The Role of DNA/Histone Modifying Enzymes and Chromatin Remodeling Complexes in Testicular Germ Cell Tumors




Abstract
1. Testicular Germ Cell Tumors in Brief
2. Protein-Coding Epigenetic Players: Their Role in Cancer
3. Protein-Coding Epigenetic Players in Testicular Germ Cell Tumors
3.1. DNA-MODIFYING ENZYMES
3.1.1. Methylation
DNA Methyltransferases (DNMTs)
DNA Demethylases (TETs)
3.2. HISTONE-MODIFYING ENZYMES
3.2.1. Acetylation
Lysine Acetyltransferases (KATs)
A. MYST Family
B. GNAT Family
C. Orphan Family
Lysine Deacetylases (KDACs)
A. Zn2+-Dependent Histone Deacetylases (HDACs)
B. NAD+-Dependent Sirtuin Deacetylases (SIRTs)
3.2.2. Methylation
Lysine Methyltransferases (KMTs)
A. SET Domain-Containing KMTs
B. DOT1-Like Family (DOT1L)
Arginine Methyltransferases (PRMTs)
Lysine Demethylases (KDMs)
A. Alpha-Ketoglutarate (2OG) and Fe2+-Dependent, Jumonji (JmjC) Domain-Containing Demethylases
B. FAD-Dependent Amine Oxidase Demethylases (LSDs)
3.2.3. Phosphorylation
Serine/Threonine/Tyrosine Kinases
3.2.4. Ubiquitination
Ubiquitin Ligases
Deubiquitinating Enzymes
3.3. CHROMATIN REMODELING ENZYMES
3.3.1. SWI/SNF Family
3.3.2. ISWI Family
3.3.3. CHD Family
3.3.4. INO80 Family
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef]
- Lobo, J.; Costa, A.L.; Vilela-Salgueiro, B.; Rodrigues, A.; Guimaraes, R.; Cantante, M.; Lopes, P.; Antunes, L.; Jeronimo, C.; Henrique, R. Testicular germ cell tumors: Revisiting a series in light of the new WHO classification and AJCC staging systems, focusing on challenges for pathologists. Hum. Pathol. 2018, 82, 113–124. [Google Scholar] [CrossRef]
- Trabert, B.; Chen, J.; Devesa, S.S.; Bray, F.; McGlynn, K.A. International patterns and trends in testicular cancer incidence, overall and by histologic subtype, 1973–2007. Andrology 2015, 3, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Beyer, J.; Albers, P.; Altena, R.; Aparicio, J.; Bokemeyer, C.; Busch, J.; Cathomas, R.; Cavallin-Stahl, E.; Clarke, N.W.; Classen, J.; et al. Maintaining success, reducing treatment burden, focusing on survivorship: Highlights from the third European consensus conference on diagnosis and treatment of germ-cell cancer. Ann. Oncol. 2013, 24, 878–888. [Google Scholar] [CrossRef]
- Henrique, R.; Jeronimo, C. Testicular Germ Cell Tumors Go Epigenetics: Will miR-371a-3p Replace Classical Serum Biomarkers? Eur. Urol. 2017, 71, 221–222. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Lobo, J.; Jeronimo, C.; Henrique, R. The epigenetics of testicular germ cell tumors: Looking for novel disease biomarkers. Epigenomics 2017, 9, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Buljubasic, R.; Buljubasic, M.; Bojanac, A.K.; Ulamec, M.; Vlahovic, M.; Jezek, D.; Bulic-Jakus, F.; Sincic, N. Epigenetics and testicular germ cell tumors. Gene 2018, 661, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Looijenga, L.H.; Stoop, H.; Biermann, K. Testicular cancer: Biology and biomarkers. Virchows Arch. 2014, 464, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Van Der Zwan, Y.G.; Stoop, H.; Rossello, F.; White, S.J.; Looijenga, L.H. Role of epigenetics in the etiology of germ cell cancer. Int. J. Dev. Biol. 2013, 57, 299–308. [Google Scholar] [CrossRef]
- Kristensen, D.G.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Almstrup, K. Epigenetic features of testicular germ cell tumours in relation to epigenetic characteristics of foetal germ cells. Int. J. Dev. Biol. 2013, 57, 309–317. [Google Scholar] [CrossRef]
- Murray, M.J.; Huddart, R.A.; Coleman, N. The present and future of serum diagnostic tests for testicular germ cell tumours. Nat. Rev. Urol. 2016, 13, 715–725. [Google Scholar] [CrossRef] [PubMed]
- DesJarlais, R.; Tummino, P.J. Role of Histone-Modifying Enzymes and Their Complexes in Regulation of Chromatin Biology. Biochemistry 2016, 55, 1584–1599. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Arfuso, F.; Arumugam, S.; Chinnathambi, A.; Jinsong, B.; Warrier, S.; Wang, L.Z.; Kumar, A.P.; Ahn, K.S.; Sethi, G.; et al. Role of novel histone modifications in cancer. Oncotarget 2018, 9, 11414–11426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, J. DNA methyltransferases and their roles in tumorigenesis. Biomark Res. 2017, 5, 1. [Google Scholar] [CrossRef]
- Tyagi, M.; Imam, N.; Verma, K.; Patel, A.K. Chromatin remodelers: We are the drivers!! Nucleus 2016, 7, 388–404. [Google Scholar] [CrossRef]
- Arrowsmith, C.H.; Bountra, C.; Fish, P.V.; Lee, K.; Schapira, M. Epigenetic protein families: A new frontier for drug discovery. Nat. Rev. Drug. Discov. 2012, 11, 384–400. [Google Scholar] [CrossRef]
- Lobo, J.; Rodrigues, A.; Antunes, L.; Graca, I.; Ramalho-Carvalho, J.; Vieira, F.Q.; Martins, A.T.; Oliveira, J.; Jeronimo, C.; Henrique, R. High immunoexpression of Ki67, EZH2, and SMYD3 in diagnostic prostate biopsies independently predicts outcome in patients with prostate cancer. Urol. Oncol. 2018, 36, 161.e7–161.e17. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Pires-Luis, A.S.; Vieira-Coimbra, M.; Costa-Pinheiro, P.; Antunes, L.; Dias, P.C.; Lobo, F.; Oliveira, J.; Goncalves, C.S.; Costa, B.M.; et al. SETDB2 and RIOX2 are differentially expressed among renal cell tumor subtypes, associating with prognosis and metastization. Epigenetics 2017, 12, 1057–1064. [Google Scholar] [CrossRef]
- Junqueira-Neto, S.; Vieira, F.Q.; Montezuma, D.; Costa, N.R.; Antunes, L.; Baptista, T.; Oliveira, A.I.; Graca, I.; Rodrigues, A.; Magalhaes, J.S.; et al. Phenotypic impact of deregulated expression of class I histone deacetylases in urothelial cell carcinoma of the bladder. Mol. Carcinog. 2015, 54, 523–531. [Google Scholar] [CrossRef]
- Carvalho, S.; Freitas, M.; Antunes, L.; Monteiro-Reis, S.; Vieira-Coimbra, M.; Tavares, A.; Paulino, S.; Videira, J.F.; Jeronimo, C.; Henrique, R. Prognostic value of histone marks H3K27me3 and H3K9me3 and modifying enzymes EZH2, SETDB1 and LSD-1 in colorectal cancer. J. Cancer Res. Clin. Oncol. 2018. [Google Scholar] [CrossRef]
- Roberts, C.W.; Orkin, S.H. The SWI/SNF complex--chromatin and cancer. Nat. Rev. Cancer 2004, 4, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Langst, G.; Manelyte, L. Chromatin Remodelers: From Function to Dysfunction. Genes (Basel) 2015, 6, 299–324. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Koutsogiannouli, E.A.; Wagner, N.; Hader, C.; Pinkerneil, M.; Hoffmann, M.J.; Schulz, W.A. Differential Effects of Histone Acetyltransferase GCN5 or PCAF Knockdown on Urothelial Carcinoma Cells. Int. J. Mol. Sci. 2017, 18, 1449. [Google Scholar] [CrossRef] [PubMed]
- Skulte, K.A.; Phan, L.; Clark, S.J.; Taberlay, P.C. Chromatin remodeler mutations in human cancers: Epigenetic implications. Epigenomics 2014, 6, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Markulin, D.; Vojta, A.; Samarzija, I.; Gamulin, M.; Beceheli, I.; Jukic, I.; Maglov, C.; Zoldos, V.; Fucic, A. Association Between RASSF1A Promoter Methylation and Testicular Germ Cell Tumor: A Meta-analysis and a Cohort Study. Cancer Genom. Proteom. 2017, 14, 363–372. [Google Scholar] [CrossRef]
- Martinelli, C.; Lengert, A.V.H.; Carcano, F.M.; Silva, E.C.A.; Brait, M.; Lopes, L.F.; Vidal, D.O. MGMT and CALCA promoter methylation are associated with poor prognosis in testicular germ cell tumor patients. Oncotarget 2017, 8, 50608–50617. [Google Scholar] [CrossRef]
- Spiller, C.M.; Gillis, A.J.; Burnet, G.; Stoop, H.; Koopman, P.; Bowles, J.; Looijenga, L.H. Cripto: Expression, epigenetic regulation and potential diagnostic use in testicular germ cell tumors. Mol. Oncol. 2016, 10, 526–537. [Google Scholar] [CrossRef]
- Rijlaarsdam, M.A.; Tax, D.M.; Gillis, A.J.; Dorssers, L.C.; Koestler, D.C.; de Ridder, J.; Looijenga, L.H. Genome wide DNA methylation profiles provide clues to the origin and pathogenesis of germ cell tumors. PLoS ONE 2015, 10, e0122146. [Google Scholar] [CrossRef]
- Vilela-Salgueiro, B.; Barros-Silva, D.; Lobo, J.; Costa, A.L.; Guimaraes, R.; Cantante, M.; Lopes, P.; Braga, I.; Oliveira, J.; Henrique, R.; et al. Germ cell tumour subtypes display differential expression of microRNA371a-3p. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- Van Agthoven, T.; Eijkenboom, W.M.H.; Looijenga, L.H.J. microRNA-371a-3p as informative biomarker for the follow-up of testicular germ cell cancer patients. Cell. Oncol. (Dordr) 2017, 40, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Leao, R.; van Agthoven, T.; Figueiredo, A.; Jewett, M.A.S.; Fadaak, K.; Sweet, J.; Ahmad, A.E.; Anson-Cartwright, L.; Chung, P.; Hansen, A.; et al. Serum miRNA Predicts Viable Disease after Chemotherapy in Patients with Testicular Nonseminoma Germ Cell Tumor. J. Urol. 2018, 200, 126–135. [Google Scholar] [CrossRef]
- Anheuser, P.; Radtke, A.; Wulfing, C.; Kranz, J.; Belge, G.; Dieckmann, K.P. Serum Levels of MicroRNA371a-3p: A Highly Sensitive Tool for Diagnosing and Staging Testicular Germ Cell Tumours: A Clinical Case Series. Urol. Int. 2017, 99, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Almstrup, K.; Nielsen, J.E.; Mlynarska, O.; Jansen, M.T.; Jorgensen, A.; Skakkebaek, N.E.; Rajpert-De Meyts, E. Carcinoma in situ testis displays permissive chromatin modifications similar to immature foetal germ cells. Br. J. Cancer 2010, 103, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Eckert, D.; Biermann, K.; Nettersheim, D.; Gillis, A.J.; Steger, K.; Jack, H.M.; Muller, A.M.; Looijenga, L.H.; Schorle, H. Expression of BLIMP1/PRMT5 and concurrent histone H2A/H4 arginine 3 dimethylation in fetal germ cells, CIS/IGCNU and germ cell tumors. BMC Dev. Biol. 2008, 8, 106. [Google Scholar] [CrossRef]
- Beyrouthy, M.J.; Garner, K.M.; Hever, M.P.; Freemantle, S.J.; Eastman, A.; Dmitrovsky, E.; Spinella, M.J. High DNA methyltransferase 3B expression mediates 5-aza-deoxycytidine hypersensitivity in testicular germ cell tumors. Cancer Res. 2009, 69, 9360–9366. [Google Scholar] [CrossRef]
- Hinz, S.; Magheli, A.; Weikert, S.; Schulze, W.; Krause, H.; Schrader, M.; Miller, K.; Kempkensteffen, C. Deregulation of EZH2 expression in human spermatogenic disorders and testicular germ cell tumors. World J. Urol. 2010, 28, 631–635. [Google Scholar] [CrossRef]
- Wang, J.; Lu, F.; Ren, Q.; Sun, H.; Xu, Z.; Lan, R.; Liu, Y.; Ward, D.; Quan, J.; Ye, T.; et al. Novel histone demethylase LSD1 inhibitors selectively target cancer cells with pluripotent stem cell properties. Cancer Res. 2011, 71, 7238–7249. [Google Scholar] [CrossRef]
- Arai, E.; Nakagawa, T.; Wakai-Ushijima, S.; Fujimoto, H.; Kanai, Y. DNA methyltransferase 3B expression is associated with poor outcome of stage I testicular seminoma. Histopathology 2012, 60, E12–E18. [Google Scholar] [CrossRef]
- Omisanjo, O.A.; Biermann, K.; Hartmann, S.; Heukamp, L.C.; Sonnack, V.; Hild, A.; Brehm, R.; Bergmann, M.; Weidner, W.; Steger, K. DNMT1 and HDAC1 gene expression in impaired spermatogenesis and testicular cancer. Histochem. Cell. Biol. 2007, 127, 175–181. [Google Scholar] [CrossRef]
- Yamada, S.; Kohu, K.; Ishii, T.; Ishidoya, S.; Hiramatsu, M.; Kanto, S.; Fukuzaki, A.; Adachi, Y.; Endoh, M.; Moriya, T.; et al. Gene expression profiling identifies a set of transcripts that are up-regulated inhuman testicular seminoma. DNA Res. 2004, 11, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Minami, K.; Chano, T.; Kawakami, T.; Ushida, H.; Kushima, R.; Okabe, H.; Okada, Y.; Okamoto, K. DNMT3L is a novel marker and is essential for the growth of human embryonal carcinoma. Clin. Cancer Res. 2010, 16, 2751–2759. [Google Scholar] [CrossRef] [PubMed]
- Bartkova, J.; Bakkenist, C.J.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Sehested, M.; Lukas, J.; Kastan, M.B.; Bartek, J. ATM activation in normal human tissues and testicular cancer. Cell. Cycle 2005, 4, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Chieffi, P.; Troncone, G.; Caleo, A.; Libertini, S.; Linardopoulos, S.; Tramontano, D.; Portella, G. Aurora B expression in normal testis and seminomas. J. Endocrinol. 2004, 181, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Benesova, M.; Trejbalova, K.; Kucerova, D.; Vernerova, Z.; Hron, T.; Szabo, A.; Amouroux, R.; Klezl, P.; Hajkova, P.; Hejnar, J. Overexpression of TET dioxygenases in seminomas associates with low levels of DNA methylation and hydroxymethylation. Mol. Carcinog. 2017, 56, 1837–1850. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, D.G.; Nielsen, J.E.; Jorgensen, A.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Almstrup, K. Evidence that active demethylation mechanisms maintain the genome of carcinoma in situ cells hypomethylated in the adult testis. Br. J. Cancer 2014, 110, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Kawai, K.; Ando, S.; Sugita, S.; Kandori, S.; Kojima, T.; Miyazaki, J.; Nishiyama, H. DNA methyltransferase-3 like protein expression in various histological types of testicular germ cell tumor. Jpn. J. Clin. Oncol. 2016, 46, 475–481. [Google Scholar] [CrossRef]
- Chen, B.F.; Suen, Y.K.; Gu, S.; Li, L.; Chan, W.Y. A miR-199a/miR-214 self-regulatory network via PSMD10, TP53 and DNMT1 in testicular germ cell tumor. Sci. Rep. 2014, 4, 6413. [Google Scholar] [CrossRef]
- Ueda, J.; Ho, J.C.; Lee, K.L.; Kitajima, S.; Yang, H.; Sun, W.; Fukuhara, N.; Zaiden, N.; Chan, S.L.; Tachibana, M.; et al. The hypoxia-inducible epigenetic regulators Jmjd1a and G9a provide a mechanistic link between angiogenesis and tumor growth. Mol. Cell. Biol. 2014, 34, 3702–3720. [Google Scholar] [CrossRef]
- Chen, B.F.; Gu, S.; Suen, Y.K.; Li, L.; Chan, W.Y. microRNA-199a-3p, DNMT3A, and aberrant DNA methylation in testicular cancer. Epigenetics 2014, 9, 119–128. [Google Scholar] [CrossRef]
- Ishii, T.; Kohu, K.; Yamada, S.; Ishidoya, S.; Kanto, S.; Fuji, H.; Moriya, T.; Satake, M.; Arai, Y. Up-regulation of DNA-methyltransferase 3A expression is associated with hypomethylation of intron 25 in human testicular germ cell tumors. Tohoku J. Exp. Med. 2007, 212, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.J.; Wang, Z.; Chiriboga, L.; Greco, M.A.; Shapiro, E.; Huang, H.; Yang, X.J.; Huang, J.; Peng, Y.; Melamed, J.; et al. The expression and function of androgen receptor coactivator p44 and protein arginine methyltransferase 5 in the developing testis and testicular tumors. J. Urol. 2007, 177, 1918–1922. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche, F.R.; Hasler, A.; Bode, P.K.; Adams, H.; Seifert, H.H.; Sulser, T.; Moch, H.; Barghorn, A.; Kristiansen, G. Expression of histone deacetylases 1, 2 and 3 in histological subtypes of testicular germ cell tumours. Histol. Histopathol. 2011, 26, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Lan, R.; Zhang, X.; Zhu, L.; Chen, F.; Xu, Z.; Liu, Y.; Ye, T.; Sun, H.; Lu, F.; et al. LSD1 regulates pluripotency of embryonic stem/carcinoma cells through histone deacetylase 1-mediated deacetylation of histone H4 at lysine 16. Mol. Cell. Biol. 2014, 34, 158–179. [Google Scholar] [CrossRef] [PubMed]
- Di Zazzo, E.; Porcile, C.; Bartollino, S.; Moncharmont, B. Critical Function of PRDM2 in the Neoplastic Growth of Testicular Germ Cell Tumors. Biology (Basel) 2016, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Baptissart, M.; Caira, F.; Brugnon, F.; Lobaccaro, J.M.; Volle, D.H. Epigenetic: A molecular link between testicular cancer and environmental exposures. Front. Endocrinol. (Lausanne) 2012, 3, 150. [Google Scholar] [CrossRef]
- Liu, S.; Brind’Amour, J.; Karimi, M.M.; Shirane, K.; Bogutz, A.; Lefebvre, L.; Sasaki, H.; Shinkai, Y.; Lorincz, M.C. Setdb1 is required for germline development and silencing of H3K9me3-marked endogenous retroviruses in primordial germ cells. Genes Dev. 2014, 28, 2041–2055. [Google Scholar] [CrossRef]
- Zhou, Z.; Yang, X.; He, J.; Liu, J.; Wu, F.; Yu, S.; Liu, Y.; Lin, R.; Liu, H.; Cui, Y.; et al. Kdm2b Regulates Somatic Reprogramming through Variant PRC1 Complex-Dependent Function. Cell. Rep. 2017, 21, 2160–2170. [Google Scholar] [CrossRef]
- Yamamoto, T.; Takenaka, C.; Yoda, Y.; Oshima, Y.; Kagawa, K.; Miyajima, H.; Sasaki, T.; Kawamata, S. Differentiation potential of Pluripotent Stem Cells correlates to the level of CHD7. Sci. Rep. 2018, 8, 241. [Google Scholar] [CrossRef]
- Deniz, O.; de la Rica, L.; Cheng, K.C.L.; Spensberger, D.; Branco, M.R. SETDB1 prevents TET2-dependent activation of IAP retroelements in naive embryonic stem cells. Genome Biol. 2018, 19, 6. [Google Scholar] [CrossRef]
- Del Mazo, J.; Garcia-Lopez, J.; Weber, M. Epigenetic traits of testicular cancer: From primordial germ cells to germ cell tumors. Epigenomics 2014, 6, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.M.; de Miguel, M.P.; Rebel, V.I.; Donovan, P.J. Identifying genes differentially expressed between PGCs and ES cells reveals a role for CREB-binding protein in germ cell survival. Dev. Biol. 2007, 311, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef]
- Schisterman, E.F.; Perkins, N.J.; Liu, A.; Bondell, H. Optimal cut-point and its corresponding Youden Index to discriminate individuals using pooled blood samples. Epidemiology 2005, 16, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Voss, A.K.; Thomas, T. MYST family histone acetyltransferases take center stage in stem cells and development. Bioessays 2009, 31, 1050–1061. [Google Scholar] [CrossRef]
- Sapountzi, V.; Cote, J. MYST-family histone acetyltransferases: Beyond chromatin. Cell. Mol. Life Sci. 2011, 68, 1147–1156. [Google Scholar] [CrossRef]
- Nagy, Z.; Tora, L. Distinct GCN5/PCAF-containing complexes function as co-activators and are involved in transcription factor and global histone acetylation. Oncogene 2007, 26, 5341–5357. [Google Scholar] [CrossRef]
- Di Martile, M.; Del Bufalo, D.; Trisciuoglio, D. The multifaceted role of lysine acetylation in cancer: Prognostic biomarker and therapeutic target. Oncotarget 2016, 7, 55789–55810. [Google Scholar] [CrossRef]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell. Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef]
- Di Cerbo, V.; Schneider, R. Cancers with wrong HATs: The impact of acetylation. Brief. Funct. Genom. 2013, 12, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Parbin, S.; Kar, S.; Shilpi, A.; Sengupta, D.; Deb, M.; Rath, S.K.; Patra, S.K. Histone deacetylases: A saga of perturbed acetylation homeostasis in cancer. J. Histochem. Cytochem. 2014, 62, 11–33. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.; Chen, W.Y. Sorting out functions of sirtuins in cancer. Oncogene 2014, 33, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zhang, S.; Liu, H.M.; Zhang, Y.B.; Blair, C.A.; Mercola, D.; Sassone-Corsi, P.; Zi, X. Histone lysine-specific methyltransferases and demethylases in carcinogenesis: New targets for cancer therapy and prevention. Curr. Cancer Drug Targets 2013, 13, 558–579. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.; Trojer, P. Targeting histone lysine methylation in cancer. Pharmacol. Ther. 2015, 150, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Choi, M.; Kim, J.E. The histone methyltransferase Dot1/DOT1L as a critical regulator of the cell cycle. Cell. Cycle 2014, 13, 726–738. [Google Scholar] [CrossRef]
- Yang, Y.; Bedford, M.T. Protein arginine methyltransferases and cancer. Nat. Rev. Cancer 2013, 13, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Park, J.W.; Chun, Y.S. Jumonji histone demethylases as emerging therapeutic targets. Pharmacol. Res. 2016, 105, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Minucci, S. A comprehensive review of lysine-specific demethylase 1 and its roles in cancer. Epigenomics 2017, 9, 1123–1142. [Google Scholar] [CrossRef]
- Rossetto, D.; Avvakumov, N.; Cote, J. Histone phosphorylation: A chromatin modification involved in diverse nuclear events. Epigenetics 2012, 7, 1098–1108. [Google Scholar] [CrossRef]
- Cao, J.; Yan, Q. Histone ubiquitination and deubiquitination in transcription, DNA damage response, and cancer. Front. Oncol. 2012, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Meas, R.; Mao, P. Histone ubiquitylation and its roles in transcription and DNA damage response. DNA Repair (Amst) 2015, 36, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Masliah-Planchon, J.; Bieche, I.; Guinebretiere, J.M.; Bourdeaut, F.; Delattre, O. SWI/SNF chromatin remodeling and human malignancies. Annu. Rev. Pathol. 2015, 10, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, B. ISWI chromatin remodeling: One primary actor or a coordinated effort? Curr. Opin. Struct. Biol. 2014, 24, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Stanley, F.K.; Moore, S.; Goodarzi, A.A. CHD chromatin remodelling enzymes and the DNA damage response. Mutat. Res. 2013, 750, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell. Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef]
- Todenhofer, T.; Hennenlotter, J.; Dorner, N.; Kuhs, U.; Aufderklamm, S.; Rausch, S.; Bier, S.; Mischinger, J.; Schellbach, D.; Hauch, S.; et al. Transcripts of circulating tumor cells detected by a breast cancer-specific platform correlate with clinical stage in bladder cancer patients. J. Cancer Res. Clin. Oncol. 2016, 142, 1013–1020. [Google Scholar] [CrossRef]
- Stein, U.; Burock, S.; Herrmann, P.; Wendler, I.; Niederstrasser, M.; Wernecke, K.D.; Schlag, P.M. Circulating MACC1 transcripts in colorectal cancer patient plasma predict metastasis and prognosis. PLoS ONE 2012, 7, e49249. [Google Scholar] [CrossRef]
- Burock, S.; Herrmann, P.; Wendler, I.; Niederstrasser, M.; Wernecke, K.D.; Stein, U. Circulating Metastasis Associated in Colon Cancer 1 transcripts in gastric cancer patient plasma as diagnostic and prognostic biomarker. World J. Gastroenterol. 2015, 21, 333–341. [Google Scholar] [CrossRef]
- Genc, C.G.; Jilesen, A.P.J.; Nieveen van Dijkum, E.J.M.; Klumpen, H.J.; van Eijck, C.H.J.; Drozdov, I.; Malczewska, A.; Kidd, M.; Modlin, I. Measurement of circulating transcript levels (NETest) to detect disease recurrence and improve follow-up after curative surgical resection of well-differentiated pancreatic neuroendocrine tumors. J. Surg. Oncol. 2018, 118, 37–48. [Google Scholar] [CrossRef]
- Marques-Magalhaes, A.; Graca, I.; Henrique, R.; Jeronimo, C. Targeting DNA Methyltranferases in Urological Tumors. Front. Pharmacol. 2018, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Heerboth, S.; Lapinska, K.; Snyder, N.; Leary, M.; Rollinson, S.; Sarkar, S. Use of epigenetic drugs in disease: An overview. Genet. Epigenet. 2014, 6, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Bae, S.C. Histone deacetylase inhibitors: Molecular mechanisms of action and clinical trials as anti-cancer drugs. Am. J. Transl. Res. 2011, 3, 166–179. [Google Scholar] [PubMed]
- Wapenaar, H.; Dekker, F.J. Histone acetyltransferases: Challenges in targeting bi-substrate enzymes. Clin. Epigenet. 2016, 8, 59. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Family | Epigenetic Players | Major Findings | Sample Type and Size | Methodology | Author |
|---|---|---|---|---|---|
| Histone Kinases | ATM | pS-ATM constitutively detected in TGCTs (↑↑ in ECs) and normal testis | TGCTs and normal testes (FFPE) | IHC | Bartkova et al. 2005 [43] |
| AURKB | ↑ in SEs (vs. normal testis) correlated with Ki67 proliferation index | 10 SEs (FFPE) | IHC | Chieffi et al. 2004 [44] | |
| DNA Methyltransferases | DNMT1 | DNMT1 ↑↑ in ECs and ↑ in TEs (vs. absent in SEs); | 32 TGCTs (FFPE) | RT-PCR IHC ISH | Omisanjo et al. 2007 [40] |
| ↑ in ECs (vs. normal testis) Evidence of a miR-199a/miR-214/PSMD10/TP53/DNMT1 self-regulatory pathway | 9 ECs + 12 normal testes (genomic RNA samples) TGCT cell lines | RT-PCR MSP WB | Chen et al. 2014 [48] | ||
| DNMT3A | ↑ in TGCTs (vs. normal testis) and associates with hypomethylation of intron 25 | 20 TGCTs + 9 adjacent tissue + 1 normal testis (FFPE and frozen tissue) | PCR IB/SB IHC | Ishii et al. 2007 [51] | |
| ↑ in SEs (vs. normal testis) | 8 SEs (frozen tissue) | Oligonucleotide-based microarray RT-PCR IHC | Yamada et al. 2004 [41] | ||
| ↑ in ECs (vs. normal testis); DNMT3A is the target of miR-199a-3p | 11 TGCTs + 14 normal testes (genomic RNA samples) TGCT cell lines | RT-PCR MSP/BSP WB | Chen et al. 2014 [50] | ||
| DNMT3B | ↑ in stage III SEs; ↑ associates with poorer relapse-free survival | 88 SEs (FFPE) | IHC | Arai et al. 2012 [39] | |
| ↑ in ECs (vs. somatic solid tumors) ↑ leads to 5AZA hypersensitivity in ECs (vs. somatic tumors) | TGCT cell lines | RT-PCR WB | Beyrouthy et al. 2009 [36] | ||
| DNMT3L | ↑ in ECs (vs. other TGCT subtypes); ↑ in advanced stage SEs | TMAs (n = 83 TGCTs) | IHC | Matsuoka et al. 2016 [47] | |
| ↑ in ECs (vs. somatic tumors and vs. other TGCT subtypes) | 53 TGCTs (FFPE, 43 with frozen tissue) TGCT cell lines | RT-PCR IHC WB | Minami et al. 2010 [42] | ||
| Histone Methyltransferases | EHMT2 | Its loss results in decreased tumor growth | TMA (67 TGCTs + 13 adjacent tissue + 4 normal testes) Cell lines Animal models | RT-PCR WB ChIP IHC | Ueda et al. 2014 [49] |
| EZH2 | ↓ in GCNIS and TGCTs (vs. normal testis) No significant differences between SEs vs. NSTs | 100 TGCTs + 4 GCNIS (frozen tissue) | RT-PCR | Hinz et al. 2010 [37] | |
| EZH2 is expressed in GCNIS cells, but only in the cytoplasm | TGCTs and GCNIS (FFPE) | IHC | Almstrup et al. 2010 [34] | ||
| Histone Deacetylases | HDAC1/2/3 | All 3 HDACs ↑ in CHs No associations with prognostic features | TMA (n = 325 TGCTs) | IHC | Fritzsche et al. 2011 [53] |
| HDAC1 | HDAC1 is expressed at low levels in TGCTs | 32 TGCTs (FFPE) | RT-PCR IHC ISH | Omisanjo et al. 2007 [40] | |
| HDAC1 regulates EC cells proliferation by establishing H4K16; HDAC1 ↑ in EC pluripotent cells (vs. non-pluripotent cells) | TGCT cell lines | RT-PCR ChIP WB | Yin et al. 2014 [54] | ||
| Histone Demethylases | JMJD1A | ↓ in TGCTs (vs. normal testis); Its loss results in increased tumor growth | TMA (67 TGCTs + 13 adjacent tissue + 4 normal testes) Cell lines Animal models | RT-PCR WB ChIP IHC | Ueda et al. 2014 [49] |
| JMJD3 | JMJD3 is absent in GCNIS | TGCTs and GCNIS (FFPE) | IHC | Almstrup et al. 2010 [34] | |
| KDM6A | KDM6A is absent in GCNIS | TGCTs and GCNIS (FFPE) | IHC | Almstrup et al. 2010 [34] | |
| LSD1 | ↑ in SEs (vs. normal testis) and pluripotent TGCT cells; LSD1 inhibitors and LSD1 knockdown impeded proliferation of pluripotent TGCT cells (vs. somatic tumors) | TMAs (n = 6 SEs) TGCT cell lines | WB IHC | Wang et al. 2011 [38] | |
| LSD1 regulates EC cells proliferation by establishing H4K16; LSD1 ↑ in EC pluripotent cells (vs. non-pluripotent cells) | TGCT cell lines | RT-PCR ChIP WB | Yin et al. 2014 [54] | ||
| Histone Methyltransferases | PRDM1 and PRMT5 | H2AR3me2 and H4R3me2 establishment; ↑ co-expression in GCNIS and SEs (vs. NSTs) → silencing of differentiation-related genes | 46 TGCTs + 15 GCNIS tissue (FFPE), 17 frozen tissue TGCT cell lines | Array RT-PCR Co-IP WB IHC | Eckert et al. 2008 [35] |
| PRDM2 | PRDM2 binds ER-α and influences proliferation, survival and apoptosis | TGCT cell lines | RT-PCR WB IP | Zazzo et al. 2016 [55] | |
| PRMT5 | PRMT5 colocalizes with p44 (AR coactivator) PRMT5 ↓ in the nucleus and ↑ in the cytoplasm (vs. normal testis) | 33 SEs + 9 normal testes + 11 LCTs (FFPE) | IHC | Liang et al. 2006 [52] | |
| DNA Demethylases | TET1/2 | TETs ↓ in GCNIS | TGCTs + normal testes (FFPE and frozen tissue) TGCT cell lines | RT-PCR ELISA WB IHC/IF | Kristensen et al. 2014 [46] |
| TET1 | ↑ in SEs (vs. normal testis and vs. NSTs) | 47 TGCTs + 7 normal testes (frozen tissue) TGCT cell lines | RT-PCR Droplet digital PCR IHC | Benesova et al. 2017 [45] |
| PLAYERS * | Most Frequently Deregulated (% of Cases) | Related Alterations (logOR) | SEvsNST | Best Performance (AUC) | Association with Stage | Survival Impact |
|---|---|---|---|---|---|---|
| DNA-modifying enzymes | ||||||
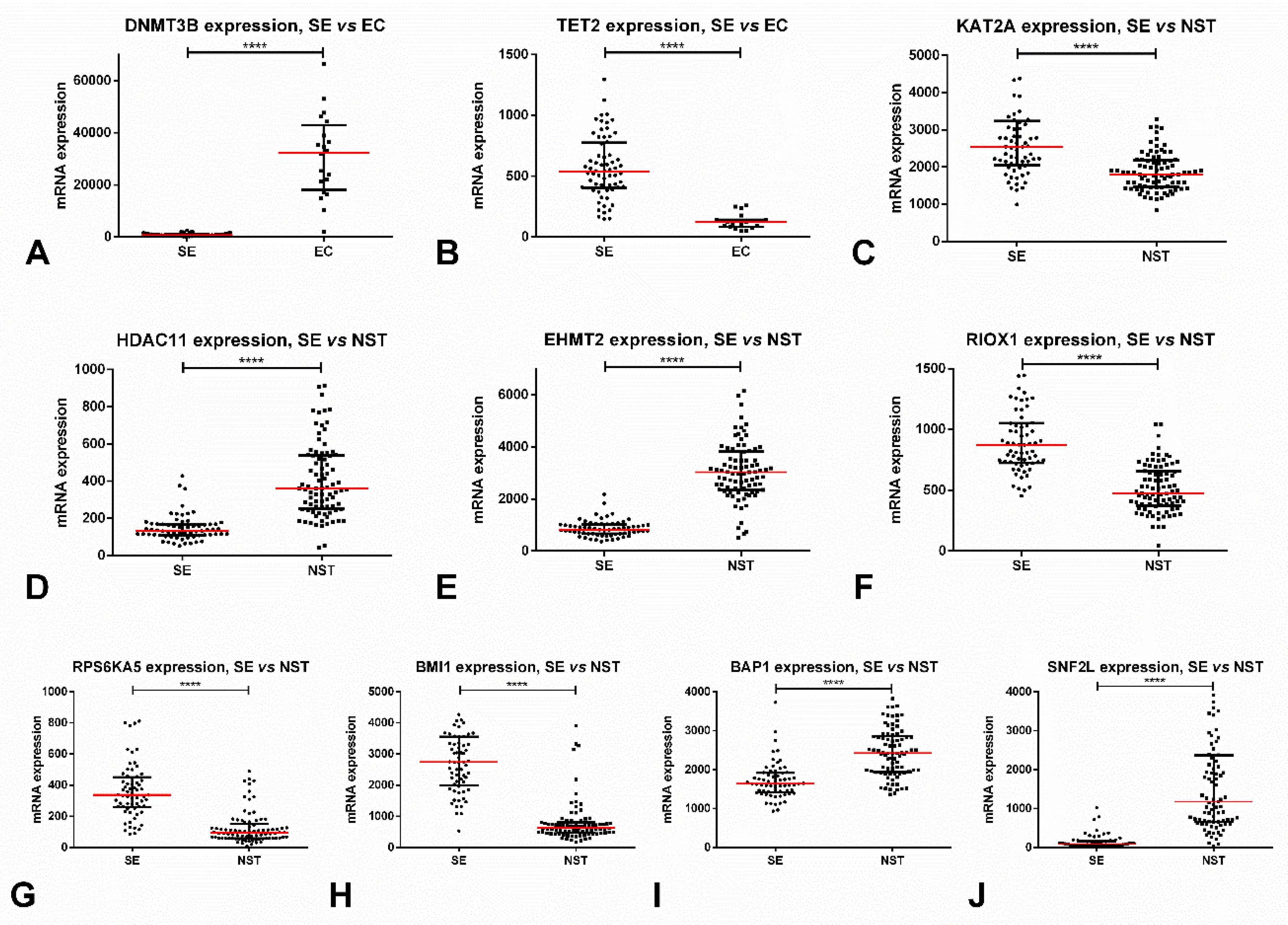
| DNMTs | DNMT3B (10) | DNMT3A and DNMT3B (co-occurrent, 2.785) | All ↓ in SE | DNMT3A (0.88) | Yes (DNMT3A/3B) | No |
| TETs | TET3 (8) | No | TET2 ↑ in SE | TET2 (0.79) | Yes (TET2) | No |
| Histone-modifying enzymes | ||||||
| ACETYLATION | ||||||
| KATs | ||||||
| MYST family | KAT6A (20) | KAT6A and KAT7 (mutually exclusive, <−3) | KAT6A/6B ↑ in SE; KAT5/8 ↓ in SE | KAT5 (0.75) | Yes (KAT6B and KAT8) | No |
| GNAT family | KAT9 (36) | KAT2A and KAT9 (mutually exclusive, <−3) | KAT1/2A/2B/9 ↑ in SE; SLC33A1 and ATAT1 ↓ in SE | KAT2A (0.78) | Yes (KAT2B) | No |
| Orphan family | KAT13C (41) | KAT3B and KAT13C; KAT13A and KAT13C (co-occurrent, >3) | KAT3B/13A/13C and CLOCK ↑ in SE | KAT13A (0.74) | Yes (KAT3B/13A/13C) | No |
| KDACs | ||||||
| HDACs | HDAC9 (14) | HDAC6 and HDAC8 (co-occurrent, 2.9) | HDAC1/2/3/8/9/11 ↓ in SE; HDAC5/6/7 ↑ in SE | HDAC11 (0.93) | Yes (HDAC1/11) | No |
| SIRTs | SIRT2/6 (10) | SIRT3 and SIRT6 (co-occurrent, >3) | SIRT4 ↓ in SE; SIRT5 ↑ in SE | SIRT4 (0.77) | Yes (SIRT4) | No |
| METHYLATION | ||||||
| KMTs | ||||||
| SET domain | SETD4 (45) EZH2 (21) | KMT2A and PRDM10; ASH1L and PRDM11; ASH1L and SMYD1 (co-occurrent, >3) | KMT2B/2C/2D, SETD1A/D4/DB2, EZH1, SMYD3 and PRDM1/2/7/15 ↑ in SE; EHMT2, MECOM, SETD7, PRDM5 and EZH2 ↓ in SE | EHMT2, PRDM1 and PRDM5 (AUC 0.96) | Yes (EHMT2, KMT2B and PRDM15) | Yes (KMT2D and PRDM2) |
| DOT1-like | DOT1L (9) | N/A | ↑ in SE | DOT1L (0.79) | No | No |
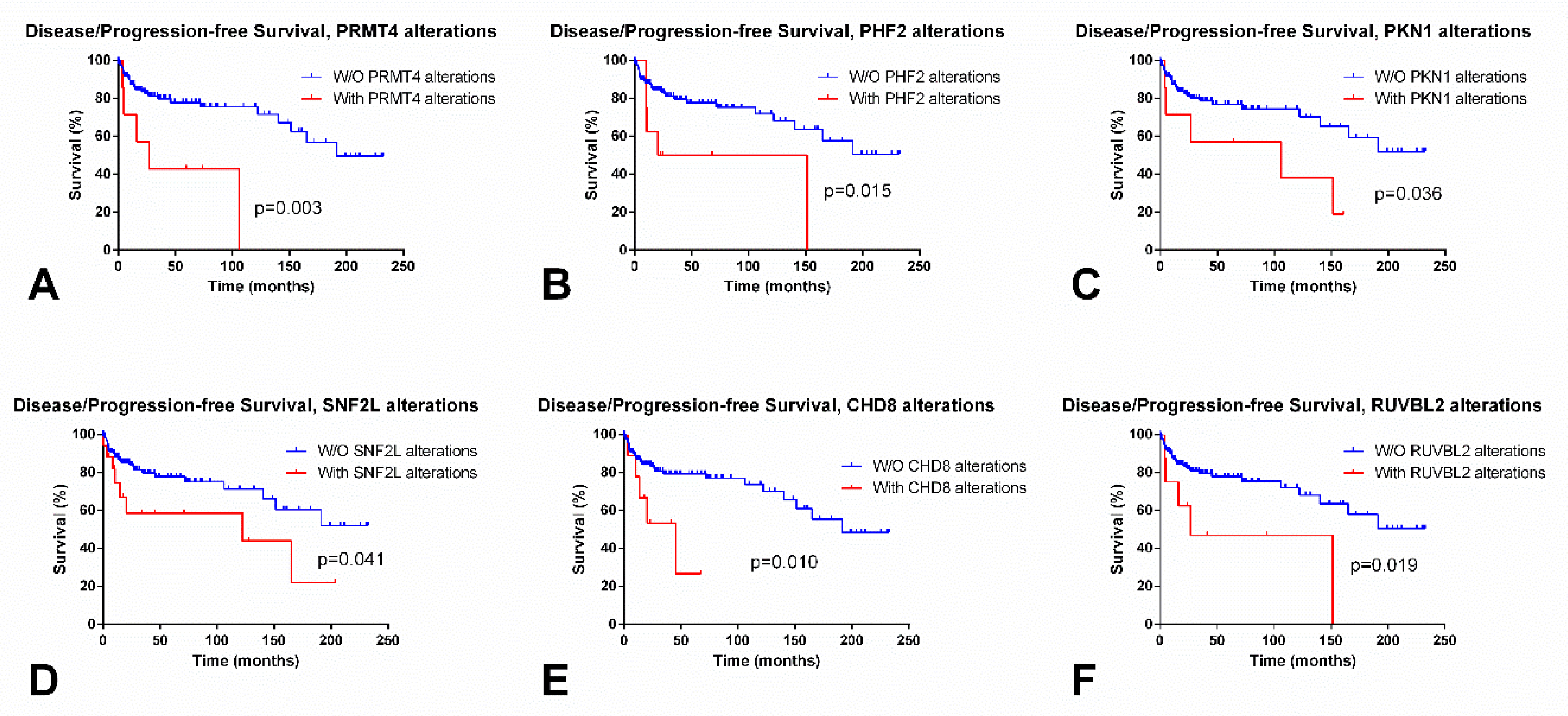
| PRMTs | PRMT8 (21) | No | PRMT8 ↓ in SE PRMT9 ↑ in SE | PRMT8 (0.83) | No | Yes (PRMT4) |
| KDMs | ||||||
| Jumonji-domain | KDM5A (21) KDM7A (19) | KDM4D and KDM4E (co-occurrent, >3) | KDM2A/3A/3B/4D/7A/8, RIOX1 and HSPBAP1 ↑ in SE; TYW5 and JMJD8 ↓ in SE | HSPBAP1 (0.93) | Yes (RIOX1 and KDM2A) | Yes (KDM6B and PHF2) |
| LSDs | LSD1 (8) | No | LSD2 ↑ in SE | LSD2 (0.73) | Yes (LSD2) | No |
| PHOSPHORYLATION | ||||||
| Kinases | BAZ1B (34) PRKDC (33) | CHEK2 and ATM (co-occurrent, 2.565) | BAZ1B, ATM, ATR and RPS6KA5 ↑ in SE; RPS6KA4, PAK2 and AURKB ↓ in SE | RPS6KA5 (0.89) | Yes (AURKB, ATM, ATR and RPS6KA4) | Yes (ATM, RPS6KA4 and PKN1) |
| UBIQUITINATION | ||||||
| Ubiquitin ligases | RING1 and RNF40 (10) | No | RNF2 and BMI1 ↑ in SE; RING1, RNF20 and EBE2A ↓ in SE | BMI1 (0.95) | Yes (BMI1, RNF2 and RNF20) | Yes (RNF40 and UBE2E1) |
| Deubiquitinating enzymes | USP16 and BAP1 (12) | No | USP16 ↑ in SE; BAP1 ↓ in SE | USP16 (0.89) | Yes (USP16) | No |
| Chromatin remodeling complexes | ||||||
| SWI/SNF | SMARCD1 (12) | ARID1B and PBRM1 (co-occurrent, >3) | BRG1 and BRM ↑ in SE; SMARCD1 ↓ in SE | BRM (0.84) | Yes (BRM) | No |
| ISWI | SNF2L (15) | SNF2L and BAZ1A (co-occurrent, 2.201) | SNF2L ↓ in SE | SNF2L (0.96) | Yes (SNF2L) | Yes (SNF2L) |
| CHD | CHD7 (28) CHD4 (21) | CHD3, CHD2 and CHD9 (co-occurrent alterations, >3) | CHD1/2/6/7 ↑ in SE; CHD4 ↓ in SE | CDH1 and CHD7 (0.81) | Yes (CHD7 and CHD8) | Yes (CHD2 and CHD8) |
| INO80 | ARP6 (12) | No | INO80 ↑ in SE | INO80 (0.88) | Yes (INO80) | Yes (SRCAP and RUVBL2) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobo, J.; Henrique, R.; Jerónimo, C. The Role of DNA/Histone Modifying Enzymes and Chromatin Remodeling Complexes in Testicular Germ Cell Tumors. Cancers 2019, 11, 6. https://doi.org/10.3390/cancers11010006
Lobo J, Henrique R, Jerónimo C. The Role of DNA/Histone Modifying Enzymes and Chromatin Remodeling Complexes in Testicular Germ Cell Tumors. Cancers. 2019; 11(1):6. https://doi.org/10.3390/cancers11010006
Chicago/Turabian StyleLobo, João, Rui Henrique, and Carmen Jerónimo. 2019. "The Role of DNA/Histone Modifying Enzymes and Chromatin Remodeling Complexes in Testicular Germ Cell Tumors" Cancers 11, no. 1: 6. https://doi.org/10.3390/cancers11010006
APA StyleLobo, J., Henrique, R., & Jerónimo, C. (2019). The Role of DNA/Histone Modifying Enzymes and Chromatin Remodeling Complexes in Testicular Germ Cell Tumors. Cancers, 11(1), 6. https://doi.org/10.3390/cancers11010006