MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells


 ,
,  ,
, 
Abstract
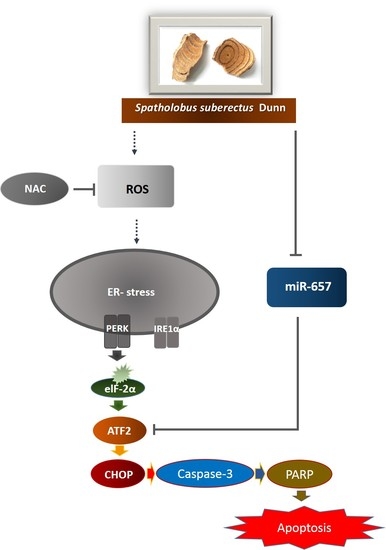
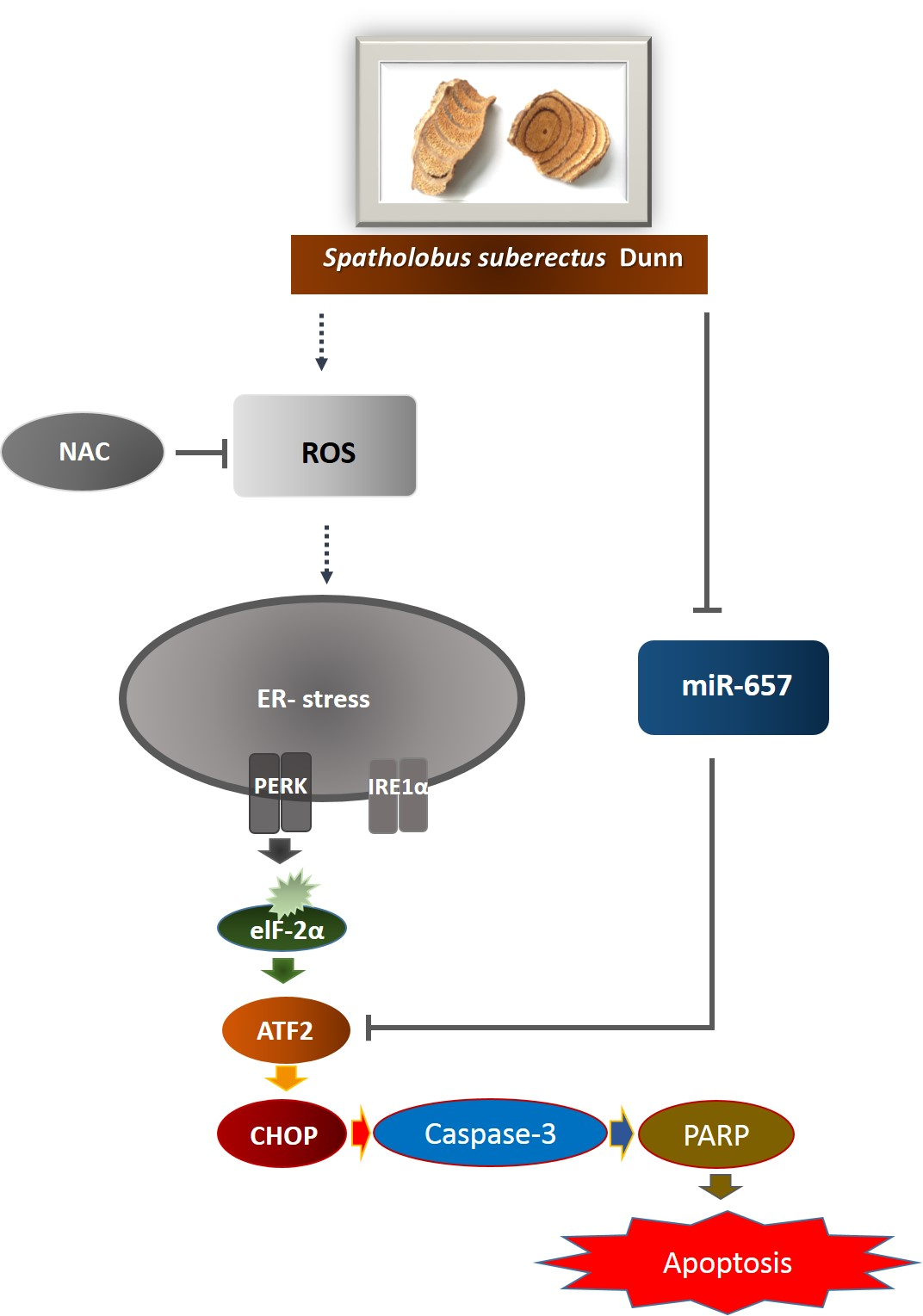
1. Introduction
2. Results
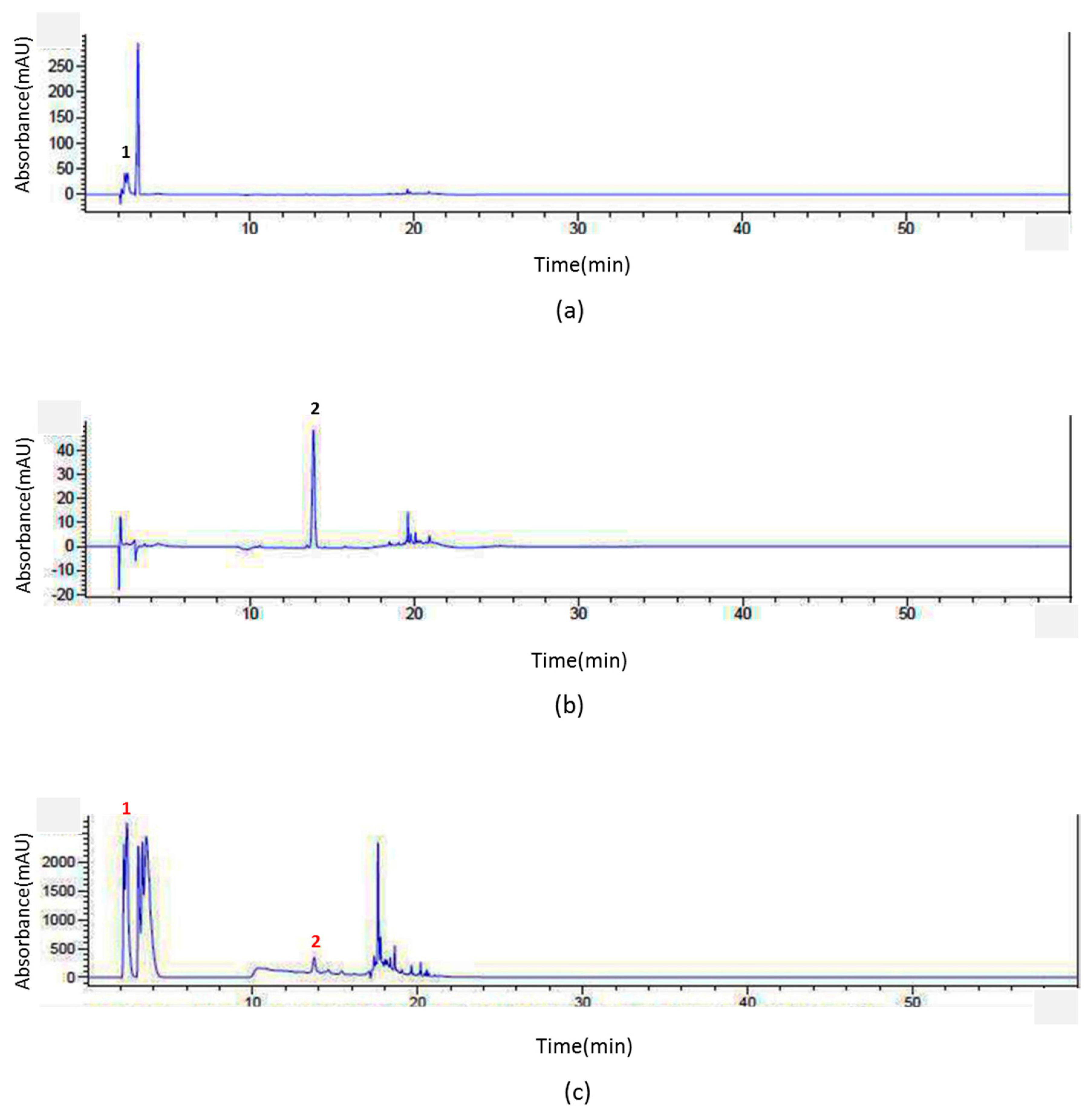
2.1. Identification of Epigallocatechin (EGC) and Genistein from SSD through HPLC Analysis
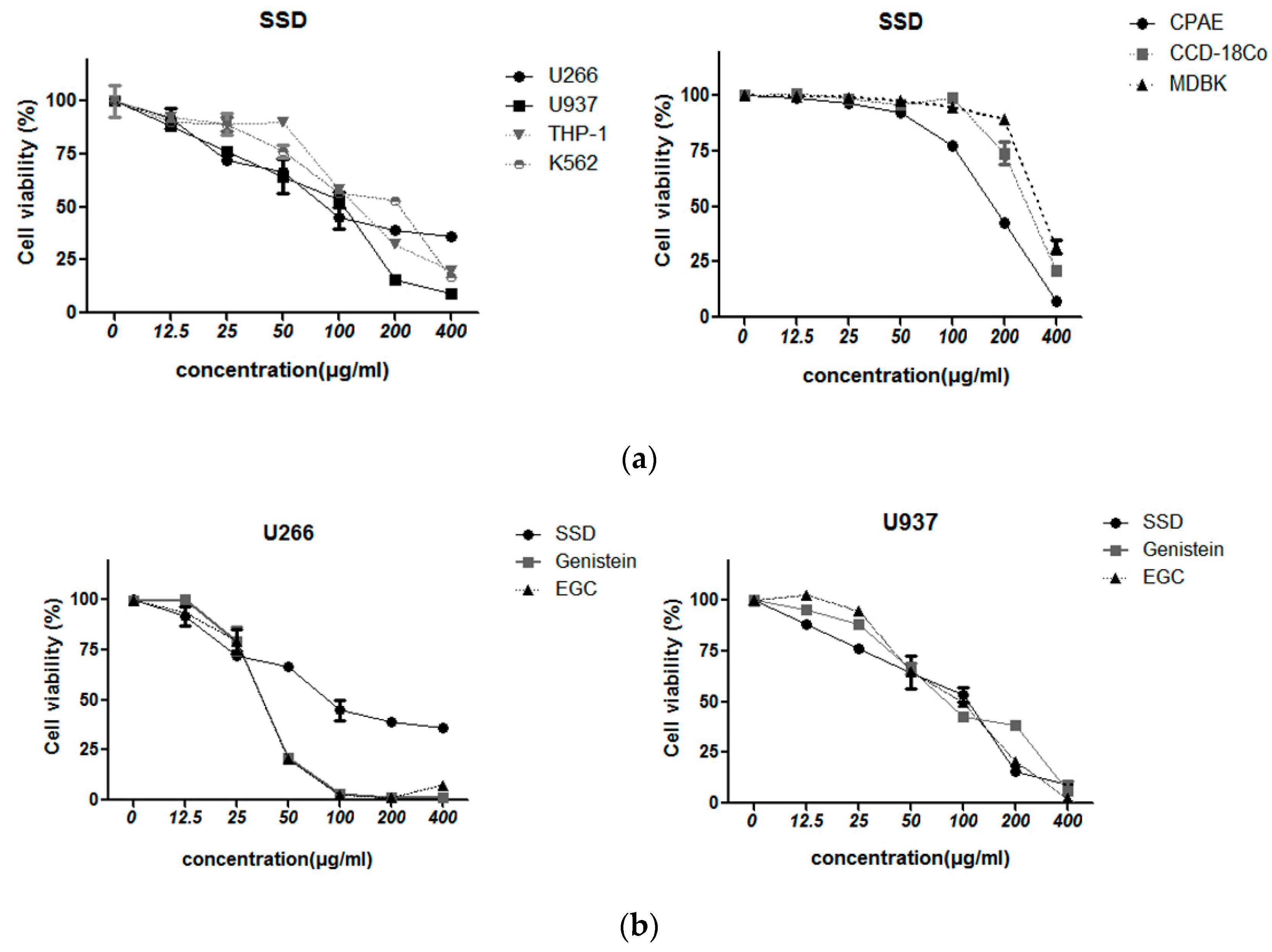
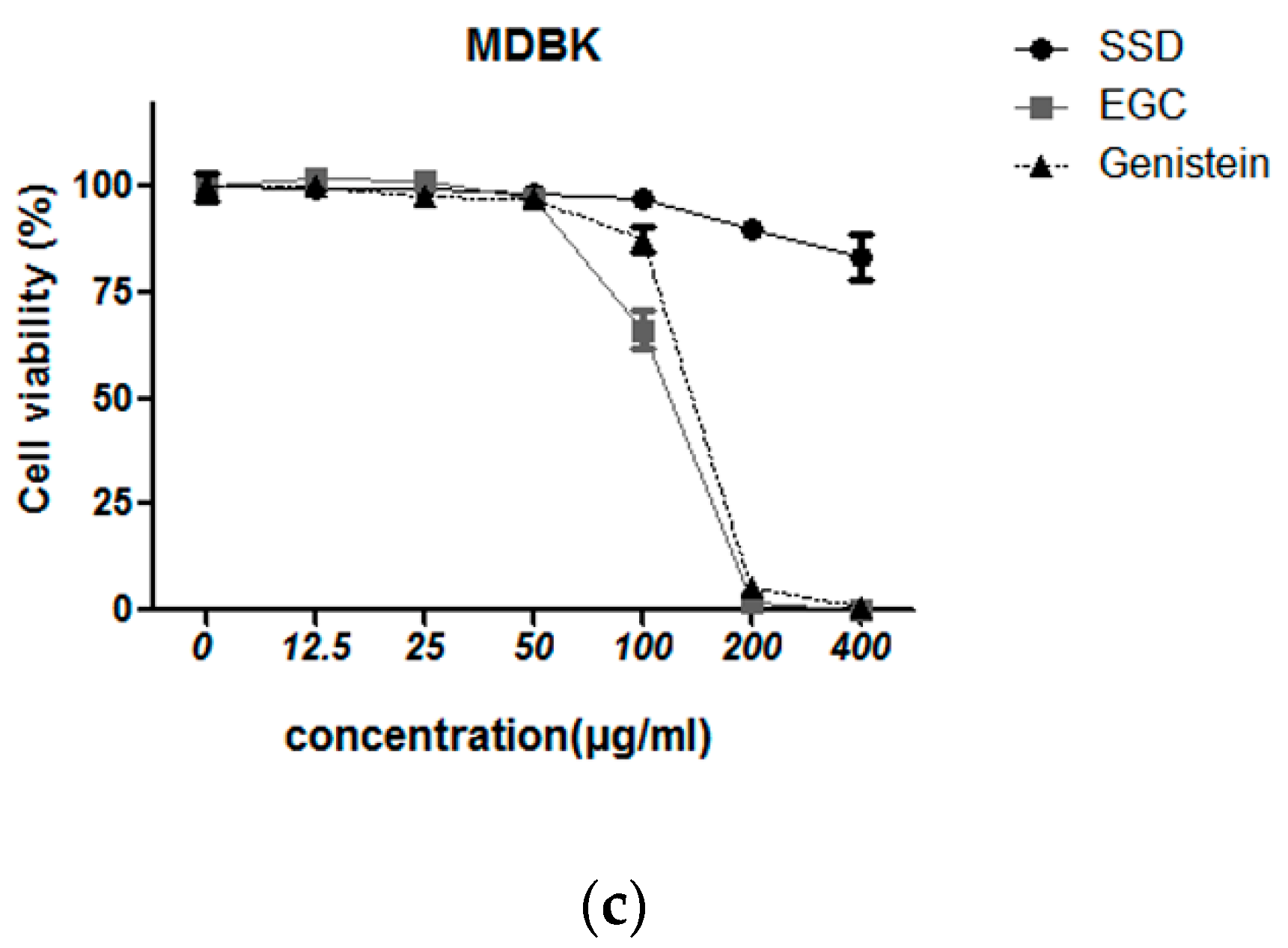
2.2. SSD Exerted Cytotoxicity in Hematological Cancers but Less in Normal Cells
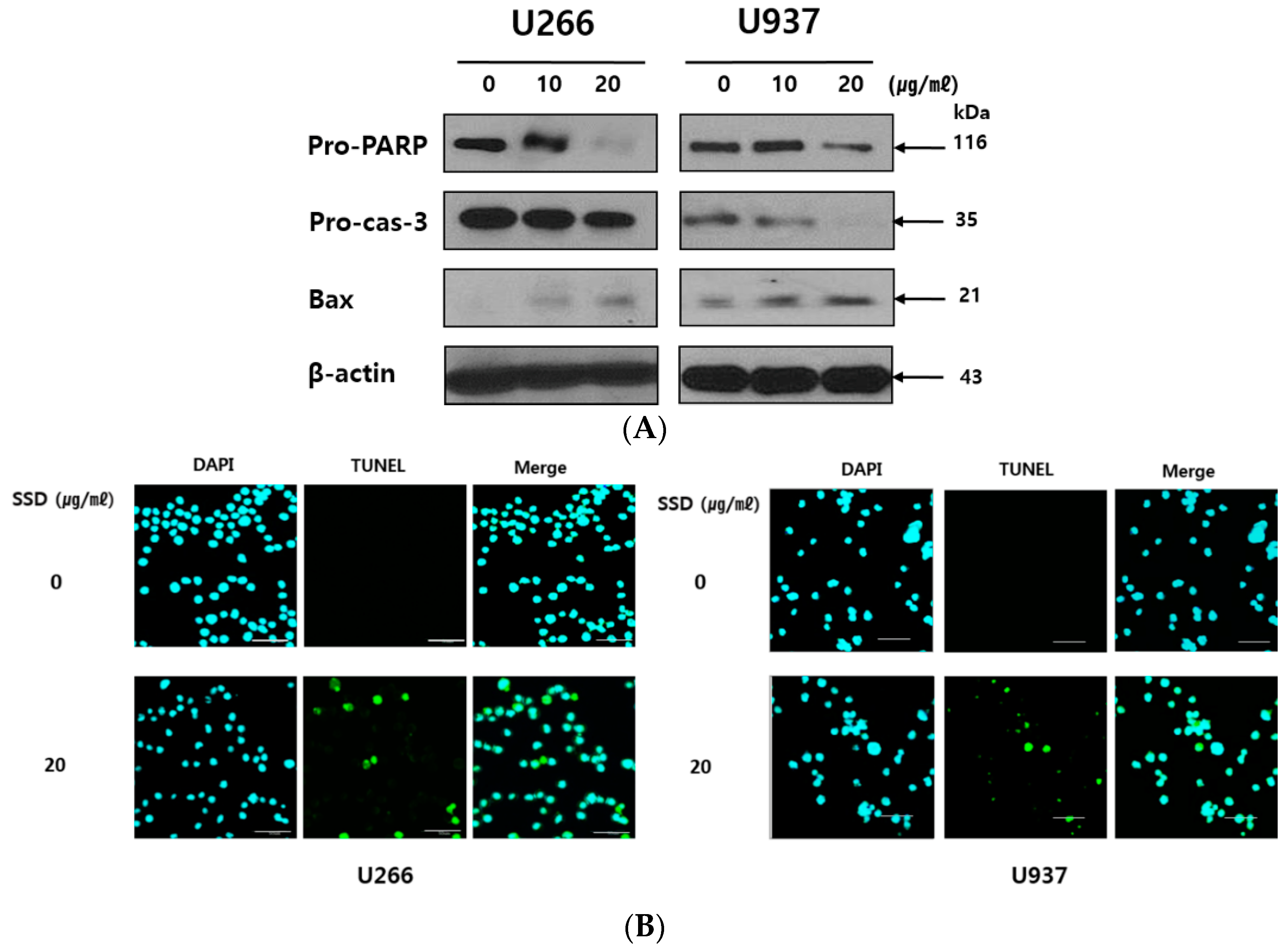
2.3. SSD Induced Apoptosis in U266 and U937 Cells
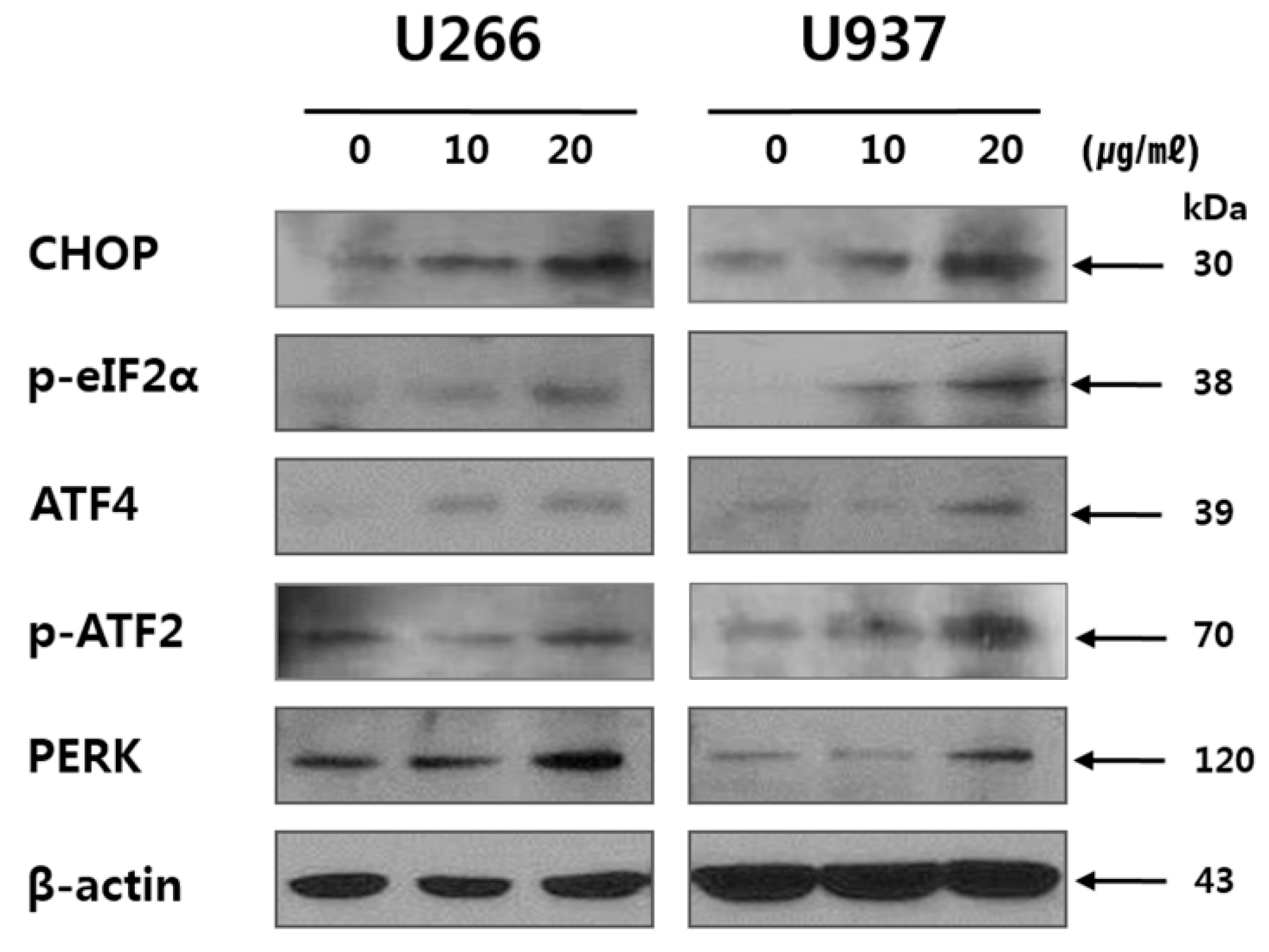
2.4. SSD Upregulated ER Stress-Related Proteins in U266 and U937 Cells
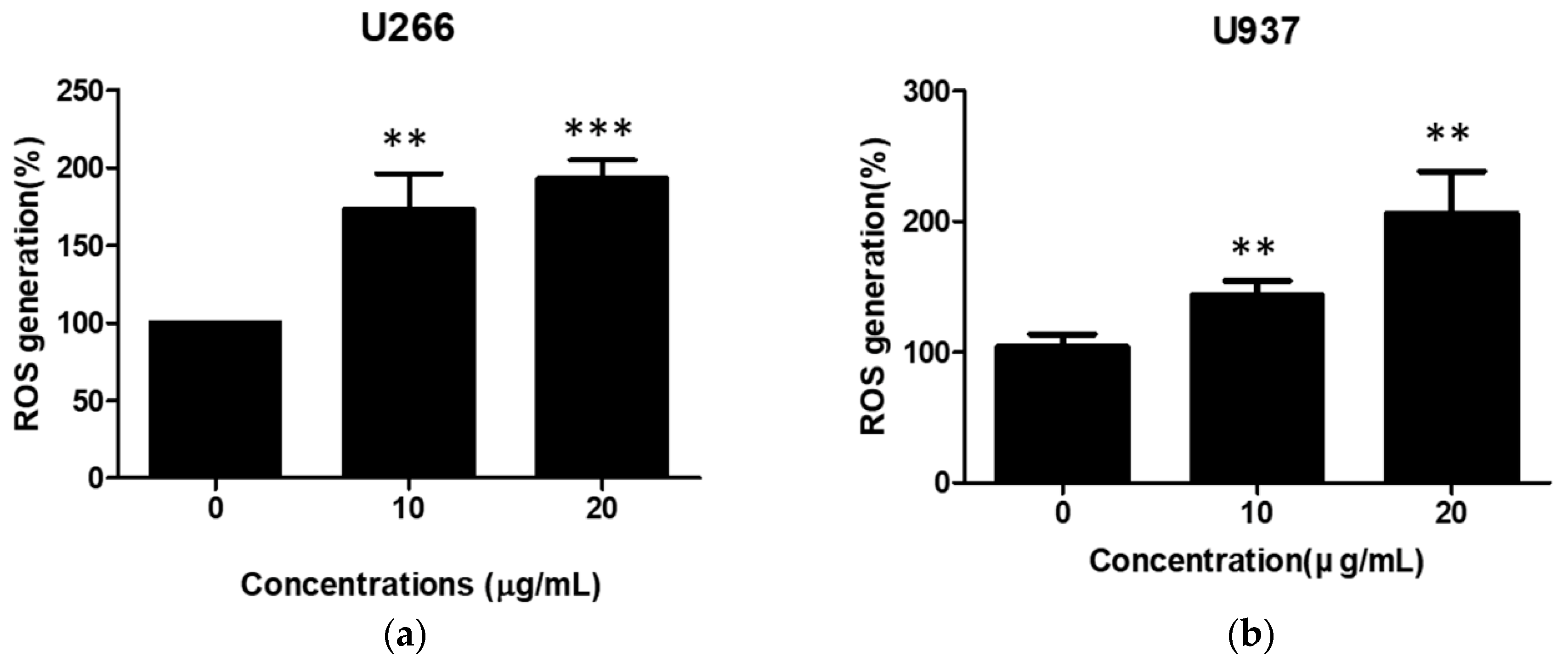
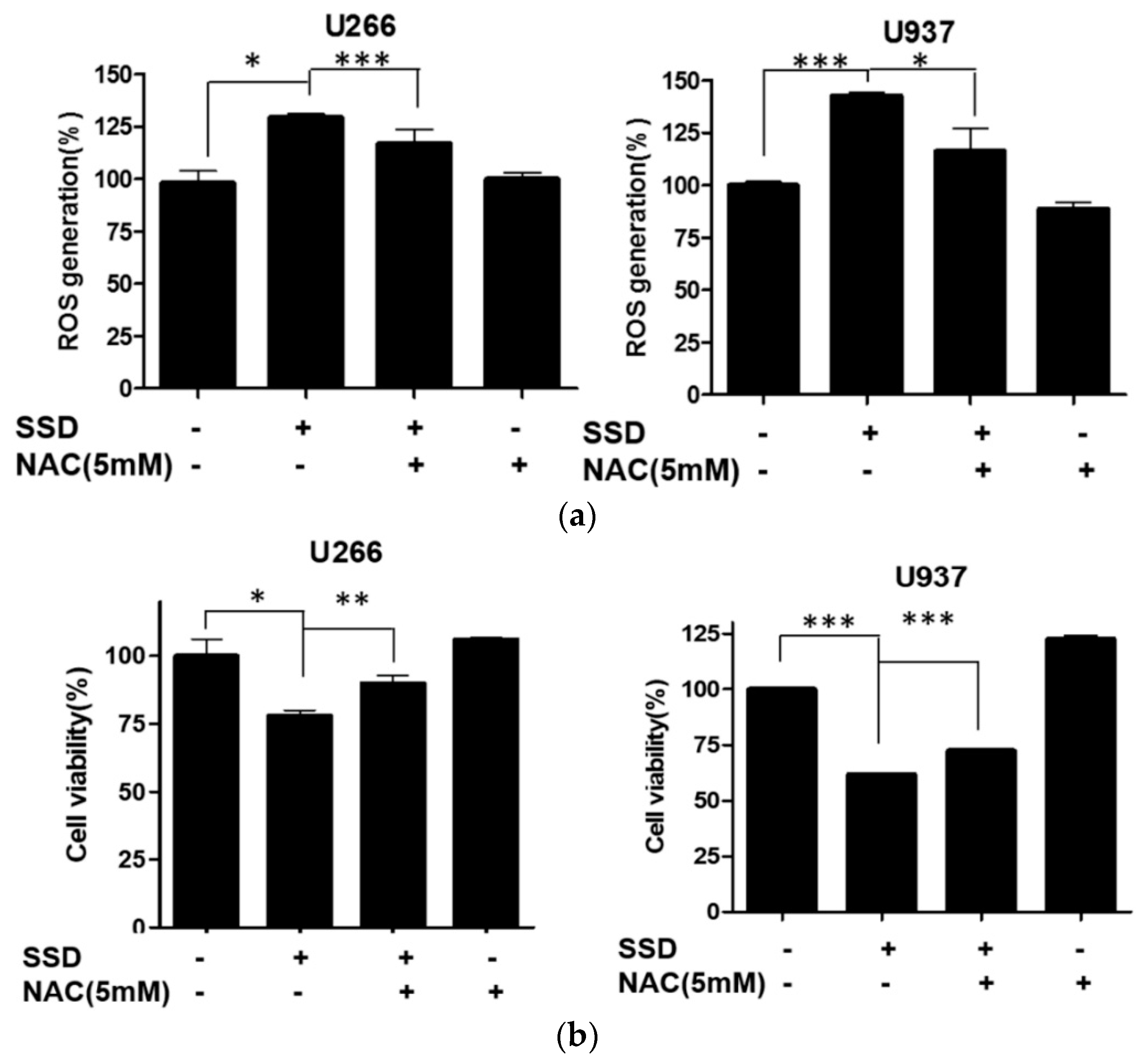
2.5. SSD Increased ROS Generation in U266 and U937 Cells
2.6. ROS Scavenger Reversed SSD-Induced Apoptosis in U266 and U937 Cells
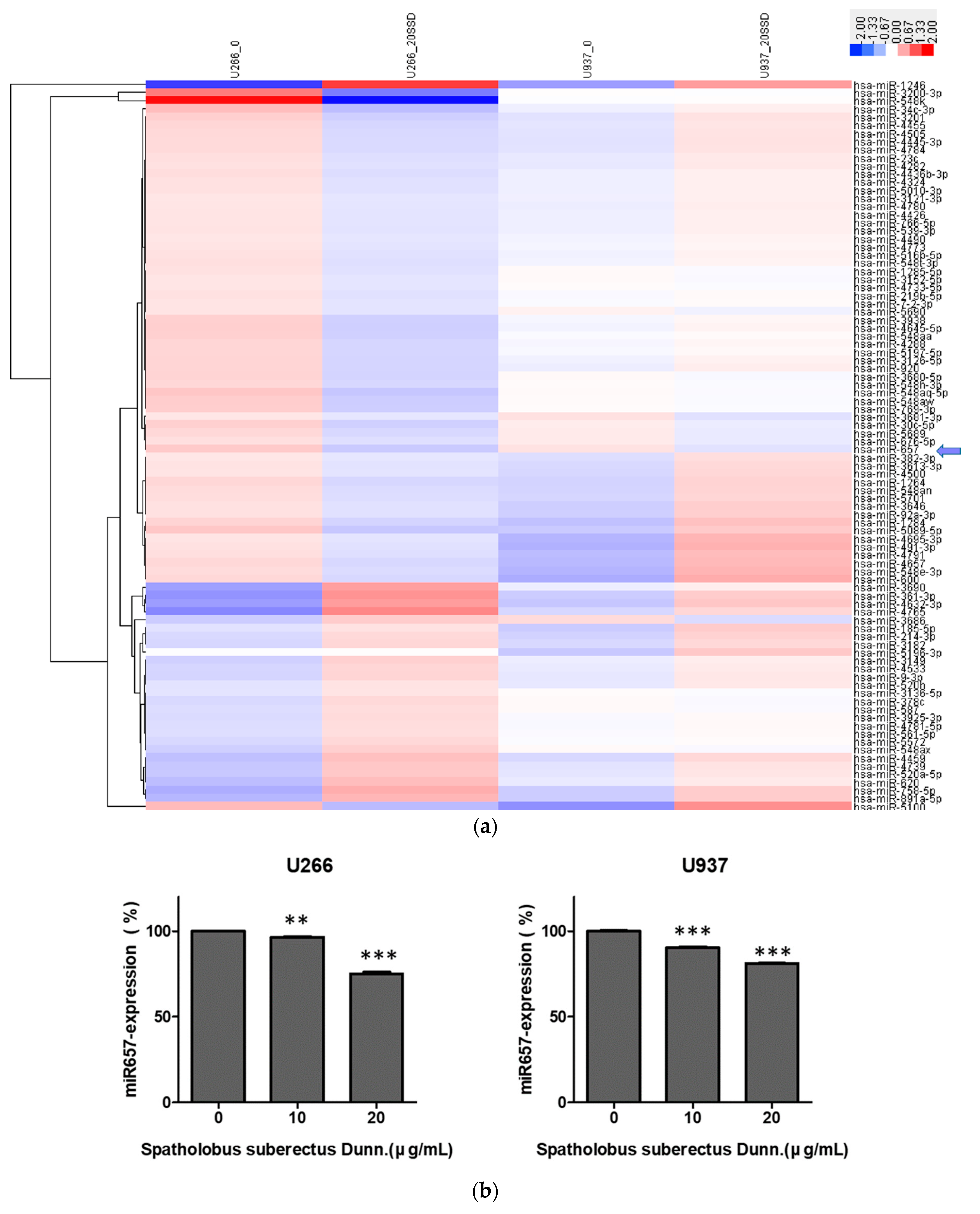
2.7. SSD Reduced the Expression of miR-657 in U266 and U937 Cells
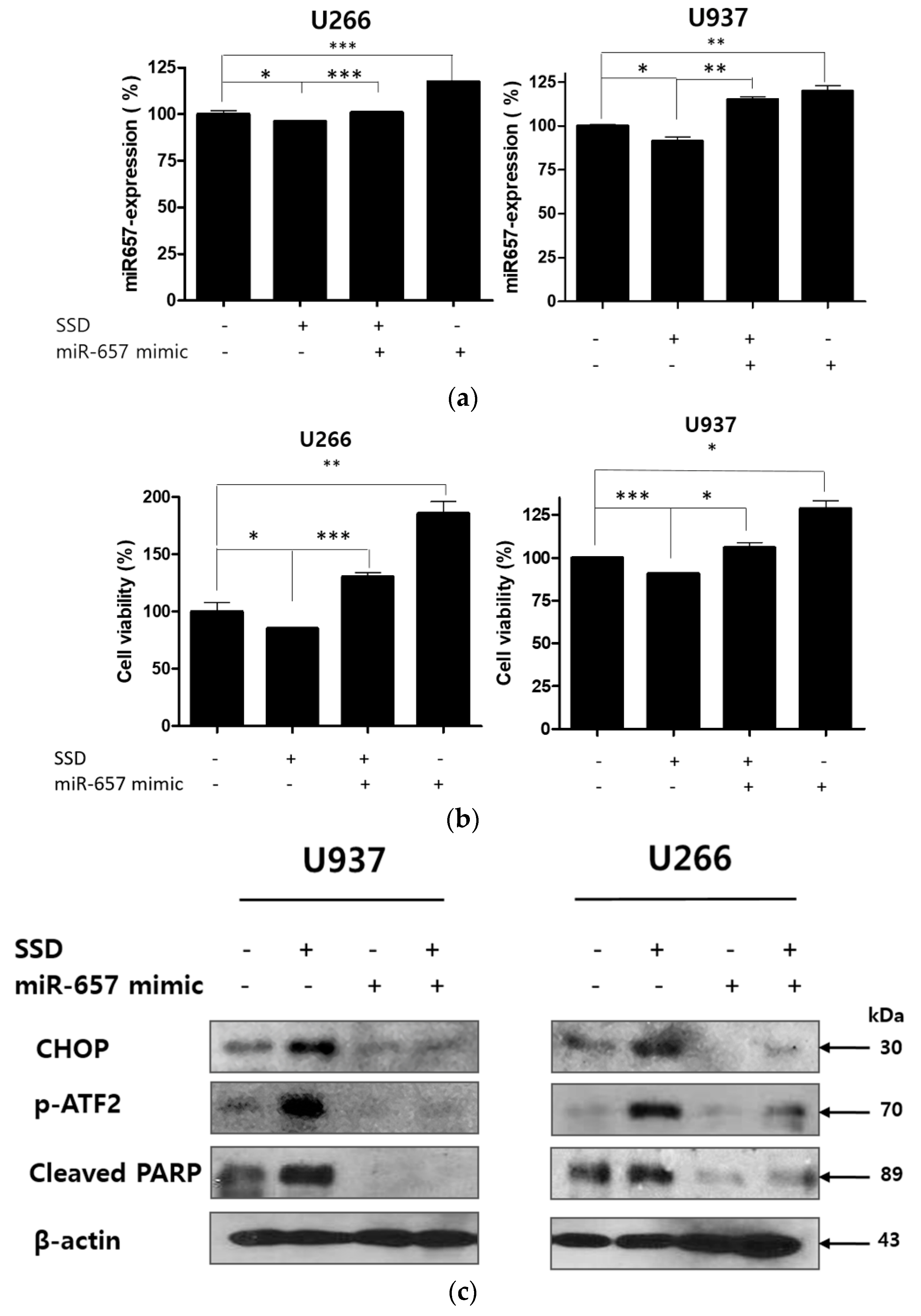
2.8. MiR-657 Plays a Role in SSD-Induced Apoptosis in U266 and U937 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. HPLC Analysis
4.3. Cell Culture
4.4. Cytotoxicity Assay
4.5. Western Blot Analysis
4.6. TUNEL Assay
4.7. Measurement of ROS
4.8. Quantitative Real-Time PCR Analyses
4.9. Microarray
4.10. Data Analysis for Microarray
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sant, M.; Allemani, C.; Tereanu, C.; De Angelis, R.; Capocaccia, R.; Visser, O.; Marcos-Gragera, R.; Maynadié, M.; Simonetti, A.; Lutz, J.-M. Incidence of hematologic malignancies in europe by morphologic subtype: Results of the haemacare project. Blood 2010, 116, 3724–3734. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Howell, D.; Patmore, R.; Jack, A.; Roman, E. Incidence of haematological malignancy by sub-type: A report from the haematological malignancy research network. Br. J. Cancer 2011, 105, 1684. [Google Scholar] [CrossRef] [PubMed]
- Kyle, R.A.; Rajkumar, S.V. Treatment of multiple myeloma: A comprehensive review. Clin. Lymphoma Myeloma 2009, 9, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Freeman, C.; Berg, J.W.; Cutler, S.J. Occurrence and prognosis of extranodal lymphomas. Cancer 1972, 29, 252–260. [Google Scholar] [CrossRef]
- Reed, J.C.; Pellecchia, M. Apoptosis-based therapies for hematologic malignancies. Blood 2005, 106, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. Bcl-2–family proteins and hematologic malignancies: History and future prospects. Blood 2008, 111, 3322–3330. [Google Scholar] [CrossRef] [PubMed]
- Herr, I.; Debatin, K.-M. Cellular stress response and apoptosis in cancer therapy. Blood 2001, 98, 2603–2614. [Google Scholar] [CrossRef]
- Reddy, N.; Czuczman, M. Enhancing activity and overcoming chemoresistance in hematologic malignancies with bortezomib: Preclinical mechanistic studies. Ann. Oncol. 2010, 21, 1756–1764. [Google Scholar] [CrossRef]
- Morishima, N.; Nakanishi, K.; Takenouchi, H.; Shibata, T.; Yasuhiko, Y. An er stress-specific caspase cascade in apoptosis: Cytochrome c-independent activation of caspase-9 by caspase-12. J. Biol. Chem. 2002, 277, 34287–34294. [Google Scholar] [CrossRef]
- Du, Y.; Wang, K.; Fang, H.; Li, J.; Xiao, D.; Zheng, P.; Chen, Y.; Fan, H.; Pan, X.; Zhao, C. Coordination of intrinsic, extrinsic, and endoplasmic reticulum-mediated apoptosis by imatinib mesylate combined with arsenic trioxide in chronic myeloid leukemia. Blood 2006, 107, 1582–1590. [Google Scholar] [CrossRef]
- Rosati, E.; Sabatini, R.; Rampino, G.; De Falco, F.; Di Ianni, M.; Falzetti, F.; Fettucciari, K.; Bartoli, A.; Screpanti, I.; Marconi, P. Novel targets for endoplasmic reticulum stress-induced apoptosis in b-cll. Blood 2010, 116, 2713–2723. [Google Scholar] [CrossRef] [PubMed]
- Sano, R.; Reed, J.C. Er stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Lim, W.; Bazer, F.W.; Song, G. Chrysin induces death of prostate cancer cells by inducing ros and er stress. J. Cell. Physiol. 2017, 232, 3786–3797. [Google Scholar] [CrossRef]
- Ozkaya, A.B.; Ak, H.; Aydin, H.H. High concentration calcitriol induces endoplasmic reticulum stress related gene profile in breast cancer cells. Biochem. Cell Biol. 2017, 95, 289–294. [Google Scholar] [CrossRef]
- Shamu, C.E.; Walter, P. Oligomerization and phosphorylation of the ire1p kinase during intracellular signaling from the endoplasmic reticulum to the nucleus. EMBO J. 1996, 15, 3028–3039. [Google Scholar] [CrossRef] [PubMed]
- Haze, K.; Yoshida, H.; Yanagi, H.; Yura, T.; Mori, K. Mammalian transcription factor atf6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol. Biol. Cell 1999, 10, 3787–3799. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Rawson, R.B.; Komuro, R.; Chen, X.; Dave, U.P.; Prywes, R.; Brown, M.S.; Goldstein, J.L. Er stress induces cleavage of membrane-bound atf6 by the same proteases that process srebps. Mol. Cell 2000, 6, 1355–1364. [Google Scholar] [CrossRef]
- Fels, D.R.; Koumenis, C. The perk/eif2alpha/atf4 module of the upr in hypoxia resistance and tumor growth. Cancer Biol. Ther. 2006, 5, 723–728. [Google Scholar] [CrossRef]
- Han, J.; Back, S.H.; Hur, J.; Lin, Y.H.; Gildersleeve, R.; Shan, J.; Yuan, C.L.; Krokowski, D.; Wang, S.; Hatzoglou, M.; et al. Er-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat. Cell Biol. 2013, 15, 481–490. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Farooqi, A.A.; Li, K.T.; Fayyaz, S.; Chang, Y.T.; Ismail, M.; Liaw, C.C.; Yuan, S.S.; Tang, J.Y.; Chang, H.W. Anticancer drugs for the modulation of endoplasmic reticulum stress and oxidative stress. Tumour Biol. 2015, 36, 5743–5752. [Google Scholar] [CrossRef] [PubMed]
- Belmont, P.J.; Chen, W.J.; Thuerauf, D.J.; Glembotski, C.C. Regulation of microrna expression in the heart by the atf6 branch of the er stress response. J. Mol. Cell. Cardiol. 2012, 52, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Upton, J.-P.; Wang, L.; Han, D.; Wang, E.S.; Huskey, N.E.; Lim, L.; Truitt, M.; McManus, M.T.; Ruggero, D.; Goga, A. Ire1α cleaves select micrornas during er stress to derepress translation of proapoptotic caspase-2. Science 2012, 338, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.-W.; Chang, T.-C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R. Therapeutic microrna delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Mendell, J. Micrornas in cell proliferation, cell death, and tumorigenesis. Br. J. Cancer 2006, 94, 776. [Google Scholar] [CrossRef] [PubMed]
- Szafranska, A.; Davison, T.; John, J.; Cannon, T.; Sipos, B.; Maghnouj, A.; Labourier, E.; Hahn, S. Microrna expression alterations are linked to tumorigenesis and non-neoplastic processes in pancreatic ductal adenocarcinoma. Oncogene 2007, 26, 4442. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Schuiki, I.; Zhang, L.; Allister, E.M.; Wheeler, M.B.; Volchuk, A. Sdf2l1 interacts with the er-associated degradation machinery and retards the degradation of mutant proinsulin in pancreatic beta-cells. J. Cell Sci. 2013, 126, 1962–1968. [Google Scholar] [CrossRef]
- Fu, Y.F.; Jiang, L.H.; Zhao, W.D.; Xi-Nan, M.; Huang, S.Q.; Yang, J.; Hu, T.J.; Chen, H.L. Immunomodulatory and antioxidant effects of total flavonoids of spatholobus suberectus dunn on pcv2 infected mice. Sci. Rep. 2017, 7, 8676. [Google Scholar] [CrossRef]
- Sun, J.Q.; Zhang, G.L.; Zhang, Y.; Nan, N.; Sun, X.; Yu, M.W.; Wang, H.; Li, J.P.; Wang, X.M. Spatholobus suberectus column extract inhibits estrogen receptor positive breast cancer via suppressing er mapk pi3k/akt pathway. Evid.-Based Complement. Altern. Med. 2016, 2016, 2934340. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, D.M.; Loo, T.Y.; Cheng, Y.; Chen, L.L.; Shen, J.G.; Yang, D.P.; Chow, L.W.; Guan, X.Y.; Chen, J.P. Spatholobus suberectus inhibits cancer cell growth by inducing apoptosis and arresting cell cycle at g2/m checkpoint. J. Ethnopharmacol. 2011, 133, 751–758. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, L.; Duan, L.; Dong, X.; Zhou, P.; Liu, E.H.; Li, P. Simultaneous determination of 16 phenolic constituents in spatholobi caulis by high performance liquid chromatography/electrospray ionization triple quadrupole mass spectrometry. J. Pharm. Biomed. Anal. 2015, 102, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Ren, Y.; Guan, L.; Wei, W.; Hua, F.; Yang, Y.; Yang, T.; Cao, T.; Dong, H.; Pan, H.; et al. Sodium selenite induces apoptosis in acute promyelocytic leukemia-derived nb4 cells through mitochondria-dependent pathway. Oncol. Res. 2009, 17, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.N.; Oldan, J.D. Pet/mr imaging of multiple myeloma. Magn. Reson. Imaging Clin. N. Am. 2017, 25, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhong, Z.; Zeng, C.; Meng, L.; Li, C.; Luo, Y.; Wang, H.; Li, W.; Wang, J.; Cheng, F. A clinical observation of chinese chronic myelogenous leukemia patients after discontinuation of tyrosine kinase inhibitors. Oncotarget 2016, 7, 58234. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.K.; Rajkumar, S.V.; Dispenzieri, A.; Lacy, M.Q.; Hayman, S.R.; Buadi, F.K.; Zeldenrust, S.R.; Dingli, D.; Russell, S.J.; Lust, J.A.; et al. Improved survival in multiple myeloma and the impact of novel therapies. Blood 2008, 111, 2516–2520. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Lee, H.J.; Vo, M.C.; Kim, H.J.; Lee, J.J. Immunotherapy for the treatment of multiple myeloma. Crit. Rev. Oncol./Hematol. 2017, 111, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Song, H.-S.; Park, H.; Kim, B. Activation of er stress dependent-mir-216b has critical role in salvia miltiorrhiza ethanol extract induced apoptosis in u266 and u937 cells. Int. J. Mol. Sci. 2018, 19, 1240. [Google Scholar] [CrossRef] [PubMed]
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute myeloid leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef]
- Gire, V.; Roux, P.; Wynford-Thomas, D.; Brondello, J.M.; Dulic, V. DNA damage checkpoint kinase chk2 triggers replicative senescence. EMBO J. 2004, 23, 2554–2563. [Google Scholar] [CrossRef]
- Nicholson, D.W.; Ali, A.; Thornberry, N.A.; Vaillancourt, J.P.; Ding, C.K.; Gallant, M.; Gareau, Y.; Griffin, P.R.; Labelle, M.; Lazebnik, Y.A.; et al. Identification and inhibition of the ice/ced-3 protease necessary for mammalian apoptosis. Nature 1995, 376, 37–43. [Google Scholar] [CrossRef]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of poly(adp-ribose) polymerase (parp) cleavage in apoptosis. Caspase 3-resistant parp mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Cao, Y.; Wei, W.; Liu, W.; Lu, S.Y.; Chen, Y.B.; Wang, Y.; Yan, H.; Wu, Y.L. Targeting poly (adp-ribose) polymerase partially contributes to bufalin-induced cell death in multiple myeloma cells. PLoS ONE 2013, 8, e66130. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Lee, H.J.; Shin, E.A.; Kim, H.; Seo, K.H.; Baek, N.I.; Kim, B.; Kim, S.H. C-jun n-terminal kinase-dependent endoplasmic reticulum stress pathway is critically involved in arjunic acid induced apoptosis in non-small cell lung cancer cells. Phytother. Res. 2016, 30, 596–603. [Google Scholar] [CrossRef]
- Kim, J.; Yun, M.; Kim, E.O.; Jung, D.B.; Won, G.; Kim, B.; Jung, J.H.; Kim, S.H. Decursin enhances trail-induced apoptosis through oxidative stress mediated- endoplasmic reticulum stress signalling in non-small cell lung cancers. Br. J. Pharmacol. 2016, 173, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Ozben, T. Oxidative stress and apoptosis: Impact on cancer therapy. J. Pharm. Sci. 2007, 96, 2181–2196. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y. N-acetylcysteine, reactive oxygen species and beyond. Cancer Biol. Ther. 2010, 9, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Guan, Y.; Sheng, H.; Liu, Y. Crosstalk in competing endogenous rna network reveals the complex molecular mechanism underlying lung cancer. Oncotarget 2017, 8, 91270–91280. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, S.K.; Zhang, K.; Zhang, H.; Pan, Q.; Liu, Z.; Pan, H.; Xue, L.; Yen, Y.; Chu, P.G. Expression of glypican 3 enriches hepatocellular carcinoma development-related genes and associates with carcinogenesis in cirrhotic livers. Carcinogenesis 2015, 36, 232–242. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, M.; Tao, Z.; Hua, Q.; Chen, S.; Xiao, B. Identification of predictive biomarkers for early diagnosis of larynx carcinoma based on microrna expression data. Cancer Genet. 2013, 206, 340–346. [Google Scholar] [CrossRef]
- Ding, H.; Wu, Y.L.; Wang, Y.X.; Zhu, F.F. Characterization of the microrna expression profile of cervical squamous cell carcinoma metastases. Asian Pac. J. Cancer Prev. 2014, 15, 1675–1679. [Google Scholar] [CrossRef]
- Kim, C.; Song, H.S.; Park, H.; Kim, B. Activation of er stress-dependent mir-216b has a critical role in salviamiltiorrhiza ethanol-extract-induced apoptosis in u266 and u937 cells. Int. J. Mol. Sci. 2018, 19. [Google Scholar]
- Cha, J.A.; Song, H.-S.; Kang, B.; Park, M.N.; Park, K.S.; Kim, S.-H.; Shim, B.-S.; Kim, B. Mir-211 plays a critical role in cnidium officinale makino extract-induced, ros/er stress-mediated apoptosis in u937 and u266 cells. Int. J. Mol. Sci. 2018, 19, 865. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.S.; Jeong, M.S.; Kim, B.; Kim, S.H. Antiangiogenic effect of ethanol extract of vigna angularis via inhibition of phosphorylation of vegfr2, erk, and akt. Evid.-Based Complement. Altern. Med. 2015, 2015, 371368. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Woo, M.J.; Park, C.S.; Lee, S.H.; Kim, J.S.; Kim, B.; An, S.; Kim, S.H. Hovenia dulcis extract reduces lipid accumulation in oleic acid-induced steatosis of hep g2 cells via activation of ampk and pparalpha/cpt-1 pathway and in acute hyperlipidemia mouse model. Phytother. Res. 2017, 31, 132–139. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inter-day | ||||
| Standards | SSD1 | |||
| RT 2 (min) | RSD 3 (%) | RT 2 (min) | SD 4 (%) | |
| EGC | 2.412 | 0.078 | 2.376 | 0.172 |
| Genistein | 13.88 | 0.102 | 13.913 | 0.500 |
| Intra-day | ||||
| Standards | SSD1 | |||
| RT 2 (min) | RSD 3 (%) | RT 2 (min) | SD 4 (%) | |
| EGC | 2.411 | 0.017 | 2.381 | 0.689 |
| Genistein | 13.888 | 0.689 | 13.844 | 0.900 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.J.; Park, M.N.; Kim, C.; Kang, B.; Song, H.-S.; Lee, H.; Kim, S.-H.; Shim, B.-S.; Kim, B. MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells. Cancers 2019, 11, 150. https://doi.org/10.3390/cancers11020150
Lim HJ, Park MN, Kim C, Kang B, Song H-S, Lee H, Kim S-H, Shim B-S, Kim B. MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells. Cancers. 2019; 11(2):150. https://doi.org/10.3390/cancers11020150
Chicago/Turabian StyleLim, Hyun Ji, Moon Nyeo Park, Changmin Kim, Beomku Kang, Hyo-Sook Song, Hyemin Lee, Sung-Hoon Kim, Bum-Sang Shim, and Bonglee Kim. 2019. "MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells" Cancers 11, no. 2: 150. https://doi.org/10.3390/cancers11020150
APA StyleLim, H. J., Park, M. N., Kim, C., Kang, B., Song, H.-S., Lee, H., Kim, S.-H., Shim, B.-S., & Kim, B. (2019). MiR-657/ATF2 Signaling Pathway Has a Critical Role in Spatholobus suberectus Dunn Extract-Induced Apoptosis in U266 and U937 Cells. Cancers, 11(2), 150. https://doi.org/10.3390/cancers11020150