarded Fish Species of the West Coast of the Iberian Peninsula. J. Aquat. Food Prod. Technol. 2016, 25, 388–399. [Google Scholar] [CrossRef]
- Gomez-Guillen, M.C.; Gimenez, B.; Lopez-Caballero, M.E.; Montero, M.P. Functional and Bioactive Properties of Collagen and Gelatin from Alternative Sources: A Review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Zhang, Y.; Olsen, K.; Grossi, A.; Otte, J. Effect of Pretreatment on Enzymatic Hydrolysis of Bovine Collagen and Formation of ACE-Inhibitory Peptides. Food Chem. 2013, 141, 2343–2354. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the Roots: A Reappraisal of Neocosmospora. Persoonia Mol. Phylogeny Evol. Fungi 2019, 43, 90–185. [Google Scholar] [CrossRef]
- Su, Y.-Y.; Qi, Y.-L.; Cai, L. Induction of Sporulation in Plant Pathogenic Fungi. Mycology 2012, 3, 195–200. [Google Scholar] [CrossRef]
- Sharma, G.; Pandey, R.R. Influence of Culture Media on Growth, Colony Character and Sporulation of Fungi Isolated from Decaying Vegetable Wastes. J. Yeast Fungal Res. 2010, 1, 157–164. [Google Scholar]
- Lazarotto, M.; Mezzomo, R.; Gonzatto, M.; Finger, G.; Brião Muniz, M.F. Mycelia Growth and Sporulation of Fusarium Chlamydosporum Species Complex under Different Culture Conditions. Amaz. J. Agric. Environ. Sci. 2014, 57, 35–40. [Google Scholar] [CrossRef]
- Li, S.; Myung, K.; Guse, D.; Donkin, B.; Proctor, R.H.; Grayburn, W.S.; Calvo, A.M. FvVE1 Regulates Filamentous Growth, the Ratio of Microconidia to Macroconidia and Cell Wall Formation in Fusarium verticillioides. Mol. Microbiol. 2006, 62, 1418–1432. [Google Scholar] [CrossRef] [PubMed]
- Chehri, K.; Salleh, B.; Zakaria, L. Morphological and Phylogenetic Analysis of Fusarium Solani Species Complex in Malaysia. Microb. Ecol. 2015, 69, 457–471. [Google Scholar] [CrossRef]
- Martins, C.; Hartmann, D.O.; Varela, A.; Coelho, J.A.S.; Lamosa, P.; Afonso, C.A.M.; Silva Pereira, C. Securing a Furan-based Biorefinery: Disclosing the Genetic Basis of the Degradation of Hydroxymethylfurfural and Its Derivatives in the Model Fungus Aspergillus nidulans. Microb. Biotechnol. 2020, 13, 1983–1996. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved Method for Determining Food Protein Degree of Hydrolysis. J. Food Sci. Food Chem. Toxicol. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Enzymic Hydrolysis of Food Proteins, 1st ed.; Adler-Nissen, J., Ed.; Elsevier Applied Science Publishers: Amsterdam, The Netherlands, 1986. [Google Scholar]
- Rutherfurd, S.M. Methodology for Determining Degree of Hydrolysis of Proteins in Hydrolysates: A Review. J. AOAC Int. 2010, 93, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, J.; Hansen, E.T.; Bredie, W.L.P.; Lametsch, R. Structural Characteristics of Low Bitter and High Umami Protein Hydrolysates Prepared from Bovine Muscle and Porcine Plasma. Food Chem. 2018, 257, 163–171. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
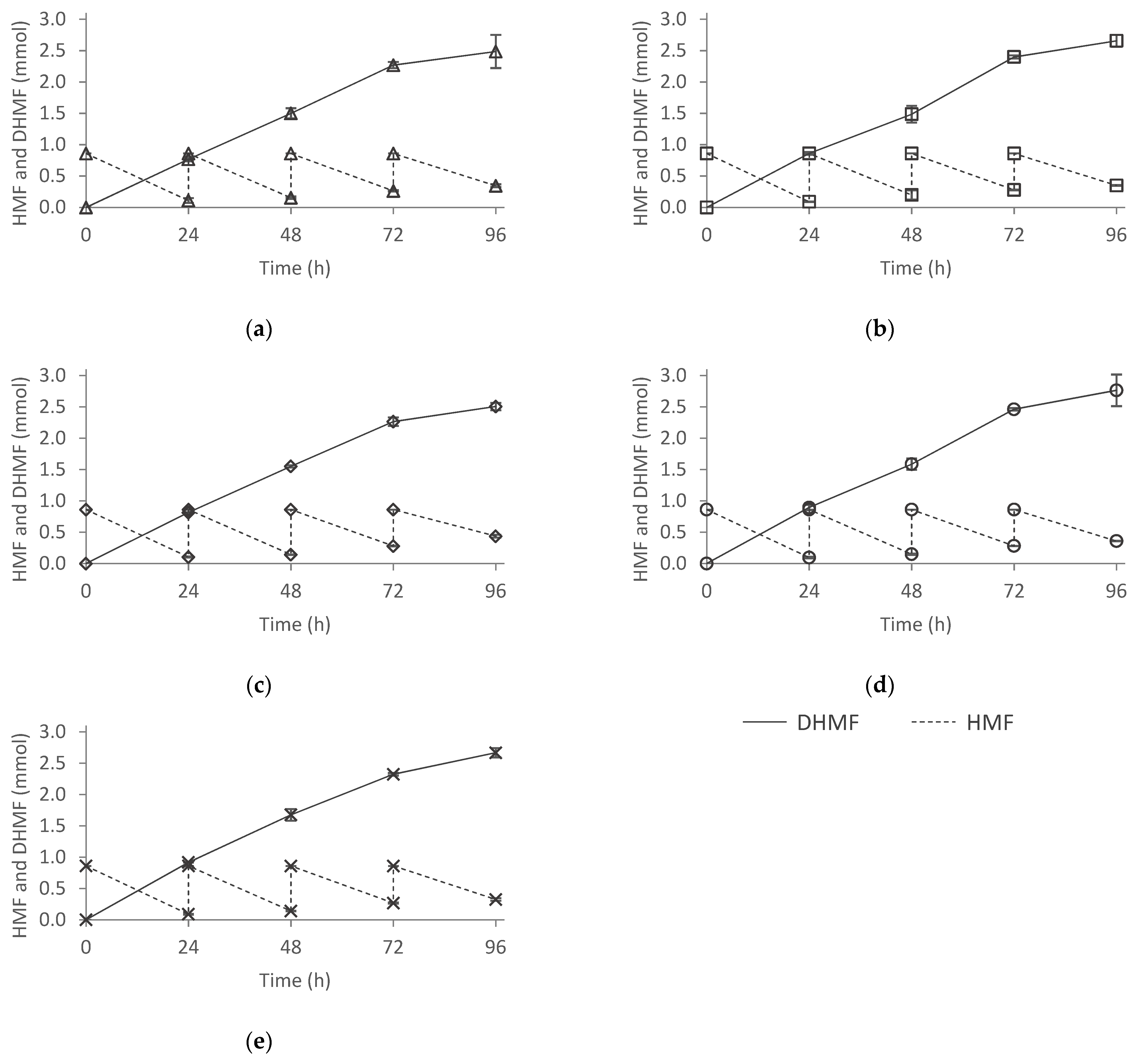{kind=link}
| Factors | Levels | ||||
|---|---|---|---|---|---|
| α (−1.48) | −1 | 0 | 1 | α (1.48) | |
| X1—E/S (%) | 2.59 | 5.0 | 10 | 15 | 17.41 |
| X2—Time (h) | 0.57 | 2.5 | 6.5 | 10.5 | 12.43 |
| X3—Initial pH | 5.52 | 6 | 7 | 8 | 8.48 |
| X4—T (°C) | 37.59 | 40 | 45 | 50 | 52.41 |
| Compounds | Culture Media | ||||||
|---|---|---|---|---|---|---|---|
| MEA | MEA-WP | MEA-50WP | MEA-HA | MEA-50HA | MEA-HN | MEA-50HN | |
| Malt extract | 20 | 20 | 10 | 20 | 10 | 20 | 10 |
| Glucose | 20 | 20 | 20 | 20 | 20 | 20 | 20 |
| Soybean peptone | 1 | - | - | - | - | - | - |
| HA | - | - | - | 1 | 11 | - | - |
| HN | - | - | - | - | - | 1 | 11 |
| Agar | 15 | 15 | 15 | 15 | 15 | 15 | 15 |
| Compound | Culture Medium | ||
|---|---|---|---|
| ME | Me-50HA | ME-50HN | |
| Malt extract | 20 | 10 | 10 |
| Glucose | 20 | 20 | 20 |
| Soybean peptone | 1 | - | - |
| HA | - | 11 | - |
| HN | - | - | 11 |
| Compound | Culture Medium | ||||
|---|---|---|---|---|---|
| ME | ME-50WP | ME-50Meat | ME-50HA | ME-50HN | |
| Malt extract | 20 | 10 | 10 | 10 | 10 |
| Glucose | 20 | 20 | 20 | 20 | 20 |
| Soybean peptone | 1 | - | - | - | - |
| Meat peptone | - | - | 11 | - | - |
| HA | - | - | - | 11 | - |
| HN | - | - | - | - | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cosovanu, D.; Millán Acosta, A.; Cabañeros López, P.; Gernaey, K.V.; Li, Q.; Lametsch, R.; Canela-Garayoa, R.; Eras, J.; Villorbina, G. Rendered-Protein Hydrolysates as a Low-Cost Nitrogen Source for the Fungal Biotransformation of 5-Hydroxymethylfurfural. Catalysts 2022, 12, 839. https://doi.org/10.3390/catal12080839
Cosovanu D, Millán Acosta A, Cabañeros López P, Gernaey KV, Li Q, Lametsch R, Canela-Garayoa R, Eras J, Villorbina G. Rendered-Protein Hydrolysates as a Low-Cost Nitrogen Source for the Fungal Biotransformation of 5-Hydroxymethylfurfural. Catalysts. 2022; 12(8):839. https://doi.org/10.3390/catal12080839
Chicago/Turabian StyleCosovanu, Diana, Alberto Millán Acosta, Pau Cabañeros López, Krist V. Gernaey, Qian Li, Rene Lametsch, Ramon Canela-Garayoa, Jordi Eras, and Gemma Villorbina. 2022. "Rendered-Protein Hydrolysates as a Low-Cost Nitrogen Source for the Fungal Biotransformation of 5-Hydroxymethylfurfural" Catalysts 12, no. 8: 839. https://doi.org/10.3390/catal12080839
APA StyleCosovanu, D., Millán Acosta, A., Cabañeros López, P., Gernaey, K. V., Li, Q., Lametsch, R., Canela-Garayoa, R., Eras, J., & Villorbina, G. (2022). Rendered-Protein Hydrolysates as a Low-Cost Nitrogen Source for the Fungal Biotransformation of 5-Hydroxymethylfurfural. Catalysts, 12(8), 839. https://doi.org/10.3390/catal12080839