Roles of Sorcin in Drug Resistance in Cancer: One Protein, Many Mechanisms, for a Novel Potential Anticancer Drug Target

 , and
, and
Abstract
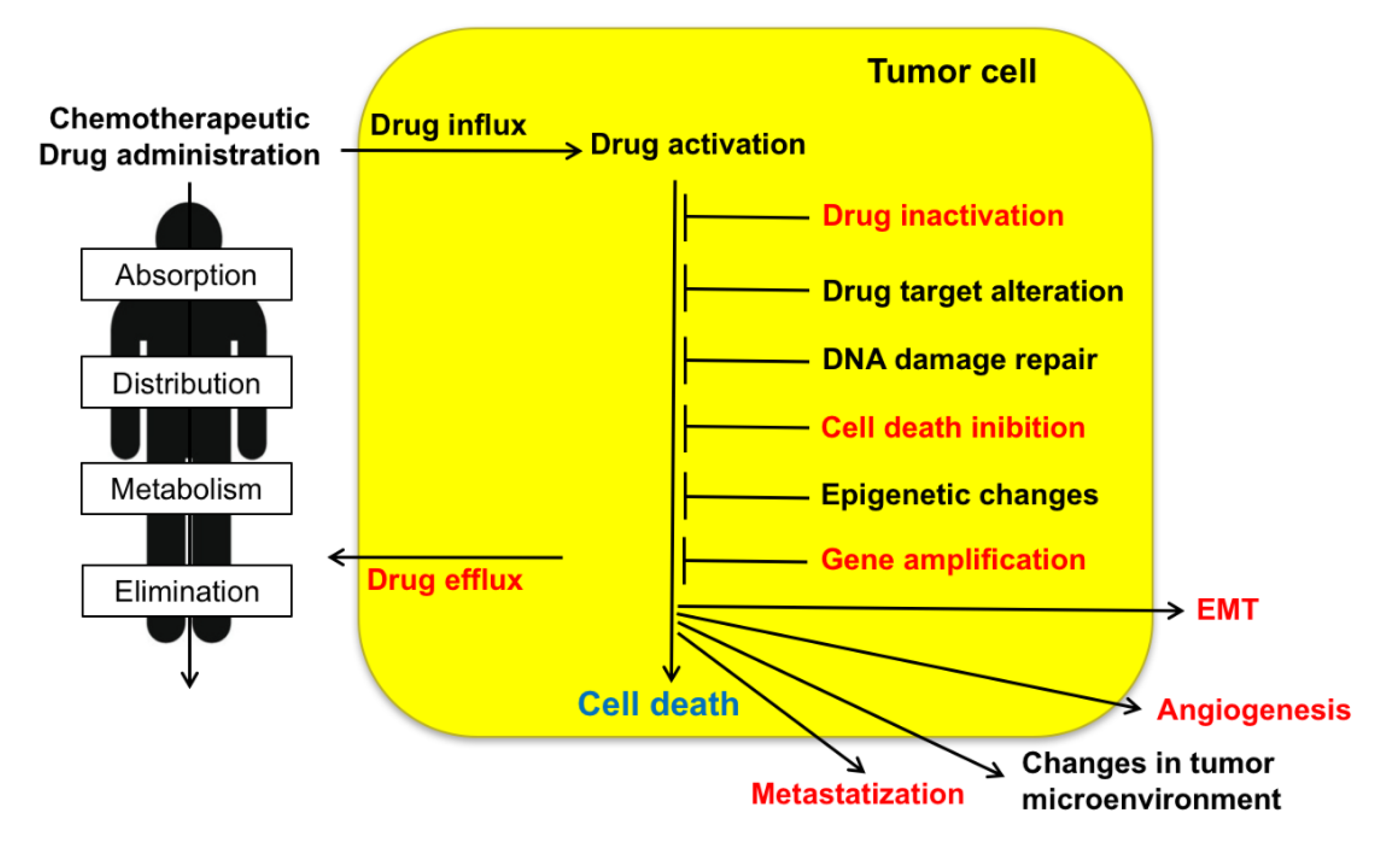
1. Introduction
2. Role and Mode of Action of Sorcin in Physiological and Pathological Processes
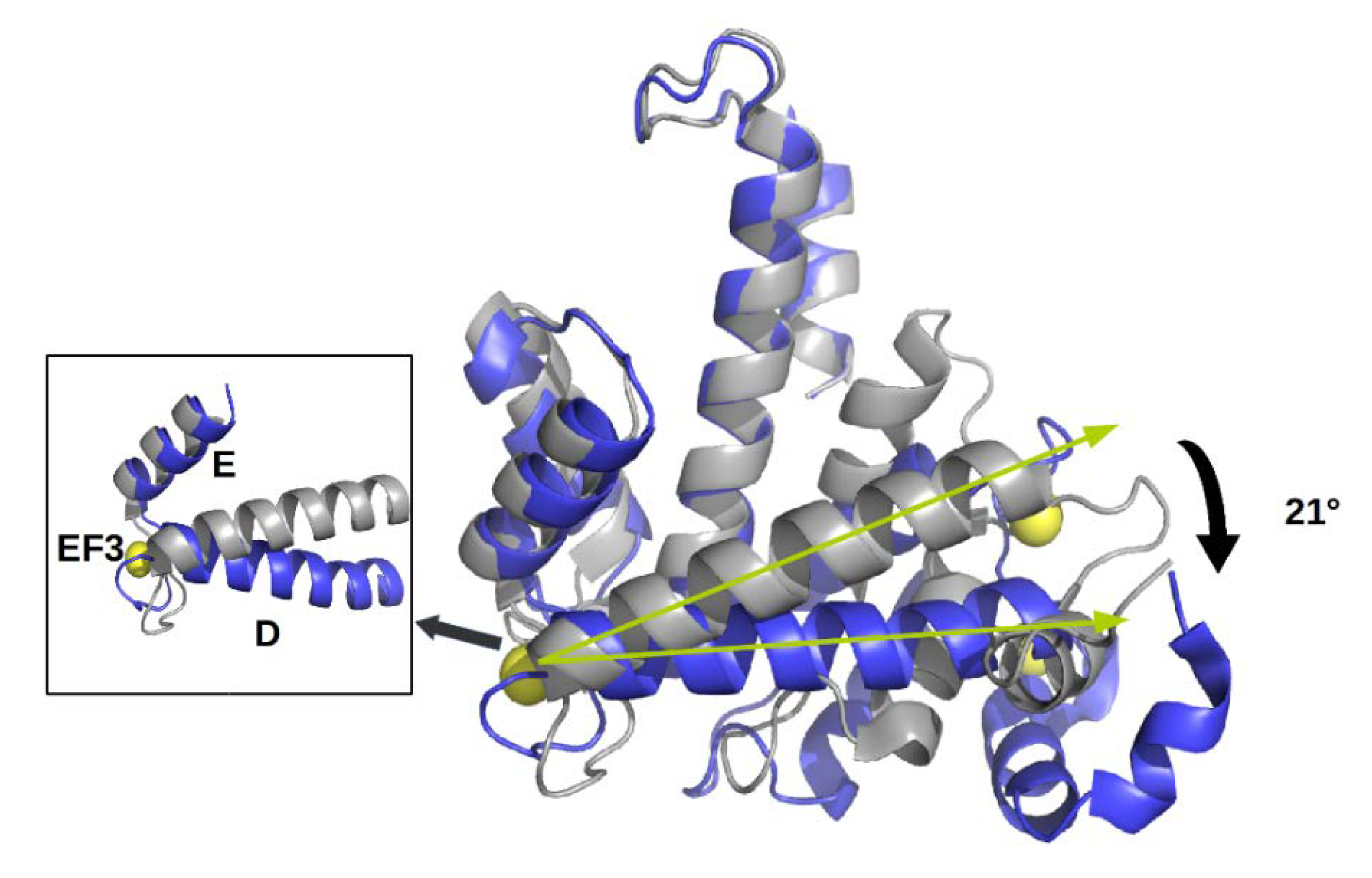
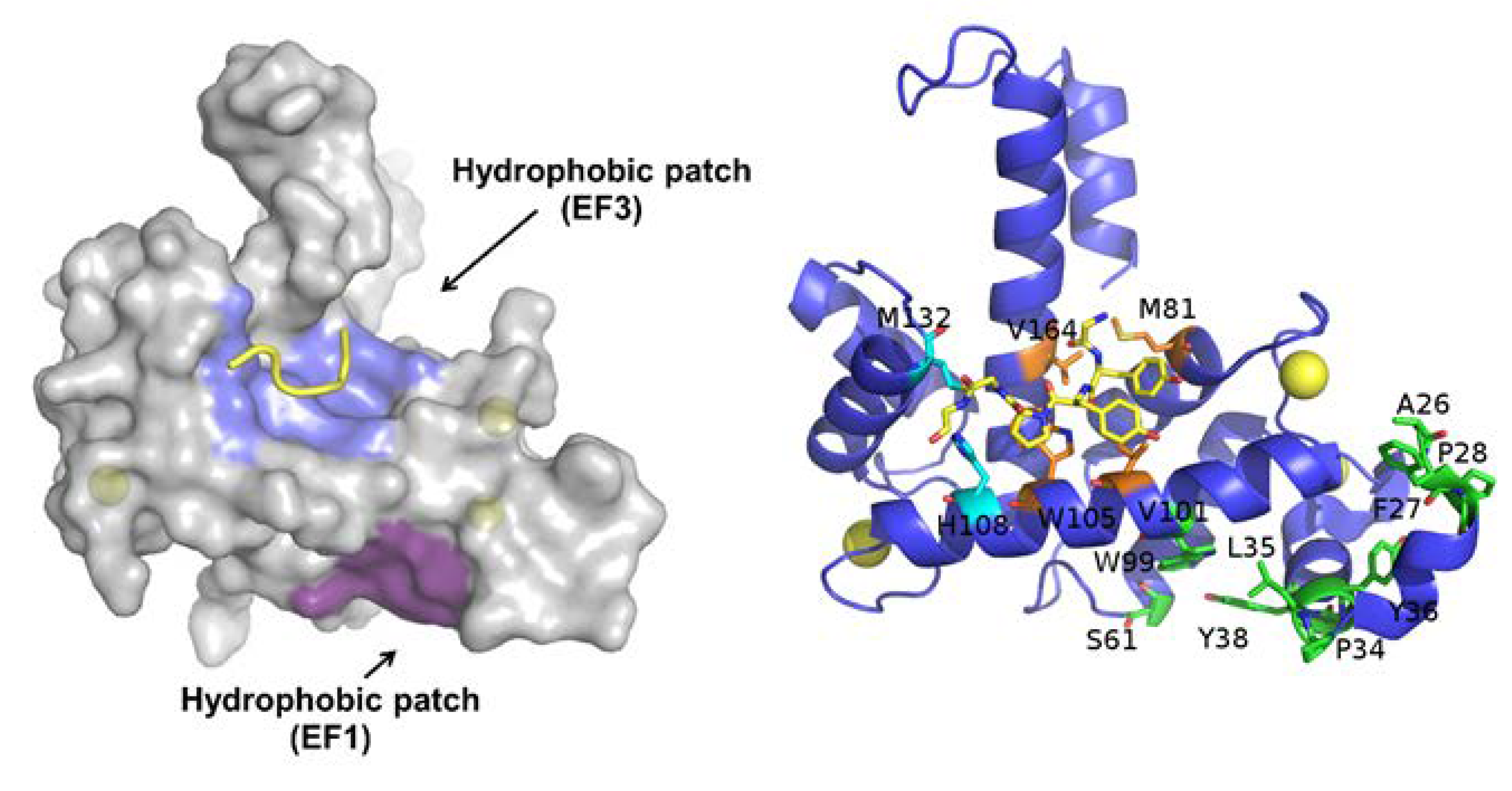
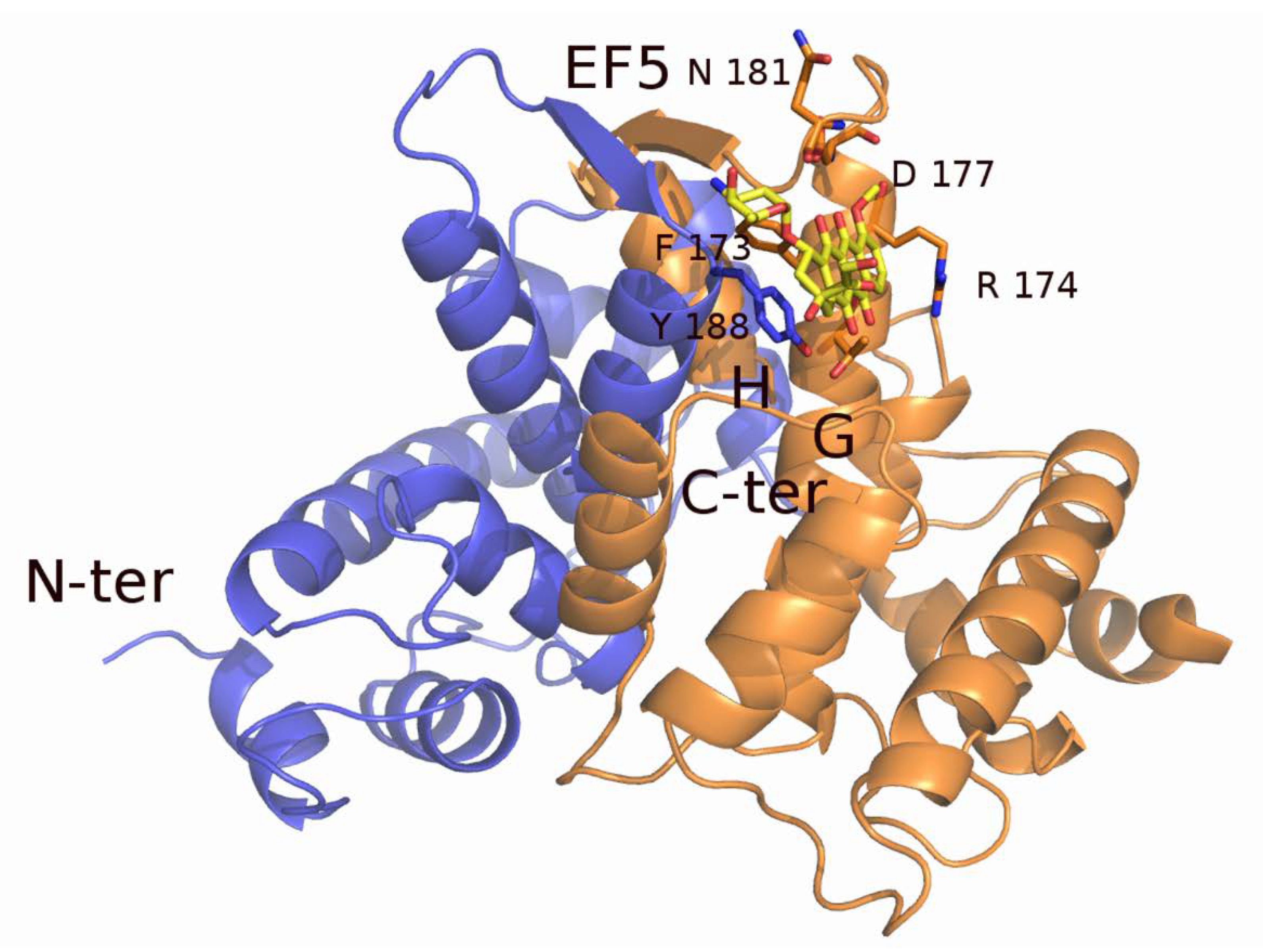
2.1. Sorcin Structure and Calcium-dependent Activation
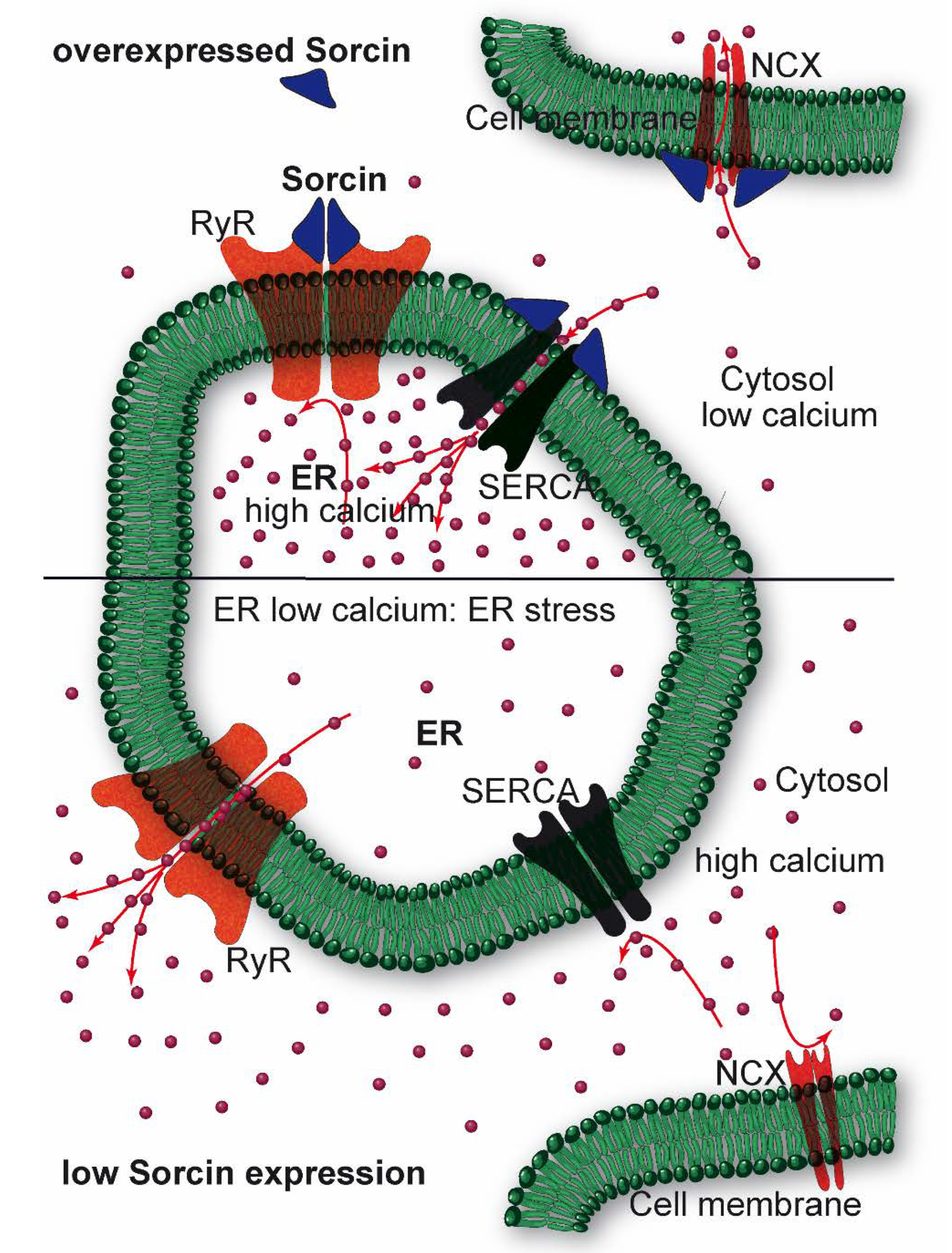
2.2. Sorcin Mechanisms of Action: Role in Calcium Homeostasis, ER Stress, and Apoptosis
2.3. Sorcin Under Cellular Stressing Conditions and in Pathologies
2.4. Sorcin in Cancer and Multidrug (MD)-resistant Tumors
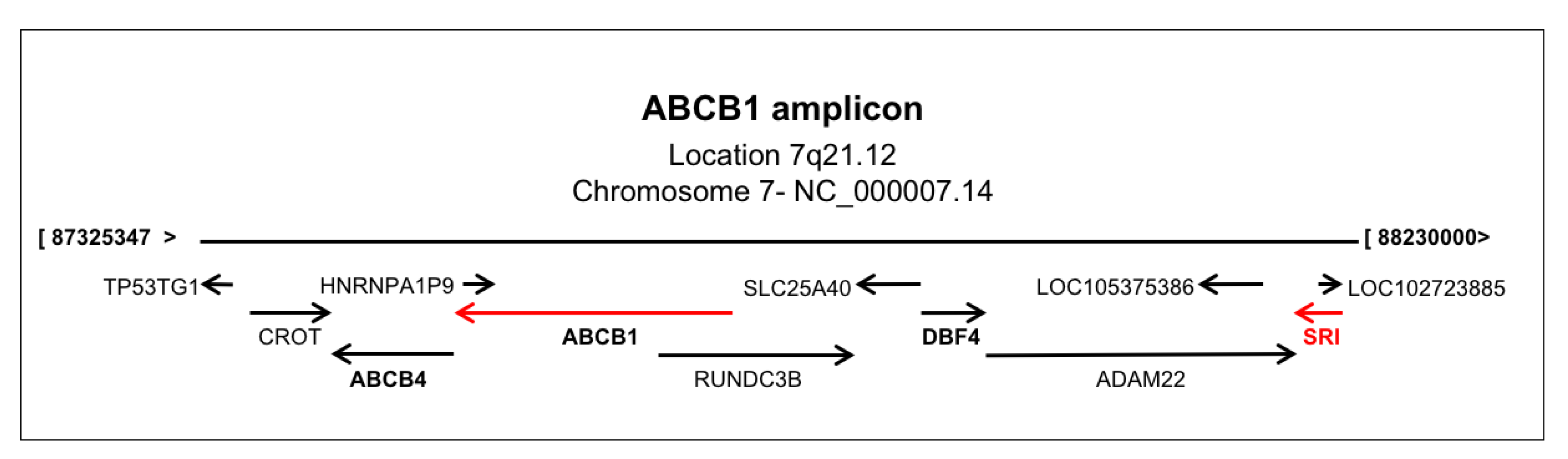
2.5. Sorcin Expression and ABCB1 Expression are Linked
2.6. Sorcin, Metastatization, and EMT
2.7. Sorcin Directly Binds Chemotherapeutic Drugs
2.8. Targeting Sorcin
3. Conclusions
Funding
Conflicts of Interest
References
- Colotti, G.; Poser, E.; Fiorillo, A.; Genovese, I.; Chiarini, V.; Ilari, A. Sorcin, a calcium binding protein involved in the multidrug resistance mechanisms in cancer cells. Molecules 2014, 19, 13976–13989. [Google Scholar] [CrossRef] [PubMed]
- Genovese, I.; Ilari, A.; Battista, T.; Chiarini, F.; Fiorillo, A.; Colotti, G. Molecular bases of Sorcin-dependent resistance to chemotherapeutic agents. Cancer Drug Resist. 2018, 1, 17. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: an overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed]
- Zamparelli, C.; Ilari, A.; Verzili, D.; Giangiacomo, L.; Colotti, G.; Pascarella, S.; Chiancone, E. Structure-function relationships in sorcin, a member of the penta EF-hand family. Interaction of sorcin fragments with the ryanodine receptor and an Escherichia coli model system. Biochemistry 2000, 39, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Colotti, G.; Zamparelli, C.; Verzili, D.; Mella, M.; Loughrey, C.M.; Smith, G.L.; Chiancone, E. The W105G and W99G sorcin mutants demonstrate the role of the D helix in the Ca(2+)-dependent interaction with annexin VII and the cardiac ryanodine receptor. Biochemistry 2006, 45, 12519–12529. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, S.; Ilari, A.; Verzili, D.; Zamparelli, C.; Antaramian, A.; Rueda, A.; Valdivia, H.H.; Chiancone, E.; Colotti, G. Molecular basis for the impaired function of the natural F112L sorcin mutant: X-ray crystal structure, calcium affinity, and interaction with annexin VII and the ryanodine receptor. FASEB J. 2008, 22, 295–306. [Google Scholar] [CrossRef]
- Meyers, M.B.; Pickel, V.M.; Sheu, S.S.; Sharma, V.K.; Scotto, K.W.; Fishman, G.I. Association of sorcin with the cardiac ryanodine receptor. J. Biol. Chem. 1995, 270, 26411–26418. [Google Scholar] [CrossRef]
- Zamparelli, C.; Macquaide, N.; Colotti, G.; Verzili, D.; Seidler, T.; Smith, G.L.; Chiancone, E. Activation of the cardiac Na(+)-Ca(2+) exchanger by sorcin via the interaction of the respective Ca(2+)-binding domains. J. Mol. Cell. Cardiol. 2010, 49, 132–141. [Google Scholar] [CrossRef]
- Farrell, E.F.; Antaramian, A.; Rueda, A.; Gomez, A.M.; Valdivia, H.H. Sorcin inhibits calcium release and modulates excitation-contraction coupling in the heart. J. Biol. Chem. 2003, 278, 34660–34666. [Google Scholar] [CrossRef]
- Lokuta, A.J.; Meyers, M.B.; Sander, P.R.; Fishman, G.I.; Valdivia, H.H. Modulation of cardiac ryanodine receptors by sorcin. J. Biol. Chem. 1997, 272, 25333–25338. [Google Scholar] [CrossRef]
- Matsumoto, T.; Hisamatsu, Y.; Ohkusa, T.; Inoue, N.; Sato, T.; Suzuki, S.; Ikeda, Y.; Matsuzaki, M. Sorcin interacts with sarcoplasmic reticulum Ca(2+)-ATPase and modulates excitation-contraction coupling in the heart. Basic Res. Cardiol. 2005, 100, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.R.; Colotti, G.; Chiancone, E.; Higuchi, Y.; Seidler, T.; Smith, G.L. Complex modulation of L-type Ca(2+) current inactivation by sorcin in isolated rabbit cardiomyocytes. Pflug. Arch. 2009, 457, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Fowler, M.R.; Colotti, G.; Chiancone, E.; Smith, G.L.; Fearon, I.M. Sorcin modulates cardiac L-type Ca2+ current by functional interaction with the alpha1C subunit in rabbits. Exp. Physiol. 2008, 93, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Frank, K.F.; Bolck, B.; Ding, Z.; Krause, D.; Hattebuhr, N.; Malik, A.; Brixius, K.; Hajjar, R.J.; Schrader, J.; Schwinger, R.H. Overexpression of sorcin enhances cardiac contractility in vivo and in vitro. J. Mol. Cell. Cardiol. 2005, 38, 607–615. [Google Scholar] [CrossRef]
- Meyers, M.B.; Fischer, A.; Sun, Y.J.; Lopes, C.M.; Rohacs, T.; Nakamura, T.Y.; Zhou, Y.Y.; Lee, P.C.; Altschuld, R.A.; McCune, S.A.; et al. Sorcin regulates excitation-contraction coupling in the heart. J. Biol. Chem. 2003, 278, 28865–28871. [Google Scholar] [CrossRef]
- Seidler, T.; Miller, S.L.; Loughrey, C.M.; Kania, A.; Burow, A.; Kettlewell, S.; Teucher, N.; Wagner, S.; Kogler, H.; Meyers, M.B.; et al. Effects of adenovirus-mediated sorcin overexpression on excitation-contraction coupling in isolated rabbit cardiomyocytes. Circ. Res. 2003, 93, 132–139. [Google Scholar] [CrossRef]
- Suarez, J.; McDonough, P.M.; Scott, B.T.; Suarez-Ramirez, A.; Wang, H.; Fricovsky, E.S.; Dillmann, W.H. Sorcin modulates mitochondrial Ca(2+) handling and reduces apoptosis in neonatal rat cardiac myocytes. Am. J. Physiol. Cell Physiol. 2013, 304, C248–C256. [Google Scholar] [CrossRef]
- Chen, X.; Weber, C.; Farrell, E.T.; Alvarado, F.J.; Zhao, Y.T.; Gomez, A.M.; Valdivia, H.H. Sorcin ablation plus beta-adrenergic stimulation generate an arrhythmogenic substrate in mouse ventricular myocytes. J. Mol. Cell. Cardiol. 2018, 114, 199–210. [Google Scholar] [CrossRef]
- Ranum, P.T.; Goodwin, A.T.; Yoshimura, H.; Kolbe, D.L.; Walls, W.D.; Koh, J.Y.; He, D.Z.Z.; Smith, R.J.H. Insights into the Biology of Hearing and Deafness Revealed by Single-Cell RNA Sequencing. Cell Rep. 2019, 26, 3160–3171. [Google Scholar] [CrossRef]
- Padar, S.; van Breemen, C.; Thomas, D.W.; Uchizono, J.A.; Livesey, J.C.; Rahimian, R. Differential regulation of calcium homeostasis in adenocarcinoma cell line A549 and its Taxol-resistant subclone. Br. J. Pharmacol. 2004, 142, 305–316. [Google Scholar] [CrossRef]
- Takeda, T.; Asahi, M.; Yamaguchi, O.; Hikoso, S.; Nakayama, H.; Kusakari, Y.; Kawai, M.; Hongo, K.; Higuchi, Y.; Kashiwase, K.; et al. Presenilin 2 regulates the systolic function of heart by modulating Ca2+ signaling. FASEB J. 2005, 19, 2069–2071. [Google Scholar] [CrossRef] [PubMed]
- Lalioti, V.S.; Ilari, A.; O’Connell, D.J.; Poser, E.; Sandoval, I.V.; Colotti, G. Sorcin links calcium signaling to vesicle trafficking, regulates Polo-like kinase 1 and is necessary for mitosis. PloS ONE 2014, 9, e85438. [Google Scholar] [CrossRef] [PubMed]
- Bouchelouche, P.; Friche, E.; Sehested, M.; Jensen, P.B.; Skovsgaard, T. Cytosolic free Ca2+ in daunorubicin and vincristine resistant Ehrlich ascites tumor cells. Drug accumulation is independent of intracellular Ca2+ changes. Biochem. Pharmacol. 1991, 41, 243–253. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Meng, Q.; Liu, Z.; Huo, X.; Sun, P.; Sun, H.; Ma, X.; Peng, J.; Liu, K. Targeting P-glycoprotein and SORCIN: Dihydromyricetin strengthens anti-proliferative efficiency of adriamycin via MAPK/ERK and Ca(2+) -mediated apoptosis pathways in MCF-7/ADR and K562/ADR. J. Cell. Physiol. 2018, 233, 3066–3079. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Liu, N.; Zhou, Y.; Tan, Y.; Cheng, Y.; Yang, C.; Zhu, Z.; Xiong, D. Overexpression of sorcin in multidrug resistant human leukemia cells and its role in regulating cell apoptosis. Biochem. Biophys. Res. Commun. 2006, 349, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Maddalena, F.; Laudiero, G.; Piscazzi, A.; Secondo, A.; Scorziello, A.; Lombardi, V.; Matassa, D.S.; Fersini, A.; Neri, V.; Esposito, F.; et al. Sorcin induces a drug-resistant phenotype in human colorectal cancer by modulating Ca(2+) homeostasis. Cancer Res. 2011, 71, 7659–7669. [Google Scholar] [CrossRef]
- Landriscina, M.; Laudiero, G.; Maddalena, F.; Amoroso, M.R.; Piscazzi, A.; Cozzolino, F.; Monti, M.; Garbi, C.; Fersini, A.; Pucci, P.; et al. Mitochondrial chaperone Trap1 and the calcium binding protein Sorcin interact and protect cells against apoptosis induced by antiblastic agents. Cancer Res. 2010, 70, 6577–6586. [Google Scholar] [CrossRef]
- Marmugi, A.; Parnis, J.; Chen, X.; Carmichael, L.; Hardy, J.; Mannan, N.; Marchetti, P.; Piemonti, L.; Bosco, D.; Johnson, P.; et al. Sorcin links pancreatic beta cell lipotoxicity to ER Ca2+ stores. Diabetes 2016, 65, 1009–1021. [Google Scholar] [CrossRef]
- Ilari, A.; Fiorillo, A.; Poser, E.; Lalioti, V.S.; Sundell, G.N.; Ivarsson, Y.; Genovese, I.; Colotti, G. Structural basis of Sorcin-mediated calcium-dependent signal transduction. Sci. Rep. 2015, 5, 16828. [Google Scholar] [CrossRef]
- Ilari, A.; Johnson, K.A.; Nastopoulos, V.; Verzili, D.; Zamparelli, C.; Colotti, G.; Tsernoglou, D.; Chiancone, E. The crystal structure of the sorcin calcium binding domain provides a model of Ca2+-dependent processes in the full-length protein. J. Mol. Biol. 2002, 317, 447–458. [Google Scholar] [CrossRef]
- Kim, S.I.; Lee, H.J.; Kim, S.S.; Kwon, Y.S.; Chun, W. Sequestration of sorcin by aberrant forms of tau results in the defective calcium homeostasis. Korean J. Physiol. Pharm. 2016, 20, 387–397. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, P.; Jiang, Y.F.; Wang, J.H. shRNA-mediated silencing of sorcin increases drug chemosensitivity in myeloma KM3/DDP and U266/ADM cell lines. Int. J. Clin. Exp. Pathol. 2015, 8, 2300–2310. [Google Scholar] [PubMed]
- Anthony, D.F.; Beattie, J.; Paul, A.; Currie, S. Interaction of calcium/calmodulin-dependent protein kinase IIdeltaC with sorcin indirectly modulates ryanodine receptor function in cardiac myocytes. J. Mol. Cell. Cardiol. 2007, 43, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, N.; Nakao, R.; Kondo, R.; Nishitsuji, M.; Saito, Y.; Kuga, T.; Hatayama, T.; Nakayama, Y. Increased expression of sorcin is associated with multidrug resistance in leukemia cells via up-regulation of MDR1 expression through cAMP response element-binding protein. Biochem. Biophys. Res. Commun. 2014, 448, 430–436. [Google Scholar] [CrossRef]
- Noordeen, N.A.; Meur, G.; Rutter, G.A.; Leclerc, I. Glucose-induced nuclear shuttling of ChREBP is mediated by sorcin and Ca(2+) ions in pancreatic beta-cells. Diabetes 2012, 61, 574–585. [Google Scholar] [CrossRef]
- Xie, H.; Chang, M.; Hu, X.; Wang, D.; Tian, M.; Li, G.; Jiang, H.; Wang, Y.; Dong, Z.; Zhang, Y.; et al. Proteomics analysis of MPP+-induced apoptosis in SH-SY5Y cells. Neurol. Sci. 2011, 32, 221–228. [Google Scholar] [CrossRef]
- Andreev, V.P.; Petyuk, V.A.; Brewer, H.M.; Karpievitch, Y.V.; Xie, F.; Clarke, J.; Camp, D.; Smith, R.D.; Lieberman, A.P.; Albin, R.L.; et al. Label-free quantitative LC-MS proteomics of Alzheimer’s disease and normally aged human brains. J. Proteome Res. 2012, 11, 3053–3067. [Google Scholar] [CrossRef]
- Tsuji, T.; Shiozaki, A.; Kohno, R.; Yoshizato, K.; Shimohama, S. Proteomic profiling and neurodegeneration in Alzheimer’s disease. Neurochem Res. 2002, 27, 1245–1253. [Google Scholar] [CrossRef]
- Seyfried, N.T.; Dammer, E.B.; Swarup, V.; Nandakumar, D.; Duong, D.M.; Yin, L.; Deng, Q.; Nguyen, T.; Hales, C.M.; Wingo, T.; et al. A Multi-network Approach Identifies Protein-Specific Co-expression in Asymptomatic and Symptomatic Alzheimer’s Disease. Cell Syst 2017, 4, 60–72. [Google Scholar] [CrossRef]
- Drummond, E.; Nayak, S.; Faustin, A.; Pires, G.; Hickman, R.A.; Askenazi, M.; Cohen, M.; Haldiman, T.; Kim, C.; Han, X.; et al. Proteomic differences in amyloid plaques in rapidly progressive and sporadic Alzheimer’s disease. Acta Neuropathol. 2017, 133, 933–954. [Google Scholar] [CrossRef]
- Hondius, D.C.; Eigenhuis, K.N.; Morrema, T.H.J.; van der Schors, R.C.; van Nierop, P.; Bugiani, M.; Li, K.W.; Hoozemans, J.J.M.; Smit, A.B.; Rozemuller, A.J.M. Proteomics analysis identifies new markers associated with capillary cerebral amyloid angiopathy in Alzheimer’s disease. Acta Neuropathol. Commun. 2018, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Umoh, M.E.; Dammer, E.B.; Dai, J.; Duong, D.M.; Lah, J.J.; Levey, A.I.; Gearing, M.; Glass, J.D.; Seyfried, N.T. A proteomic network approach across the ALS-FTD disease spectrum resolves clinical phenotypes and genetic vulnerability in human brain. Embo Mol. Med. 2018, 10, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.J.; Heyny-von Haussen, R.; Mall, G.; Wolf, S. Proteome analysis of human substantia nigra in Parkinson’s disease. Proteome Sci. 2008, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Hulette, C.; Wang, Y.; Zhang, T.; Pan, C.; Wadhwa, R.; Zhang, J. Proteomic identification of a stress protein, mortalin/mthsp70/GRP75: relevance to Parkinson disease. Mol. Cell. Proteom. 2006, 5, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- George, G.; Singh, S.; Lokappa, S.B.; Varkey, J. Gene co-expression network analysis for identifying genetic markers in Parkinson’s disease - a three-way comparative approach. Genomics 2019, 111, 819–830. [Google Scholar] [CrossRef]
- Kalathur, R.K.; Giner-Lamia, J.; Machado, S.; Barata, T.; Ayasolla, K.R.; Futschik, M.E. The unfolded protein response and its potential role in Huntington’s disease elucidated by a systems biology approach. F1000Res 2015, 4, 103. [Google Scholar] [CrossRef]
- Pack-Chung, E.; Meyers, M.B.; Pettingell, W.P.; Moir, R.D.; Brownawell, A.M.; Cheng, I.; Tanzi, R.E.; Kim, T.W. Presenilin 2 interacts with sorcin, a modulator of the ryanodine receptor. J. Biol. Chem. 2000, 275, 14440–14445. [Google Scholar] [CrossRef]
- Woods, W.S.; Boettcher, J.M.; Zhou, D.H.; Kloepper, K.D.; Hartman, K.L.; Ladror, D.T.; Qi, Z.; Rienstra, C.M.; George, J.M. Conformation-specific binding of alpha-synuclein to novel protein partners detected by phage display and NMR spectroscopy. J. Biol. Chem. 2007, 282, 34555–34567. [Google Scholar] [CrossRef]
- Meyers, M.B.; Biedler, J.L. Increased synthesis of a low molecular weight protein in vincristine-resistant cells. Biochem. Biophys. Res. Commun. 1981, 99, 228–235. [Google Scholar] [CrossRef]
- Van der Bliek, A.M.; Baas, F.; Van der Velde-Koerts, T.; Biedler, J.L.; Meyers, M.B.; Ozols, R.F.; Hamilton, T.C.; Joenje, H.; Borst, P. Genes amplified and overexpressed in human multidrug-resistant cell lines. Cancer Res. 1988, 48, 5927–5932. [Google Scholar]
- Chen, J.; Watanabe, M.; Huang, P.; Sakaguchi, M.; Ochiai, K.; Nasu, Y.; Ouchida, M.; Huh, N.H.; Shimizu, K.; Kashiwakura, Y.; et al. REIC/Dkk-3 stable transfection reduces the malignant phenotype of mouse prostate cancer RM9 cells. Int J. Mol. Med. 2009, 24, 789–794. [Google Scholar] [PubMed]
- Dabaghi, M.; Rahgozar, S.; Moshtaghian, J.; Moafi, A.; Abedi, M.; Pourabutaleb, E. Overexpression of SORCIN is a Prognostic Biomarker for Multidrug-Resistant Pediatric Acute Lymphoblastic Leukemia and Correlates with Upregulated MDR1/P-gp. Genet. Test. Mol. Biomark. 2016, 20, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Su, T.; Leng, A.; Zhang, X.; Xu, M.; Yan, L.; Gu, H.; Zhang, G. Upregulation of soluble resistance-related calcium-binding protein (sorcin) in gastric cancer. Med. Oncol. 2010, 27, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, W.; Liu, X.; Gao, F.; Zhao, X. Reversing effect and mechanism of soluble resistance-related calcium-binding protein on multidrug resistance in human lung cancer A549/DDP cells. Mol. Med. Rep. 2015, 11, 2118–2124. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nagpal, J.K.; Das, B.R. Identification of differentially expressed genes in tobacco chewing-mediated oral cancer by differential display-polymerase chain reaction. Eur. J. Clin. Investig. 2007, 37, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Yang, Y.; Liu, B.; Xiao, W. Comparative proteomic profiling identified sorcin being associated with gemcitabine resistance in non-small cell lung cancer. Med Oncol. 2010, 27, 1303–1308. [Google Scholar] [CrossRef]
- Tan, Y.; Li, G.; Zhao, C.; Wang, J.; Zhao, H.; Xue, Y.; Han, M.; Yang, C. Expression of sorcin predicts poor outcome in acute myeloid leukemia. Leuk. Res. 2003, 27, 125–131. [Google Scholar] [CrossRef]
- Tong, W.; Sun, D.; Wang, Q.; Suo, J. Sorcin Enhances Metastasis and Promotes Epithelial-to-Mesenchymal Transition of Colorectal Cancer. Cell Biochem. Biophys. 2015, 72, 453–459. [Google Scholar] [CrossRef]
- Yang, Y.X.; Chen, Z.C.; Zhang, G.Y.; Yi, H.; Xiao, Z.Q. A subcelluar proteomic investigation into vincristine-resistant gastric cancer cell line. J. Cell. Biochem. 2008, 104, 1010–1021. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, Y.; Tan, Y.; Qi, J.; Xiao, Y.; Yang, C.; Zhu, Z.; Xiong, D. Sorcin, an important gene associated with multidrug-resistance in human leukemia cells. Leuk. Res. 2006, 30, 469–476. [Google Scholar] [CrossRef]
- Pomeroy, S.L.; Tamayo, P.; Gaasenbeek, M.; Sturla, L.M.; Angelo, M.; McLaughlin, M.E.; Kim, J.Y.; Goumnerova, L.C.; Black, P.M.; Lau, C.; et al. Prediction of central nervous system embryonal tumour outcome based on gene expression. Nature 2002, 415, 436–442. [Google Scholar] [CrossRef] [PubMed]
- French, P.J.; Swagemakers, S.M.; Nagel, J.H.; Kouwenhoven, M.C.; Brouwer, E.; van der Spek, P.; Luider, T.M.; Kros, J.M.; van den Bent, M.J.; Sillevis Smitt, P.A. Gene expression profiles associated with treatment response in oligodendrogliomas. Cancer Res. 2005, 65, 11335–11344. [Google Scholar] [CrossRef] [PubMed]
- Shai, R.; Shi, T.; Kremen, T.J.; Horvath, S.; Liau, L.M.; Cloughesy, T.F.; Mischel, P.S.; Nelson, S.F. Gene expression profiling identifies molecular subtypes of gliomas. Oncogene 2003, 22, 4918–4923. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Hui, A.M.; Su, Q.; Vortmeyer, A.; Kotliarov, Y.; Pastorino, S.; Passaniti, A.; Menon, J.; Walling, J.; Bailey, R.; et al. Neuronal and glioma-derived stem cell factor induces angiogenesis within the brain. Cancer Cell 2006, 9, 287–300. [Google Scholar] [CrossRef]
- Yokota, T.; Kouno, J.; Adachi, K.; Takahashi, H.; Teramoto, A.; Matsumoto, K.; Sugisaki, Y.; Onda, M.; Tsunoda, T. Identification of histological markers for malignant glioma by genome-wide expression analysis: dynein, alpha-PIX and sorcin. Acta Neuropathol. 2006, 111, 29–38. [Google Scholar] [CrossRef]
- He, Q.; Zhang, G.; Hou, D.; Leng, A.; Xu, M.; Peng, J.; Liu, T. Overexpression of sorcin results in multidrug resistance in gastric cancer cells with up-regulation of P-gp. Oncol. Rep. 2011, 25, 237–243. [Google Scholar]
- Hu, Y.; Li, S.; Yang, M.; Yan, C.; Fan, D.; Zhou, Y.; Zhang, Y.; Yague, E.; Xiong, D. Sorcin silencing inhibits epithelial-to-mesenchymal transition and suppresses breast cancer metastasis in vivo. Breast Cancer Res. Treat. 2014, 143, 287–299. [Google Scholar] [CrossRef]
- Kawakami, M.; Nakamura, T.; Okamura, N.; Komoto, C.; Markova, S.; Kobayashi, H.; Hashimoto, N.; Okumura, K.; Sakaeda, T. Knock-down of sorcin induces up-regulation of MDR1 in HeLa cells. Biol. Pharm. Bull. 2007, 30, 1065–1073. [Google Scholar] [CrossRef]
- Liu, X.; Chen, L.; Feng, B.; Liu, G. Reversing effect of sorcin in the drug resistance of human nasopharyngeal carcinoma. Anat. Rec. 2014, 297, 215–221. [Google Scholar] [CrossRef]
- Parekh, H.K.; Deng, H.B.; Choudhary, K.; Houser, S.R.; Simpkins, H. Overexpression of sorcin, a calcium-binding protein, induces a low level of paclitaxel resistance in human ovarian and breast cancer cells. Biochem. Pharmacol. 2002, 63, 1149–1158. [Google Scholar] [CrossRef]
- Demidova, N.S.; Ilyinskaya, G.V.; Shiryaeva, O.A.; Chernova, O.B.; Goncharova, S.A.; Kopnin, B.P. Decreased sensitivity of multidrug-resistant tumor cells to cisplatin is correlated with sorcin gene co-amplification. Neoplasma 1995, 42, 195–201. [Google Scholar] [PubMed]
- Liang, B.; Peng, P.; Chen, S.; Li, L.; Zhang, M.; Cao, D.; Yang, J.; Li, H.; Gui, T.; Li, X.; et al. Characterization and proteomic analysis of ovarian cancer-derived exosomes. J. Proteom. 2013, 80, 171–182. [Google Scholar] [CrossRef]
- Van der Bliek, A M.; Meyers, M.B.; Biedler, J.L.; Hes, E.; Borst, P. A 22-kd protein (sorcin/V19) encoded by an amplified gene in multidrug-resistant cells, is homologous to the calcium-binding light chain of calpain. Embo J. 1986, 5, 3201–3208. [Google Scholar] [CrossRef] [PubMed]
- Genovese, I.; Fiorillo, A.; Ilari, A.; Masciarelli, S.; Fazi, F.; Colotti, G. Binding of doxorubicin to Sorcin impairs cell death and increases drug resistance in cancer cells. Cell Death Dis. 2017, 8, e2950. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.; Wang, C.; Meng, Q.; Liu, Z.; Huo, X.; Yang, X.; Sun, P.; Sun, H.; Ma, X.; et al. Combination of dihydromyricetin and ondansetron strengthens antiproliferative efficiency of adriamycin in K562/ADR through downregulation of SORCIN: A new strategy of inhibiting P-glycoprotein. J. Cell. Physiol. 2019, 234, 3685–3696. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Sirohi, V.K.; Kumari, S.; Shukla, V.; Manohar, M.; Popli, P.; Dwivedi, A. Sorcin is involved during embryo implantation via activating VEGF/PI3K/Akt pathway in mice. J. Mol. Endocrinol. 2018, 60, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Khan, H.; Sirohi, V.K.; Das, V.; Agarwal, A.; Pandey, A.; Siddiqui, W.A.; Dwivedi, A. Alteration in endometrial proteins during early- and mid-secretory phases of the cycle in women with unexplained infertility. PloS ONE 2014, 9, e111687. [Google Scholar] [CrossRef]
- Tuo, H.; Shu, F.; She, S.; Yang, M.; Zou, X.Q.; Huang, J.; Hu, H.D.; Hu, P.; Ren, H.; Peng, S.F.; et al. Sorcin induces gastric cancer cell migration and invasion contributing to STAT3 activation. Oncotarget 2017, 8, 104258–104271. [Google Scholar] [CrossRef]
- Kawasaki, H.; Mizutome, H.; Kretsinger, R.H. Interaction sites of PEF proteins for recognition of their targets. Int. J. Biol. Macromol 2019, 133, 1035–1041. [Google Scholar] [CrossRef]
- Mella, M.; Colotti, G.; Zamparelli, C.; Verzili, D.; Ilari, A.; Chiancone, E. Information transfer in the penta-EF-hand protein sorcin does not operate via the canonical structural/functional pairing. A study with site-specific mutants. J. Biol. Chem. 2003, 278, 24921–24928. [Google Scholar] [CrossRef]
- Bers, D.M.; Despa, S.; Bossuyt, J. Regulation of Ca2+ and Na+ in normal and failing cardiac myocytes. Ann. N. Y. Acad. Sci. 2006, 1080, 165–177. [Google Scholar] [CrossRef]
- Brownawell, A.M.; Creutz, C.E. Calcium-dependent binding of sorcin to the N-terminal domain of synexin (annexin VII). J. Biol. Chem. 1997, 272, 22182–22190. [Google Scholar] [CrossRef] [PubMed]
- Tomas, A.; Futter, C.; Moss, S.E. Annexin 11 is required for midbody formation and completion of the terminal phase of cytokinesis. J. Cell Biol. 2004, 165, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Salzer, U.; Hinterdorfer, P.; Hunger, U.; Borken, C.; Prohaska, R. Ca(++)-dependent vesicle release from erythrocytes involves stomatin-specific lipid rafts, synexin (annexin VII), and sorcin. Blood 2002, 99, 2569–2577. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.C.; Chen, T.S.; Lim, S.K. Mesenchymal stem cell exosome: a novel stem cell-based therapy for cardiovascular disease. Regen. Med. 2011, 6, 481–492. [Google Scholar] [CrossRef]
- Buschow, S.I.; van Balkom, B.W.; Aalberts, M.; Heck, A.J.; Wauben, M.; Stoorvogel, W. MHC class II-associated proteins in B-cell exosomes and potential functional implications for exosome biogenesis. Immunol. Cell Biol. 2010, 88, 851–856. [Google Scholar] [CrossRef]
- Demory Beckler, M.; Higginbotham, J.N.; Franklin, J.L.; Ham, A.J.; Halvey, P.J.; Imasuen, I.E.; Whitwell, C.; Li, M.; Liebler, D.C.; Coffey, R.J. Proteomic analysis of exosomes from mutant KRAS colon cancer cells identifies intercellular transfer of mutant KRAS. Mol. Cell. Proteom. 2013, 12, 343–355. [Google Scholar] [CrossRef]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-scale proteomics and phosphoproteomics of urinary exosomes. J. Am. Soc. Nephrol. 2009, 20, 363–379. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Gangoda, L.; Liem, M.; Fonseka, P.; Atukorala, I.; Ozcitti, C.; Mechler, A.; Adda, C.G.; Ang, C.S.; Mathivanan, S. Proteogenomic analysis reveals exosomes are more oncogenic than ectosomes. Oncotarget 2015, 6, 15375–15396. [Google Scholar] [CrossRef]
- Kharaziha, P.; Chioureas, D.; Rutishauser, D.; Baltatzis, G.; Lennartsson, L.; Fonseca, P.; Azimi, A.; Hultenby, K.; Zubarev, R.; Ullen, A.; et al. Molecular profiling of prostate cancer derived exosomes may reveal a predictive signature for response to docetaxel. Oncotarget 2015, 6, 21740–21754. [Google Scholar] [CrossRef]
- Pienimaeki-Roemer, A.; Kuhlmann, K.; Bottcher, A.; Konovalova, T.; Black, A.; Orso, E.; Liebisch, G.; Ahrens, M.; Eisenacher, M.; Meyer, H.E.; et al. Lipidomic and proteomic characterization of platelet extracellular vesicle subfractions from senescent platelets. Transfusion 2015, 55, 507–521. [Google Scholar] [CrossRef]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [PubMed]
- Rutti, S.; Arous, C.; Schvartz, D.; Timper, K.; Sanchez, J.C.; Dermitzakis, E.; Donath, M.Y.; Halban, P.A.; Bouzakri, K. Fractalkine (CX3CL1), a new factor protecting beta-cells against TNFalpha. Mol. Metab. 2014, 3, 731–741. [Google Scholar] [CrossRef]
- Tu, H.; Nelson, O.; Bezprozvanny, A.; Wang, Z.; Lee, S.F.; Hao, Y.H.; Serneels, L.; De Strooper, B.; Yu, G.; Bezprozvanny, I. Presenilins form ER Ca2+ leak channels, a function disrupted by familial Alzheimer’s disease-linked mutations. Cell 2006, 126, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Gracy, K.N.; Clarke, C.L.; Meyers, M.B.; Pickel, V.M. N-methyl-D-aspartate receptor 1 in the caudate-putamen nucleus: Ultrastructural localization and co-expression with sorcin, a 22,000 mol. wt calcium binding protein. Neuroscience 1999, 90, 107–117. [Google Scholar] [CrossRef]
- Clemen, C.S.; Herr, C.; Hovelmeyer, N.; Noegel, A.A. The lack of annexin A7 affects functions of primary astrocytes. Exp. Cell Res. 2003, 291, 406–414. [Google Scholar] [CrossRef]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Reviews. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Deng, L.M.; Tan, T.; Zhang, T.Y.; Xiao, X.F.; Gu, H. miR1 reverses multidrug resistance in gastric cancer cells via downregulation of sorcin through promoting the accumulation of intracellular drugs and apoptosis of cells. Int. J. Oncol. 2019, 55, 451–461. [Google Scholar]
- Hu, Y.; Cheng, X.; Li, S.; Zhou, Y.; Wang, J.; Cheng, T.; Yang, M.; Xiong, D. Inhibition of sorcin reverses multidrug resistance of K562/A02 cells and MCF-7/A02 cells via regulating apoptosis-related proteins. Cancer Chemother. Pharmacol. 2013, 72, 789–798. [Google Scholar] [CrossRef]
- Fletcher, J.I.; Williams, R.T.; Henderson, M.J.; Norris, M.D.; Haber, M. ABC transporters as mediators of drug resistance and contributors to cancer cell biology. Drug Resist. Updates 2016, 26, 1–9. [Google Scholar] [CrossRef]
- Genovese, I.; Ilari, A.; Assaraf, Y.G.; Fazi, F.; Colotti, G. Not only P-glycoprotein: Amplification of the ABCB1-containing chromosome region 7q21 confers multidrug resistance upon cancer cells by coordinated overexpression of an assortment of resistance-related proteins. Drug Resist. Updates 2017, 32, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Abolhoda, A.; Wilson, A.E.; Ross, H.; Danenberg, P.V.; Burt, M.; Scotto, K.W. Rapid activation of MDR1 gene expression in human metastatic sarcoma after in vivo exposure to doxorubicin. Clin. Cancer Res. 1999, 5, 3352–3356. [Google Scholar]
- Brugger, D.; Brischwein, K.; Liu, C.; Bader, P.; Niethammer, D.; Gekeler, V.; Beck, J.F. Induction of drug resistance and protein kinase C genes in A2780 ovarian cancer cells after incubation with antineoplastic agents at sublethal concentrations. Anticancer Res. 2002, 22, 4229–4232. [Google Scholar] [PubMed]
- Chin, K.V.; Tanaka, S.; Darlington, G.; Pastan, I.; Gottesman, M.M. Heat shock and arsenite increase expression of the multidrug resistance (MDR1) gene in human renal carcinoma cells. J. Biol. Chem. 1990, 265, 221–226. [Google Scholar] [PubMed]
- Fojo, A.T.; Ueda, K.; Slamon, D.J.; Poplack, D.G.; Gottesman, M.M.; Pastan, I. Expression of a multidrug-resistance gene in human tumors and tissues. Proc. Natl. Acad. Sci. USA 1987, 84, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Gekeler, V.; Frese, G.; Diddens, H.; Probst, H. Expression of a P-glycoprotein gene is inducible in a multidrug-resistant human leukemia cell line. Biochem. Biophys. Res. Commun. 1988, 155, 754–760. [Google Scholar] [CrossRef]
- Hu, X.F.; Slater, A.; Wall, D.M.; Kantharidis, P.; Parkin, J.D.; Cowman, A.; Zalcberg, J.R. Rapid up-regulation of mdr1 expression by anthracyclines in a classical multidrug-resistant cell line. Br. J. Cancer 1995, 71, 931–936. [Google Scholar] [CrossRef]
- Liu, Z.L.; Onda, K.; Tanaka, S.; Toma, T.; Hirano, T.; Oka, K. Induction of multidrug resistance in MOLT-4 cells by anticancer agents is closely related to increased expression of functional P-glycoprotein and MDR1 mRNA. Cancer Chemother. Pharmacol. 2002, 49, 391–397. [Google Scholar] [CrossRef]
- Park, J.; Shinohara, N.; Liebert, M.; Noto, L.; Flint, A.; Grossman, H.B. P-glycoprotein expression in bladder cancer. J. Urol. 1994, 151, 43–46. [Google Scholar] [CrossRef]
- Schneider, J.; Efferth, T.; Centeno, M.M.; Mattern, J.; Rodriguez-Escudero, F.J.; Volm, M. High rate of expression of multidrug resistance-associated P-glycoprotein in human endometrial carcinoma and normal endometrial tissue. Eur. J. Cancer 1993, 29A, 554–558. [Google Scholar] [CrossRef]
- Schoenlein, P.V. Molecular cytogenetics of multiple drug resistance. Cytotechnology 1993, 12, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Scotto, K.W.; Biedler, J.L.; Melera, P.W. Amplification and expression of genes associated with multidrug resistance in mammalian cells. Science 1986, 232, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.K.; Lacayo, N.J.; Duran, G.E.; Wang, Y.; Bangs, C.D.; Rea, S.; Kovacs, M.; Cherry, A.M.; Brown, J.M.; Sikic, B.I. Preferential expression of a mutant allele of the amplified MDR1 (ABCB1) gene in drug-resistant variants of a human sarcoma. Gene Chromosomes Cancer 2002, 34, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Duesberg, P.; Li, R.; Sachs, R.; Fabarius, A.; Upender, M.B.; Hehlmann, R. Cancer drug resistance: the central role of the karyotype. Drug Resist. Updates 2007, 10, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Shibata, T.; Kokubu, A.; Ojima, H.; Loukopoulos, P.; Kanai, Y.; Kosuge, T.; Fukayama, M.; Kondo, T.; Sakamoto, M.; et al. Genetic profile of hepatocellular carcinoma revealed by array-based comparative genomic hybridization: identification of genetic indicators to predict patient outcome. J. Hepatol. 2005, 43, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.W.; Han, N.; Kim, M.G.; Kim, T.; Oh, J.M. Copy number variability analysis of pharmacogenes in patients with lymphoma, leukemia, hepatocellular, and lung carcinoma using The Cancer Genome Atlas data. Pharm. Genom. 2015, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mickley, L.A.; Spengler, B.A.; Knutsen, T.A.; Biedler, J.L.; Fojo, T. Gene rearrangement: a novel mechanism for MDR-1 gene activation. J. Clin. Investig. 1997, 99, 1947–1957. [Google Scholar] [CrossRef]
- Pang, E.; Hu, Y.; Chan, K.Y.; Lai, P.B.; Squire, J.A.; Macgregor, P.F.; Beheshti, B.; Albert, M.; Leung, T.W.; Wong, N. Karyotypic imbalances and differential gene expressions in the acquired doxorubicin resistance of hepatocellular carcinoma cells. Lab. Investig. 2005, 85, 664–674. [Google Scholar] [CrossRef]
- Bonte, D.; Lindvall, C.; Liu, H.; Dykema, K.; Furge, K.; Weinreich, M. Cdc7-Dbf4 kinase overexpression in multiple cancers and tumor cell lines is correlated with p53 inactivation. Neoplasia 2008, 10, 920–931. [Google Scholar] [CrossRef]
- Chao, C.C.; Ma, C.M.; Lin-Chao, S. Co-amplification and over-expression of two mdr genes in a multidrug-resistant human colon carcinoma cell line. FEBS Lett. 1991, 291, 214–218. [Google Scholar] [CrossRef]
- Finalet Ferreiro, J.; Rouhigharabaei, L.; Urbankova, H.; van der Krogt, J.A.; Michaux, L.; Shetty, S.; Krenacs, L.; Tousseyn, T.; De Paepe, P.; Uyttebroeck, A.; et al. Integrative genomic and transcriptomic analysis identified candidate genes implicated in the pathogenesis of hepatosplenic T-cell lymphoma. PloS ONE 2014, 9, e102977. [Google Scholar] [CrossRef] [PubMed]
- Flahaut, M.; Muhlethaler-Mottet, A.; Martinet, D.; Fattet, S.; Bourloud, K.B.; Auderset, K.; Meier, R.; Schmutz, N.B.; Delattre, O.; Joseph, J.M.; et al. Molecular cytogenetic characterization of doxorubicin-resistant neuroblastoma cell lines: evidence that acquired multidrug resistance results from a unique large amplification of the 7q21 region. Gene Chromosomes Cancer 2006, 45, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.N.; Ehlers, N.S.; Zhu, S.; Thomsen, M.B.; Nielsen, R.L.; Liu, D.; Wang, G.; Hou, Y.; Zhang, X.; Xu, X.; et al. The stepwise evolution of the exome during acquisition of docetaxel resistance in breast cancer cells. Bmc Genom. 2016, 17, 442. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Sterzynska, K.; Zawierucha, P.; Rucinski, M.; Swierczewska, M.; Partyka, M.; Bednarek-Rajewska, K.; Brazert, M.; Nowicki, M.; Zabel, M.; et al. Microarray-based detection and expression analysis of new genes associated with drug resistance in ovarian cancer cell lines. Oncotarget 2017, 8, 49944–49958. [Google Scholar] [CrossRef] [PubMed]
- Kitada, K.; Yamasaki, T. The MDR1/ABCB1 regional amplification in large inverted repeats with asymmetric sequences and microhomologies at the junction sites. Cancer Genet. Cytogenet. 2007, 178, 120–127. [Google Scholar] [CrossRef]
- Lee, S.; Kim, K.; Ho, J.N.; Jin, H.; Byun, S.S.; Lee, E. Analysis of resistance-associated gene expression in docetaxel-resistant prostate cancer cells. Oncol. Lett. 2017, 14, 3011–3018. [Google Scholar] [CrossRef]
- Litviakov, N.V.; Cherdyntseva, N.V.; Tsyganov, M.M.; Slonimskaya, E.M.; Ibragimova, M.K.; Kazantseva, P.V.; Kzhyshkowska, J.; Choinzonov, E.L. Deletions of multidrug resistance gene loci in breast cancer leads to the down-regulation of its expression and predict tumor response to neoadjuvant chemotherapy. Oncotarget 2016, 7, 7829–7841. [Google Scholar] [CrossRef]
- Patch, A.M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole-genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Torigoe, K.; Sato, S.; Kusaba, H.; Kohno, K.; Kuwano, M.; Okumura, K.; Green, E.D.; Tsui, L.C.; Scherer, S.W.; Schlessinger, D.; et al. A YAC-based contig of 1.5 Mb spanning the human multidrug resistance gene region and delineating the amplification unit in three human multidrug-resistant cell lines. Genome Res. 1995, 5, 233–244. [Google Scholar] [CrossRef]
- Van der Bliek, A.M.; Van der Velde-Koerts, T.; Ling, V.; Borst, P. Overexpression and amplification of five genes in a multidrug-resistant Chinese hamster ovary cell line. Mol. Cell. Biol. 1986, 6, 1671–1678. [Google Scholar] [CrossRef][Green Version]
- Yabuki, N.; Sakata, K.; Yamasaki, T.; Terashima, H.; Mio, T.; Miyazaki, Y.; Fujii, T.; Kitada, K. Gene amplification and expression in lung cancer cells with acquired paclitaxel resistance. Cancer Genet. Cytogenet. 2007, 173, 1–9. [Google Scholar] [CrossRef]
- Kadioglu, O.; Efferth, T. Peptide aptamer identified by molecular docking targeting translationally controlled tumor protein in leukemia cells. Investig. New Drugs 2016, 34, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.N.; Jiang, S.S.; Fan, C.C.; Lo, Y.K.; Kuo, C.Y.; Chen, C.H.; Liu, Y.L.; Lee, C.C.; Chen, W.S.; Huang, T.S.; et al. Increased Cdc7 expression is a marker of oral squamous cell carcinoma and overexpression of Cdc7 contributes to the resistance to DNA-damaging agents. Cancer Lett. 2013, 337, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Choschzick, M.; Lebeau, A.; Marx, A.H.; Tharun, L.; Terracciano, L.; Heilenkotter, U.; Jaenicke, F.; Bokemeyer, C.; Simon, R.; Sauter, G.; et al. Overexpression of cell division cycle 7 homolog is associated with gene amplification frequency in breast cancer. Hum. Pathol. 2010, 41, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; Fountaine, T.J.; Hennessy, J.; Bruggeman, R.D.; Clarke, J.T.; Mauger, D.T.; Helm, K.F. Cdc7 expression in melanomas, Spitz tumors and melanocytic nevi. J. Cutan. Pathol. 2009, 36, 433–438. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, H.Q.; Ba, Y. High expression of cell division cycle 7 protein correlates with poor prognosis in patients with diffuse large B-cell lymphoma. Med Oncol. 2012, 29, 3498–3503. [Google Scholar] [CrossRef]
- Kulkarni, A.A.; Kingsbury, S.R.; Tudzarova, S.; Hong, H.K.; Loddo, M.; Rashid, M.; Rodriguez-Acebes, S.; Prevost, A.T.; Ledermann, J.A.; Stoeber, K.; et al. Cdc7 kinase is a predictor of survival and a novel therapeutic target in epithelial ovarian carcinoma. Clin. Cancer Res. 2009, 15, 2417–2425. [Google Scholar] [CrossRef]
- Nambiar, S.; Mirmohammadsadegh, A.; Hassan, M.; Mota, R.; Marini, A.; Alaoui, A.; Tannapfel, A.; Hegemann, J.H.; Hengge, U.R. Identification and functional characterization of ASK/Dbf4, a novel cell survival gene in cutaneous melanoma with prognostic relevance. Carcinogenesis 2007, 28, 2501–2510. [Google Scholar] [CrossRef]
- Sasi, N.K.; Bhutkar, A.; Lanning, N.J.; MacKeigan, J.P.; Weinreich, M. DDK Promotes Tumor Chemoresistance and Survival via Multiple Pathways. Neoplasia 2017, 19, 439–450. [Google Scholar] [CrossRef]
- Ali, R.; Huang, Y.; Maher, S.E.; Kim, R.W.; Giordano, F.J.; Tellides, G.; Geirsson, A. miR-1 mediated suppression of Sorcin regulates myocardial contractility through modulation of Ca2+ signaling. J. Mol. Cell. Cardiol. 2012, 52, 1027–1037. [Google Scholar] [CrossRef]
- Shan, Z.X.; Lin, Q.X.; Deng, C.Y.; Zhu, J.N.; Mai, L.P.; Liu, J.L.; Fu, Y.H.; Liu, X.Y.; Li, Y.X.; Zhang, Y.Y.; et al. miR-1/miR-206 regulate Hsp60 expression contributing to glucose-mediated apoptosis in cardiomyocytes. Febs Lett. 2010, 584, 3592–3600. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Hu, Z.; Fu, H.; Zhang, H.; Wu, Y.; Zheng, X. MicroRNA-1 and microRNA-499 downregulate the expression of the ets1 proto-oncogene in HepG2 cells. Oncol. Rep. 2012, 28, 701–706. [Google Scholar] [CrossRef]
- Li, G.Y.; Liu, J.Z.; Zhang, B.; Yang, M.; Chen, S.G.; Hou, M.; Wang, L.X. Tegillarca granosa extract Haishengsu (HSS) suppresses expression of mdr1, BCR/ABL and sorcin in drug-resistant K562/ADM tumors in mice. Adv. Med Sci. 2013, 58, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Zhang, L.; Liu, J.Z.; Chen, S.G.; Xiao, T.W.; Liu, G.Z.; Wang, J.X.; Wang, L.X.; Hou, M. Marine drug Haishengsu increases chemosensitivity to conventional chemotherapy and improves quality of life in patients with acute leukemia. Biomed. Pharmacother. 2016, 81, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.J.; Tchack, L.; Angelo, G.; Pratt, R.E.; Sonna, L.A. DNA microarray analysis of vitamin D-induced gene expression in a human colon carcinoma cell line. Physiol. Genom. 2004, 17, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Groebe, K.; Cen, J.; Schvartz, D.; Sargsyan, E.; Chowdhury, A.; Roomp, K.; Schneider, R.; Alderborn, A.; Sanchez, J.C.; Bergsten, P. Palmitate-Induced Insulin Hypersecretion and Later Secretory Decline Associated with Changes in Protein Expression Patterns in Human Pancreatic Islets. J. Proteome Res. 2018, 17, 3824–3836. [Google Scholar] [CrossRef]
- Cen, J.; Sargsyan, E.; Forslund, A.; Bergsten, P. Mechanisms of beneficial effects of metformin on fatty acid-treated human islets. J. Mol. Endocrinol.s 2018, 61, 91–99. [Google Scholar] [CrossRef]
- Hu, H.; Zhu, S.; Tong, Y.; Huang, G.; Tan, B.; Yang, L. Antitumor activity of triptolide in SKOV3 cells and SKOV3/DDP in vivo and in vitro. Anticancer Drugs 2020. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Events: | Sorcin Role: | References Relevant Studies |
|---|---|---|
| Calcium homeostasis | Regulation of Ca2+ channels, pumps, exchangers | [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21] |
| Regulation of ER and cytosolic Ca2+ concentration | [4,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28] | |
| Regulation of heart-muscle contraction | [6,7,8,9,10,11,12,13,14,15,16,17,18,29,30] | |
| Cellular metabolism | ER stress sensor, unfolded protein response regulation | [20,22,26,27,28,31] |
| Regulation of mitosis, cytokinesis, cell cycle | [17,22,26,27,32] | |
| Regulation of kinases | [22,30,33,34] | |
| Regulation of glucose metabolism | [28,35] | |
| Neurodegeneration | Overexpression in neurodegenerative diseases | [31,36,37,38,39,40,41,42,43,44,45,46,47,48] |
| Cancer | Overexpression in tumors | [20,23,24,25,26,34,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65] |
| Increase of MDR | [24,25,26,32,34,52,54,56,60,66,67,68,69,70,71] | |
| Cell death | Regulation of cell death | [9,15,25,42,43,44,45,46,47,48] |
| Drug elimination | Drug binding-elimination | [72] |
| Efflux pumps | Increased expression of ABCB1 | [24,32,34,52,57,60,66,68,69,73,74,75] |
| EMT | Increase of EMT | [58,67] |
| Angiogenesis | Increase of invasion and angiogenesis | [67,76,77] |
| Metastatization | Increase of metastatization | [53,67,78] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battista, T.; Fiorillo, A.; Chiarini, V.; Genovese, I.; Ilari, A.; Colotti, G. Roles of Sorcin in Drug Resistance in Cancer: One Protein, Many Mechanisms, for a Novel Potential Anticancer Drug Target. Cancers 2020, 12, 887. https://doi.org/10.3390/cancers12040887
Battista T, Fiorillo A, Chiarini V, Genovese I, Ilari A, Colotti G. Roles of Sorcin in Drug Resistance in Cancer: One Protein, Many Mechanisms, for a Novel Potential Anticancer Drug Target. Cancers. 2020; 12(4):887. https://doi.org/10.3390/cancers12040887
Chicago/Turabian StyleBattista, Theo, Annarita Fiorillo, Valerio Chiarini, Ilaria Genovese, Andrea Ilari, and Gianni Colotti. 2020. "Roles of Sorcin in Drug Resistance in Cancer: One Protein, Many Mechanisms, for a Novel Potential Anticancer Drug Target" Cancers 12, no. 4: 887. https://doi.org/10.3390/cancers12040887
APA StyleBattista, T., Fiorillo, A., Chiarini, V., Genovese, I., Ilari, A., & Colotti, G. (2020). Roles of Sorcin in Drug Resistance in Cancer: One Protein, Many Mechanisms, for a Novel Potential Anticancer Drug Target. Cancers, 12(4), 887. https://doi.org/10.3390/cancers12040887