Determination of the Membrane Transport Properties of Jurkat Cells with a Microfluidic Device

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Cells
2.2. Preparation of Cells
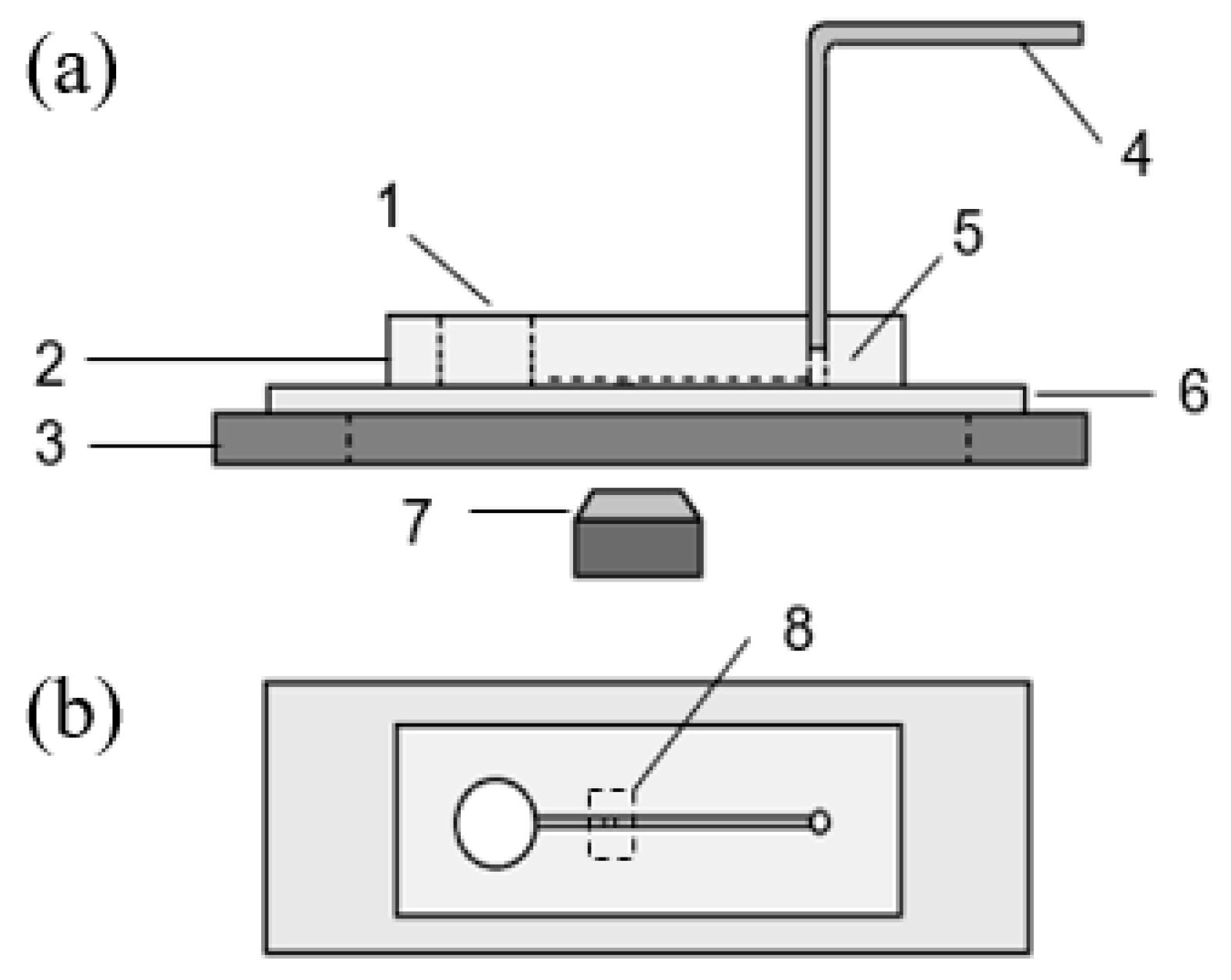
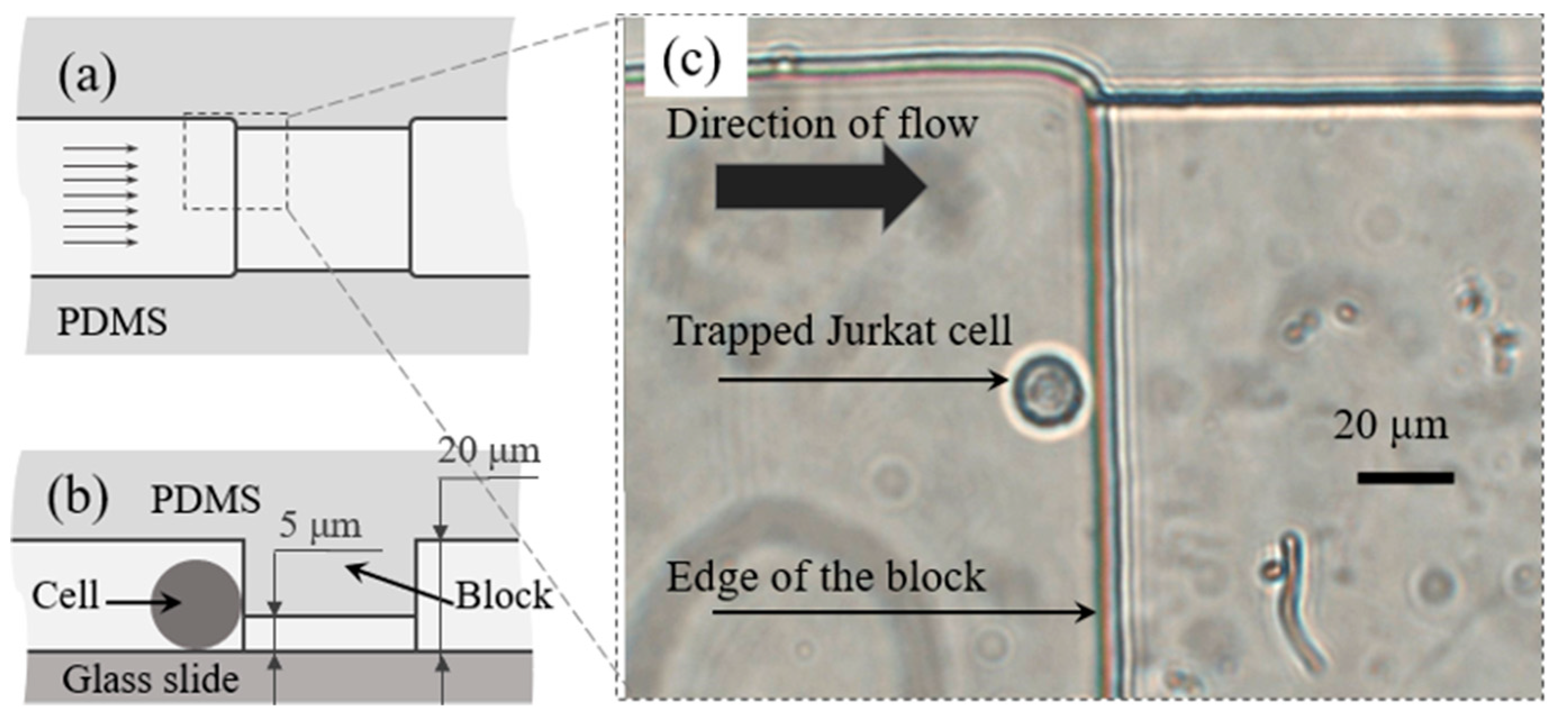
2.3. Design and Fabrication of the Microfluidic Chip
2.4. Setup of the Device and Operation Procedure
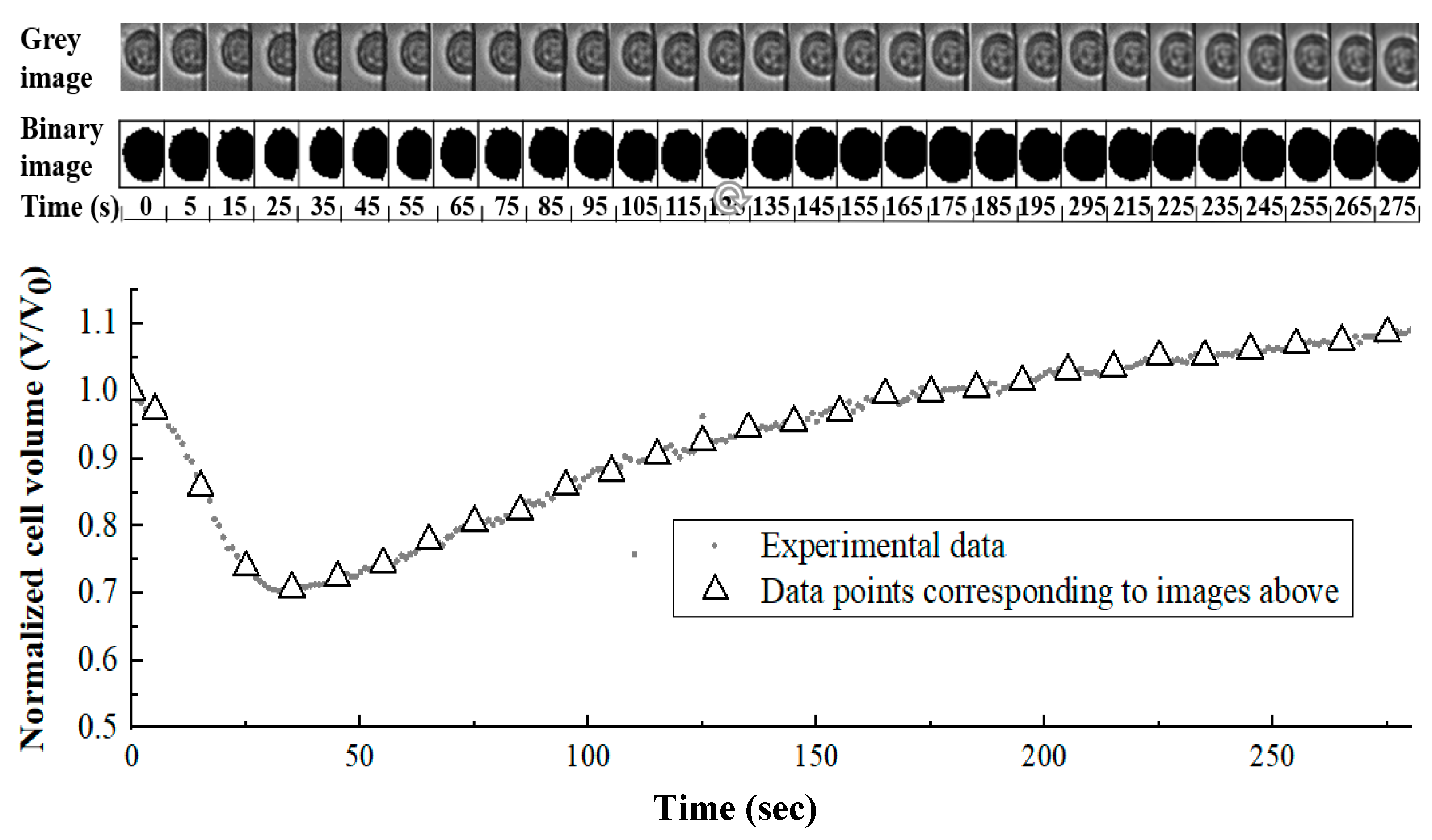
2.5. Image Analysis and Data Processing
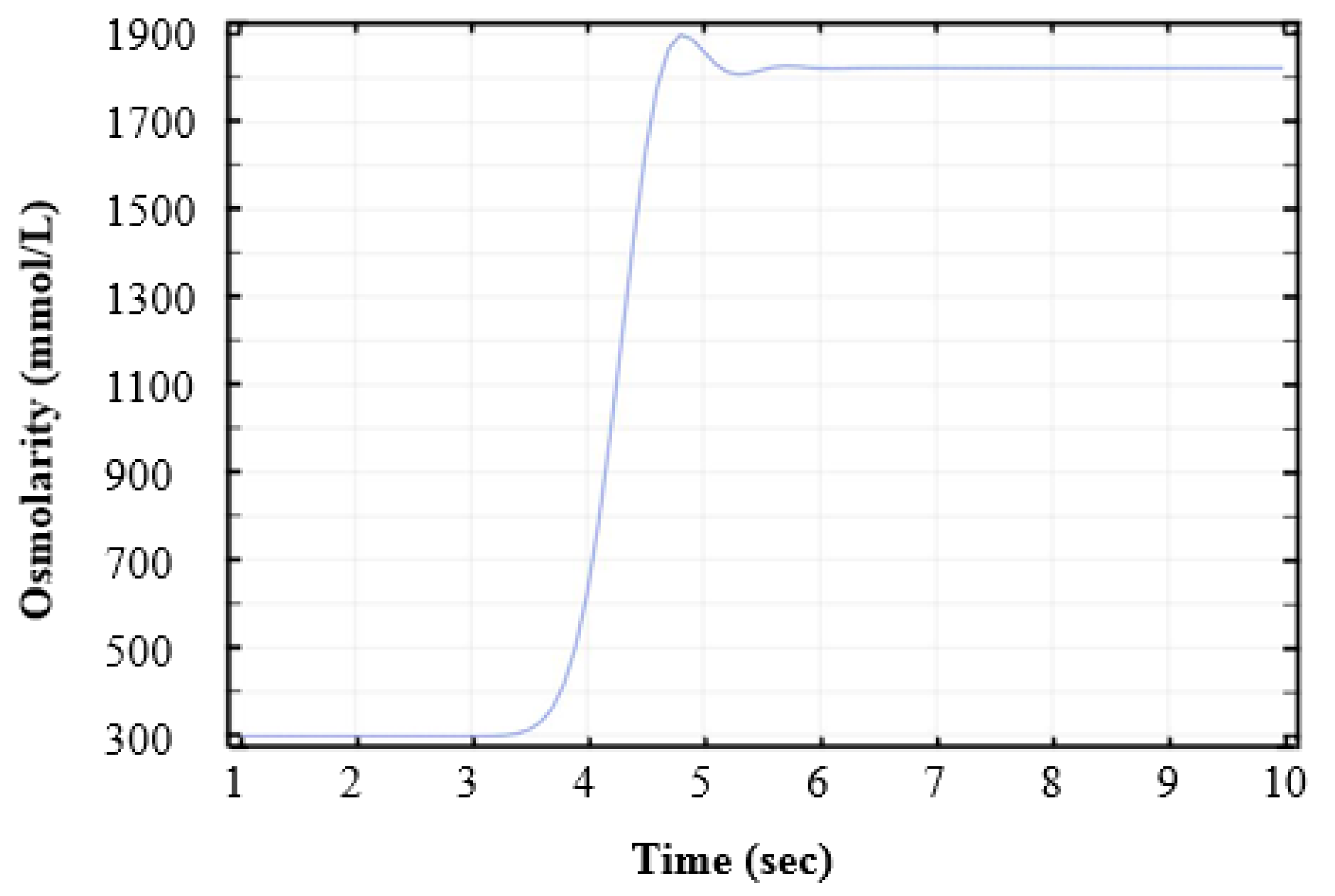
2.6. Validation of Replacement Time between Different Media
2.7. Mathematical Model of Cell Volume Change to Cryoprotective Agent (CPA) Addition
3. Results
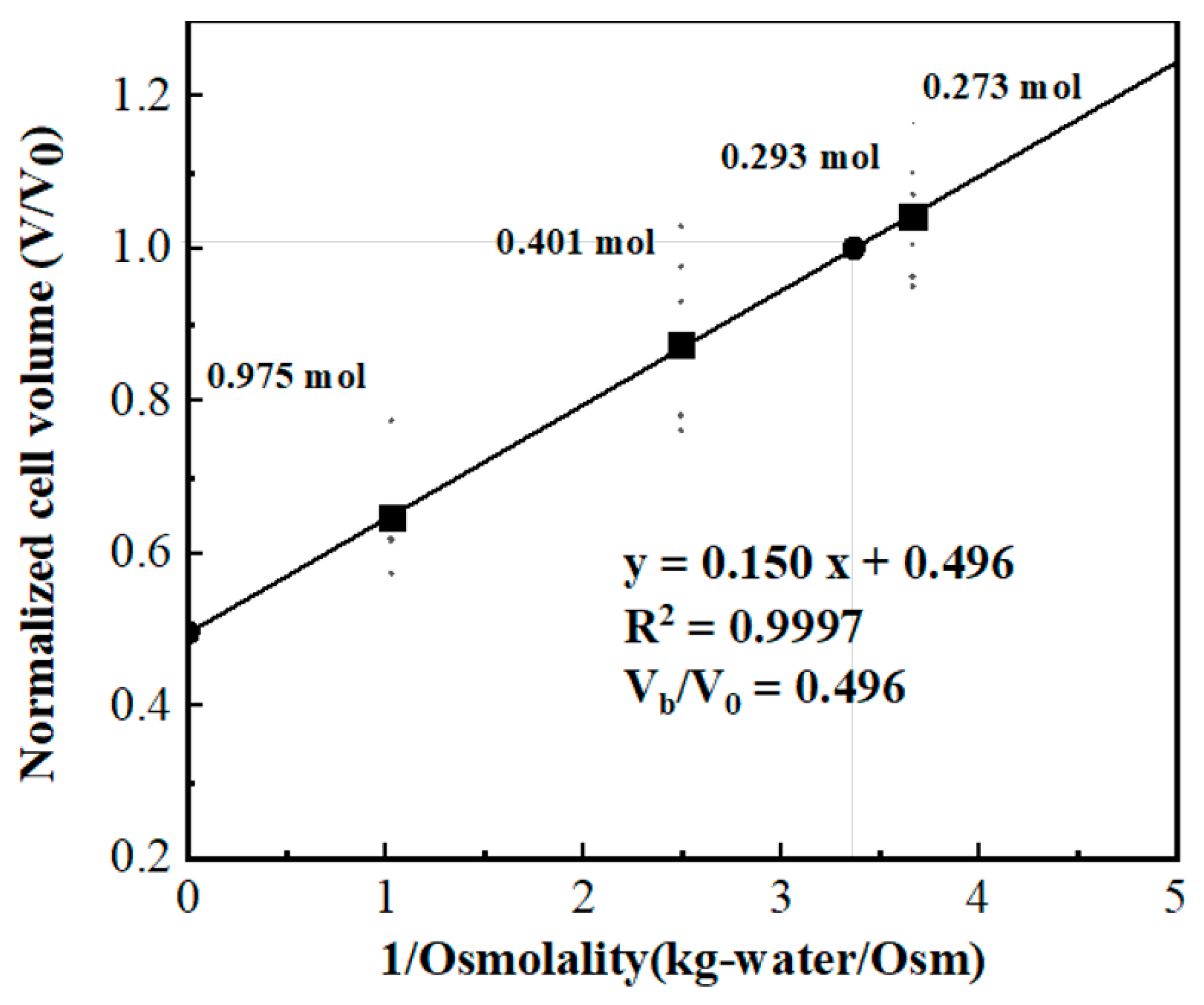
3.1. Obtaining Osmotically Inactive Cell Volume (Vb) of Jurkat Cell
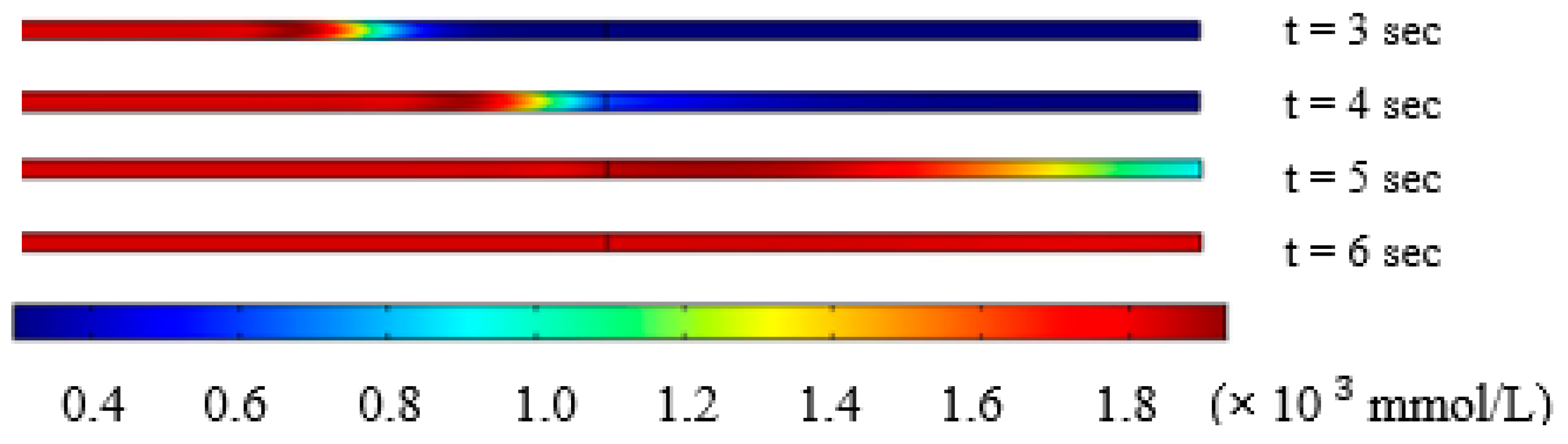
3.2. Solution Switching in the Microchannel
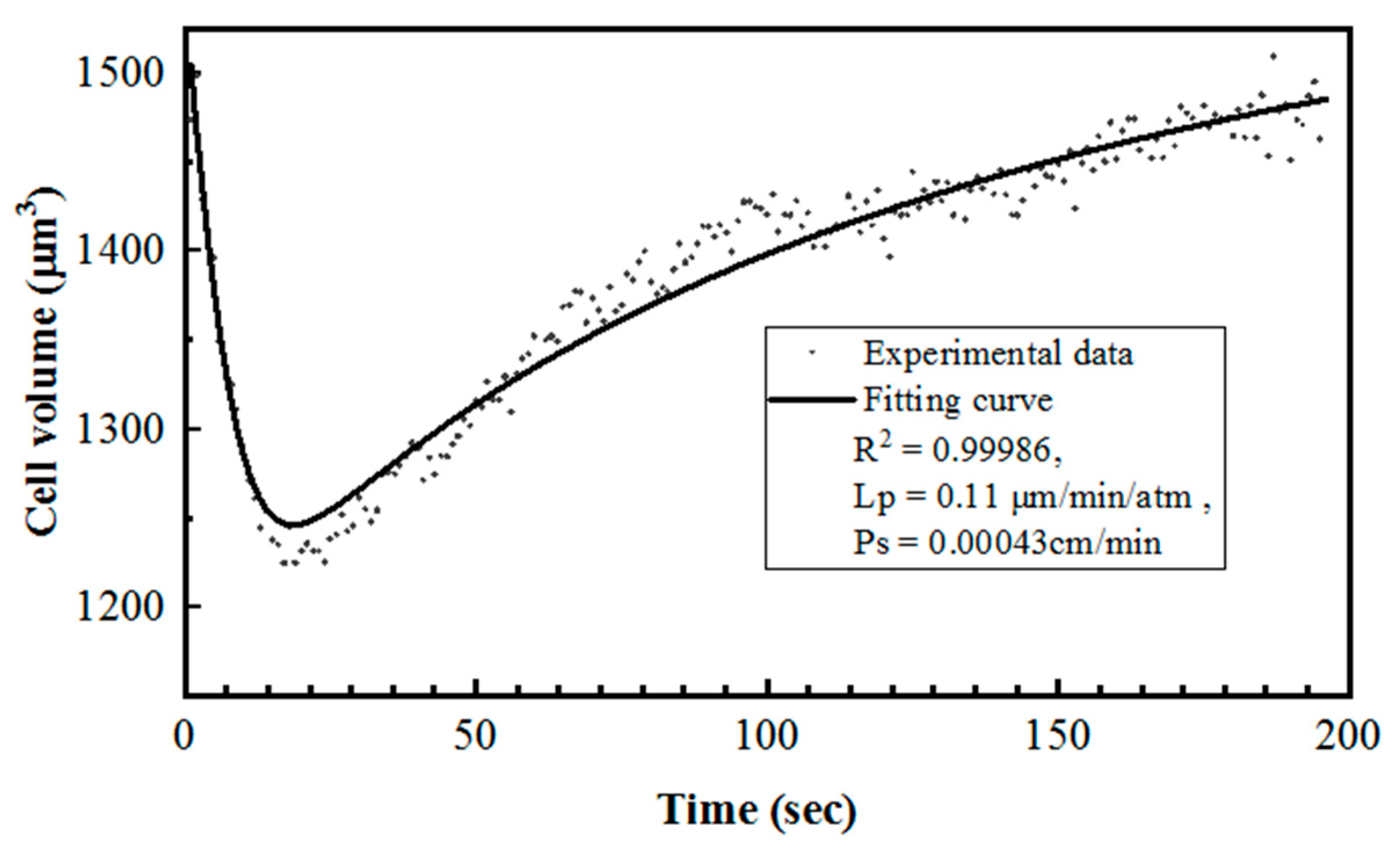
3.3. Cell Membrane Permeability Coefficient to Water (Lp) and CPAs (Ps) by Curve-Fitting
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mazur, P. Freezing of living cells: Mechanisms and implications. Am. J. Physiol. Cell Physiol. 1984, 247, C125–C142. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P. Cryobiology: The freezing of biological systems. Science 1970, 168, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Demirer, T.; Shu, Z.; Heimfeld, S.; Huang, Z.; Liu, C.; Gao, D. Progress in Stem Cell Transplantation; IntechOpen: London, UK, 2015; Volume 2, pp. 23–42. ISBN 978-953-51-2227-2. [Google Scholar]
- Mazur, P. Equilibrium, quasi-equilibrium, and nonequilibrium freezing of mammalian embryos. Cell Biophys. 1990, 17, 53–92. [Google Scholar] [CrossRef] [PubMed]
- Katkov, I.I.; Liebermann, J. Current Frontiers in Cryobiology; IntechOpen: London, UK, 2012; Volume 6, pp. 169–184. ISBN 978-953-51-0191-8. [Google Scholar]
- McGann, L.E.; Yang, H.; Walterson, M. Manifestations of cell damage after freezing and thawing. Cryobiology 1988, 25, 178–185. [Google Scholar] [CrossRef]
- Mazur, P. The role of intracellular freezing in the death of cells cooled at supraoptimal rates. Cryobiology 1977, 14, 251–272. [Google Scholar] [CrossRef]
- Mazur, P.; Koshimoto, C. Is intracellular ice formation the cause of death of mouse sperm frozen at high cooling rates? Biol. Reprod. 2002, 66, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P.; Leibo, S.; Chu, E. A two-factor hypothesis of freezing injury: Evidence from Chinese hamster tissue-culture cells. Exp. Cell Res. 1972, 71, 345–355. [Google Scholar] [CrossRef]
- Mazur, P.; Schmidt, J.J. Interactions of cooling velocity, temperature, and warming velocity on the survival of frozen and thawed yeast. Cryobiology 1968, 5, 1–17. [Google Scholar] [CrossRef]
- Łapińska, U.; Glover, G.; Capilla-Lasheras, P.; Young, A.J.; Pagliara, S. Bacterial ageing in the absence of external stressors. Philos. Trans. R. Soc. B 2019, 374, 20180442. [Google Scholar] [CrossRef]
- Pagliara, S.; Dettmer, S.L.; Keyser, U.F. Channel-Facilitated Diffusion Boosted by Particle Binding at the Channel Entrance. Phys. Rev. Lett. 2014, 113, 048102. [Google Scholar] [CrossRef]
- Nestorovich, E.M.; Danelon, C.; Winterhalter, M.; Bezrukov, S.M. Designed to penetrate: Time-resolved interaction of single antibiotic molecules with bacterial pores. Proc. Natl. Acad. Sci. USA 2002, 99, 9789–9794. [Google Scholar] [CrossRef] [PubMed]
- Gravelle, S.; Joly, L.; Detcheverry, F.; Ybert, C.; Bocquet, L. Optimizing water permeability through the hourglass shape of aquaporins. Proc. Natl. Acad. Sci. USA 2013, 110, 16367–16372. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J. Quantitative measurement of cell membrane transport: Technology and applications. Cryobiology 1997, 34, 315–334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Fu, J. Microfluidics for cryopreservation. Biotechnol. Adv. 2017, 35, 323–336. [Google Scholar] [CrossRef]
- Casquillas, G.V.; Fu, C.; Le Berre, M.; Cramer, J.; Meance, S.; Plecis, A.; Baigl, D.; Greffet, J.-J.; Chen, Y.; Piel, M. Fast microfluidic temperature control for high resolution live cell imaging. Lab Chip 2011, 11, 484–489. [Google Scholar] [CrossRef]
- Gao, D.; McGrath, J.; Tao, J.; Benson, C.; Critser, E.; Critser, J. Membrane transport properties of mammalian oocytes: A micropipette perfusion technique. Reproduction 1994, 102, 385–392. [Google Scholar] [CrossRef]
- McGrath, J. A microscope diffusion chamber for the determination of the equilibrium and non-equilibrium osmotic response of individual cells. J. Microsc. 1985, 139, 249–263. [Google Scholar] [CrossRef]
- Tseng, H.-Y.; Sun, S.; Shu, Z.; Ding, W.; Reems, J.-A.; Gao, D. A microfluidic study of megakaryocytes membrane transport properties to water and dimethyl sulfoxide at suprazero and subzero temperatures. Biopreserv. Biobank. 2011, 9, 355–362. [Google Scholar] [CrossRef]
- Guo, F.; Mao, Z.; Chen, Y.; Xie, Z.; Lata, J.P.; Li, P.; Ren, L.; Liu, J.; Yang, J.; Dao, M. Three-dimensional manipulation of single cells using surface acoustic waves. Proc. Natl. Acad. Sci. USA 2016, 113, 1522–1527. [Google Scholar] [CrossRef]
- Gossett, D.R.; Tse, H.T.K.; Lee, S.A.; Ying, Y.; Lindgren, A.; Yang, O.O.; Rao, J.; Clark, A.T.; Carlo, D.D. Hydrodynamic stretching of single cells for large population mechanical phenotyping. Proc. Natl. Acad. Sci. USA 2012, 109, 7630–7635. [Google Scholar] [CrossRef]
- Fang, C.; Ji, F.; Shu, Z.; Gao, D. Determination of the temperature-dependent cell membrane permeabilities using microfluidics with integrated flow and temperature control. Lab Chip 2017, 17, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, S.; Franze, K.; McClain, C.R.; Wylde, G.; Fisher, C.L.; Franklin, R.J.M.; Kabla, A.J.; Keyser, U.F.; Chalut, K.J. Transition from pluripotency in embryonic stem cells distinguished by an auxetic nucleus. Nat. Mater. 2014, 13, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Bamford, R.A.; Smith, A.; Metz, J.; Glover, G.; Titball, R.W.; Pagliara, S. Investigating the physiology of viable but non-culturable bacteria by microfluidics and time-lapse microscopy. BMC Biol. 2017, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S.; Van Hoek, A.; Ma, T.; Frigeri, A.; Skach, W.R.; Mitra, A.K.; Tamarappoo, B.K.; Farinas, J. Water transport across mammalian cell membranes. Am. J. Physiol. Cell Physiol. 1996, 270, C12–C30. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.M.; Samper, V.; Chen, Y.; Heng, C.K.; Lim, T.M.; Yobas, L. Silicon-based microfilters for whole blood cell separation. Biomed. Microdevices 2008, 10, 251–257. [Google Scholar] [CrossRef]
- Kedem, O. Commentary on ‘Thermodynamic analysis of the permeability of biological membranes to non-electrolytes’ by O. Kedem and A. Katchalsky Biochim. Biophys. Acta 27 (1958) 229–246. Biochim. Biophys. Acta 1989, 1000, 411–430. [Google Scholar] [CrossRef]
- Gossett, D.R.; Weaver, W.M.; Mach, A.J.; Hur, S.C.; Tse, H.; Lee, W.; Amini, H.; Carlo, D.D. Label-free cell separation and sorting in microfluidic systems. Anal. Bioanal. Chem. 2010, 397, 3249–3267. [Google Scholar] [CrossRef]
- Chen, H.; Purtteman, J.J.; Heimfeld, S.; Folch, A.; Gao, D. Development of a microfluidic device for determination of cell osmotic behavior and membrane transport properties. Cryobiology 2007, 55, 200–209. [Google Scholar] [CrossRef]
- Di Carlo, D.; Wu, L.Y.; Lee, L.P. Dynamic single cell culture array. Lab Chip 2006, 6, 1445–1449. [Google Scholar] [CrossRef]
- Heo, Y.S.; Lee, H.J.; Hassell, B.A.; Irimia, D.; Toth, T.L.; Elmoazzen, H.; Toner, M. Controlled loading of cryoprotectants (CPAs) to oocyte with linear and complex CPA profiles on a microfluidic platform. Lab Chip 2011, 11, 3530–3537. [Google Scholar] [CrossRef]
- Schneider, U.; Schwenk, H.U.; Bornkamm, G. Characterization of EBV-genome negative “null” and “T” cell lines derived from children with acute lymphoblastic leukemia and leukemic transformed non-Hodgkin lymphoma. Int. J. Cancer 2010, 19, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Wu, J.; Liao, H.; Chen, Y.; Kuo, C. N-Farnesyloxy-norcantharimide inhibits progression of human leukemic Jurkat T cells through regulation of mitogen-activated protein kinase and interleukin-2 production. Anticancer Drugs 2015, 26, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, Y.; Zhang, Y.; Zhang, C. T Cell Receptor Signaling Pathways: New Targets for Herpes Simplex Virus. Virol. Sin. 2008, 23, 429–437. [Google Scholar] [CrossRef]
- Abraham, R.T.; Arthur, W. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nat. Rev. Immunol. 2004, 4, 301–308. [Google Scholar] [CrossRef]
- Wang, X.; Viswanath, R.; Zhao, J.; Tang, S.; Hewlett, I. Changes in the level of apoptosis-related proteins in Jurkat cells infected with HIV-1 versus HIV-2. Mol. Cell. Biochem. 2010, 337, 175–183. [Google Scholar] [CrossRef]
- Moore, B.E. Survival of human immunodeficiency virus (HIV), HIV-infected lymphocytes, and poliovirus in water. Appl. Environ. Microbiol. 1993, 59, 1437–1443. [Google Scholar]
- Ryu, B.Y.; Evans-Galea, M.V.; Gray, J.T.; Bodine, D.M.; Persons, D.A.; Nienhuis, A.W. An experimental system for the evaluation of retroviral vector design to diminish the risk for proto-oncogene activation. Blood 2008, 111, 1866–1875. [Google Scholar] [CrossRef]
- Takahashi, K.; Murakami, M.; Hosaka, K.; Kikuchi, H.; Oshima, Y.; Kubohara, Y. Regulation of IL-2 production in Jurkat cells by Dictyostelium-derived factors. Life Sci. 2009, 85, 438–443. [Google Scholar] [CrossRef]
- Shu, Z.; Heimfeld, S.; Gao, D. Hematopoietic SCT with cryopreserved grafts: Adverse reactions after transplantation and cryoprotectant removal before infusion. Bone Marrow Transp. 2013, 49, 469–476. [Google Scholar] [CrossRef]
- Phillips, K.G.; Jacques, S.L.; Mccarty, O.J.T. Measurement of single cell refractive index, dry mass, volume, and density using a transillumination microscope. Phys. Rev. Lett. 2012, 109, 118105. [Google Scholar] [CrossRef]
- Curl, C.L.; Bellair, C.J.; Harris, P.J.; Allman, B.E.; Roberts, A.; Nugent, K.A.; Delbridge, L.M. Single cell volume measurement by quantitative phase microscopy (QPM): A case study of erythrocyte morphology. J. Cell Physiol. 2006, 17, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Hai, F.; Liu, X.; Li, S. Mixing indexes considering the combination of mean and dispersion information from intensity images for the performance estimation of micromixing. RSC Adv. 2017, 7, 10906–10914. [Google Scholar]
- Gao, D.; Benson, C.; Liu, C.; McGrath, J.; Critser, E.; Critser, J. Development of a novel microperfusion chamber for determination of cell membrane transport properties. Biophys. J. 1996, 71, 443–450. [Google Scholar] [CrossRef]
- Holz, M.; Heil, S.R.; Sacco, A. Temperature-dependent self-diffusion coefficients of water and six selected molecular liquids for calibration in accurate 1H NMR PFG measurements. Phys. Chem. Chem. Phys. 2000, 2, 4740–4742. [Google Scholar] [CrossRef]
- Kleinhans, F. Membrane permeability modeling: Kedem–Katchalsky vs a two-parameter formalism. Cryobiology 1998, 37, 271–289. [Google Scholar] [CrossRef]
- Vian, A.M.; Higgins, A.Z. Membrane permeability of the human granulocyte to water, dimethyl sulfoxide, glycerol, propylene glycol and ethylene glycol. Cryobiology 2014, 68, 35–42. [Google Scholar] [CrossRef]
- Shu, Z.; Hughes, S.M.; Fang, C.; Huang, J.; Fu, B.; Zhao, G.; Fialkow, M.; Lentz, G.; Hladik, F.; Gao, D. A study of the osmotic characteristics, water permeability, and cryoprotectant permeability of human vaginal immune cells. Cryobiology 2016, 72, 93–99. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Peng, J.; Shu, Z.; Sekar, P.K.; Li, S.; Gao, D. Determination of the Membrane Transport Properties of Jurkat Cells with a Microfluidic Device. Micromachines 2019, 10, 832. https://doi.org/10.3390/mi10120832
Yang T, Peng J, Shu Z, Sekar PK, Li S, Gao D. Determination of the Membrane Transport Properties of Jurkat Cells with a Microfluidic Device. Micromachines. 2019; 10(12):832. https://doi.org/10.3390/mi10120832
Chicago/Turabian StyleYang, Tianhang, Ji Peng, Zhiquan Shu, Praveen K. Sekar, Songjing Li, and Dayong Gao. 2019. "Determination of the Membrane Transport Properties of Jurkat Cells with a Microfluidic Device" Micromachines 10, no. 12: 832. https://doi.org/10.3390/mi10120832
APA StyleYang, T., Peng, J., Shu, Z., Sekar, P. K., Li, S., & Gao, D. (2019). Determination of the Membrane Transport Properties of Jurkat Cells with a Microfluidic Device. Micromachines, 10(12), 832. https://doi.org/10.3390/mi10120832