
The miR-19a/Cylindromatosis Axis Regulates Pituitary Adenoma Bone Invasion by Promoting Osteoclast Differentiation



Abstract
Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Fluorescence In Situ Hybridization (FISH)
2.2. Animal Experiments
2.3. Cell Culture and Reagents
2.4. Osteoclastogenesis Assay
2.5. F-Actin Ring Assay
2.6. Bone Resorption Assay
2.7. Quantitative Real-Time PCR (qRT–PCR)
2.8. Dual-Luciferase Reporter Gene Assay
2.9. Western Blotting (WB) and Co-Immunoprecipitation (Co-IP)
2.10. Electrophoretic Mobility Shift Assay (EMSA)
2.11. Statistical Analysis
3. Results
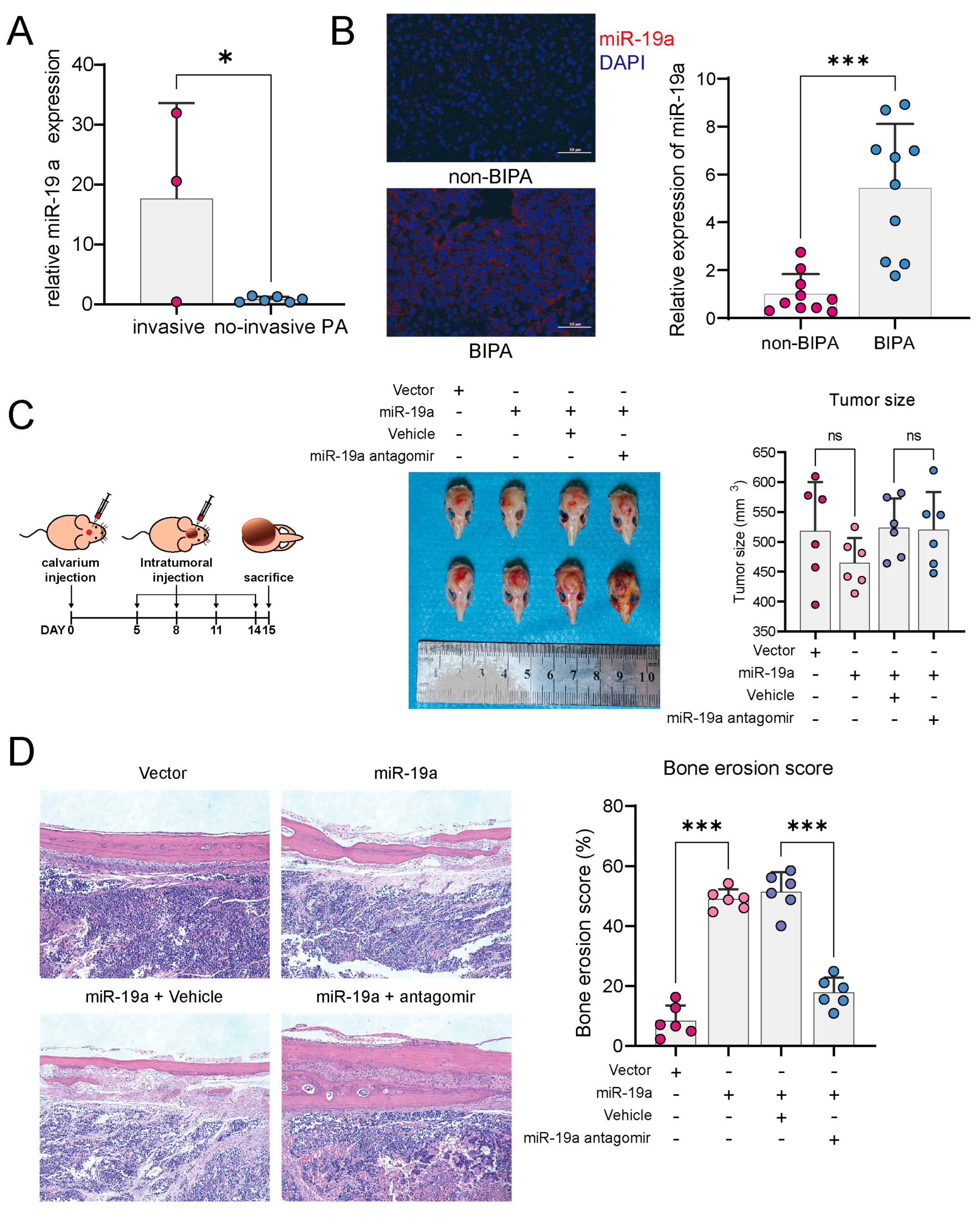
3.1. miR-19a Promotes PA Bone Invasion
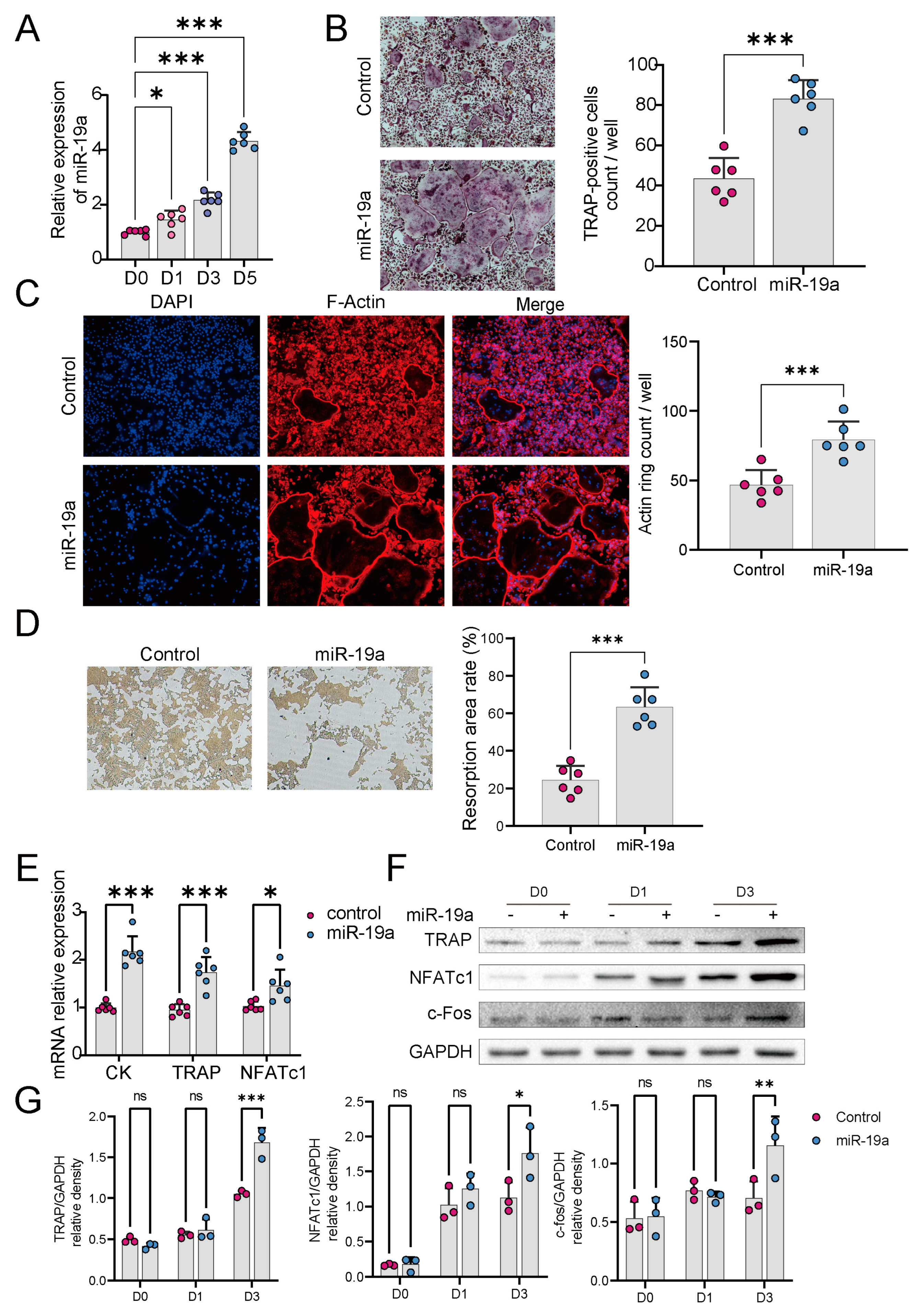
3.2. miR-19a Accelerates Osteoclast Differentiation and Function
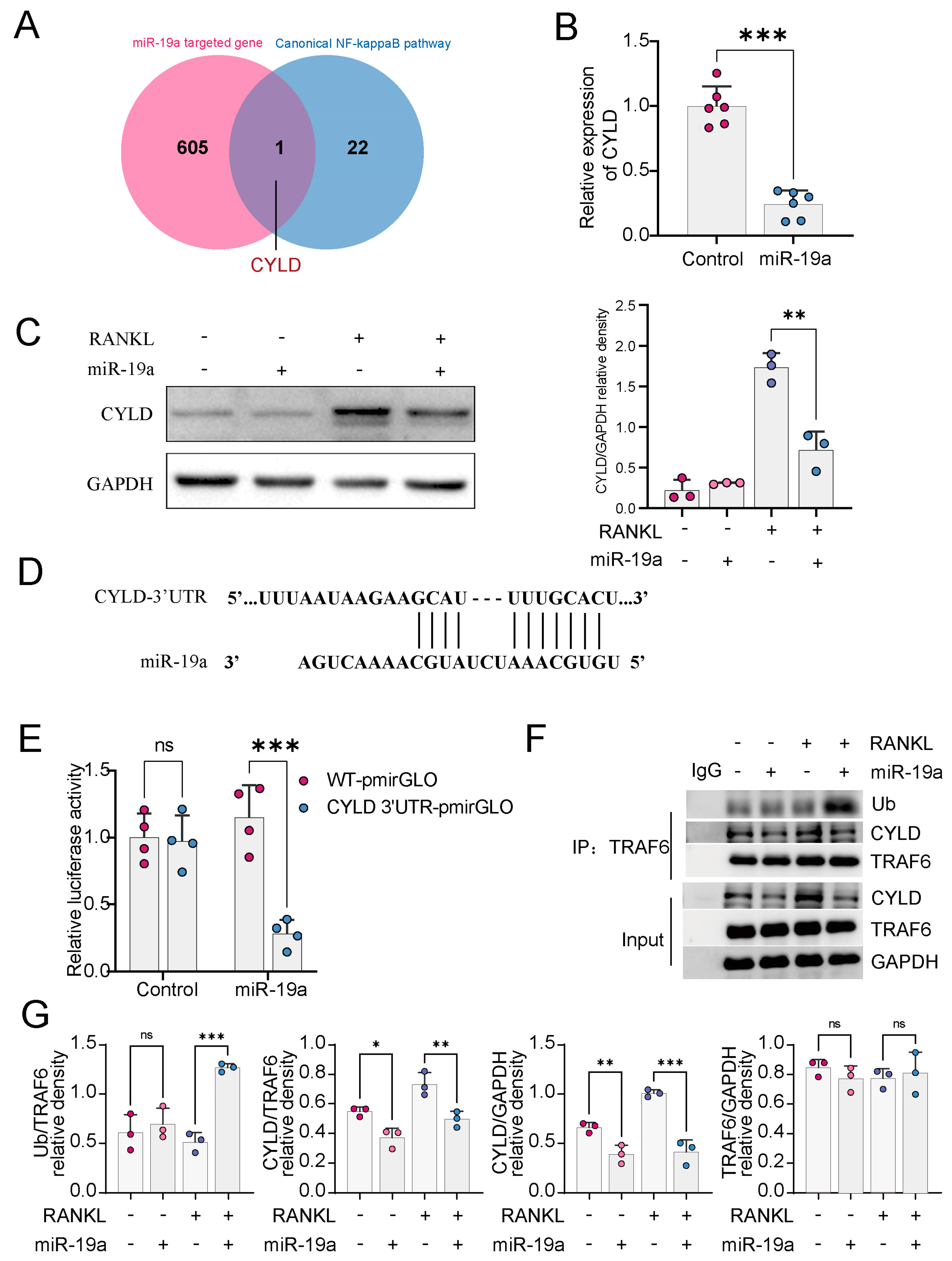
3.3. miR-19a Inhibits CYLD Expression and, Thus, Promotes TRAF6 Ubiquitination
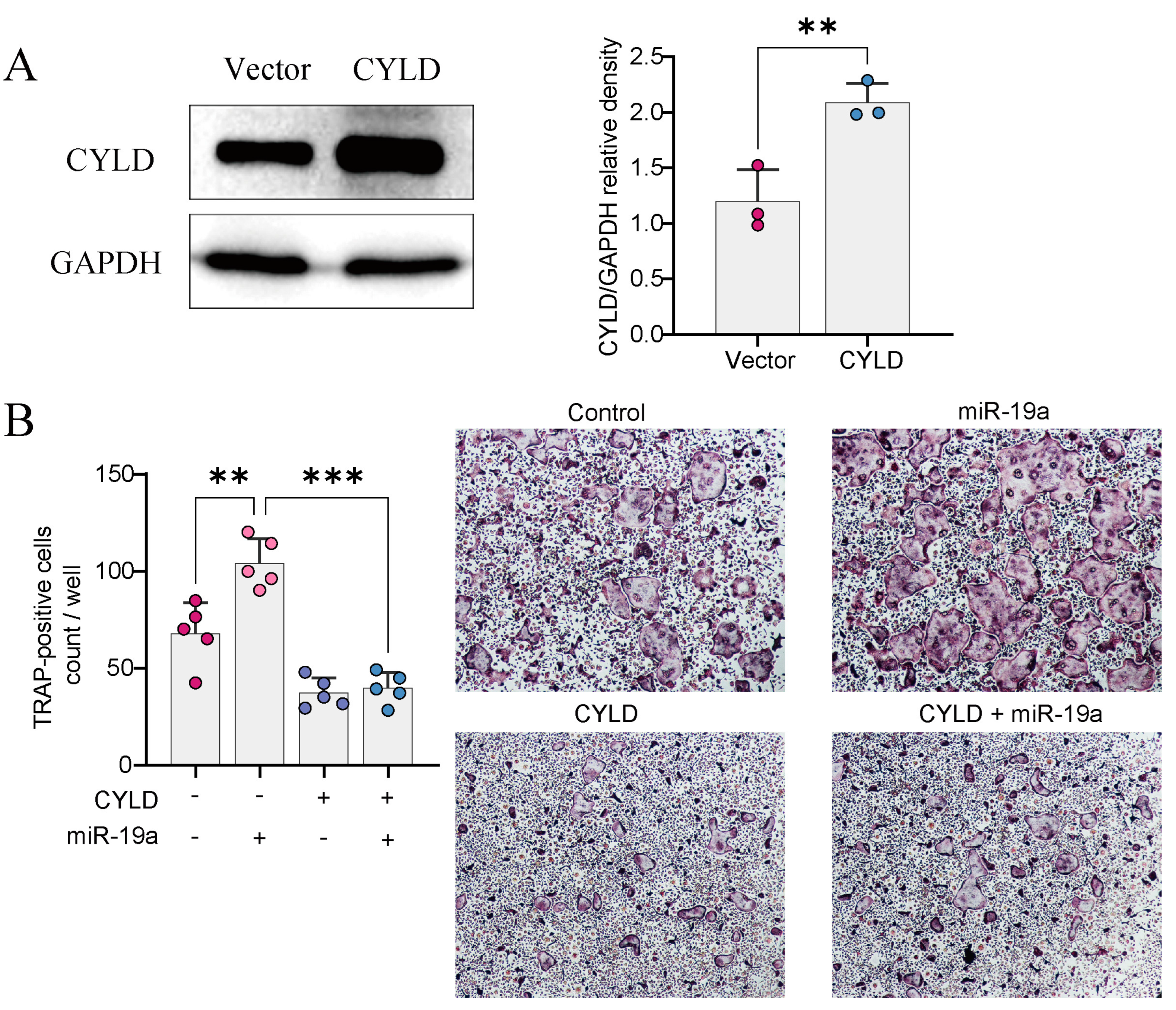
3.4. CYLD Is Required for miRNA-Mediated Regulation of Osteoclast Formation
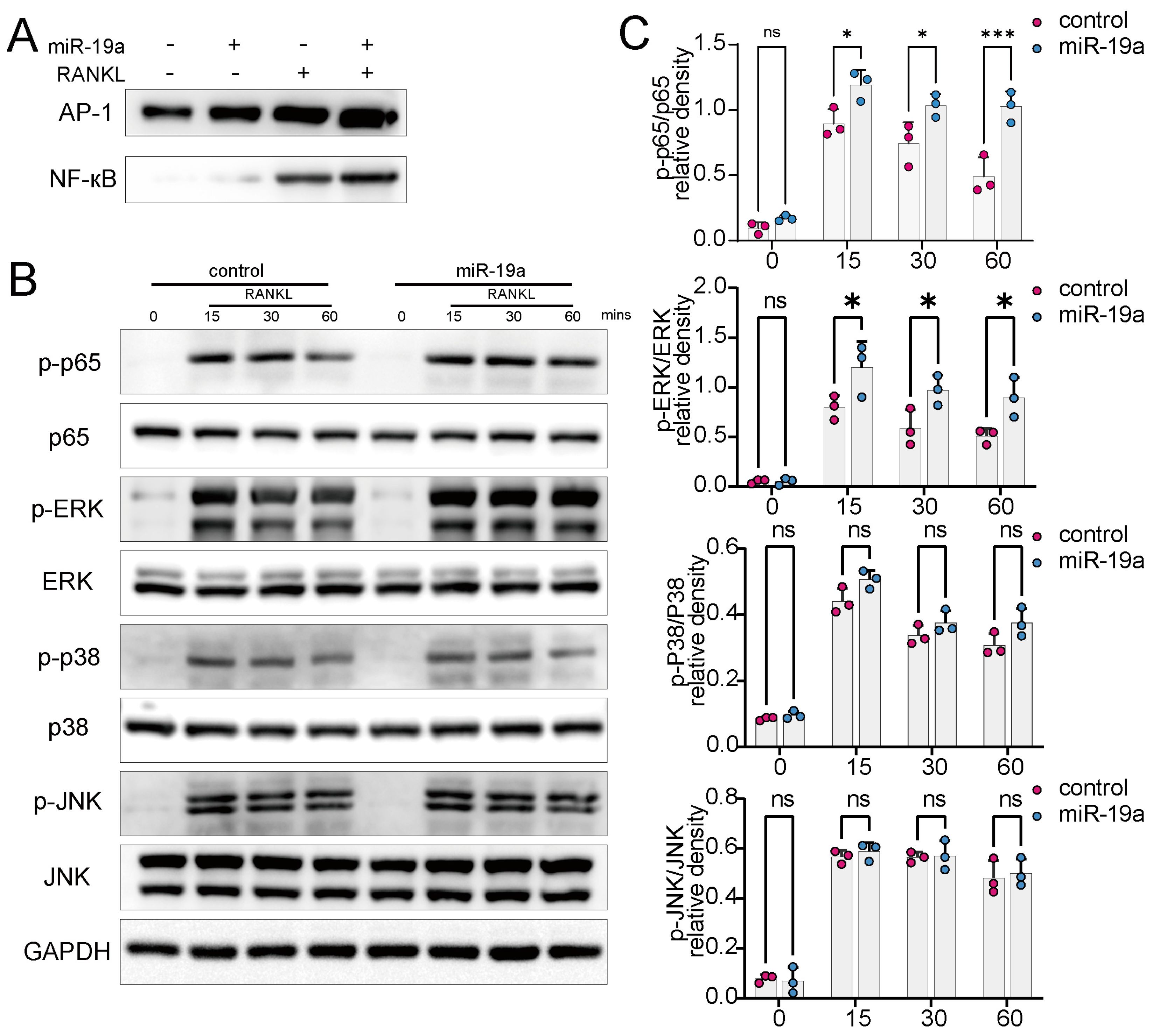
3.5. miR-19a Promotes RANKL-Induced NF-кB and p-ERK Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of bone resorption and formation by RANKL reverse signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Liu, X.; He, Z.; Han, X.; Yan, M.; Qu, X.; Li, X.; Yu, Z. Articular Cartilage Degradation and Aberrant Subchondral Bone Remodeling in Patients with Osteoarthritis and Osteoporosis. J. Bone Miner. Res. 2020, 35, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Emami, A.J.; Toupadakis, C.A.; Telek, S.M.; Fyhrie, D.P.; Yellowley, C.E.; Christiansen, B.A. Age Dependence of Systemic Bone Loss and Recovery Following Femur Fracture in Mice. J. Bone Miner. Res. 2019, 34, 157–170. [Google Scholar] [CrossRef]
- Wang, Q.; Lei, Z.; Wang, Z.; Jiang, Q.; Zhang, Z.; Liu, X.; Xing, B.; Li, S.; Guo, X.; Liu, Y.; et al. PKCtheta Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-kappaB/IL-1beta-Dependent Manner. Cancers 2023, 15, 1624. [Google Scholar] [CrossRef]
- Lu, L.; Wan, X.; Xu, Y.; Chen, J.; Shu, K.; Lei, T. Classifying Pituitary Adenoma Invasiveness Based on Radiological, Surgical and Histological Features: A Retrospective Assessment of 903 Cases. J. Clin. Med. 2022, 11, 2464. [Google Scholar] [CrossRef]
- Kodama, J.; Kaito, T. Osteoclast Multinucleation: Review of Current Literature. Int. J. Mol. Sci. 2020, 21, 5685. [Google Scholar] [CrossRef]
- Kobayashi, N.; Kadono, Y.; Naito, A.; Matsumoto, K.; Yamamoto, T.; Tanaka, S.; Inoue, J. Segregation of TRAF6-mediated signaling pathways clarifies its role in osteoclastogenesis. EMBO J. 2001, 20, 1271–1280. [Google Scholar] [CrossRef]
- Jin, W.; Chang, M.; Paul, E.M.; Babu, G.; Lee, A.J.; Reiley, W.; Wright, A.; Zhang, M.; You, J.; Sun, S.C. Deubiquitinating enzyme CYLD negatively regulates RANK signaling and osteoclastogenesis in mice. J. Clin. Investig. 2008, 118, 1858–1866. [Google Scholar] [CrossRef]
- Lee, S.E.; Woo, K.M.; Kim, S.Y.; Kim, H.M.; Kwack, K.; Lee, Z.H.; Kim, H.H. The phosphatidylinositol 3-kinase, p38, and extracellular signal-regulated kinase pathways are involved in osteoclast differentiation. Bone 2002, 30, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.H.; Kim, H.H. Signal transduction by receptor activator of nuclear factor kappa B in osteoclasts. Biochem. Biophys. Res. Commun. 2003, 305, 211–214. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, N. Signaling Pathways in Osteoclast Differentiation. Chonnam Med. J. 2016, 52, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Nishimura, R.; Matsubara, T.; Tanaka, S.; Inoue, J.; Reddy, S.V.; Hata, K.; Yamashita, K.; Hiraga, T.; Watanabe, T.; et al. Critical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulated osteoclast differentiation. J. Clin. Investig. 2004, 114, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Taipaleenmaki, H.; Saito, H.; Schroder, S.; Maeda, M.; Mettler, R.; Ring, M.; Rollmann, E.; Gasser, A.; Haasper, C.; Gehrke, T.; et al. Antagonizing microRNA-19a/b augments PTH anabolic action and restores bone mass in osteoporosis in mice. EMBO Mol. Med. 2022, 14, e13617. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, B.; Qiu, M.; Huang, Y. MiR-19b-3p accelerates bone loss after spinal cord injury by suppressing osteogenesis via regulating PTEN/Akt/mTOR signalling. J. Cell Mol. Med. 2021, 25, 990–1000. [Google Scholar] [CrossRef]
- Jiang, Q.; Lei, Z.; Wang, Z.; Wang, Q.; Zhang, Z.; Liu, X.; Xing, B.; Li, S.; Guo, X.; Liu, Y.; et al. Tumor-Associated Fibroblast-Derived Exosomal circDennd1b Promotes Pituitary Adenoma Progression by Modulating the miR-145-5p/ONECUT2 Axis and Activating the MAPK Pathway. Cancers 2023, 15, 3375. [Google Scholar] [CrossRef]
- Zhang, Z.; Schäfer, A.; Voellger, B.; Wang, J.-W.; Lei, T.; Nimsky, C.; Bartsch, J.W. MicroRNA-149 Regulates Proliferation, Migration, and Invasion of Pituitary Adenoma Cells by Targeting ADAM12 and MMP14. Curr. Med. Sci. 2022, 42, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Mu, P.; Han, Y.C.; Betel, D.; Yao, E.; Squatrito, M.; Ogrodowski, P.; de Stanchina, E.; D’Andrea, A.; Sander, C.; Ventura, A. Genetic dissection of the miR-17~92 cluster of microRNAs in Myc-induced B-cell lymphomas. Genes. Dev. 2009, 23, 2806–2811. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, M.; Guo, Q.; Lou, J.; Wang, L. Non-small cell lung cancer cell-derived exosomal miR-17-5p promotes osteoclast differentiation by targeting PTEN. Exp. Cell Res. 2021, 408, 112834. [Google Scholar] [CrossRef]
- Wu, K.; Feng, J.; Lyu, F.; Xing, F.; Sharma, S.; Liu, Y.; Wu, S.Y.; Zhao, D.; Tyagi, A.; Deshpande, R.P.; et al. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat. Commun. 2021, 12, 5196. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fang, Z.; Song, C.; Kang, H.; Guo, Q.; Dong, Y.; Zhang, Y.; Peng, R.; Guan, H.; Li, F. Schisandrin B Inhibits Osteoclastogenesis and Protects Against Ovariectomy-Induced Bone Loss. Front. Pharmacol. 2020, 11, 1175. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Tan, P.; Zhang, Z.; Wu, W.; Dong, Y.; Zhao, L.; Liu, H.; Guan, H.; Li, F. REV-ERB agonism suppresses osteoclastogenesis and prevents ovariectomy-induced bone loss partially via FABP4 upregulation. FASEB J. 2018, 32, 3215–3228. [Google Scholar] [CrossRef]
- Lei, Z.; Wang, J.; Sun, W.; Chen, X.; Jiao, W.; Zhang, H.; Lei, T.; Li, F. PKCdelta reveals a tumor promoter function by promoting cell proliferation and migration in somatotropinomas. Int. J. Clin. Exp. Pathol. 2018, 11, 208–215. [Google Scholar]
- Liu, L.; Jin, R.; Duan, J.; Yang, L.; Cai, Z.; Zhu, W.; Nie, Y.; He, J.; Xia, C.; Gong, Q.; et al. Bioactive iron oxide nanoparticles suppress osteoclastogenesis and ovariectomy-induced bone loss through regulating the TRAF6-p62-CYLD signaling complex. Acta Biomater. 2020, 103, 281–292. [Google Scholar] [CrossRef]
- Lin, F.T.; Lin, V.Y.; Lin, V.T.; Lin, W.C. TRIP6 antagonizes the recruitment of A20 and CYLD to TRAF6 to promote the LPA2 receptor-mediated TRAF6 activation. Cell Discov. 2016, 2, 15048. [Google Scholar] [CrossRef]
- Sun, K.T.; Chen, M.Y.; Tu, M.G.; Wang, I.K.; Chang, S.S.; Li, C.Y. MicroRNA-20a regulates autophagy related protein-ATG16L1 in hypoxia-induced osteoclast differentiation. Bone 2015, 73, 145–153. [Google Scholar] [CrossRef]
- Zhao, M.; Dong, J.; Liao, Y.; Lu, G.; Pan, W.; Zhou, H.; Zuo, X.; Shan, B. MicroRNA miR-18a-3p promotes osteoporosis and possibly contributes to spinal fracture by inhibiting the glutamate AMPA receptor subunit 1 gene (GRIA1). Bioengineered 2021, 13, 370–382. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, Y.; Feng, S.; He, P.; Sheng, B.; Ni, J. miR-19b enhances osteogenic differentiation of mesenchymal stem cells and promotes fracture healing through the WWP1/Smurf2-mediated KLF5/beta-catenin signaling pathway. Exp. Mol. Med. 2021, 53, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Xiaoling, G.; Shuaibin, L.; Kailu, L. MicroRNA-19b-3p promotes cell proliferation and osteogenic differentiation of BMSCs by interacting with lncRNA H19. BMC Med. Genet. 2020, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Wa, Q.; Li, L.; Lin, H.; Peng, X.; Ren, D.; Huang, Y.; He, P.; Huang, S. Downregulation of miR-19a-3p promotes invasion, migration and bone metastasis via activating TGF-beta signaling in prostate cancer. Oncol. Rep. 2018, 39, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Brummelkamp, T.R.; Nijman, S.M.; Dirac, A.M.; Bernards, R. Loss of the cylindromatosis tumour suppressor inhibits apoptosis by activating NF-kappaB. Nature 2003, 424, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Herlyn, M.; Yang, X. TRIM15 and CYLD regulate ERK activation via lysine-63-linked polyubiquitination. Nat. Cell Biol. 2021, 23, 978–991. [Google Scholar] [CrossRef]
- Nguyen, J.; Massoumi, R.; Alliston, T. CYLD, a mechanosensitive deubiquitinase, regulates TGFbeta signaling in load-induced bone formation. Bone 2020, 131, 115148. [Google Scholar] [CrossRef]
- Ye, H.; Liu, X.; Lv, M.; Wu, Y.; Kuang, S.; Gong, J.; Yuan, P.; Zhong, Z.; Li, Q.; Jia, H.; et al. MicroRNA and transcription factor co-regulatory network analysis reveals miR-19 inhibits CYLD in T-cell acute lymphoblastic leukemia. Nucleic Acids Res. 2012, 40, 5201–5214. [Google Scholar] [CrossRef]
- Yu, J.; Yun, H.; Shin, B.; Kim, Y.; Park, E.S.; Choi, S.; Yu, J.; Amarasekara, D.S.; Kim, S.; Inoue, J.; et al. Interaction of Tumor Necrosis Factor Receptor-associated Factor 6 (TRAF6) and Vav3 in the Receptor Activator of Nuclear Factor kappaB (RANK) Signaling Complex Enhances Osteoclastogenesis. J. Biol. Chem. 2016, 291, 20643–20660. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Z.; Wang, Q.; Jiang, Q.; Liu, H.; Xu, L.; Kang, H.; Li, F.; Huang, Y.; Lei, T. The miR-19a/Cylindromatosis Axis Regulates Pituitary Adenoma Bone Invasion by Promoting Osteoclast Differentiation. Cancers 2024, 16, 302. https://doi.org/10.3390/cancers16020302
Lei Z, Wang Q, Jiang Q, Liu H, Xu L, Kang H, Li F, Huang Y, Lei T. The miR-19a/Cylindromatosis Axis Regulates Pituitary Adenoma Bone Invasion by Promoting Osteoclast Differentiation. Cancers. 2024; 16(2):302. https://doi.org/10.3390/cancers16020302
Chicago/Turabian StyleLei, Zhuowei, Quanji Wang, Qian Jiang, Huiyong Liu, Linpeng Xu, Honglei Kang, Feng Li, Yimin Huang, and Ting Lei. 2024. "The miR-19a/Cylindromatosis Axis Regulates Pituitary Adenoma Bone Invasion by Promoting Osteoclast Differentiation" Cancers 16, no. 2: 302. https://doi.org/10.3390/cancers16020302
APA StyleLei, Z., Wang, Q., Jiang, Q., Liu, H., Xu, L., Kang, H., Li, F., Huang, Y., & Lei, T. (2024). The miR-19a/Cylindromatosis Axis Regulates Pituitary Adenoma Bone Invasion by Promoting Osteoclast Differentiation. Cancers, 16(2), 302. https://doi.org/10.3390/cancers16020302