

Exercise Promotes Pro-Apoptotic Ceramide Signaling in a Mouse Melanoma Model
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Animal Experimental Protocol
2.3. Tumor Volume Assessment
2.4. Reverse-Phase Protein Array (RPPA)
2.5. Lipid Extraction and Sphingolipid Analysis
2.6. Immunofluorescence Staining
2.7. TUNEL Staining
2.8. Western Blotting
2.9. TNM Plot Analysis
2.10. Statistical Analysis
3. Results
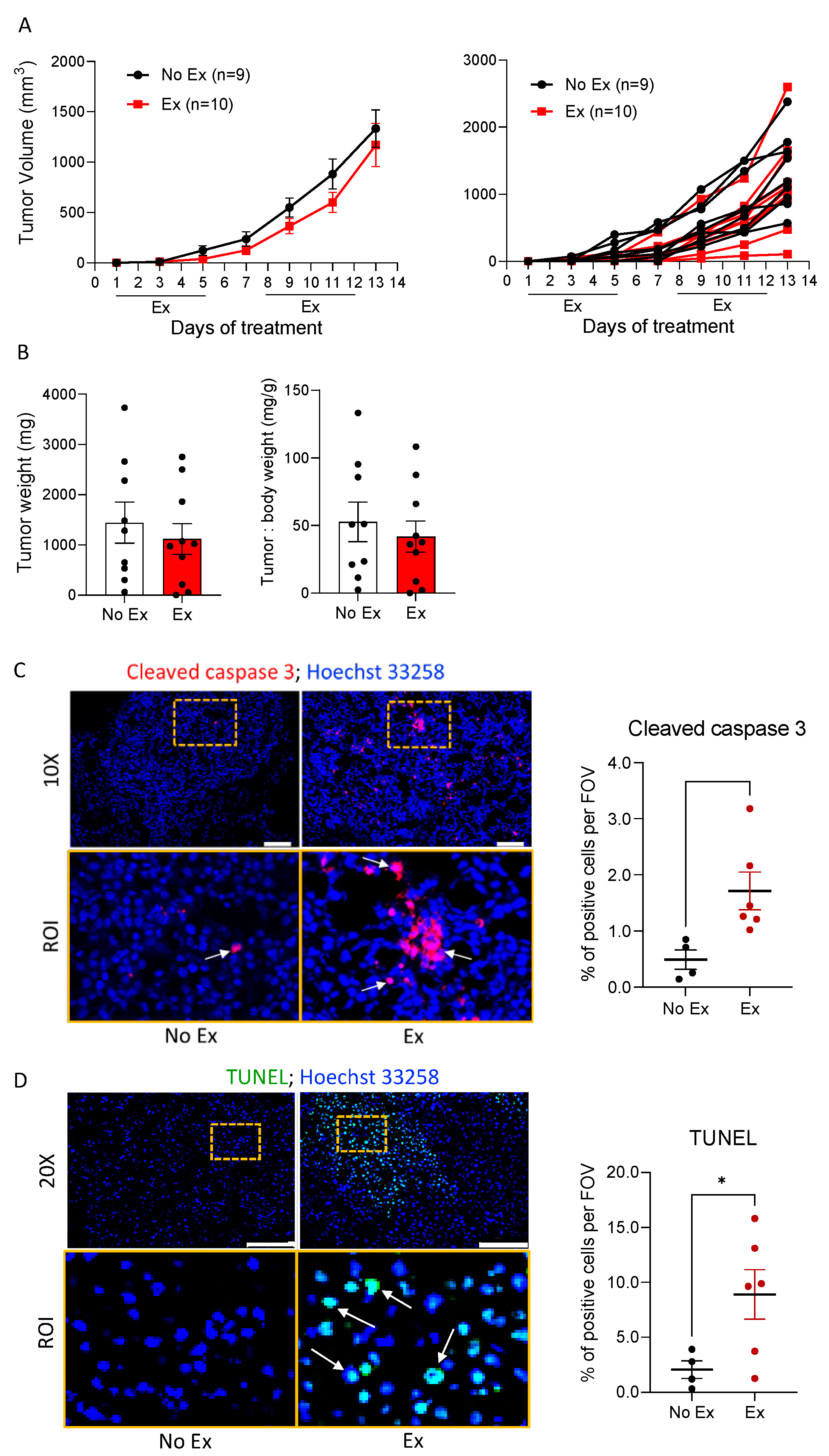
3.1. Exercise Induces Apoptosis in B16F10 Melanoma
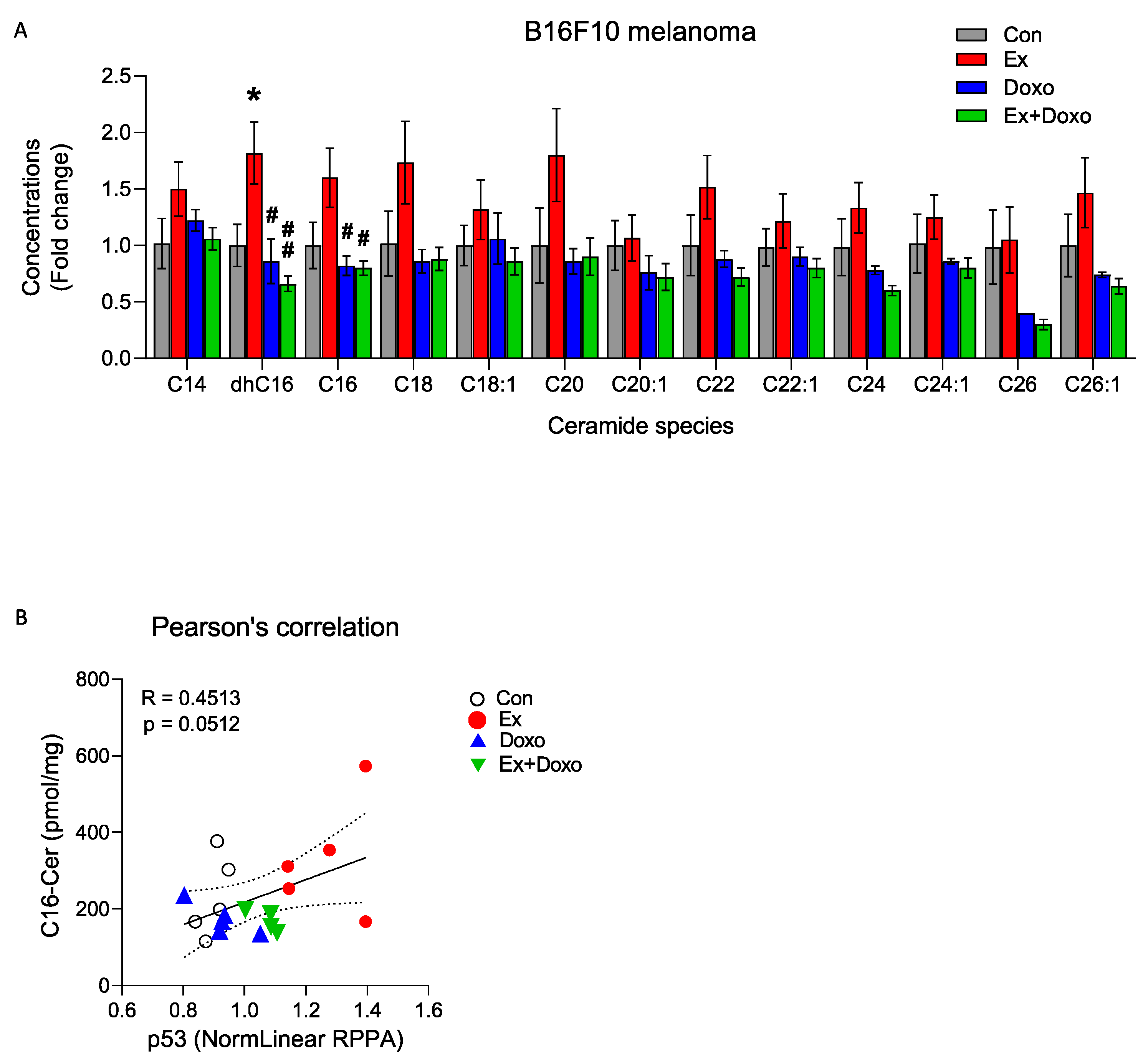
3.2. Multiple Ceramide Species Are Increased within B16F10 Tumors by Exercise
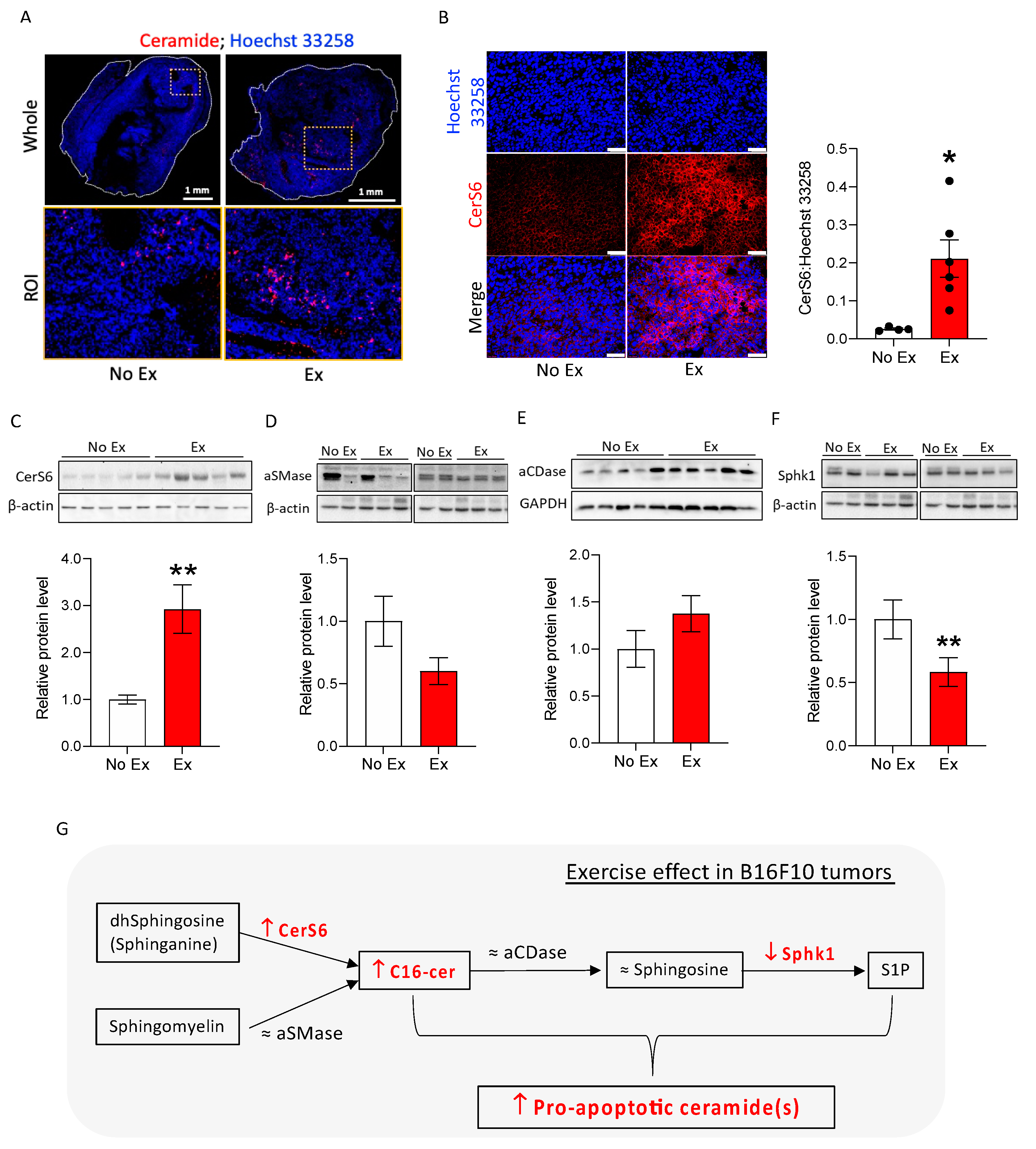
3.3. Exercise Increases Apoptosis-Related Ceramide Accumulation and Upregulates CerS6 and Downregulates Sphk1 in B16F10 Melanoma in Mice
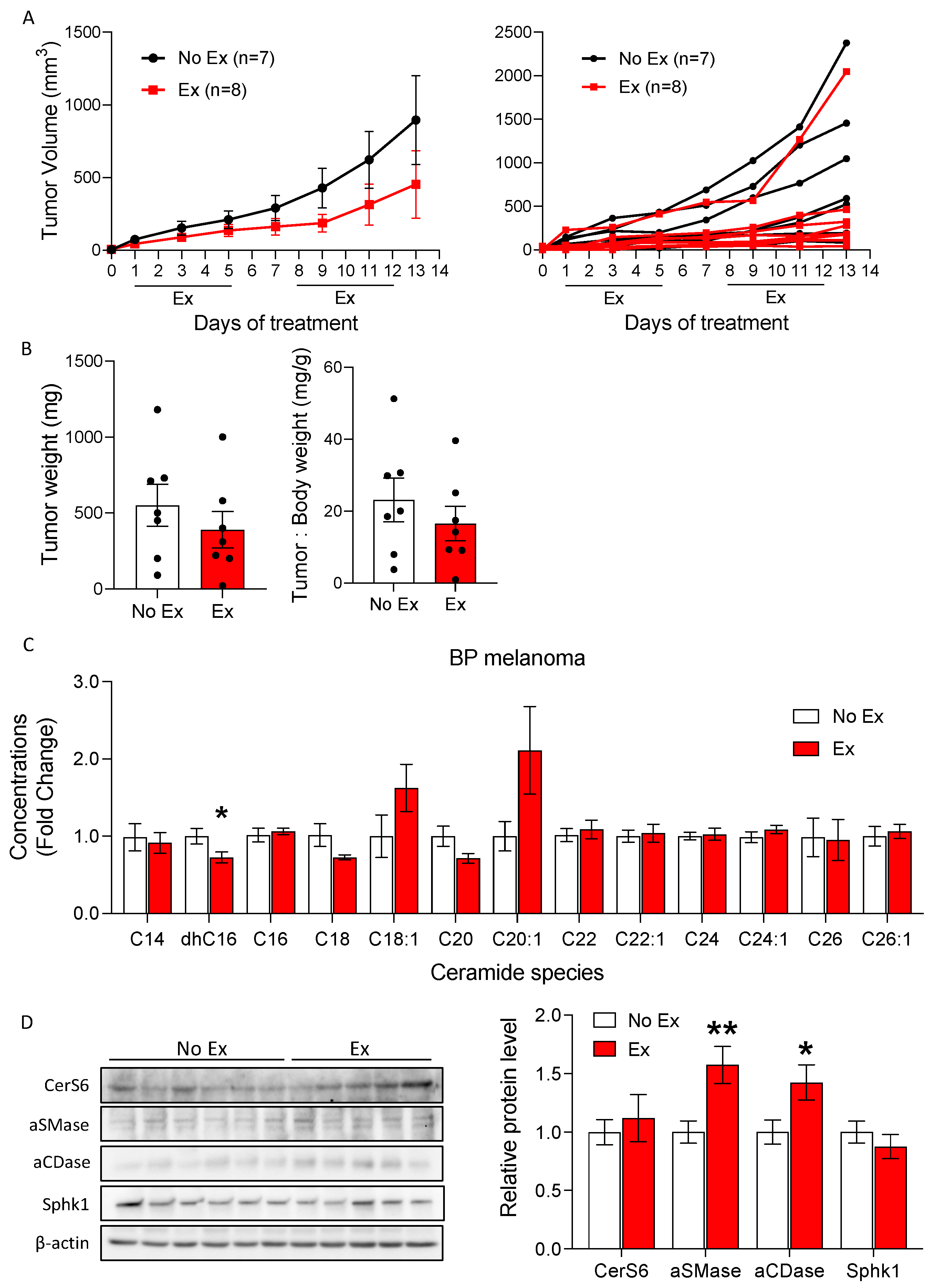
3.4. Exercise Did Not Increase Ceramide Accumulation in the BP Melanoma Model
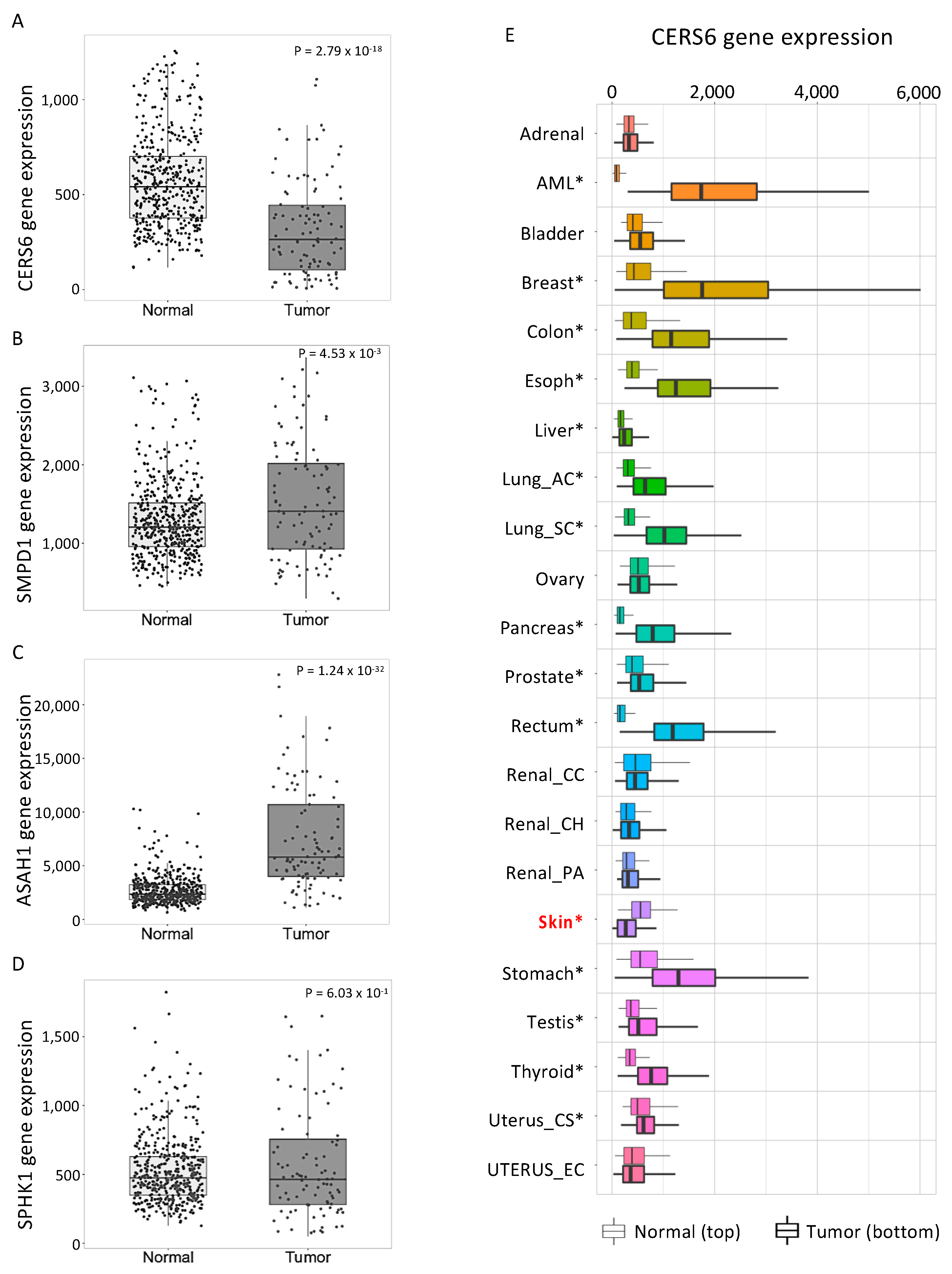
3.5. Human Skin Cancers Have Low CERS6 and High ASAH1 Gene Expression Relative to Normal Skin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brachtendorf, S.; El-Hindi, K.; Grösch, S. Ceramide synthases in cancer therapy and chemoresistance. Prog. Lipid Res. 2019, 74, 160–185. [Google Scholar] [CrossRef] [PubMed]
- Senkal, C.E.; Ponnusamy, S.; Rossi, M.J.; Bialewski, J.; Sinha, D.; Jiang, J.C.; Jazwinski, S.M.; Hannun, Y.A.; Ogretmen, B. Role of human longevity assurance gene 1 and C18-ceramide in chemotherapy-induced cell death in human head and neck squamous cell carcinomas. Mol. Cancer Ther. 2007, 6, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Devalapally, H.; Duan, Z.; Seiden, M.V.; Amiji, M.M. Modulation of drug resistance in ovarian adenocarcinoma by enhancing intracellular ceramide using tamoxifen-loaded biodegradable polymeric nanoparticles. Clin. Cancer Res. 2008, 14, 3193–3203. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, Y.; Liu, Z.; Liu, X.; Chen, L.; Yan, W. Bioactive lipids-based pH sensitive micelles for co-delivery of doxorubicin and ceramide to overcome multidrug resistance in leukemia. Pharm. Res. 2013, 30, 2902–2916. [Google Scholar] [CrossRef]
- Dany, M.; Ogretmen, B. Ceramide induced mitophagy and tumor suppression. Biochim. Biophys. Acta 2015, 1853 Pt B, 2834–2845. [Google Scholar] [CrossRef]
- Li, R.Z.; Wang, X.R.; Wang, J.; Xie, C.; Wang, X.X.; Pan, H.D.; Meng, W.Y.; Liang, T.L.; Li, J.X.; Yan, P.Y.; et al. The key role of sphingolipid metabolism in cancer: New therapeutic targets, diagnostic and prognostic values, and anti-tumor immunotherapy resistance. Front. Oncol. 2022, 12, 941643. [Google Scholar] [CrossRef]
- Sheridan, M.; Ogretmen, B. The Role of Ceramide Metabolism and Signaling in the Regulation of Mitophagy and Cancer Therapy. Cancers 2021, 13, 2475. [Google Scholar] [CrossRef]
- Tang, Y.; Cao, K.; Wang, Q.; Chen, J.; Liu, R.; Wang, S.; Zhou, J.; Xie, H. Silencing of CerS6 increases the invasion and glycolysis of melanoma WM35, WM451 and SK28 cell lines via increased GLUT1-induced downregulation of WNT5A. Oncol. Rep. 2016, 35, 2907–2915. [Google Scholar] [CrossRef]
- Realini, N.; Palese, F.; Pizzirani, D.; Pontis, S.; Basit, A.; Bach, A.; Ganesan, A.; Piomelli, D. Acid Ceramidase in Melanoma: Expression, localization, and effects of pharmacological inhibition. J. Biol. Chem. 2016, 291, 2422–2434. [Google Scholar] [CrossRef]
- Schmitz, K.H.; Campbell, A.M.; Stuiver, M.M.; Pinto, B.M.; Schwartz, A.L.; Morris, G.S.; Ligibel, J.A.; Cheville, A.; Galvão, D.A.; Alfano, C.M.; et al. Exercise is medicine in oncology: Engaging clinicians to help patients move through cancer. CA Cancer J. Clin. 2019, 69, 468–484. [Google Scholar] [CrossRef] [Green Version]
- McTiernan, A.; Friedenreich, C.M.; Katzmarzyk, P.T.; Powell, K.E.; Macko, R.; Buchner, D.; Pescatello, L.S.; Bloodgood, B.; Tennant, B.; Vaux-Bjerke, A.; et al. Physical Activity in Cancer Prevention and Survival: A Systematic Review. Med. Sci. Sports Exerc. 2019, 51, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Schadler, K.L.; Thomas, N.J.; Galie, P.A.; Bhang, D.H.; Roby, K.C.; Addai, P.; Till, J.E.; Sturgeon, K.; Zaslavsky, A.; Chen, C.S.; et al. Tumor vessel normalization after aerobic exercise enhances chemotherapeutic efficacy. Oncotarget 2016, 7, 65429–65440. [Google Scholar] [CrossRef] [PubMed]
- Morrell, M.B.G.; Alvarez-Florez, C.; Zhang, A.; Kleinerman, E.S.; Savage, H.; Marmonti, E.; Park, M.; Shaw, A.; Schadler, K.L. Vascular modulation through exercise improves chemotherapy efficacy in Ewing sarcoma. Pediatr. Blood Cancer 2019, 66, e27835. [Google Scholar] [CrossRef] [PubMed]
- Florez Bedoya, C.A.; Cardoso, A.C.F.; Parker, N.; Ngo-Huang, A.; Petzel, M.Q.; Kim, M.P.; Fogelman, D.; Romero, S.G.; Wang, H.; Park, M.; et al. Exercise during preoperative therapy increases tumor vascularity in pancreatic tumor patients. Sci. Rep. 2019, 9, 13966. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Jonat, W.; Wesch, D.; Oberg, H.H.; Adam-Klages, S.; Keller, L.; Röcken, C.; Mundhenke, C. Influence of physical activity on the immune system in breast cancer patients during chemotherapy. J. Cancer Res. Clin. Oncol 2018, 144, 579–586. [Google Scholar] [CrossRef]
- Wiggins, J.M.; Opoku-Acheampong, A.B.; Baumfalk, D.R.; Siemann, D.W.; Behnke, B.J. Exercise and the Tumor Microenvironment: Potential Therapeutic Implications. Exerc. Sport Sci. Rev. 2018, 46, 56–64. [Google Scholar] [CrossRef]
- Hannah Savage, K.L.S. Mechanisms of Exercise in Cancer Prevention, Treatment, and Survivorship. In Exercise Oncology; Springer: Cham, Switzerland, 2020; pp. 61–83. [Google Scholar]
- Helge, J.W.; Dobrzyn, A.; Saltin, B.; Gorski, J. Exercise and training effects on ceramide metabolism in human skeletal muscle. Exp. Physiol. 2004, 89, 119–127. [Google Scholar] [CrossRef]
- Esteves, M.; Silva, C.; Pereira, S.S.; Morais, T.; Moreira, Â.; Costa, M.M.; Monteiro, M.P.; Duarte, J.A. Regular Voluntary Running Inhibits Androgen-Independent Prostate Cancer Growth in Mice. J. Phys. Act. Health 2021, 18, 653–659. [Google Scholar] [CrossRef]
- Ahmadabadi, F.; Saghebjoo, M.; Hedayati, M.; Hoshyar, R.; Huang, C.J. Treatment-induced tumor cell apoptosis following high-intensity interval training and saffron aqueous extract in mice with breast cancer. Physiol. Int. 2021, 108, 19–26. [Google Scholar] [CrossRef]
- Kim, J.S.; Galvão, D.A.; Newton, R.U.; Gray, E.; Taaffe, D.R. Exercise-induced myokines and their effect on prostate cancer. Nat. Rev. Urol. 2021, 18, 519–542. [Google Scholar] [CrossRef]
- Iadevaia, S.; Lu, Y.; Morales, F.C.; Mills, G.B.; Ram, P.T. Identification of optimal drug combinations targeting cellular networks: Integrating phospho-proteomics and computational network analysis. Cancer Res. 2010, 70, 6704–6714. [Google Scholar] [CrossRef] [PubMed]
- Bielawski, J.; Szulc, Z.M.; Hannun, Y.A.; Bielawska, A. Simultaneous quantitative analysis of bioactive sphingolipids by high-performance liquid chromatography-tandem mass spectrometry. Methods 2006, 39, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Bartha, Á.; Győrffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int J. Mol. Sci 2021, 22, 2622. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.; Amato, R.; La Rocca, V.; Bilgin, M.; Freer, G.; Spezia, P.; Quaranta, P.; Piomelli, D.; Pistello, M. Acid ceramidase controls apoptosis and increases autophagy in human melanoma cells treated with doxorubicin. Sci. Rep. 2021, 11, 11221. [Google Scholar] [CrossRef] [PubMed]
- Senkal, C.E.; Ponnusamy, S.; Manevich, Y.; Meyers-Needham, M.; Saddoughi, S.A.; Mukhopadyay, A.; Dent, P.; Bielawski, J.; Ogretmen, B. Alteration of ceramide synthase 6/C16-ceramide induces activating transcription factor 6-mediated endoplasmic reticulum (ER) stress and apoptosis via perturbation of cellular Ca2+ and ER/Golgi membrane network. J. Biol. Chem. 2011, 286, 42446–42458. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Leonhardt, L.G.; Obeid, L.M. Cell-cycle-dependent changes in ceramide levels preceding retinoblastoma protein dephosphorylation in G2/M. Biochem. J. 1998, 334 Pt 2, 457–461. [Google Scholar] [CrossRef]
- Phillips, D.C.; Hunt, J.T.; Moneypenny, C.G.; Maclean, K.H.; McKenzie, P.P.; Harris, L.C.; Houghton, J.A. Ceramide-induced G2 arrest in rhabdomyosarcoma (RMS) cells requires p21Cip1/Waf1 induction and is prevented by MDM2 overexpression. Cell Death Differ. 2007, 14, 1780–1791. [Google Scholar] [CrossRef]
- Hoeferlin, L.A.; Fekry, B.; Ogretmen, B.; Krupenko, S.A.; Krupenko, N.I. Folate stress induces apoptosis via p53-dependent de novo ceramide synthesis and up-regulation of ceramide synthase 6. J. Biol. Chem. 2013, 288, 12880–12890. [Google Scholar] [CrossRef]
- Fekry, B.; Jeffries, K.A.; Esmaeilniakooshkghazi, A.; Szulc, Z.M.; Knagge, K.J.; Kirchner, D.R.; Horita, D.A.; Krupenko, S.A.; Krupenko, N.I. C(16)-ceramide is a natural regulatory ligand of p53 in cellular stress response. Nat. Commun. 2018, 9, 4149. [Google Scholar] [CrossRef]
- Cooper, Z.A.; Juneja, V.R.; Sage, P.T.; Frederick, D.T.; Piris, A.; Mitra, D.; Lo, J.A.; Hodi, F.S.; Freeman, G.J.; Bosenberg, M.W.; et al. Response to BRAF inhibition in melanoma is enhanced when combined with immune checkpoint blockade. Cancer Immunol. Res. 2014, 2, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Sharpless, N.E.; DePinho, R.A. p53: Good cop/bad cop. Cell 2002, 110, 9–12. [Google Scholar] [CrossRef]
- Fekry, B.; Esmaeilniakooshkghazi, A.; Krupenko, S.A.; Krupenko, N.I. Ceramide Synthase 6 Is a Novel Target of Methotrexate Mediating Its Antiproliferative Effect in a p53-Dependent Manner. PLoS ONE 2016, 11, e0146618. [Google Scholar] [CrossRef] [PubMed]
- Fekry, B.; Jeffries, K.A.; Esmaeilniakooshkghazi, A.; Ogretmen, B.; Krupenko, S.A.; Krupenko, N.I. CerS6 Is a Novel Transcriptional Target of p53 Protein Activated by Non-genotoxic Stress. J. Biol. Chem. 2016, 291, 16586–16596. [Google Scholar] [CrossRef]
- Lu, P.; White-Gilbertson, S.; Beeson, G.; Beeson, C.; Ogretmen, B.; Norris, J.; Voelkel-Johnson, C. Ceramide Synthase 6 Maximizes p53 Function to Prevent Progeny Formation from Polyploid Giant Cancer Cells. Cancers 2021, 13, 2212. [Google Scholar] [CrossRef]
- Yu, M.; King, B.; Ewert, E.; Su, X.; Mardiyati, N.; Zhao, Z.; Wang, W. Exercise Activates p53 and Negatively Regulates IGF-1 Pathway in Epidermis within a Skin Cancer Model. PLoS ONE 2016, 11, e0160939. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S.; Aronson, W.J.; Ngo, T.H.; Golding, L.A.; Barnard, R.J. Exercise alters the IGF axis in vivo and increases p53 protein in prostate tumor cells in vitro. J. Appl. Physiol. (1985) 2004, 96, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, K.; Schadler, K.; Muthukumaran, G.; Ding, D.; Bajulaiye, A.; Thomas, N.J.; Ferrari, V.; Ryeom, S.; Libonati, J.R. Concomitant low-dose doxorubicin treatment and exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R685-92. [Google Scholar] [CrossRef]
- White-Gilbertson, S.; Mullen, T.; Senkal, C.; Lu, P.; Ogretmen, B.; Obeid, L.; Voelkel-Johnson, C. Ceramide synthase 6 modulates TRAIL sensitivity and nuclear translocation of active caspase-3 in colon cancer cells. Oncogene 2009, 28, 1132–1141. [Google Scholar] [CrossRef]
- Lee, H.; Rotolo, J.A.; Mesicek, J.; Penate-Medina, T.; Rimner, A.; Liao, W.C.; Yin, X.; Ragupathi, G.; Ehleiter, D.; Gulbins, E.; et al. Mitochondrial ceramide-rich macrodomains functionalize Bax upon irradiation. PLoS ONE 2011, 6, e19783. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ceramide | Con (n = 6) | Ex (n = 6) | Doxo (n = 5) | Ex + Doxo (n = 5) |
|---|---|---|---|---|
| C14 | 5.7 ± 1.3 | 8.6 ± 1.4 | 6.9 ± 0.5 | 6.1 ± 0.6 |
| dhC16 | 24.5 ± 4.6 | 44.7 ± 6.7 * | 21.4 ± 5.0 # | 16.3 ± 1.8 ## |
| C16 | 213.0 ± 43.1 | 336.0 ± 55.7 | 172.8 ± 17.9 # | 171.9 ± 11.2 # |
| C18 | 62.5 ± 18.0 | 108.6 ± 23.1 | 54.4 ± 5.6 # | 53.7 ± 6.3 # |
| C18:1 | 3.8 ± 0.7 | 5.0 ± 1.0 | 4.0 ± 0.9 | 3.2 ± 0.5 |
| C20 | 91.6 ± 29.5 | 165.0 ± 37.2 | 77.6 ± 10.9 | 82.0 ± 14.9 |
| C20:1 | 1.9 ± 0.4 | 2.0 ± 0.4 | 1.4 ± 0.3 | 1.4 ± 0.2 |
| C22 | 262.0 ± 71.1 | 390.2 ± 72.2 | 229.9 ± 21.6 | 197.1 ± 20.3 |
| C22:1 | 24.7 ± 4.3 | 29.5 ± 6.0 | 22.5 ± 2.0 | 19.8 ± 1.8 |
| C24 | 633.3 ± 161.8 | 841.7 ± 142.0 | 481.2 ± 24.0 | 384.7 ± 38.2 |
| C24:1 | 456.7 ± 118.1 | 571.6 ± 86.5 | 396.3 ± 17.5 | 365.7 ± 43.0 |
| C26 | 16.0 ± 5.2 | 17.0 ± 4.6 | 6.4 ± 0.2 | 4.6 ± 0.5 |
| C26:1 | 8.7 ± 2.4 | 12.7 ± 2.6 | 6.4 ± 0.2 | 5.6 ± 0.6 |
| Sum | 1804.3 ± 441.4 | 2532.5 ± 431.4 | 1481.0 ± 58.5 | 1312.1 ± 130.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Savage, H.; Maegawa, S.; Ballarò, R.; Pareek, S.; Guerrouahen, B.S.; Gopalakrishnan, V.; Schadler, K. Exercise Promotes Pro-Apoptotic Ceramide Signaling in a Mouse Melanoma Model. Cancers 2022, 14, 4306. https://doi.org/10.3390/cancers14174306
Lee J, Savage H, Maegawa S, Ballarò R, Pareek S, Guerrouahen BS, Gopalakrishnan V, Schadler K. Exercise Promotes Pro-Apoptotic Ceramide Signaling in a Mouse Melanoma Model. Cancers. 2022; 14(17):4306. https://doi.org/10.3390/cancers14174306
Chicago/Turabian StyleLee, Jonghae, Hannah Savage, Shinji Maegawa, Riccardo Ballarò, Sumedha Pareek, Bella Samia Guerrouahen, Vidya Gopalakrishnan, and Keri Schadler. 2022. "Exercise Promotes Pro-Apoptotic Ceramide Signaling in a Mouse Melanoma Model" Cancers 14, no. 17: 4306. https://doi.org/10.3390/cancers14174306
APA StyleLee, J., Savage, H., Maegawa, S., Ballarò, R., Pareek, S., Guerrouahen, B. S., Gopalakrishnan, V., & Schadler, K. (2022). Exercise Promotes Pro-Apoptotic Ceramide Signaling in a Mouse Melanoma Model. Cancers, 14(17), 4306. https://doi.org/10.3390/cancers14174306