
Exercise Suppresses Head and Neck Squamous Cell Carcinoma Growth via Oncostatin M
 , ,
, ,  , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
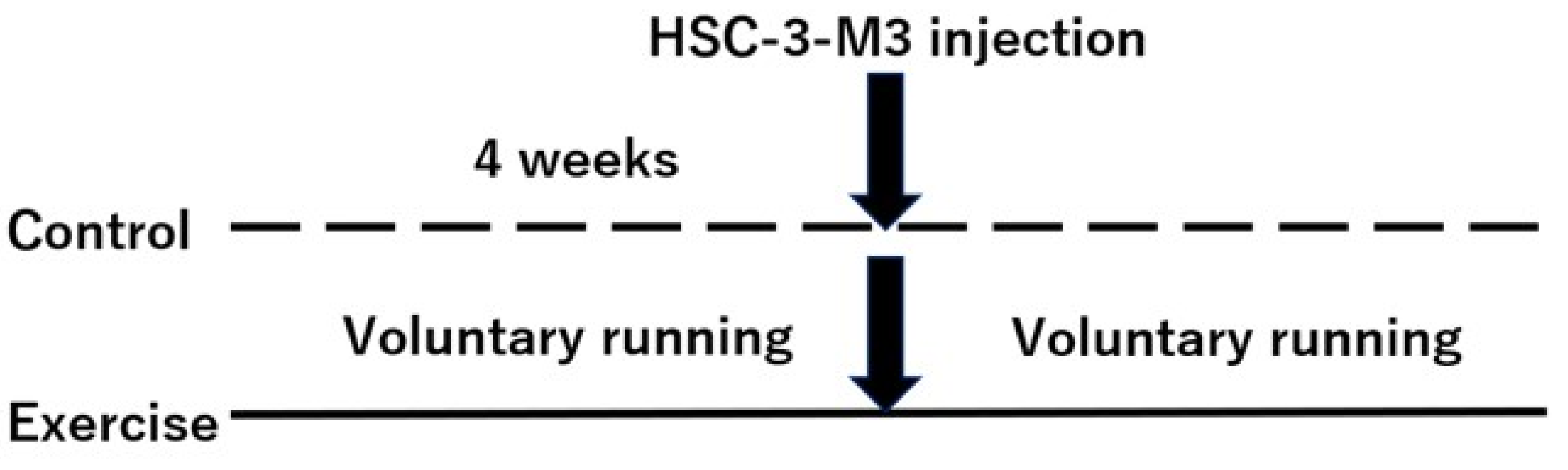
2.2. Mouse Models
2.3. ELISA
2.4. Cell Viability and Proliferation
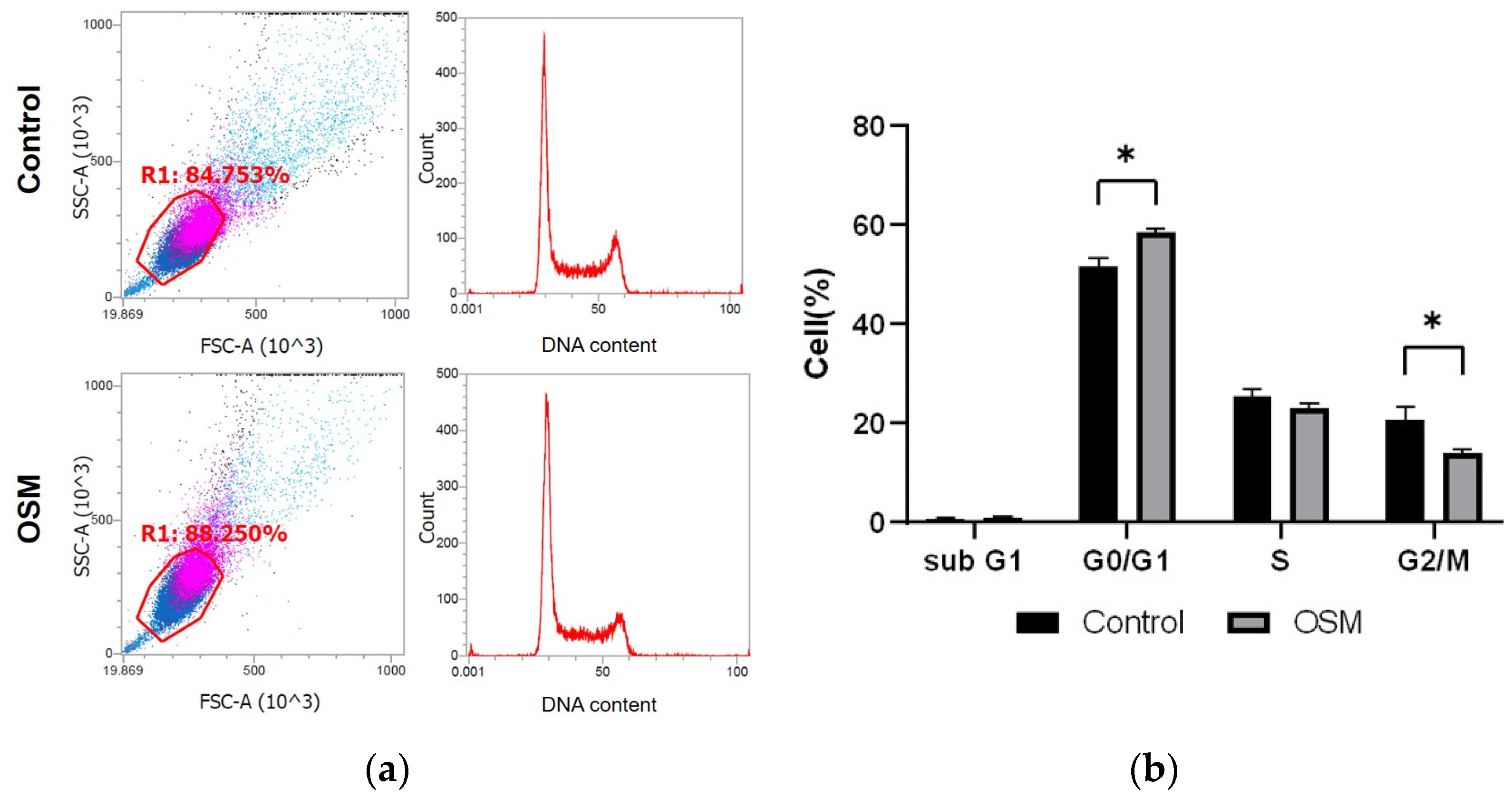
2.5. Cell Cycle Analysis
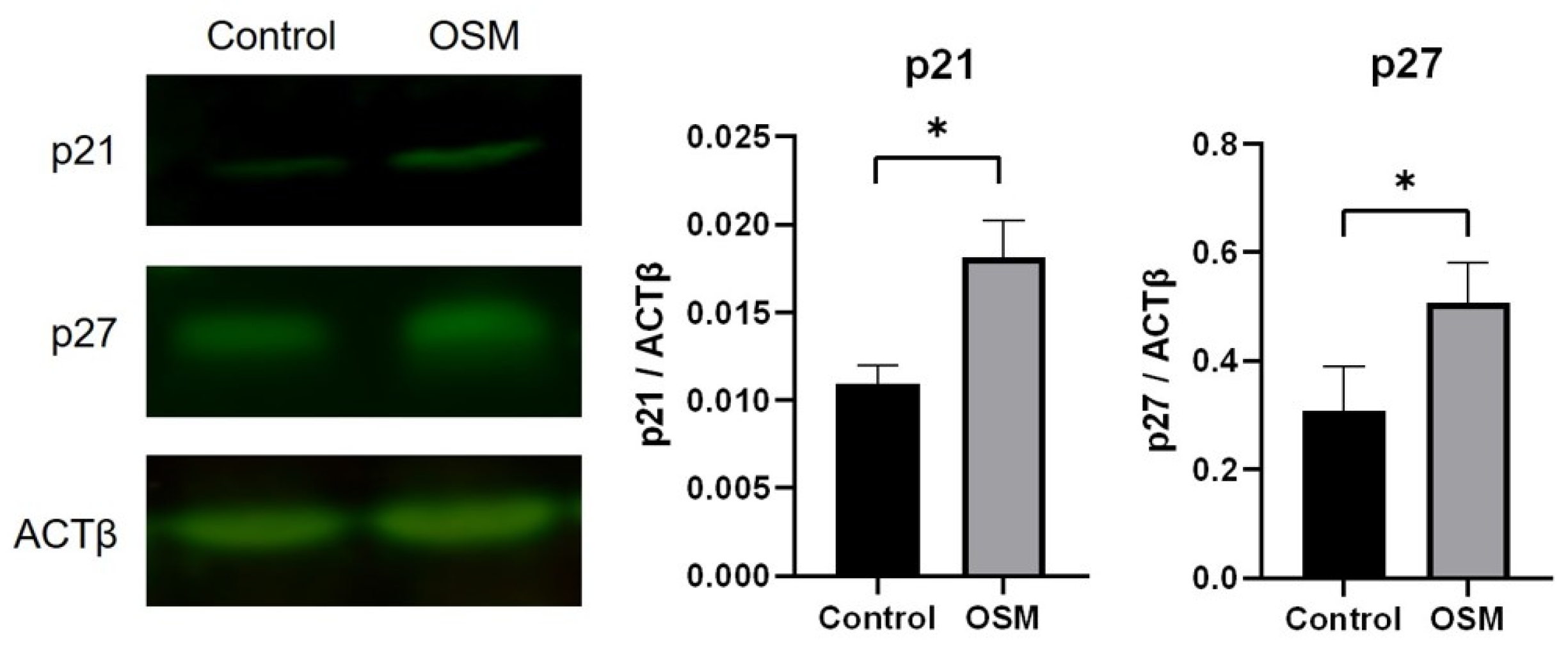
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
3.1. Effects of Exercise on HNSCC-Bearing Mice In Vivo
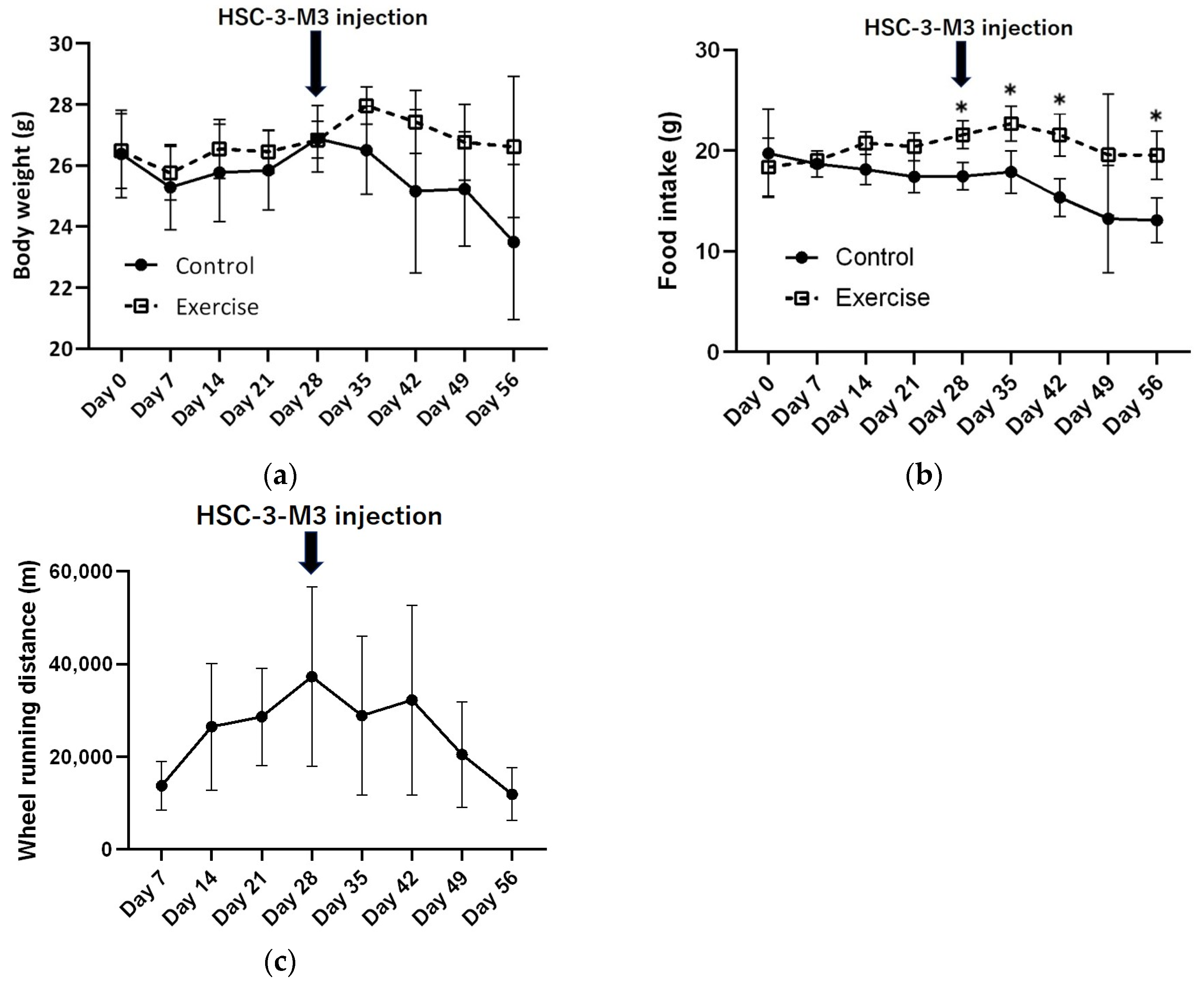
3.1.1. Changes in Body Weight, Food Intake, and Wheel Running Distance
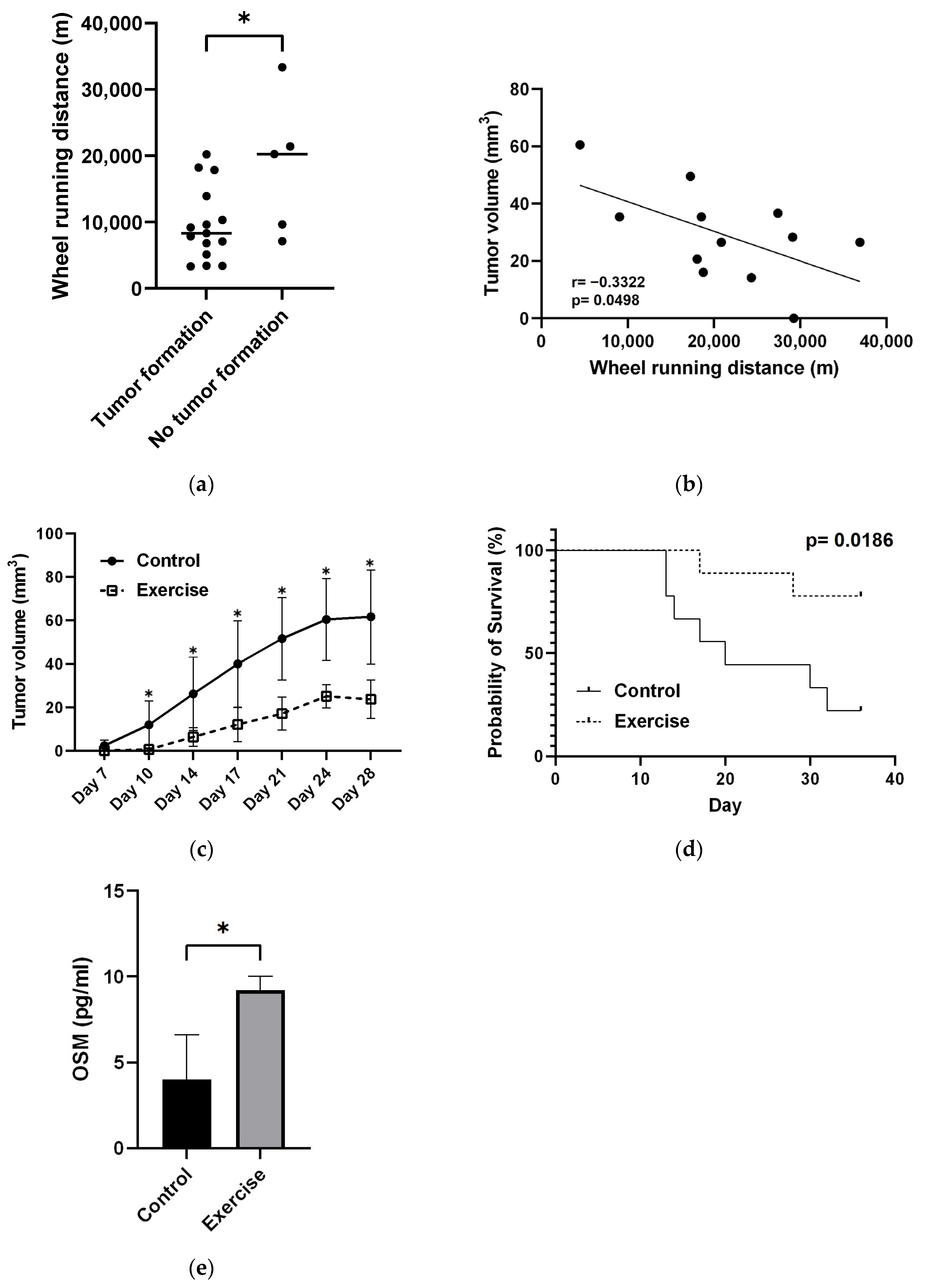
3.1.2. Changes in Wheel Running Distance and Tumor Volume
3.1.3. Exercise Significantly Suppressed Tumor Growth and Prolonged Survival
3.1.4. Exercise Increases OSM Levels
3.2. Direct Impact of OSM on HNSCC
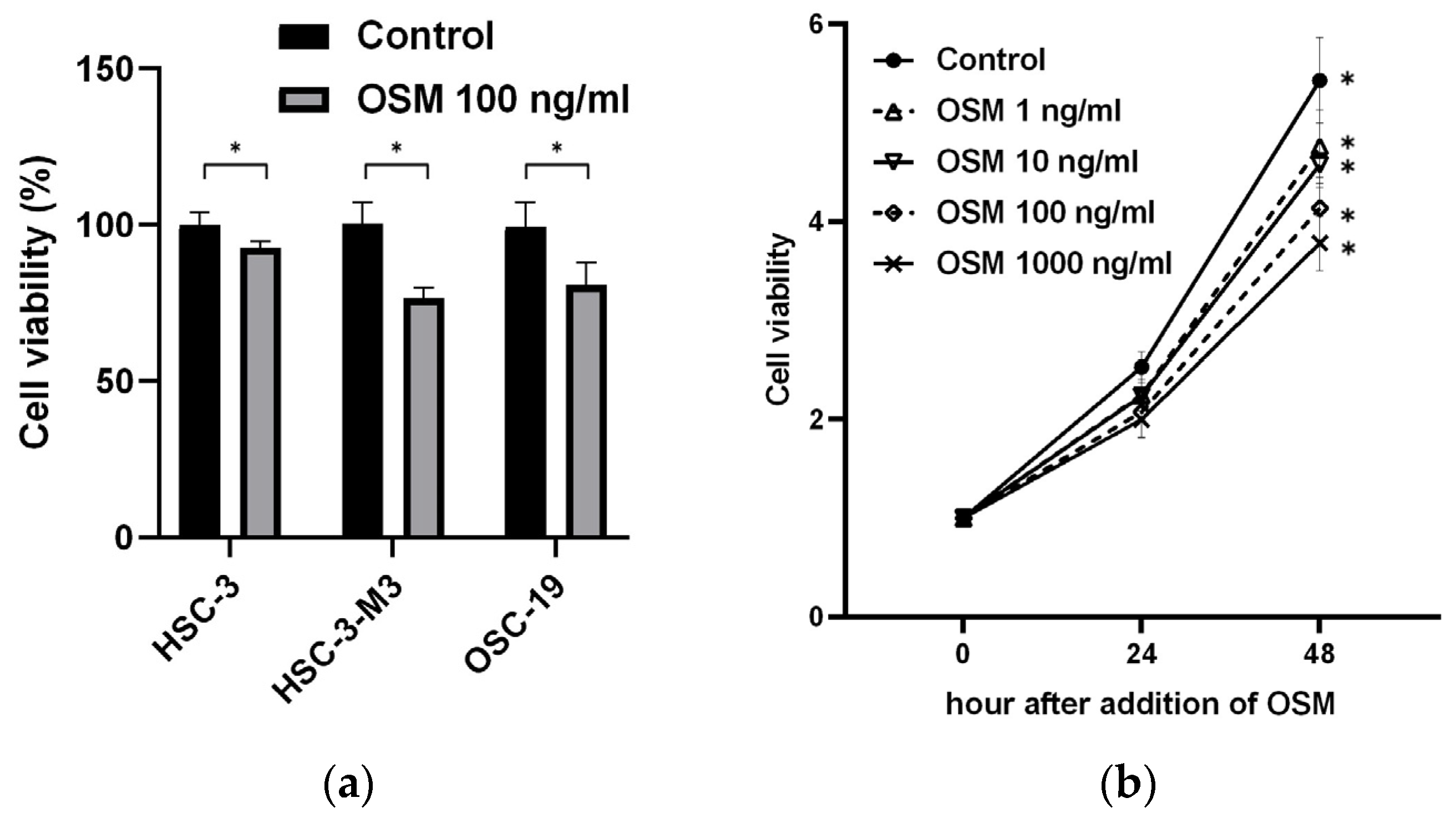
3.2.1. OSM Inhibits Cancer Cell Growth
3.2.2. OSM Inhibits Cell Cycle Progression in HSC-3-M3 Cells
3.2.3. OSM Increases the Expression of the CDK Inhibitors p21 and p27 in HSC-3-M3 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pallis, A.G.; Fortpied, C.; Wedding, U.; Van Nes, M.C.; Penninckx, B.; Ring, A.; Lacombe, D.; Monfardini, S.; Scalliet, P.; Wildiers, H. EORTC elderly task force position paper: Approach to the older cancer patient. Eur. J. Cancer 2010, 46, 1502–1513. [Google Scholar] [CrossRef]
- Boyle, H.J.; Alibhai, S.; Decoster, L.; Efstathiou, E.; Fizazi, K.; Mottet, N.; Oudard, S.; Payne, H.; Prentice, M.; Puts, M.; et al. Updated recommendations of the International Society of Geriatric Oncology on prostate cancer management in older patients. Eur. J. Cancer 2019, 116, 116–136. [Google Scholar] [CrossRef]
- Mizutani, T.; Nakamura, K.; Fukuda, H.; Ogawa, A.; Hamaguchi, T.; Nagashima, F. Geriatric research policy: Japan clinical oncology group (JCOG) policy. Jpn. J. Clin. Oncol. 2019, 49, 901–910. [Google Scholar] [CrossRef]
- Chindapasirt, J. Sarcopenia in cancer patients. Asian Pac. J. Cancer Prev. 2015, 16, 8075–8077. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.N.; Choi, K.M. Sarcopenia: Definition, epidemiology, and pathophysiology. J. Bone Metab. 2013, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.T.; Quinn, L.S. Sarcopenia, obesity, and natural killer cell immune senescence in aging: Altered cytokine levels as a common mechanism. Aging 2012, 4, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Dou, Q.L.; Zeng, Y.; Yang, Y.; Cheng, A.S.K.; Zhang, W.W. Sarcopenia as a predictor of mortality in women with breast cancer: A meta-analysis and systematic review. BMC Cancer 2020, 20, 172. [Google Scholar] [CrossRef]
- Takiguchi, K.; Furuya, S.; Sudo, M.; Saito, R.; Yamamoto, A.; Ashizawa, N.; Hirayama, K.; Shoda, K.; Akaike, H.; Hosomura, N.; et al. Prognostic effect of sarcopenia in colorectal cancer recurrence. Nutrition 2021, 91–92, 11136. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yagyu, T.; Uchinaka, E.; Miyatani, K.; Hanaki, T.; Kihara, K.; Matsunaga, T.; Yamamoto, M.; Tokuyasu, N.; Honjo, S.; et al. Sarcopenia as a prognostic factor in patients with recurrent pancreatic cancer: A retrospective study. World J. Surg. Oncol. 2020, 18, 221. [Google Scholar] [CrossRef]
- Katsui, K.; Ogata, T.; Sugiyama, S.; Yoshio, K.; Kuroda, M.; Hiraki, T.; Kiura, K.; Maeda, Y.; Toyooka, S.; Kanazawa, S. Sarcopenia is associated with poor prognosis after chemoradiotherapy in patients with stage iii non-small-cell lung cancer: A retrospective analysis. Sci. Rep. 2021, 11, 11882. [Google Scholar] [CrossRef]
- Yoshimura, T.; Suzuki, H.; Takayama, H.; Higashi, S.; Hirano, Y.; Tezuka, M.; Ishida, T.; Ishihata, K.; Nishi, Y.; Nakamura, Y.; et al. Impact of preoperative low prognostic nutritional index and high intramuscular adipose tissue content on outcomes of patients with oral squamous cell carcinoma. Cancers 2020, 12, 3167. [Google Scholar] [CrossRef]
- Yoshimura, T.; Suzuki, H.; Takayama, H.; Higashi, S.; Hirano, Y.; Tezuka, M.; Ishida, T.; Ishihata, K.; Nishi, Y.; Nakamura, Y.; et al. Prognostic role of preoperative sarcopenia evaluation of cervical muscles with long-term outcomes of patients with oral squamous cell carcinoma. Cancers 2021, 13, 4725. [Google Scholar] [CrossRef]
- Olson, B.; Edwards, J.; Degnin, C.; Santucci, N.; Buncke, M.; Hu, J.; Chen, Y.; Fuller, C.D.; Geltzeiler, M.; Grossberg, A.J.; et al. Establishment and Validation of Pre-Therapy Cervical Vertebrae Muscle Quantification as a Prognostic Marker of Sarcopenia in Patients With Head and Neck Cancer. Front. Oncol. 2022, 14, 812159. [Google Scholar] [CrossRef] [PubMed]
- Barajas-Galindo, D.E.; Arnáiz, E.G.; Vicente, P.F.; Ballesteros-Pomar, M.D. Effects of physical exercise in sarcopenia. A systematic review. Endocrinol. Diabetes Nutr. 2021, 68, 159–169. [Google Scholar] [CrossRef]
- Pedersen, L.; Idorn, M.; Olofsson, G.H.; Lauenborg, B.; Nookaew, I.; Hansen, R.H.; Johannesen, H.H.; Becker, J.C.; Pedersen, K.S.; Dethlefsen, C.; et al. Voluntary running suppresses tumor growth through epinephrine-and IL-6-dependent NK cell mobilization and redistribution. Cell Metab. 2016, 23, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Ashcraft, K.A.; Peace, R.M.; Betof, A.S.; Dewhirst, M.W.; Jones, L.W. Efficacy and mechanisms of aerobic exercise on cancer initiation, progression, and metastasis: A critical systematic review of in vivo preclinical data. Cancer Res. 2016, 76, 4032–4050. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef] [PubMed]
- Bigley, A.B.; Rezvani, K.; Chew, C.; Sekine, T.; Pistillo, M.; Crucian, B.; Bollard, C.M.; Simpson, R.J. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Brain Behav. Immun. 2014, 39, 160–171. [Google Scholar] [CrossRef]
- Dethlefsen, C.; Hansen, L.S.; Lillelund, C.; Andersen, C.; Gehl, J.; Christensen, J.F.; Pedersen, B.K.; Hojman, P. Exercise-Induced Catecholamines Activate the Hippo Tumor Suppressor Pathway to Reduce Risks of Breast Cancer Development. Cancer Res. 2017, 77, 4894–4904. [Google Scholar] [CrossRef]
- Han, L.; Wu, S.; Hu, P. The functions of sarcopenia related myokines. Transl. Med. Aging 2018, 2, 38–41. [Google Scholar] [CrossRef]
- Gomarasca, M.; Banfi, G.; Lombardi, G. Myokines: The endocrine coupling of skeletal muscle and bone. Adv. Clin. Chem. 2020, 94, 155–218. [Google Scholar] [CrossRef]
- Kim, J.S.; Taaffe, D.R.; Galvão, D.A.; Hart, N.H.; Gray, E.; Ryan, C.J.; Kenfield, S.A.; Saad, F.; Newton, R.U. Exercise in advanced prostate cancer elevates myokine levels and suppresses in-vitro cell growth. Prostate Cancer Prostatic Dis. 2022, 25, 86–92. [Google Scholar] [CrossRef]
- Kim, J.S.; Galvão, D.A.; Newton, R.U.; Gray, E.; Taaffe, D.R. Exercise-induced myokines and their effect on prostate cancer. Nat. Rev. Urol. 2021, 18, 519–542. [Google Scholar] [CrossRef]
- Gannon, N.P.; Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Trujillo, K.A. Effects of the exercise-inducible myokine irisin on malignant and non-malignant breast epithelial cell behavior in vitro. Int. J. Cancer 2015, 136, E197–E202. [Google Scholar] [CrossRef]
- Hoffmann, C.; Weigert, C. Skeletal muscle as an endocrine organ: The role of myokines in exercise adaptations. Cold Spring Harb. Perspect. Med. 2017, 7, a029793. [Google Scholar] [CrossRef]
- Lee, J.H.; Jun, H.S. Role of myokines in regulating skeletal muscle mass and function. Front. Physiol. 2019, 10, 42. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Neto, E.V.P.; Goulart, R.D.A.; Bechara, M.D.; Chagas, E.F.B.; Audi, M.; Campos, L.M.G.; Guiger, E.L.; Buchaim, R.L.; Buchaim, D.V.; et al. Myokines: A descriptive review. J. Sports Med. Phys. Fit. 2020, 60, 1583–1590. [Google Scholar] [CrossRef]
- Masjedi, A.; Hajizadeh, F.; Dargani, F.B.; Beyzai, B.; Aksoun, M.; Hojjat-Farsangi, M.; Zekiy, A.; Jadidi-Niaragh, F. Oncostatin M: A mysterious cytokine in cancers. Int. Immunopharmacol. 2021, 90, 107158. [Google Scholar] [CrossRef]
- Li, C.; Ahlborn, T.E.; Kraemer, F.B.; Liu, J. Oncostatin M-induced growth inhibition and morphological changes of MDA-MB231 breast cancer cells are abolished by blocking the MEK/ERK signaling pathway. Breast Cancer Res. Treat. 2001, 66, 111–121. [Google Scholar] [CrossRef]
- Pan, C.M.; Wang, M.L.; Chiou, S.H.; Chen, H.Y.; Wu, C.W. Oncostatin M suppresses metastasis of lung adenocarcinoma by inhibiting SLUG expression through coordination of STATs and PIASs signalings. Oncotarget 2016, 7, 60395–60406. [Google Scholar] [CrossRef]
- David, E.; Guihard, P.; Brounais, B.; Riet, A.; Charrier, C.; Battaglia, S.; Gouin, F.; Ponsolle, S.; Bot, R.L.; Richards, C.D.; et al. Direct anti-cancer effect of oncostatin M on chondrosarcoma. Int. J. Cancer 2011, 128, 1822–1835. [Google Scholar] [CrossRef] [PubMed]
- Chuerduangphui, J.; Ekalaksananan, T.; Chaiyarit, P.; Patarapadungkit, N.; Chotiyano, A.; Kongyingyoes, B.; Promthet, S.; Pientong, C. Effects of arecoline on proliferation of oral squamous cell carcinoma cells by dysregulating c-Myc and miR-22, directly targeting oncostatin M. PLoS ONE 2018, 13, e0192009. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xiao, W.; Liu, Y.; Liu, R.; Kramer, R.; Li, X.; Ajena, Y.; Baehr, C.M.; Rojalin, T.; Zhang, H.; et al. One-bead one-compound combinatorial library derived targeting ligands for detection and treatment of oral squamous cancer. Oncotarget 2019, 10, 5468–5479. [Google Scholar] [CrossRef]
- Hojman, P.; Stagaard, R.; Adachi-Fernandez, E.; Deshmukh, A.S.; Mund, A.; Olsen, C.H.; Keller, L.; Pedersen, B.K.; Gehl, J. Exercise suppresses tumor growth independent of high fat food intake and associated immune dysfunction. Sci. Rep. 2022, 12, 5476. [Google Scholar] [CrossRef]
- Piguet, A.C.; Saran, U.; Simillion, C.; Piguet, A.C.; Saran, U.; Simillion, C.; Keller, I.; Terracciano, L.; Reeves, H.L.; Dufour, J.F. Regular exercise decreases liver tumors development in hepatocyte-specific PTEN-deficient mice independently of steatosis. J. Hepatol. 2015, 62, 1296–1303. [Google Scholar] [CrossRef]
- Gomes-Santos, I.L.; Amoozgar, Z.; Kumar, A.S.; Ho, W.W.; Roh, K.; Talele, N.P.; Curtis, H.; Kawaguchi, K.; Jain, R.K.; Fukumura, D. Exercise Training Improves Tumor Control by Increasing CD8+ T-cell Infiltration via CXCR3 Signaling and Sensitizes Breast Cancer to Immune Checkpoint Blockade. Cancer Immunol. Res. 2021, 9, 765–778. [Google Scholar] [CrossRef]
- Paceli, R.B.; Cal, R.N.; Dos Santos, C.H.; Cordeiro, J.A.; Neiva, C.M.; Kawano Nagamine, K.; Cury, P.M. The influence of physical activity in the progression of experimental lung cancer in mice. Pathol. Res. Pract. 2012, 208, 377–381. [Google Scholar] [CrossRef]
- Akdeniz, N.; Kaplan, M.A.; Küçüköner, M.; Urakçı, Z.; Laçin, S.; Ceylan, E.H.; Işıkdoğan, A. The effect of exercise on disease-free survival and overall survival in patients with breast cancer. Ir. J. Med. Sci. 2022, 191, 1587–1597. [Google Scholar] [CrossRef]
- Catoire, M.; Kersten, S. The search for exercise factors in humans. FASEB J. 2015, 29, 1615–1628. [Google Scholar] [CrossRef]
- Hojman, P.; Dethlefsen, C.; Brandt, C.; Hansen, J.; Pedersen, L.; Pedersen, B.K. Exercise-induced muscle-derived cytokines inhibit mammary cancer cell growth. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E504–E510. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Taaffe, D.R.; Galvão, D.A.; Clay, T.D.; Redfern, A.D.; Hart, N.H.; Gray, E.S.; Ryan, C.J.; Kenfield, S.A.; Saad, F.; et al. Acute effect of high-intensity interval aerobic exercise on serum myokine levels and resulting tumour-suppressive effect in trained patients with advanced prostate cancer. Prostate Cancer Prostatic Dis. 2023, 26, 795–801. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Yang, G.; Meng, B.; Yi, Z.; Yang, G.; Chen, M.; Hou, P.; Wang, H.; Xu, X. Moderate-Intensity Physical Exercise Affects the Exercise Performance and Gut Microbiota of Mice. Front. Cell. Infect. Microbiol. 2021, 24, 712381. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Exercise | |
|---|---|---|
| Tumor formation rate | 100% (20/20) | 75% (15/20) |
| Tumor volume (mm3) | 26.2 ± 16.9 | 6.4 ± 4.3 |
| Control | OSM | p Value | |
|---|---|---|---|
| Sub-G1 | 0.628333 | 0.892333 | 0.304222 |
| G0/G1 | 51.74 | 58.68867 | 0.002743 |
| S | 25.42567 | 23.195 | 0.078736 |
| G2/M | 20.63833 | 13.94633 | 0.016104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshimura, T.; Hirano, Y.; Hamada, T.; Yokoyama, S.; Suzuki, H.; Takayama, H.; Migita, H.; Ishida, T.; Nakamura, Y.; Ohsawa, M.; et al. Exercise Suppresses Head and Neck Squamous Cell Carcinoma Growth via Oncostatin M. Cancers 2024, 16, 1187. https://doi.org/10.3390/cancers16061187
Yoshimura T, Hirano Y, Hamada T, Yokoyama S, Suzuki H, Takayama H, Migita H, Ishida T, Nakamura Y, Ohsawa M, et al. Exercise Suppresses Head and Neck Squamous Cell Carcinoma Growth via Oncostatin M. Cancers. 2024; 16(6):1187. https://doi.org/10.3390/cancers16061187
Chicago/Turabian StyleYoshimura, Takuya, Yuka Hirano, Taiji Hamada, Seiya Yokoyama, Hajime Suzuki, Hirotaka Takayama, Hirono Migita, Takayuki Ishida, Yasunori Nakamura, Masahiro Ohsawa, and et al. 2024. "Exercise Suppresses Head and Neck Squamous Cell Carcinoma Growth via Oncostatin M" Cancers 16, no. 6: 1187. https://doi.org/10.3390/cancers16061187
APA StyleYoshimura, T., Hirano, Y., Hamada, T., Yokoyama, S., Suzuki, H., Takayama, H., Migita, H., Ishida, T., Nakamura, Y., Ohsawa, M., Asakawa, A., Ishihata, K., & Tanimoto, A. (2024). Exercise Suppresses Head and Neck Squamous Cell Carcinoma Growth via Oncostatin M. Cancers, 16(6), 1187. https://doi.org/10.3390/cancers16061187