
Immune Checkpoint Inhibitors and Male Fertility: Should Fertility Preservation Options Be Considered before Treatment?


Abstract
Simple Summary
Abstract
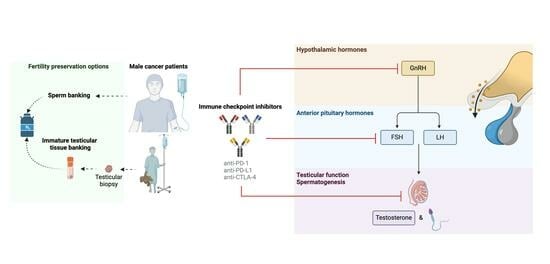
1. Introduction
2. Immune Checkpoint Inhibitors
2.1. Mechanism of Action
2.2. Available Therapies
3. Immunoregulation of the Testis
3.1. The Blood–Testis Barrier
3.2. Immune Cell Repertoire in the Testis
3.3. Testicular Cells
4. Endocrinological and Testicular Changes during Development
5. PD-1/PD-L1 and CTLA-4 Expression in the Testis
6. Direct and Indirect Effects on Testicular Function
7. Discussion: Fertility Preservation before ICI Treatment and Future Perspectives
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Chen, J. Current status and future directions of cancer immunotherapy. J. Canc. 2018, 9, 1773–1781. [Google Scholar] [CrossRef]
- Smyth, M.J.; Teng, M.W. 2018 Nobel Prize in physiology or medicine. CTI 2018, 7, e1041. [Google Scholar] [CrossRef]
- Postow, M.A.; Sidlow, R.; Hellmann, M.D. Immune-related adverse events associated with immune checkpoint blockade. N. Engl. J. Med. 2018, 378, 158–168. [Google Scholar] [CrossRef]
- Mulder, R.L.; Font-Gonzalez, A.; Green, D.M.; Loeffen, E.A.H.; Hudson, M.M.; Loonen, J.; Yu, R.; Ginsberg, J.P.; Mitchell, R.T.; Byrne, J.; et al. Fertility preservation for male patients with childhood, adolescent, and young adult cancer: Recommendations from the PanCareLIFE Consortium and the International Late Effects of Childhood Cancer Guideline Harmonization Group. Lancet Oncol. 2021, 22, e57–e67. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A new member of the immunoglobulin superfamily—CTLA-4. Nature 1987, 328, 267. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; See, A.P.; Phallen, J.; Jackson, C.M.; Belcaid, Z.; Ruzevick, J.; Durham, N.; Meyer, C.; Harris, T.J.; Albesiano, E.; et al. Anti-PD-1 blockade and stereotactic radiation produce long-term survival in mice with intracranial gliomas. Int. J. Radiat. Oncol. Biol. Phys. 2013, 86, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.; Pauken, K. The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 2018, 18, 153–167. [Google Scholar] [CrossRef]
- Webb, E.S.; Liu, P.; Baleeiro, R.; Lemoine, N.R.; Yuan, M.; Wang, Y. Immune checkpoint inhibitors in cancer therapy. J. Biomed. Res. 2018, 32, 317–326. [Google Scholar] [PubMed]
- Wei, S.C.; Duffy, C.R.; Allison, J.P. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov. 2018, 8, 1069–1086. [Google Scholar] [CrossRef] [PubMed]
- Pawłowska, A.; Suszczyk, D.; Okła, K.; Barczynski, B.; Kotarski, J.; Wertel, I. Immunotherapies based on PD-1/PD-L1 pathway inhibitors in ovarian cancer treatment. Clin. Exp. Immunol. 2019, 195, 334–344. [Google Scholar] [CrossRef]
- Egen, J.; Kuhns, M.; Allison, J. CTLA-4: New insights into its biological function and use in tumor immunotherapy. Nat. Immunol. 2002, 3, 611–618. [Google Scholar] [CrossRef]
- Hoos, A. Development of immuno-oncology drugs—From CTLA4 to PD1 to the next generations. Nat. Rev. Drug. Discov. 2016, 15, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Goswami, S.; Raychaudhuri, D.; Siddiqui, B.A.; Singh, P.; Nagarajan, A.; Liu, J.; Subudhi, S.K.; Poon, C.; Gant, K.L.; et al. Immune checkpoint therapy-current perspectives and future directions. Cell 2023, 186, 1652–1669. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 23 January 2024).
- Vaddepally, R.K.; Kharel, P.; Pandey, R.; Garje, R.; Chandra, A.B. Review of indications of FDA-approved immune checkpoint inhibitors per NCCN guidelines with the level of evidence. Cancers 2020, 12, 738. [Google Scholar] [CrossRef] [PubMed]
- Tanda, E.T.; Croce, E.; Spagnolo, F.; Zullo, L.; Spinaci, S.; Genova, C.; Rossi, G. Immunotherapy in adolescents and young adults: What remains in cancer survivors? Front. Oncol. 2021, 11, 736123. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Day, D.; Nicholls, S.J.; Segelov, E. Immune checkpoint inhibitor therapy in oncology: Current uses and future directions. Cardio Oncol. 2022, 4, 579–597. [Google Scholar]
- Drugs@FDA: FDA-Approved Drugs. Available online: https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm (accessed on 11 January 2024).
- Pediatric Oncology Drug Approvals. Available online: https://www.fda.gov/about-fda/oncology-center-excellence/pediatric-oncology-drug-approvals (accessed on 11 January 2024).
- Chen, Q.; Deng, T.; Han, D. Testicular immunoregulation and spermatogenesis. Semin. Cell Dev. Biol. 2016, 59, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhu, W.; Xue, S.; Han, D. Testicular defense systems: Immune privilege and innate immunity. Cell. Mol. Immunol. 2014, 11, 428–437. [Google Scholar] [CrossRef]
- Khambata, K.; Modi, D.N.; Gupta, S.K. Immunoregulation in the testis and its implication in fertility and infections. Explor. Immunol. 2021, 1, 309–324. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.D. The blood–testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef]
- Fijak, M.; Bhushan, S.; Meinhardt, A. Immunoprivileged sites: The testis. Methods Mol. Biol. 2011, 677, 459–470. [Google Scholar] [PubMed]
- Bhushan, S.; Theas, M.S.; Guazzone, V.A.; Jacobo, P.; Wang, M.; Fijak, M.; Meinhardt, A.; Lustig, L. Immune cell subtypes and their function in the testis. Front. Immunol. 2020, 11, 583304. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P. Macrophages and the immune responsiveness of the testis. J. Reprod. Immunol. 2002, 57, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fijak, M.; Hossain, H.; Markmann, M.; Nüsing, R.M.; Lochnit, G.; Hartmann, M.F.; Wudy, S.A.; Zhang, L.; Gu, H.; et al. Characterization of the micro-environment of the testis that shapes the phenotype and function of testicular macrophages. J. Immunol. 2017, 198, 4327–4340. [Google Scholar] [CrossRef] [PubMed]
- Winnall, W.R.; Muir, J.A.; Hedger, M.P. Rat resident testicular macrophages have an alternatively activated phenotype and constitutively produce interleukin-10 in vitro. J. Leukoc. Biol. 2011, 90, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Mossadegh-Keller, N.; Sieweke, M.H. Testicular macrophages: Guardians of fertility. Cell. Immunol. 2018, 330, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Rival, C.; Guazzone, V.A.; von Wulffen, W.; Hackstein, H.; Schneider, E.; Lustig, L.; Meinhardt, A.; Fijak, M. Expression of co-stimulatory molecules, chemokine receptors and proinflammatory cytokines in dendritic cells from normal and chronically inflamed rat testis. Mol. Hum. Reprod. 2007, 13, 853–861. [Google Scholar] [CrossRef]
- Duan, Y.G.; Yu, C.F.; Novak, N.; Bieber, T.; Zhu, C.H.; Schuppe, H.C.; Haidl, G.; Allam, J.P. Immunodeviation towards a Th17 immune response associated with testicular damage in azoospermic men. Int. J. Androl. 2011, 34, e536–e545. [Google Scholar] [CrossRef]
- Haidl, G.; Duan, Y.G.; Chen, S.J.; Kohn, F.M.; Schuppe, H.C.; Allam, J.P. The role of mast cells in male infertility. Expert Rev. Clin. Immunol. 2011, 7, 627–634. [Google Scholar] [CrossRef]
- Mayerhofer, A.; Walenta, L.; Mayer, C.; Eubler, K.; Welter, H. Human testicular peritubular cells, mast cells and testicular inflammation. Andrologia 2018, 50, e13055. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.; Sobarzo, C.M.; Lustig, L.; Rodriguez Pena, M.G.; Guazzone, V.A. Effect of ketotifen fumarate on experimental autoimmune orchitis and torsion of the spermatic cord. Asian J. Androl. 2020, 22, 112–117. [Google Scholar] [PubMed]
- Khan, U.; Ghazanfar, H. T lymphocytes and autoimmunity. Int. Rev. Cell. Mol. Biol. 2018, 341, 125–168. [Google Scholar] [PubMed]
- Hedger, M.P. Testicular leukocytes: What are they doing? Rev. Reprod. 1997, 2, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zeng, Q.; Yu, D.; Duan, Y.G. T lymphocytes and testicular immunity: A new insight into immune regulation in testes. Int. J. Mol. Sci. 2020, 22, 57. [Google Scholar] [CrossRef] [PubMed]
- Garza, K.M.; Agersborg, S.S.; Baker, E.; Tung, K.S. Persistence of physiological self-antigen is required for the regulation of self-tolerance. J. Immunol. 2000, 164, 3982–3989. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, K.; Tardif, S.; Rival, C.; Luu, B.; Bui, E.; Del Rio, R.; Teuscher, C.; Sparwasser, T.; Hardy, D.; Tung, K.S. Regulatory T cells control tolerogenic versus autoimmune response to sperm in vasectomy. Proc. Natl. Acad. Sci. USA 2011, 108, 7511–7516. [Google Scholar] [CrossRef]
- Nasr, I.W.; Wang, Y.; Gao, G.; Deng, S.; Diggs, L.; Rothstein, D.M.; Tellides, G.; Lakkis, F.G.; Dai, Z. Testicular immune privilege promotes transplantation tolerance by altering the balance between memory and regulatory T cells. J. Immunol. 2005, 174, 6161–6168. [Google Scholar] [CrossRef]
- Jacobo, P.; Guazzone, V.A.; Jarazo-Dietrich, S.; Theas, M.S.; Lustig, L. Differential changes in CD4+ and CD8+ effector and regulatory T lymphocyte subsets in the testis of rats undergoing autoimmune orchitis. J. Reprod. Immunol. 2009, 81, 44–54. [Google Scholar] [CrossRef]
- Jacobo, P. The role of regulatory T Cells in autoimmune orchitis. Andrologia 2018, 50, e13092. [Google Scholar] [CrossRef]
- Jacobo, P.; Perez, C.V.; Theas, M.S.; Guazzone, V.A.; Lustig, L. CD4+ and CD8+ T cells producing Th1 and Th17 cytokines are involved in the pathogenesis of autoimmune orchitis. Reproduction 2011, 141, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P.; Meinhardt, A. Local regulation of T cell numbers and lymphocyte-inhibiting activity in the interstitial tissue of the adult rat testis. J. Reprod. Immunol. 2000, 48, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Hedger, M.P.; Winnall, W.R. Regulation of activin and inhibin in the adult testis and the evidence for functional roles in spermatogenesis and immunoregulation. Mol. Cell. Endocrinol. 2012, 359, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Pinzon, W.; Korbutt, G.S.; Power, R.; Hooton, J.; Rajotte, R.V.; Rabinovitch, A. Testicular Sertoli cells protect islet beta-cells from autoimmune destruction in NOD mice by a transforming growth factor-beta1-dependent mechanism. Diabetes 2000, 49, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, T.; Deng, T.; Xiong, W.; Lui, P.; Li, N.; Chen, Y.; Han, D. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Mol. Cell. Endocrinol. 2013, 365, 162–173. [Google Scholar] [CrossRef]
- Fijak, M.; Schneider, E.; Klug, J.; Bhushan, S.; Hackstein, H.; Schuler, G.; Wygrecka, M.; Gromoll, J.; Meinhardt, A. Testosterone replacement effectively inhibits the development of experimental autoimmune orchitis in rats: Evidence for a direct role of testosterone on regulatory T cell expansion. J. Immunol. 2011, 186, 5162–5172. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Adam, M.; Glashauser, L.; Dietrich, K.; Schwarzer, J.U.; Köhn, F.M.; Strauss, L.; Welter, H.; Poutanen, M.; Mayerhofer, A. Sterile inflammation as a factor in human male infertility: Involvement of Toll-like receptor 2, biglycan and peritubular cells. Sci. Rep. 2016, 6, 37128. [Google Scholar] [CrossRef]
- Haugen, T.B.; Landmark, B.F.; Josefsen, G.M.; Hansson, V.; Hogset, A. The mature form of interleukin-1 alpha is constitutively expressed in immature male germ cells from rat. Mol. Cell. Endocrinol. 1994, 105, R19–R23. [Google Scholar] [CrossRef]
- Rey, R.A. Mini-puberty and true puberty: Differences in testicular function. Ann. Endocrinol. 2014, 75, 58–63. [Google Scholar] [CrossRef]
- Mitchell, R.T.; O’Hara, L.; Smith, L.B. Gonadotropin and steroid hormone control of spermatogonial differentiation. In The Biology of Mammalian Spermatogonia, 1st ed.; Oatley, J.M., Griswold, M.D., Eds.; Springer Press: New York, NY, USA, 2017; pp. 4555–4570. [Google Scholar]
- Hermann, B.P.; Sukhwani, M.; Hansel, M.C.; Orwig, K.E. Spermatogonial stem cells in higher primates: Are there differences from those in rodents? Reproduction 2010, 139, 479–493. [Google Scholar] [CrossRef]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and functional maturation of Sertoli cells, and their relevance to disorders of testis function in adulthood. Reproduction 2003, 125, 769–784. [Google Scholar] [CrossRef]
- de Kretser, D.M.; Loveland, K.; O’Bryan, M. Spermatogenesis. In Endocrinology: Adult and Pediatric, 7th ed.; Jameson, J.L., De Groot, L.J., de Kretser, D.D., Giudice, L.C., Grossman, A.B., Melmed, S., Potts, J.T., Weir, G.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 3, pp. 2325–2353.e9. [Google Scholar]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Dai, H.; Wan, N.; Moore, Y.; Vankayalapati, R.; Dai, Z. Interaction of programmed death-1 and programmed death-1 ligand-1 contributes to testicular immune privilege. Transplantation 2009, 87, 1778–1786. [Google Scholar] [CrossRef] [PubMed]
- Dal Secco, V.; Riccioli, A.; Padula, F.; Ziparo, E.; Filippini, A. Mouse Sertoli cells display phenotypical and functional traits of antigen-presenting cells in response to interferon gamma. Biol. Reprod. 2008, 78, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Li, Z.H.; Duan, Y.G.; Yuan, S.Q.; Mor, G.; Liao, A.H. Identification of programmed cell death 1 and its ligand in the testicular tissue of mice. Am. J. Reprod. Immunol. 2019, 81, e13079. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, C.D.; Honecker, F.; Beyer, J.; Bode, P.K. Emerging therapeutic targets for male germ cell tumors. Curr. Oncol. Rep. 2015, 17, 54. [Google Scholar] [CrossRef] [PubMed]
- Cierna, Z.; Mego, M.; Miskovska, V.; Machalekova, K.; Chovanec, M.; Svetlovska, D.; Hainova, K.; Rejlekova, K.; Macak, D.; Spanik, S.; et al. Prognostic value of programmed-death-1 receptor (PD-1) and its ligand 1 (PD-L1) in testicular germ cell tumors. Ann. Oncol. 2016, 27, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, L.; Bartsch, G.; Gust, K.; Kvasnicka, H.M.; Haferkamp, A.; Blaheta, R.; Mittelbronn, M.; Harter, P.N.; Mani, J. Increased tumor vascularization is associated with the amount of immune competent PD-1 positive cells in testicular germ cell tumors. Oncol. Lett. 2018, 15, 9852–9860. [Google Scholar] [CrossRef]
- Fang, L.; Feng, R.; Liang, W.; Liu, F.F.; Bian, G.L.; Yu, C.; Guo, H.; Cao, Y.; Liu, M.; Zuo, J.; et al. Overexpression of PD-L1 causes germ cells to slough from mouse seminiferous tubules via the PD-L1/PD-L1 interaction. J. Cell. Mol. Med. 2022, 26, 2908–2920. [Google Scholar] [CrossRef]
- Shinohara, T.; Yamamoto, T.; Morimoto, H.; Shiromoto, Y.; Kanatsu-Shinohara, M. Allogeneic offspring produced by induction of PD-L1 in spermatogonial stem cells via self-renewal stimulation. Stem Cell Rep. 2023, 18, 985–998. [Google Scholar] [CrossRef]
- Garutti, M.; Lambertini, M.; Puglisi, F. Checkpoint inhibitors, fertility, pregnancy, and sexual life: A systematic review. ESMO Open 2021, 6, 100276. [Google Scholar] [CrossRef]
- Santaballa, A.; Márquez-Vega, C.; Rodríguez-Lescure, Á.; Rovirosa, Á.; Vázquez, L.; Zeberio-Etxetxipia, I.; Andrés, M.; Bassas, L.; Ceballos-Garcia, E.; Domingo, J.; et al. Multidisciplinary consensus on the criteria for fertility preservation in cancer patients. Clin. Transl. Oncol. 2022, 24, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Scovell, J.M.; Benz, K.; Samarska, I.; Kohn, T.P.; Hooper, J.E.; Matoso, A.; Herati, A.S. Association of impaired spermatogenesis with the use of immune checkpoint inhibitors in patients with metastatic melanoma. JAMA Oncol. 2020, 6, 1297–1299. [Google Scholar] [CrossRef]
- Rabinowitz, M.J.; Kohn, T.P.; Peña, V.N.; Samarska, I.V.; Matoso, A.; Herati, A.S. Onset of azoospermia in man treated with ipilimumab/nivolumab for BRAF negative metastatic melanoma. Urol. Case Rep. 2020, 34, 101488. [Google Scholar] [CrossRef] [PubMed]
- Salzmann, M.; Tosev, G.; Heck, M.; Schadendorf, D.; Maatouk, I.; Enk, A.H.; Hartmann, M.; Hassel, J.C. Male fertility during and after immune checkpoint inhibitor therapy: A cross-sectional pilot study. Eur. J. Cancer 2021, 152, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, B.C. Immune checkpoint inhibitor-related hypogonadism and infertility: A neglected issue in immuno-oncology. J. ImmunoTher. Cancer 2021, 9, e002220. [Google Scholar] [CrossRef]
- Brunet-Possenti, F.; Opsomer, M.A.; Gomez, L.; Ouzaid, I.; Descamps, V. Immune checkpoint inhibitors-related orchitis. Ann. Oncol. 2017, 28, 906–907. [Google Scholar] [CrossRef]
- Quach, H.T.; Robbins, C.J.; Balko, J.M.; Chiu, C.Y.; Miller, S.; Wilson, M.R.; Nelson, G.E.; Johnson, D.B. Severe epididymo-orchitis and encephalitis complicating anti-PD-1 therapy. Oncologist 2019, 24, 872–876. [Google Scholar] [CrossRef]
- Ryder, M.; Callahan, M.; Postow, M.A.; Wolchok, J.; Fagin, J.A. Endocrine-related adverse events following ipilimumab in patients with advanced melanoma: A comprehensive retrospective review from a single institution. Endocr. Relat. Cancer 2014, 21, 371–381. [Google Scholar] [CrossRef]
- Nogueira, E.; Newsom-Davis, T.; Morganstein, D.L. Immunotherapy-induced endocrinopathies: Assessment, management, and monitoring. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018819896182. [Google Scholar] [CrossRef]
- Bai, X.; Lin, X.; Zheng, K.; Chen, X.; Wu, X.; Huang, Y.; Zhuang, Y. Mapping endocrine toxicity spectrum of immune checkpoint inhibitors: A disproportionality analysis using the WHO adverse drug reaction database, VigiBase. Endocrine 2020, 69, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Eigentler, T.K.; Hassel, J.C.; Berking, C.; Aberle, J.; Bachmann, O.; Grünwald, V.; Kähler, K.C.; Loquai, C.; Reinmuth, N.; Steins, M.; et al. Diagnosis, monitoring and management of immune-related adverse drug reactions of anti-PD-1 antibody therapy. Cancer Treat. Rev. 2016, 45, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Hassel, J.C.; Heinzerling, L.; Aberle, J.; Bähr, O.; Eigentler, T.K.; Grimm, M.O.; Grünwald, V.; Leipe, J.; Reinmuth, N.; Tietze, J.K.; et al. Combined immune checkpoint blockade (anti-PD-1/anti-CTLA-4): Evaluation and management of adverse drug reactions. Cancer Treat. Rev. 2017, 57, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Faje, A.T.; Sullivan, R.; Lawrence, D.; Tritos, N.A.; Fadden, R.; Klibanski, A.; Nachtigall, L. Ipilimumab-induced hypophysitis: A detailed longitudinal analysis in a large cohort of patients with metastatic melanoma. J. Clin. Endocrinol. Metab. 2014, 99, 4078–4085. [Google Scholar] [CrossRef]
- Albarel, F.; Gaudy, C.; Castinetti, F.; Carré, T.; Morange, I.; Conte-Devolx, B.; Grob, J.J.; Brue, T. Long-Term follow-up of ipilimumab-induced hypophysitis, a common adverse event of the anti-CTLA-4 antibody in melanoma. Eur. J. Endocrinol. 2015, 172, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Sousa, R.; Barry, W.T.; Garrido-Castro, A.C.; Hodi, F.S.; Min, L.; Krop, I.E.; Tolaney, S.M. Incidence of endocrine dysfunction following the use of different immune checkpoint inhibitor regimens: A systematic review and meta-analysis. JAMA Oncol. 2018, 4, 173–182. [Google Scholar] [CrossRef]
- Garon-Czmil, J.; Petitpain, N.; Rouby, F.; Sassier, M.; Babai, S.; Yéléhé-Okouma, M.; Weryha, G.; Klein, M.; Gillet, P. mmune checkpoint inhibitors-induced hypophysitis: A retrospective analysis of the French Pharmacovigilance database. Sci. Rep. 2019, 9, 19419. [Google Scholar] [CrossRef]
- Peters, M.; Pearlman, A.; Terry, W.; Mott, S.L.; Monga, V. Testosterone deficiency in men receiving immunotherapy for malignant melanoma. Oncotarget 2021, 12, 199–208. [Google Scholar] [CrossRef]
- Poorvu, P.D.; Frazier, A.L.; Feraco, A.M.; Manley, P.E.; Ginsburg, E.S.; Laufer, M.R.; LaCasce, A.S.; Diller, L.R.; Partridge, A.H. Cancer treatment-related infertility: A critical review of the evidence. JNCI Cancer Spectr. 2019, 3, pkz008. [Google Scholar] [CrossRef]
- Meistrich, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis in humans. Fertil. Steril. 2013, 100, 1180–1186. [Google Scholar] [CrossRef]
- Delgouffe, E.; Braye, A.; Goossens, E. Testicular tissue banking for fertility preservation in young boys: Which patients should be included? Front. Endocrinol. 2022, 13, 854186. [Google Scholar] [CrossRef]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European perspective on testicular tissue cryopreservation for fertility preservation in prepubertal and adolescent boys. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef]
- Goossens, E.; Jahnukainen, K.; Mitchell, R.T.; van Pelt, A.; Pennings, G.; Rives, N.; Poels, J.; Wyns, C.; Lane, S.; Rodriguez-Wallberg, K.A.; et al. Fertility preservation in boys: Recent developments and new insights. Hum. Reprod. Open 2020, 2020, hoaa016. [Google Scholar] [CrossRef] [PubMed]
- Cosci, I.; Grande, G.; Di Nisio, A.; Rocca, M.S.; Del Fiore, P.; Benna, C.; Mocellin, S.; Ferlin, A. Cutaneous Melanoma and hormones: Focus on sex differences and the testis. Int. J. Mol. Sci. 2022, 24, 599. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Crawford, J.D. The relationship of gonadal activity and chemotherapy-induced gonadal damage. JAMA 1988, 259, 2123–2125. [Google Scholar] [CrossRef] [PubMed]
- de Rooij, D.G.; Creemers, L.B.; den Ouden, K.; Izadyar, F. Spermatogonial stem cell development. In Testicular Tangrams, 1st ed.; Rommerts, F.F.G., Teerds, K.J., Eds.; Springer Press: Berlin/Heidelberg, Germany, 2002; pp. 121–138. [Google Scholar]
- Jahnukainen, K.; Ehmcke, J.; Hou, M.; Schlatt, S. Testicular function and fertility preservation in male cancer patients. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Bahadur, G.; Ling, K.L.; Hart, R.; Ralph, D.; Wafa, R.; Ashraf, A.; Jaman, N.; Mahmud, S.; Oyede, A.W. Semen quality and cryopreservation in adolescent cancer patients. Hum. Reprod. 2002, 17, 3157–3161. [Google Scholar] [CrossRef]
- Hagenäs, I.; Jørgensen, N.; Rechnitzer, C.; Sommer, P.; Holm, M.; Schmiegelow, K.; Daugaard, G.; Jacobsen, N.; Juul, A. Clinical and biochemical correlates of successful semen collection for cryopreservation from 12-18-year-old patients: A single-center study of 86 adolescents. Hum. Reprod. 2010, 25, 2031–2038. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Drug (Brand Name) | First Approval Date | Drug Target | Indications |
|---|---|---|---|
| Ipilimumab (Yervoy) | 25 March 2011 | CTLA-4 | Melanoma, renal cell carcinoma, colorectal cancer, hepatocellular carcinoma, and non-small cell lung cancer |
| Pembrolizumab (Keytruda) | 4 September 2014 | PD-1 | Melanoma, non-small cell lung cancer, head and neck squamous cell cancer, classical Hodgkin lymphoma, primary mediastinal large B-cell lymphoma, urothelial carcinoma, microsatellite instability-high or mismatch repair deficient cancer, microsatellite instability-high or mismatch repair deficient colorectal cancer, gastric cancer, esophageal cancer, cervical cancer, hepatocellular carcinoma, Merkel cell carcinoma, renal cell carcinoma, endometrial carcinoma, tumor mutational burden-high cancer, cutaneous squamous cell carcinoma, and triple-negative breast cancer |
| Nivolumab (Opdivo) | 22 December 2014 | PD-1 | Melanoma, non-small cell lung cancer, malignant pleural mesothelioma, renal cell carcinoma, classical Hodgkin lymphoma, squamous cell carcinoma of the head and neck, urothelial carcinoma, colorectal cancer, hepatocellular carcinoma, esophageal cancer, gastric cancer, gastroesophageal junction cancer, and esophageal adenocarcinoma |
| Atezolizumab (Tecentriq) | 18 May 2016 | PD-L1 | Urothelial carcinoma, non-small cell lung cancer, small cell lung cancer, hepatocellular carcinoma, and melanoma |
| Avelumab (Bavencio) | 23 March 2017 | PD-L1 | Merkel cell carcinoma, urothelial carcinoma, and renal cell carcinoma |
| Durvalumab (Imfinzi) | 1 May 2017 | PD-L1 | Non-small cell lung cancer, small cell lung cancer, biliary tract cancer, and hepatocellular carcinoma |
| Cemiplimab (Libtayo) | 28 September 2018 | PD-1 | Cutaneous squamous cell carcinoma, basal cell carcinoma, and non-small cell lung cancer |
| Dostarlimab (Jemperli) | 22 April 2021 | PD-1 | Endometrial cancer |
| Nivolumab and Relatlimab (Opdualag) | 18 March 2022 | PD-1, LAG-3 | Melanoma |
| Tremelimumab (Imjudo) | 21 October 2022 | CTLA-4 | Hepatocellular carcinoma |
| Retifanlimab (Zynyz) | 22 March 2023 | PD-1 | Merkel cell carcinoma |
| Toripalimab (Loqtorzi) | 27 October 2023 | PD-1 | Nasopharyngeal carcinoma |
| Drug (Brand Name) | First Approval Date | Drug Target | Indications | Age Range |
|---|---|---|---|---|
| Avelumab (Bavencio) | 23 March 2017 | PD-L1 | Merkel cell carcinoma | 12 years and older |
| Pembrolizumab (Keytruda) | 23 May 2017 | PD-1 | Melanoma, classical Hodgkin lymphoma, primary mediastinal B-cell lymphoma, microsatellite instability-high or mismatch repair deficient cancer, Merkel cell carcinoma, and tumor mutational burden-high cancer | 12 years and older |
| Ipilimumab (Yervoy) | 10 July 2018 | CTLA-4 | Melanoma and colorectal cancer | 12 years and older |
| Nivolumab (Opdivo) | 10 July 2018 | PD-1 | Melanoma and colorectal cancer | 12 years and older |
| Nivolumab and Relatlimab (Opdualag) | 18 March 2022 | PD-1, LAG-3 | Melanoma | 12 years and older |
| Atezolizumab (Tecentriq) | 9 December 2022 | PD-L1 | Alveolar soft part sarcoma | 2 years and older |
| Drug (Brand Name) | Drug Target | Reproductive Findings |
|---|---|---|
| Ipilimumab (Yervoy) | CTLA-4 | No fertility studies performed |
| Pembrolizumab (Keytruda) | PD-1 | No notable effects in male reproductive organs in 1- and 6-month repeat-dose toxicology studies on monkeys; however, most animals in these studies were not sexually mature |
| Nivolumab (Opdivo) | PD-1 | No significant effects on male reproductive organs in 1- and 3-month toxicology studies on monkeys; however, most animals in these studies were not sexually mature |
| Atezolizumab (Tecentriq) | PD-L1 | No notable effects in male reproductive organs in a 26-week repeat-dose toxicity study in monkeys |
| Avelumab (Bavencio) | PD-L1 | No notable effects in male reproductive organs in a 3-month repeat-dose toxicity study in monkeys |
| Durvalumab (Imfinzi) | PD-L1 | No notable effects in male reproductive organs in 3-month repeat-dose toxicology studies on sexually mature monkeys |
| Cemiplimab (Libtayo) | PD-1 | No effects on fertility parameters (semen analysis or testicular measurements) or in male reproductive organs in a 3-month repeat-dose toxicology study in sexually mature monkeys |
| Dostarlimab (Jemperli) | PD-1 | No significant effects on male reproductive organs in 1- and 3-month toxicology studies on monkeys; however, most animals in these studies were not sexually mature |
| Nivolumab and Relatlimab (Opdualag) | PD-1, LAG-3 | Inflammation within the reproductive tract (epididymis, seminal vesicles, and testes) was observed in a 1-month study in monkeys |
| Tremelimumab (Imjudo) | CTLA-4 | No fertility studies performed |
| Retifanlimab (Zynyz) | PD-1 | No significant effects on male reproductive organs in 1- and 3-month toxicology studies on monkeys; however, most animals in these studies were not sexually mature |
| Toripalimab (Loqtorzi) | PD-1 | No notable effects in male reproductive organs in 4- and 26-week repeat-dose toxicology studies in sexually mature monkeys |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntemou, E.; Delgouffe, E.; Goossens, E. Immune Checkpoint Inhibitors and Male Fertility: Should Fertility Preservation Options Be Considered before Treatment? Cancers 2024, 16, 1176. https://doi.org/10.3390/cancers16061176
Ntemou E, Delgouffe E, Goossens E. Immune Checkpoint Inhibitors and Male Fertility: Should Fertility Preservation Options Be Considered before Treatment? Cancers. 2024; 16(6):1176. https://doi.org/10.3390/cancers16061176
Chicago/Turabian StyleNtemou, Elissavet, Emily Delgouffe, and Ellen Goossens. 2024. "Immune Checkpoint Inhibitors and Male Fertility: Should Fertility Preservation Options Be Considered before Treatment?" Cancers 16, no. 6: 1176. https://doi.org/10.3390/cancers16061176
APA StyleNtemou, E., Delgouffe, E., & Goossens, E. (2024). Immune Checkpoint Inhibitors and Male Fertility: Should Fertility Preservation Options Be Considered before Treatment? Cancers, 16(6), 1176. https://doi.org/10.3390/cancers16061176