Zinc in Infection and Inflammation


Abstract
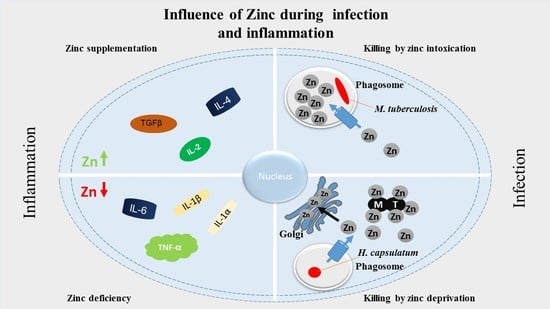
:
1. Introduction
2. Zinc and Nutrition
3. Zinc Homeostasis
3.1. Zinc Transporters
3.2. Metallothioneins and Other Zinc Binding Proteins
4. Zinc and Immunity
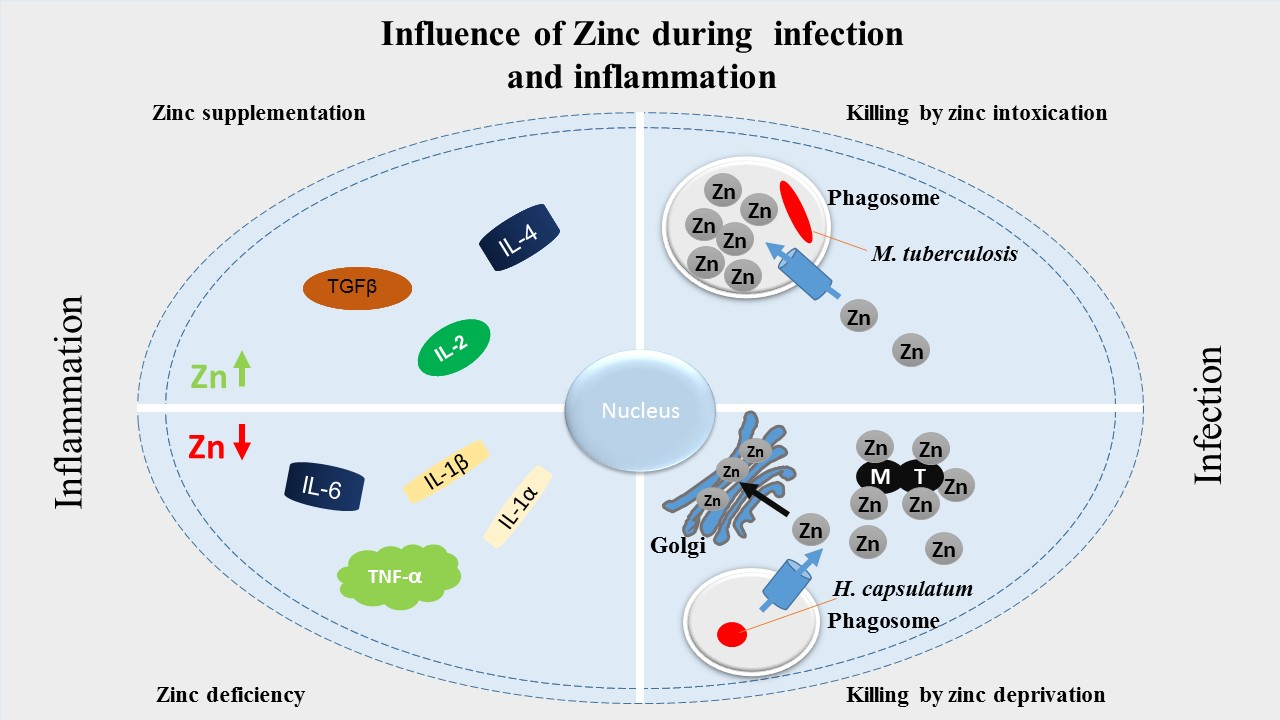
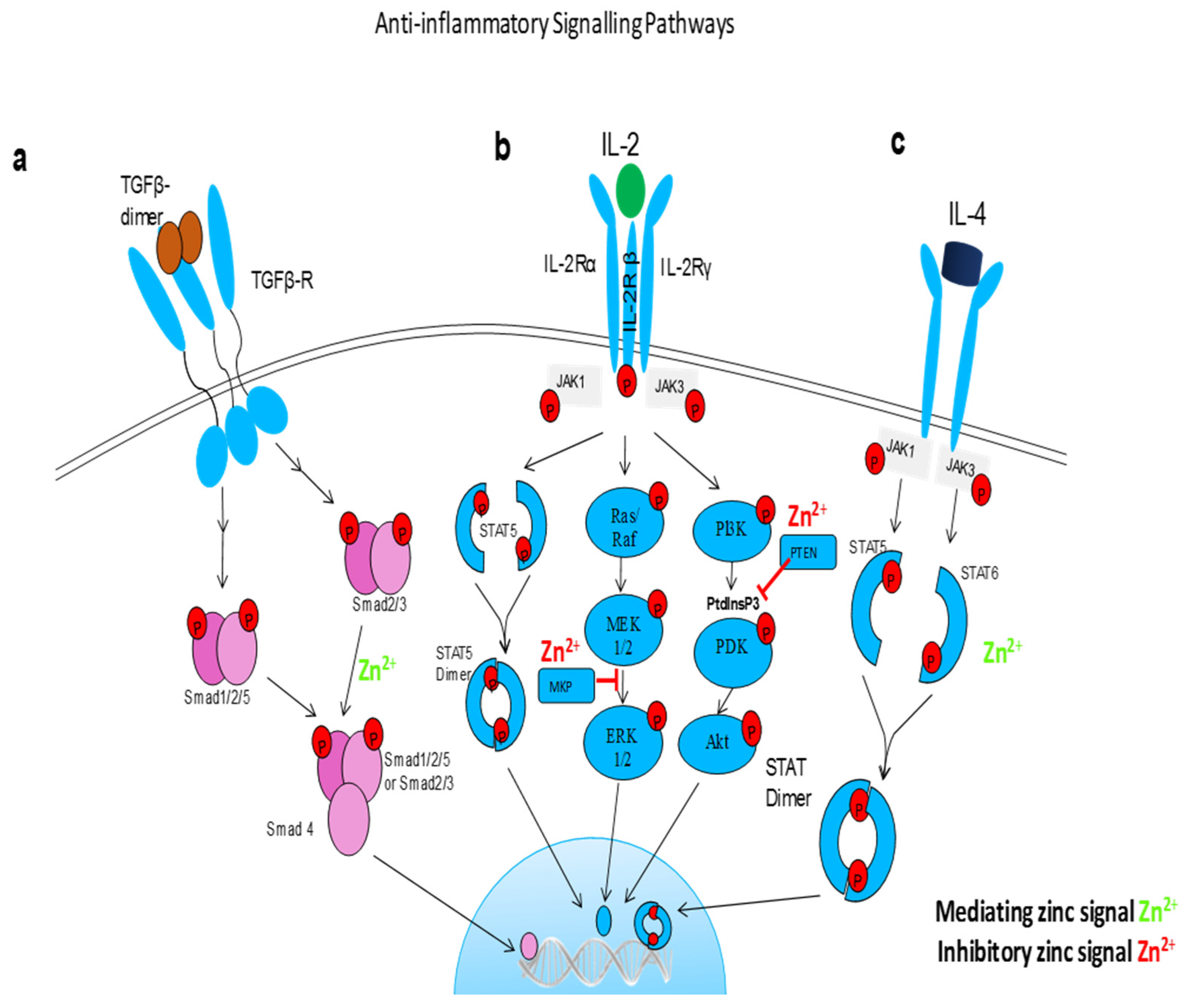
5. Zinc in Inflammation
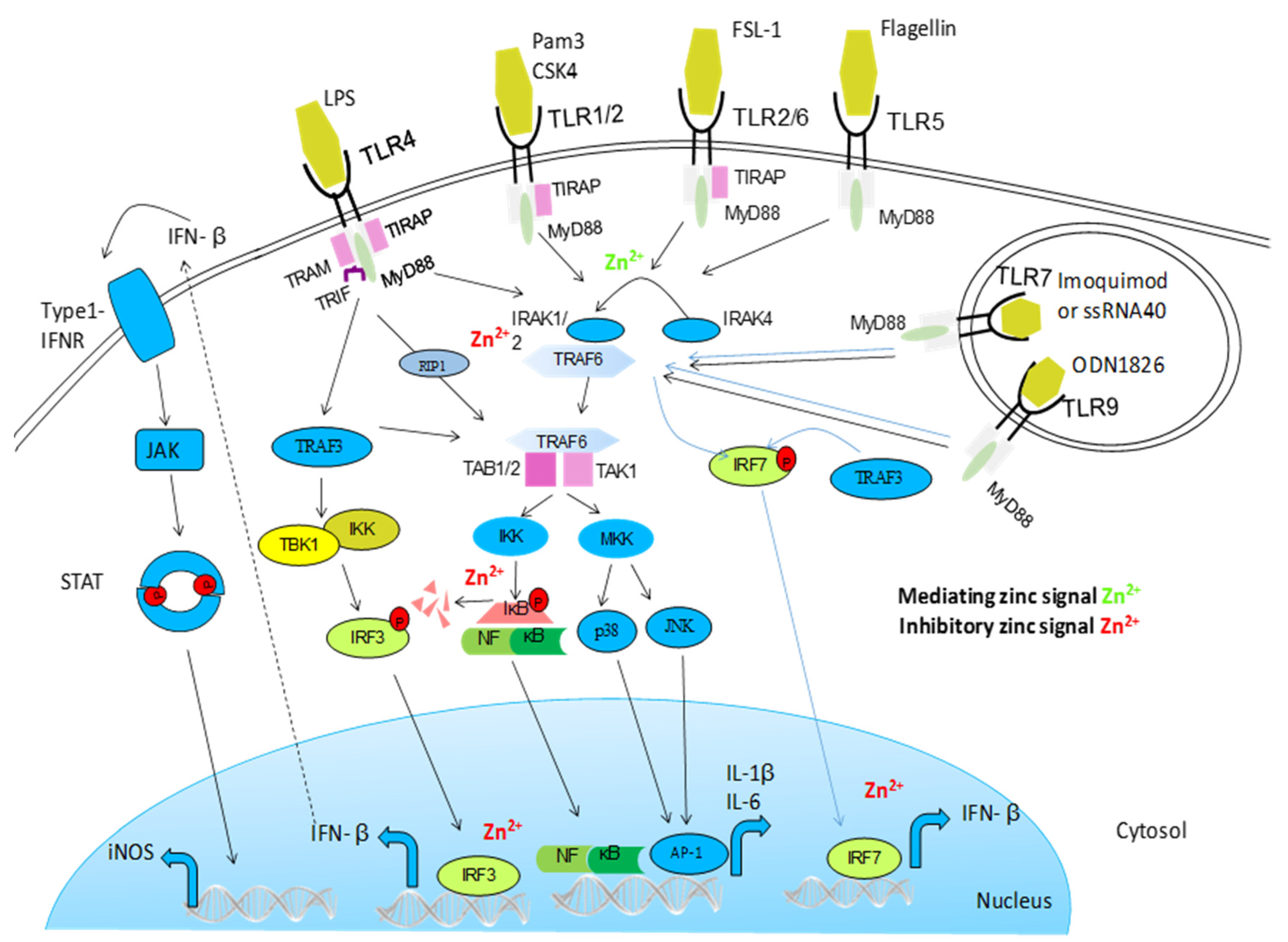
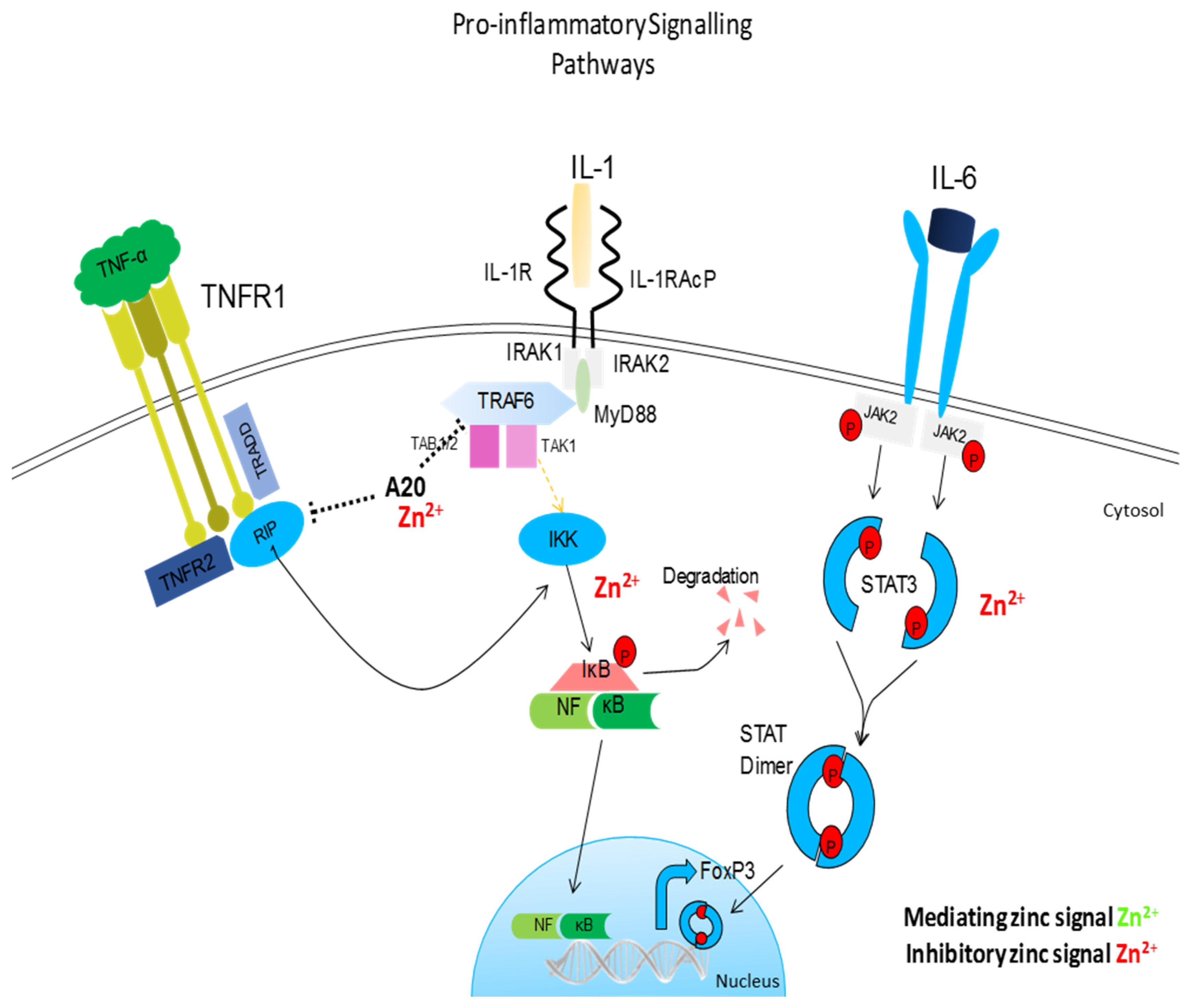
5.1. NF-κB and Other Signalling Pathways
5.2. Oxidative Stress
5.3. Zinc Status and Inflammatory Cytokines
6. Zinc in Infection
6.1. Nutritional Immunity
6.2. Zinc as a Critical Component of the Membrane Barrier
6.3. Peptidoglycan Regulation Proteins (PGLYRPs)
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prasad, A.S.; Miale, A., Jr.; Farid, Z.; Sandstead, H.H.; Schulert, A.R. Zinc metabolism in patients with the syndrome of iron deficiency anemia, hepatosplenomegaly, dwarfism, and hypogonadism. J. Lab. Clin. Med. 1963, 61, 537–549. [Google Scholar] [PubMed]
- Rink, L.; Gabriel, P. Zinc and the immune system. Proc. Nutr. Soc. 2000, 59, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [PubMed]
- Maret, W.; Li, Y. Coordination dynamics of zinc in proteins. Chem. Rev. 2009, 109, 4682–4707. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Rink, L. Multiple impacts of zinc on immune function. Metall. Integr. Biomet. Sci. 2014, 6, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc and Human Disease. In Interrelations between Essential Metal Ions and Human Diseases; Sigel, A., Sigel, H., Sigel, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 389–414. [Google Scholar]
- Mills, C.F. (Ed.) Zinc in Human Biology. In Physiology of Zinc: General Aspects; Springer: London, UK, 1989. [Google Scholar] [CrossRef]
- Favier, A.; Favier, M. Effects of zinc deficiency in pregnancy on the mother and the newborn infant. Rev. Fr. Gynecol. Obstet. 1990, 85, 13–27. [Google Scholar] [PubMed]
- Otten, J.J.; Pitzi Hellwig, J.; Meyers, L.D. (Eds.) Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; The National Academies Press: Washington, DC, USA, 2006; Available online: https://doi.org/10.17226/11537 (accessed on 7 July 2017).
- Deutsche Gesellschaft für Ernährung. Österreichische Gesellschaft für Ernährung; Schweizerische Gesellschaft für Ernährungsforschung; Schweizerische Vereinigung für Ernährung; Referenzwerte für die Nährstoffzufuhr: Bonn, Germany, 2016. (In German) [Google Scholar]
- World Health Organization (WHO). Trace Elements in Human Nutrition and Health; World Health Organization: Geneva, Switzerland, 1996. [Google Scholar]
- EFSA Panel on Dietetic Products Nutrition and Allergies. Scientific Opinion on Dietary Reference Values for zinc. EFSA J. 2014, 12. [Google Scholar] [CrossRef]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)—Zinc Review. J. Nutr. 2016, 146, 858S–885S. [Google Scholar] [CrossRef] [PubMed]
- Brieger, A.; Rink, L. Zink und Immunfunktionen. Ernährung Medizin 2010, 25, 156–160. [Google Scholar] [CrossRef]
- Sandstead, H.H. Zinc Deficiency. Am. J. Dis. Child. 1991, 145, 853–859. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. The World Health Report; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Wellenreuther, G.; Cianci, M.; Tucoulou, R.; Meyer-Klaucke, W.; Haase, H. The ligand environment of zinc stored in vesicles. Biochem. Biophys. Res. Commun. 2009, 380, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Hashimoto, A.; Fujimoto, S. Current understanding of ZIP and ZnT zinc transporters in human health and diseases. Cell. Mol. Life Sci. 2014, 71, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Rink, L. Functional significance of zinc-related signaling pathways in immune cells. Annu. Rev. Nutr. 2009, 29, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Kambe, T. The Functions of Metallothionein and ZIP and ZnT Transporters: An Overview and Perspective. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Discovery of human zinc deficiency: Its impact on human health and disease. Adv. Nutr. 2013, 4, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Eide, D.J. The SLC39 family of zinc transporters. Mol. Asp. Med. 2013, 34, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Tepaamorndech, S. The SLC30 family of zinc transporters—A review of current understanding of their biological and pathophysiological roles. Mol. Asp. Med. 2013, 34, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Murgia, C.; Leong, M.; Tan, L.-W.; Perozzi, G.; Knight, D.; Ruffin, R.; Zalewski, P. Anti-inflammatory effects of zinc and alterations in zinc transporter mRNA in mouse models of allergic inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L577–L584. [Google Scholar] [CrossRef] [PubMed]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Caruso, J.A.; Deepe, G.S., Jr. Granulocyte macrophage-colony stimulating factor induced Zn sequestration enhances macrophage superoxide and limits intracellular pathogen survival. Immunity 2013, 39, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. The function of zinc metallothionein: A link between cellular zinc and redox state. J. Nutr. 2000, 130, 1455S–1458S. [Google Scholar] [PubMed]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Stewart, A.J.; Sadler, P.J.; Pinheiro, T.J.; Blindauer, C.A. Albumin as a zinc carrier: Properties of its high-affinity zinc-binding site. Biochem. Soc. Trans. 2008, 36, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.; Cox, J.A. New perspectives on S100 proteins: A multi-functional Ca(2+)-, Zn(2+)- and Cu(2+)-binding protein family. Biometals 1998, 11, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Gilston, B.A.; Skaar, E.P.; Chazin, W.J. Binding of transition metals to S100 proteins. Sci. China Life Sci. 2016, 59, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Costarelli, L.; Giacconi, R.; Cipriano, C.; Muti, E.; Malavolta, M. Zinc-binding proteins (metallothionein and alpha-2 macroglobulin) and immunosenescence. Exp. Gerontol. 2006, 41, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, J.R.; Oteiza, P.I. Zinc and the aging brain. Genes Nutr. 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- DeCoursey, T.E.; Morgan, D.; Cherny, V.V. The voltage dependence of NADPH oxidase reveals why phagocytes need proton channels. Nature 2003, 422, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Suzuki, K.; Nakaji, S.; Sugawara, K. Effects of zinc on the reactive oxygen species generating capacity of human neutrophils and on the serum opsonic activity in vitro. Luminescence 2000, 15, 321–327. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Rink, L. Zinc signals and immune function. BioFactors 2014, 40, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Gaetke, L.M.; Frederich, R.C.; Oz, H.S.; McClain, C.J. Decreased food intake rather than zinc deficiency is associated with changes in plasma leptin, metabolic rate, and activity levels in zinc deficient rats. J. Nutr. Biochem. 2002, 13, 237–244. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology 2017, 25, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.; Samman, S. Zinc and Regulation of Inflammatory Cytokines: Implications for Cardiometabolic Disease. Nutrients 2012, 4, 676–694. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Ober-Blöbaum, J.L.; Engelhardt, G.; Hebel, S.; Heit, A.; Heine, H.; Rink, L. Zinc Signals Are Essential for Lipopolysaccharide-Induced Signal Transduction in Monocytes. J. Immunol. 2008, 181, 6491–6502. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Bao, B.; Beck, F.W.; Sarkar, F.H. Zinc-suppressed inflammatory cytokines by induction of A20-mediated inhibition of nuclear factor-κB. Nutrition 2011, 27, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Bao, B.; Beck, F.W.; Kucuk, O.; Sarkar, F.H. Antioxidant effect of zinc in humans. Free Radic. Biol. Med. 2004, 37, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Brieger, A.; Rink, L.; Haase, H. Differential Regulation of TLR-Dependent MyD88 and TRIF Signaling Pathways by Free Zinc Ions. J. Immunol. 2013, 191, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.I.; Ledford, J.R.; Zhou, P.; Page, K. Zinc supplementation alters airway inflammation and airway hyperresponsiveness to a common allergen. J. Inflamm. 2011, 8, 36. [Google Scholar] [CrossRef]
- Yan, Y.-W.; Fan, J.; Bai, S.-L.; Hou, W.-J.; Li, X.; Tong, H. Zinc Prevents Abdominal Aortic Aneurysm Formation by Induction of A20-Mediated Suppression of NF-κB Pathway. PLoS ONE 2016, 11, e0148536. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Guo, S.; Gao, J.; Guo, Y.; Du, E.; Lv, Z.; Zhang, B. Maternal high-zinc diet attenuates intestinal inflammation by reducing DNA methylation and elevating H3K9 acetylation in the A20 promoter of offspring chicks. J. Nutr. Biochem. 2015, 26, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Prasad, A.S.; Beck, F.W.J.; Fitzgerald, J.T.; Snell, D.; Bao, G.W.; Singh, T.; Cardozo, L.J. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: A potential implication of zinc as an atheroprotective agent. Am. J. Clin. Nutr. 2010, 91, 1634–1641. [Google Scholar] [CrossRef] [PubMed]
- Von Bulow, V.; Dubben, S.; Engelhardt, G.; Hebel, S.; Plumakers, B.; Heine, H.; Rink, L.; Haase, H. Zinc-dependent suppression of TNF-alpha production is mediated by protein kinase A-induced inhibition of Raf-1, I kappa B kinase beta, and NF-kappa B. J. Immunol. 2007, 179, 4180–4186. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Hasegawa, A.; Nakae, S.; Oboki, K.; Saito, H.; Yamasaki, S.; Hirano, T. Zinc transporter Znt5/Slc30a5 is required for the mast cell-mediated delayed-type allergic reaction but not the immediate-type reaction. J. Exp. Med. 2009, 206, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Truong-Tran, A.Q.; Carter, J.; Ruffin, R.E.; Zalewski, P.D. The role of zinc in caspase activation and apoptotic cell death. Biometals 2001, 14, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed]
- Stennicke, H.R.; Salvesen, G.S. Biochemical Characteristics of Caspases-3, -6, -7, and -8. J. Biol. Chem. 1997, 272, 25719–25723. [Google Scholar] [CrossRef] [PubMed]
- Huber, K.L.; Hardy, J.A. Mechanism of zinc-mediated inhibition of caspase-9. Protein Sci. 2012, 21, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Delgado, E.M.; Hardy, J.A. Zinc-mediated allosteric inhibition of caspase-6. J. Biol. Chem. 2012, 287, 36000–36011. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Jacob, C.; Vallee, B.L.; Fischer, E.H. Inhibitory sites in enzymes: Zinc removal and reactivation by thionein. Proc. Natl. Acad. Sci. USA 1999, 96, 1936–1940. [Google Scholar] [CrossRef] [PubMed]
- Muroi, M.; Tanamoto, K.-I. Zinc- and oxidative property-dependent degradation of pro-caspase-1 and NLRP3 by ziram in mouse macrophages. Toxicol. Lett. 2015, 235, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.P.-Y.; Willingham, S.B.; Bergstralh, D.T. NLRs at the intersection of cell death and immunity. Nat. Rev. Immunol. 2008, 8, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-J.; Bao, S.; Galvez-Peralta, M.; Pyle, C.J.; Rudawsky, A.C.; Pavlovicz, R.E.; Killilea, D.W.; Li, C.; Nebert, D.W.; Wewers, M.D.; et al. ZIP8 regulates host defense through zinc-mediated inhibition of NF-kappaB. Cell Rep. 2013, 3, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Singh, S.; Siddiqi, N.J. Biomedical Implications of Heavy Metals Induced Imbalances in Redox Systems. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar] [PubMed]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014, 28, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, P.N.; Gore, M.G.; Jordan, P.M. Investigation of the effect of metal ions on the reactivity of thiol groups in human 5-aminolaevulinate dehydratase. Biochem. J. 1985, 225, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.R. The antioxidant properties of zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [PubMed]
- Prasad, A.S.; Beck, F.W.J.; Bao, B.; Fitzgerald, J.T.; Snell, D.C.; Steinberg, J.D.; Cardozo, L.J. Zinc supplementation decreases incidence of infections in the elderly: Effect of zinc on generation of cytokines and oxidative stress. Am. J. Clin. Nutr. 2007, 85, 837–844. [Google Scholar] [PubMed]
- Kloubert, V.; Rink, L. Zinc as a micronutrient and its preventive role of oxidative damage in cells. Food Funct. 2015, 6, 3195–3204. [Google Scholar] [CrossRef] [PubMed]
- Young, B.; Ott, L.; Kasarskis, E.; Rapp, R.; Moles, K.; Dempsey, R.J.; Tibbs, P.A.; Kryscio, R.; McClain, C. Zinc supplementation is associated with improved neurologic recovery rate and visceral protein levels of patients with severe closed head injury. J. Neurotrauma 1996, 13, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Besecker, B.Y.; Exline, M.C.; Hollyfield, J.; Phillips, G.; DiSilvestro, R.A.; Wewers, M.D.; Knoell, D.L. A comparison of zinc metabolism, inflammation, and disease severity in critically ill infected and noninfected adults early after intensive care unit admission. Am. J. Clin. Nutr. 2011, 93, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Costarelli, L.; Muti, E.; Malavolta, M.; Cipriano, C.; Giacconi, R.; Tesei, S.; Piacenza, F.; Pierpaoli, S.; Gasparini, N.; Faloia, E.; et al. Distinctive modulation of inflammatory and metabolic parameters in relation to zinc nutritional status in adult overweight/obese subjects. J. Nutr. Biochem. 2010, 21, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-L.; Hung, T.-C.; Hsieh, B.-S.; Chen, Y.-H.; Chen, T.-F.; Cheng, H.-L. Zinc at pharmacologic concentrations affects cytokine expression and induces apoptosis of human peripheral blood mononuclear cells. Nutrition 2006, 22, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Tsou, T.-C.; Chao, H.-R.; Yeh, S.-C.; Tsai, F.-Y.; Lin, H.-J. Zinc induces chemokine and inflammatory cytokine release from human promonocytes. J. Hazard. Mater. 2011, 196, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Beck, F.W.J.; Li, Y.; Bao, B.; Prasad, A.S.; Sarkar, F.H. Evidence for reprogramming global gene expression during zinc deficiency in the HUT-78 cell line. Nutrition 2006, 22, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Wessels, I.; Haase, H.; Engelhardt, G.; Rink, L.; Uciechowski, P. Zinc deficiency induces production of the proinflammatory cytokines IL-1beta and TNFalpha in promyeloid cells via epigenetic and redox-dependent mechanisms. J. Nutr. Biochem. 2013, 24, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Beck, F.W.; Grabowski, S.M.; Kaplan, J.; Mathog, R.H. Zinc deficiency: Changes in cytokine production and T-cell subpopulations in patients with head and neck cancer and in noncancer subjects. Proc. Assoc. Am. Physicians 1997, 109, 68–77. [Google Scholar] [PubMed]
- Prasad, A.S.; Bao, B.; Beck, F.W.J.; Sarkar, F.H. Correction of interleukin-2 gene expression by in vitro zinc addition to mononuclear cells from zinc-deficient human subjects: A specific test for zinc deficiency in humans. Transl. Res. J. Lab. Clin. Med. 2006, 148, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Pinna, K.; Kelley, D.S.; Taylor, P.C.; King, J.C. Immune functions are maintained in healthy men with low zinc intake. J. Nutr. 2002, 132, 2033–2036. [Google Scholar] [PubMed]
- Kahmann, L.; Uciechowski, P.; Warmuth, S.; Plumakers, B.; Gressner, A.M.; Malavolta, M.; Mocchegiani, E.; Rink, L. Zinc supplementation in the elderly reduces spontaneous inflammatory cytokine release and restores T cell functions. Rejuvenation Res. 2008, 11, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.P.; Rinaldi, N.A.; Ho, E. Zinc deficiency enhanced inflammatory response by increasing immune cell activation and inducing IL6 promoter demethylation. Mol. Nutr. Food Res. 2015, 59, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: Mechanisms of Host Defense. J. Nutr. 2007, 137, 1345–1349. [Google Scholar] [PubMed]
- Beck, F.W.; Prasad, A.S.; Kaplan, J.; Fitzgerald, J.T.; Brewer, G.J. Changes in cytokine production and T cell subpopulations in experimentally induced zinc-deficient humans. Am. J. Physiol. 1997, 272, E1002–E1007. [Google Scholar] [PubMed]
- Tapazoglou, E.; Prasad, A.S.; Hill, G.; Brewer, G.J.; Kaplan, J. Decreased natural killer cell activity in patients with zinc deficiency with sickle cell disease. J. Lab. Clin. Med. 1985, 105, 19–22. [Google Scholar] [PubMed]
- Honscheid, A.; Rink, L.; Haase, H. T-lymphocytes: A target for stimulatory and inhibitory effects of zinc ions. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Wellinghausen, N.; Martin, M.; Rink, L. Zinc inhibits interleukin-1-dependent T cell stimulation. Eur. J. Immunol. 1997, 27, 2529–2535. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, E.; Hilgers, R.-D.; Uciechowski, P.; Petersen, A.; Plumakers, B.; Rink, L. Zinc enhances the number of regulatory T cells in allergen-stimulated cells from atopic subjects. Eur. J. Nutr. 2017, 56, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, E.; Maywald, M.; Hilgers, R.-D.; Brieger, A.; Clarner, T.; Kipp, M.; Plümäkers, B.; Meyer, S.; Schwerdtle, T.; Rink, L. Induction of regulatory T cells in Th1-/Th17-driven experimental autoimmune encephalomyelitis by zinc administration. J. Nutr. Biochem. 2016, 29, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Maywald, M.; Meurer, S.K.; Weiskirchen, R.; Rink, L. Zinc supplementation augments TGF-β1-dependent regulatory T cell induction. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, E.; Metz, C.H.; Maywald, M.; Hilgers, R.; Weßels, I.; Senff, T.; Haase, H.; Jäger, M.; Ott, M.; Aspinall, R.; et al. Zinc supplementation induces regulatory T cells by inhibition of Sirt-1 deacetylase in mixed lymphocyte cultures. Mol. Nutr. Food Res. 2016, 60. [Google Scholar] [CrossRef] [PubMed]
- Hennigar, S.R.; McClung, J.P. Nutritional Immunity. Am. J. Lifestyle Med. 2016, 10, 170–173. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Chang, S.-M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.-S.; Karabiyik, A.; Cousins, R.J. Zinc transporter ZIP14 functions in hepatic zinc, iron and glucose homeostasis during the innate immune response (endotoxemia). PLoS ONE 2012, 7, e48679. [Google Scholar] [CrossRef]
- Glaser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schroder, J.-M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Botella, H.; Stadthagen, G.; Lugo-Villarino, G.; Chastellier, C.; de Neyrolles, O. Metallobiology of host-pathogen interactions: An intoxicating new insight. Trends Microbiol. 2012, 20, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Lappann, M.; Danhof, S.; Guenther, F.; Olivares-Florez, S.; Mordhorst, I.L.; Vogel, U. In vitro resistance mechanisms of Neisseria meningitidis against neutrophil extracellular traps. Mol. Microbiol. 2013, 89, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Stork, M.; Grijpstra, J.; Bos, M.P.; Manas Torres, C.; Devos, N.; Poolman, J.T.; Chazin, W.J.; Tommassen, J. Zinc piracy as a mechanism of Neisseria meningitidis for evasion of nutritional immunity. PLoS Pathog. 2013, 9, e1003733. [Google Scholar] [CrossRef] [PubMed]
- Bobrov, A.G.; Kirillina, O.; Fetherston, J.D.; Miller, M.C.; Burlison, J.A.; Perry, R.D. The Yersinia pestis siderophore, yersiniabactin, and the ZnuABC system both contribute to zinc acquisition and the development of lethal septicaemic plague in mice. Mol. Microbiol. 2014, 93, 759–775. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Jellbauer, S.; Poe, A.J.; Ton, V.; Pesciaroli, M.; Kehl-Fie, T.E.; Restrepo, N.A.; Hosking, M.P.; Edwards, R.A.; Battistoni, A.; et al. Zinc sequestration by the neutrophil protein calprotectin enhances Salmonella growth in the inflamed gut. Cell Host Microbe 2012, 11, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.E.; Harmon, K.; Caldwell, C.C.; Wong, H.R. Prophylactic zinc supplementation reduces bacterial load and improves survival in a murine model of sepsis. Pediatr. Crit. Care Med. 2012, 13, e323–e329. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Das, R.R. Zinc for the common cold. Cochrane Database Syst. Rev. 2013, CD001364. [Google Scholar] [CrossRef]
- Mezzetti, A.; Pierdomenico, S.D.; Costantini, F.; Romano, F.; de Cesare, D.; Cuccurullo, F.; Imbastaro, T.; Riario-Sforza, G.; Di Giacomo, F.; Zuliani, G.; et al. Copper/zinc ratio and systemic oxidant load: Effect of aging and aging-related degenerative diseases. Free Radic. Biol. Med. 1998, 25, 676–681. [Google Scholar] [CrossRef]
- Kozlowski, H.; Luczkowski, M.; Remelli, M.; Valensin, D. Copper, zinc and iron in neurodegenerative diseases (Alzheimer’s, Parkinson’s and prion diseases). Coord. Chem. Rev. 2012, 256, 2129–2141. [Google Scholar] [CrossRef]
- Malavolta, M.; Giacconi, R.; Piacenza, F.; Santarelli, L.; Cipriano, C.; Costarelli, L.; Tesei, S.; Pierpaoli, S.; Basso, A.; Galeazzi, R.; et al. Plasma copper/zinc ratio: An inflammatory/nutritional biomarker as predictor of all-cause mortality in elderly population. Biogerontology 2010, 11, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Overbeck, S.; Rink, L.; Haase, H. Modulating the immune response by oral zinc supplementation: A single approach for multiple diseases. Arch. Immunol. Ther. Exp. 2008, 56, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Hulisz, D. Efficacy of zinc against common cold viruses: An overview. J. Am. Pharm. Assoc. 2004, 44, 594–603. [Google Scholar] [CrossRef]
- Kurugol, Z.; Akilli, M.; Bayram, N.; Koturoglu, G. The prophylactic and therapeutic effectiveness of zinc sulphate on common cold in children. Acta Paediatr. 2006, 95, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.K.; Lai, S.; Sales, S.; Page, J.B.; Campa, A. Randomized Controlled Clinical Trial of Zinc Supplementation to Prevent Immunological Failure in HIV-Positive Adults1,2. Clin. Infect. Dis. 2010, 50, 1653–1660. [Google Scholar] [CrossRef] [PubMed]
- Asdamongkol, N.; Phanachet, P.; Sungkanuparph, S. Low Plasma Zinc Levels and Immunological Responses to Zinc Supplementation in HIV-Infected Patients with Immunological Discordance after Antiretroviral Therapy. Jpn. J. Infect. Dis. 2013, 66, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Lodha, R.; Shah, N.; Mohari, N.; Mukherjee, A.; Vajpayee, M.; Singh, R.; Singla, M.; Saini, S.; Bhatnagar, S.; Kabra, S.K. Immunologic effect of zinc supplementation in HIV-infected children receiving highly active antiretroviral therapy: A randomized, double-blind, placebo-controlled trial. J. Acquir. Immune Defic. Syndr. 2014, 66, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Veccia, S.; Ancarani, F.; Scalise, G.; Fabris, N. Benefit of oral zinc supplementation as an adjunct to zidovudine (AZT) therapy against opportunistic infections in AIDS. Int. J. Immunopharmacol. 1995, 17, 719–727. [Google Scholar] [CrossRef]
- Zazzo, J.F.; Rouveix, B.; Rajagopalon, P.; Levacher, M.; Girard, P.M. Effect of zinc on the immune status of zinc-depleted AIDS related complex patients. Clin. Nutr. 1989, 8, 259–261. [Google Scholar] [CrossRef]
- Bobat, R.; Coovadia, H.; Stephen, C.; Naidoo, K.L.; McKerrow, N.; Black, R.E.; Moss, W.J. Safety and efficacy of zinc supplementation for children with HIV-1 infection in South Africa: A randomised double-blind placebo-controlled trial. Lancet 2005, 366, 1862–1867. [Google Scholar] [CrossRef]
- Green, J.A.; Lewin, S.R.; Wightman, F.; Lee, M.; Ravindran, T.S.; Paton, N.I. A randomised controlled trial of oral zinc on the immune response to tuberculosis in HIV-infected patients. Int. J. Tuberc. Lung Dis. 2005, 9, 1378–1384. [Google Scholar] [PubMed]
- Fawzi, W.W.; Villamor, E.; Msamanga, G.I.; Antelman, G.; Aboud, S.; Urassa, W.; Hunter, D. Trial of zinc supplements in relation to pregnancy outcomes, hematologic indicators, and T cell counts among HIV-1-infected women in Tanzania. Am. J. Clin. Nutr. 2005, 81, 161–167. [Google Scholar] [PubMed]
- Villamor, E.; Aboud, S.; Koulinska, I.N.; Kupka, R.; Urassa, W.; Chaplin, B.; Msamanga, G.; Fawzi, W.W. Zinc supplementation to HIV-1-infected pregnant women: Effects on maternal anthropometry, viral load, and early mother-to-child transmission. Eur. J. Clin. Nutr. 2006, 60, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Deloria-Knoll, M.; Steinhoff, M.; Semba, R.D.; Nelson, K.; Vlahov, D.; Meinert, C.L. Effect of zinc and vitamin A supplementation on antibody responses to a pneumococcal conjugate vaccine in HIV-positive injection drug users: A randomized trial. Vaccine 2006, 24, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Nagao, Y.; Abe, K.; Imazeki, F.; Honda, K.; Yamasaki, K.; Miyanishi, K.; Taniguchi, E.; Kakuma, T.; Kato, J.; et al. Effects of branched-chain amino acids and zinc-enriched nutrients on prognosticators in HCV-infected patients: A multicenter randomized controlled trial. Mol. Med. Rep. 2015, 11, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Koyabu, T.; Kawashima, A.; Kakibuchi, N.; Kawakami, T.; Takaguchi, K.; Kita, K.; Okita, M. Zinc supplementation prevents the increase of transaminase in chronic hepatitis C patients during combination therapy with pegylated interferon alpha-2b and ribavirin. J. Nutr. Sci. Vitaminol. 2007, 53, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Nagamine, T.; Abe, T.; Takayama, H.; Sato, K.; Otsuka, T.; Kakizaki, S.; Hashimoto, Y.; Matsumoto, T.; Kojima, A.; et al. Zinc supplementation enhances the response to interferon therapy in patients with chronic hepatitis C. J. Viral Hepat. 2001, 8, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.-S.; Guo, C.-H.; Hsu, G.-S.W.; Chiou, Y.-L.; Yeh, M.-S.; Yaun, S.-R. The effect of zinc supplementation on the treatment of chronic hepatitis C patients with interferon and ribavirin. Clin. Biochem. 2005, 38, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Hoque, K.M.; Binder, H.J. Zinc in the treatment of acute diarrhea: Current status and assessment. Gastroenterology 2006, 130, 2201–2205. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Mitra, U.; Dutta, S.; Naik, T.N.; Rajendran, K.; Chatterjee, M.K. Zinc, vitamin A, and micronutrient supplementation in children with diarrhea: A randomized controlled clinical trial of combination therapy versus monotherapy. J. Pediatr. 2011, 159, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Taneja, D.K.; Devasenapathy, N.; Rajeshwari, K. Zinc supplementation for prevention of acute respiratory infections in infants: A randomized controlled trial. Indian Pediatr. 2014, 51, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Shah, U.H.; Abu-Shaheen, A.K.; Malik, M.A.; Alam, S.; Riaz, M.; AL-Tannir, M.A. The efficacy of zinc supplementation in young children with acute lower respiratory infections: A randomized double-blind controlled trial. Clin. Nutr. 2013, 32, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Estevez, N.S.; Alvarez-Guevara, A.N.; Rodriguez-Martinez, C.E. Effects of zinc supplementation in the prevention of respiratory tract infections and diarrheal disease in Colombian children: A 12-month randomised controlled trial. Allergol. Immunopathol. 2016, 44, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Sazawal, S.; Black, R.E.; Jalla, S.; Mazumdar, S.; Sinha, A.; Bhan, M.K. Zinc supplementation reduces the incidence of acute lower respiratory infections in infants and preschool children: A double-blind, controlled trial. Pediatrics 1998, 102, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Mahalanabis, D.; Lahiri, M.; Paul, D.; Gupta, S.; Gupta, A.; Wahed, M.A.; Khaled, M.A. Randomized, double-blind, placebo-controlled clinical trial of the efficacy of treatment with zinc or vitamin A in infants and young children with severe acute lower respiratory infection. Am. J. Clin. Nutr. 2004, 79, 430–436. [Google Scholar] [PubMed]
- Karyadi, E.; West, C.E.; Schultink, W.; Nelwan, R.H.H.; Gross, R.; Amin, Z.; Dolmans, W.M.V.; Schlebusch, H.; van der Meer, J.W.M. A double-blind, placebo-controlled study of vitamin A and zinc supplementation in persons with tuberculosis in Indonesia: Effects on clinical response and nutritional status. Am. J. Clin. Nutr. 2002, 75, 720–727. [Google Scholar] [PubMed]
- Mathur, N.K.; Bumb, R.A.; Mangal, H.N. Oral zinc in recurrent Erythema Nodosum Leprosum reaction. Lepr. India 1983, 55, 547–552. [Google Scholar] [PubMed]
- Mathur, N.K.; Bumb, R.A.; Mangal, H.N.; Sharma, M.L. Oral zinc as an adjunct to dapsone in lepromatous leprosy. Int. J. Lepr. Other Mycobact. Dis. Off. Org. Int. Lepr. Assoc. 1984, 52, 331–338. [Google Scholar]
- El-Shafei, M.M.; Kamal, A.A.; Soliman, H.; el-Shayeb, F.; Abdel Baqui, M.S.; Faragalla, S.; Sabry, M.K. Effect of oral zinc supplementation on the cell mediated immunity in lepromatous leprosy. J. Egypt. Public Health Assoc. 1988, 63, 311–336. [Google Scholar] [PubMed]
- Mahajan, P.M.; Jadhav, V.H.; Patki, A.H.; Jogaikar, D.G.; Mehta, J.M. Oral zinc therapy in recurrent erythema nodosum leprosum: A clinical study. Indian J. Lepr. 1994, 66, 51–57. [Google Scholar] [PubMed]
- Alam, A.N.; Sarker, S.A.; Wahed, M.A.; Khatun, M.; Rahaman, M.M. Enteric protein loss and intestinal permeability changes in children during acute shigellosis and after recovery: Effect of zinc supplementation. Gut 1994, 35, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Raqib, R.; Roy, S.K.; Rahman, M.J.; Azim, T.; Ameer, S.S.; Chisti, J.; Andersson, J. Effect of zinc supplementation on immune and inflammatory responses in pediatric patients with shigellosis. Am. J. Clin. Nutr. 2004, 79, 444–450. [Google Scholar] [PubMed]
- Rahman, M.J.; Sarker, P.; Roy, S.K.; Ahmad, S.M.; Chisti, J.; Azim, T.; Mathan, M.; Sack, D.; Andersson, J.; Raqib, R. Effects of zinc supplementation as adjunct therapy on the systemic immune responses in shigellosis. Am. J. Clin. Nutr. 2005, 81, 495–502. [Google Scholar] [PubMed]
- Roy, S.K.; Raqib, R.; Khatun, W.; Azim, T.; Chowdhury, R.; Fuchs, G.J.; Sack, D.A. Zinc supplementation in the management of shigellosis in malnourished children in Bangladesh. Eur. J. Clin. Nutr. 2008, 62, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Kashimura, H.; Suzuki, K.; Hassan, M.; Ikezawa, K.; Sawahata, T.; Watanabe, T.; Nakahara, A.; Mutoh, H.; Tanaka, N. Polaprezinc, a mucosal protective agent, in combination with lansoprazole, amoxycillin and clarithromycin increases the cure rate of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 1999, 13, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Zeba, A.N.; Sorgho, H.; Rouamba, N.; Zongo, I.; Rouamba, J.; Guiguemdé, R.T.; Hamer, D.H.; Mokhtar, N.; Ouedraogo, J.-B. Major reduction of malaria morbidity with combined vitamin A and zinc supplementation in young children in Burkina Faso: A randomized double blind trial. Nutr. J. 2008, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Shankar, A.H.; Genton, B.; Baisor, M.; Paino, J.; Tamja, S.; Adiguma, T.; Wu, L.; Rare, L.; Bannon, D.; Tielsch, J.M.; et al. The influence of zinc supplementation on morbidity due to Plasmodium falciparum: A randomized trial in preschool children in Papua New Guinea. Am. J. Trop. Med. Hyg. 2000, 62, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.J.; Evans, P.H.; Dardenne, M.; Prentice, A.; Lunn, P.G.; Northrop-Clewes, C.A.; Hoare, S.; Cole, T.J.; Horan, S.J.; Longman, S.C. A trial of zinc supplementation in young rural Gambian children. Br. J. Nutr. 1993, 69, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Muller, O.; Becher, H.; van Zweeden, A.B.; Ye, Y.; Diallo, D.A.; Konate, A.T.; Gbangou, A.; Kouyate, B.; Garenne, M. Effect of zinc supplementation on malaria and other causes of morbidity in west African children: Randomised double blind placebo controlled trial. BMJ Clin. Res. 2001, 322, 1567. [Google Scholar] [CrossRef]
- Zinc Against Plasmodium Study Group. Effect of zinc on the treatment of Plasmodium falciparum malaria in children: A randomized controlled trial. Am. J. Clin. Nutr. 2002, 76, 805–812. [Google Scholar]
- Richard, S.A.; Zavaleta, N.; Caulfield, L.E.; Black, R.E.; Witzig, R.S.; Shankar, A.H. Zinc and iron supplementation and malaria, diarrhea, and respiratory infections in children in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2006, 75, 126–132. [Google Scholar] [PubMed]
- Lazzerini, M.; Wanzira, H. Oral zinc for treating diarrhoea in children. Cochrane Database Syst. Rev. 2016. [CrossRef]
- World Health Organisation. Zinc Supplementation in the Management of Diarrhoea; World Health Organisation: Geneva, Switzerland, 2017. [Google Scholar]
- Finamore, A.; Massimi, M.; Conti Devirgiliis, L.; Mengheri, E. Zinc deficiency induces membrane barrier damage and increases neutrophil transmigration in Caco-2 cells. J. Nutr. 2008, 138, 1664–1670. [Google Scholar] [PubMed]
- Bao, S.; Knoell, D.L. Zinc modulates cytokine-induced lung epithelial cell barrier permeability. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L1132-41. [Google Scholar] [CrossRef] [PubMed]
- Lansdown, A.B.G.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Agren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Gosain, A.; DiPietro, L.A. Aging and Wound Healing. World J. Surg. 2004, 28, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wang, M.; Qi, J.; Wang, H.; Li, X.; Gupta, D.; Dziarski, R. Peptidoglycan Recognition Proteins Are a New Class of Human Bactericidal Proteins. J. Biol. Chem. 2006, 281, 5895–5907. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, L.-H.; Wang, S.; Li, X.; Lu, X.; Gupta, D.; Dziarski, R. Human Peptidoglycan Recognition Proteins Require Zinc to Kill Both Gram-Positive and Gram-Negative Bacteria and Are Synergistic with Antibacterial Peptides. J. Immunol. 2007, 178, 3116–3125. [Google Scholar] [CrossRef] [PubMed]
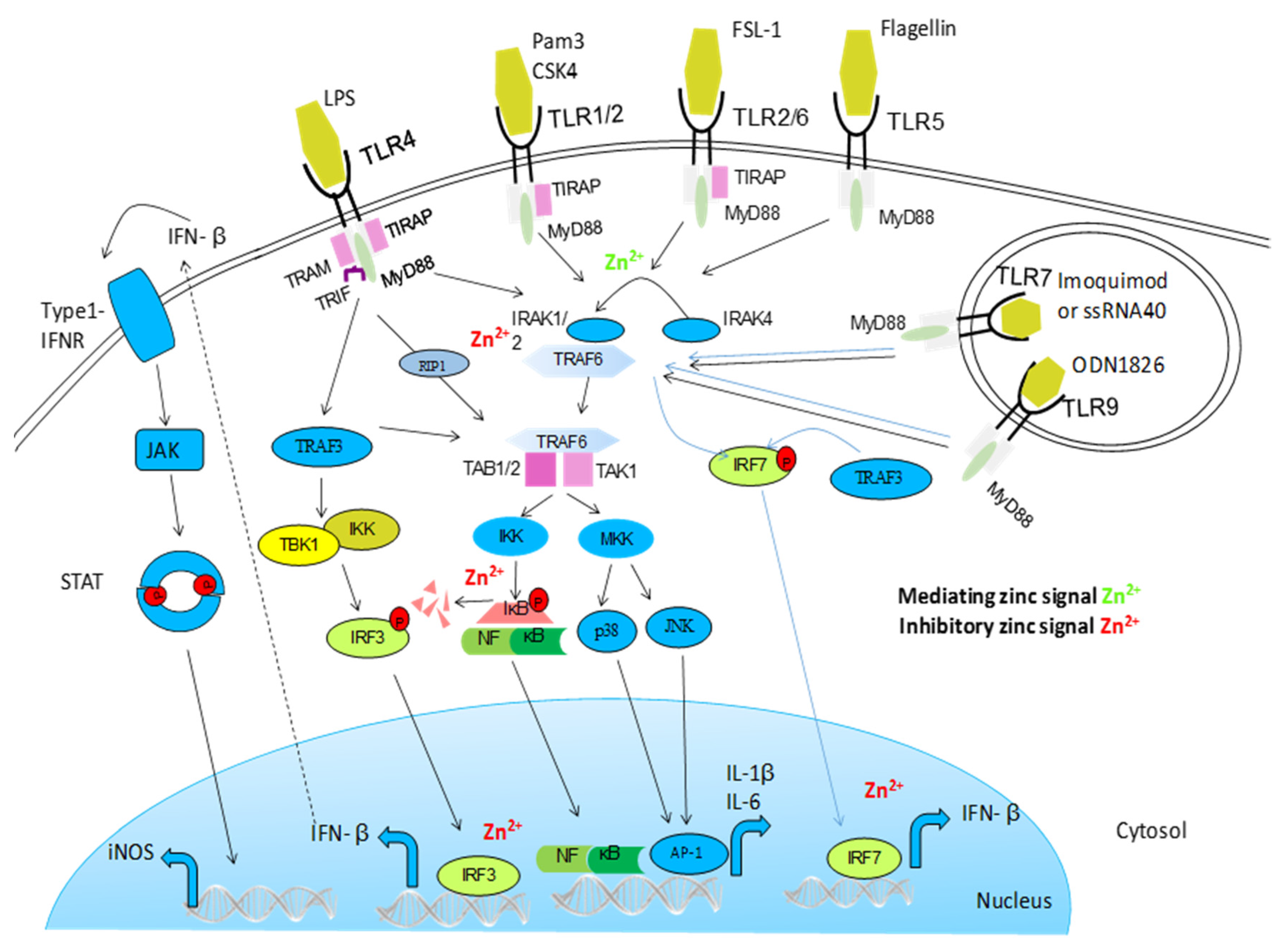
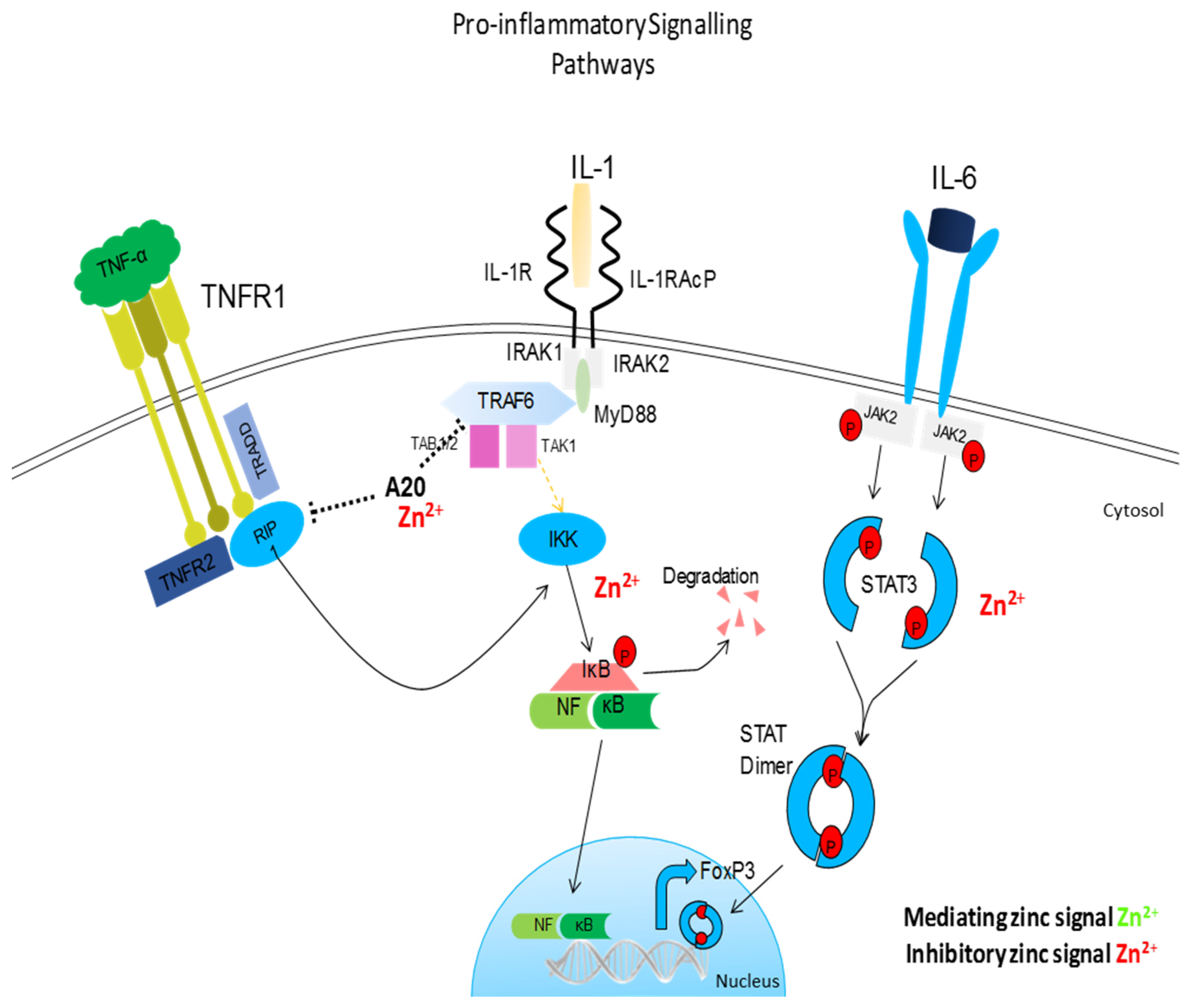
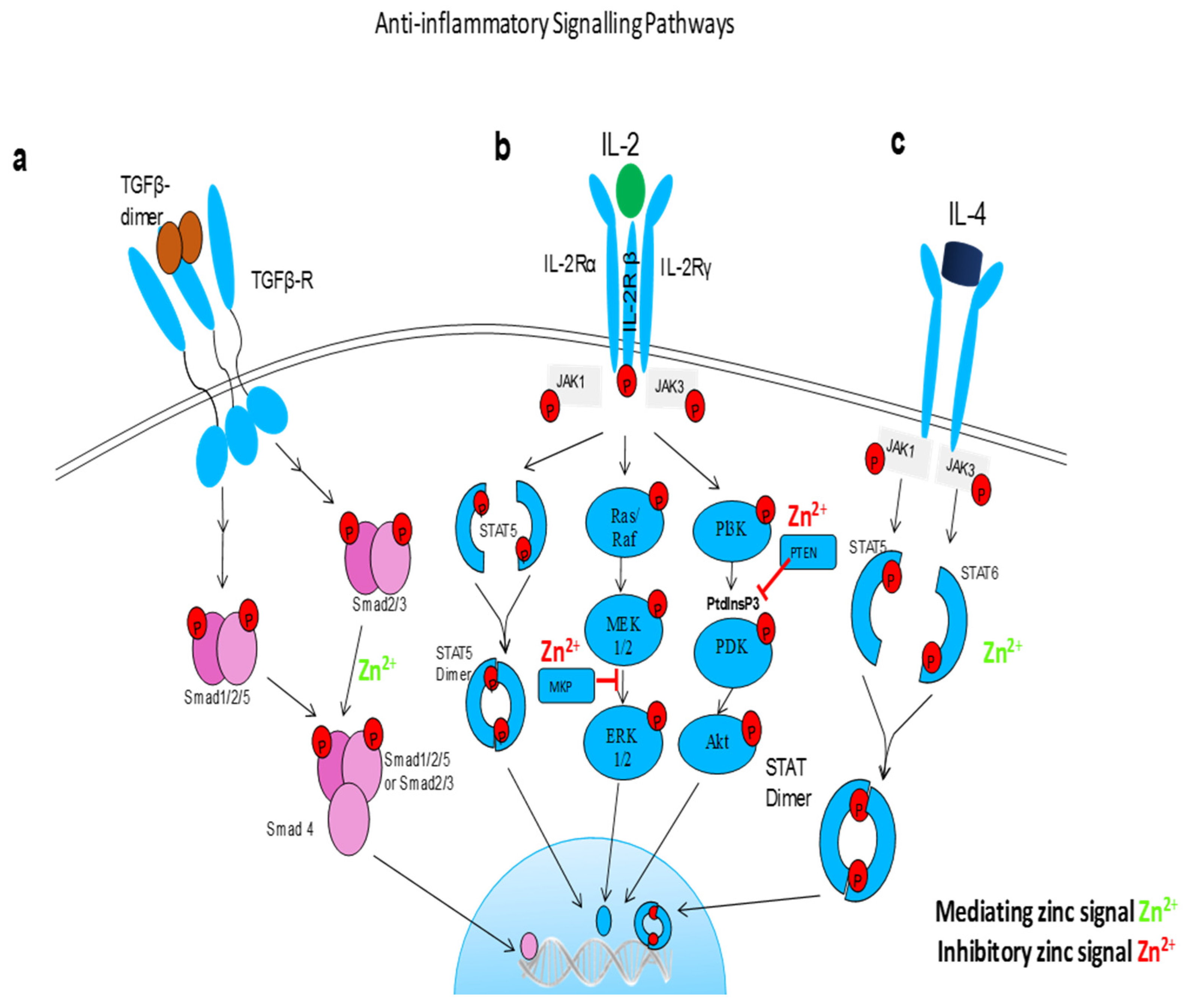

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Zinc Species | Zinc Dosage | Period | Participation | Effect of Zinc Supplementation | Reference |
|---|---|---|---|---|---|---|
| Common cold | more than 12 different studies, analyzing the therapeutic effects of zinc | variable results, reduced duration of symptoms if administered within 24 h of onset | [107] | |||
| zinc sulfate | 15 mg daily | 7 months | 100 (Z) 100 (P) | lower mean number of colds demonstrating the prophylactic effect of zinc | [108] | |
| HIV/AIDS | Not specified | 12 mg for women and 15 mg for men/day | 18 months | 115 (Z) 116 (P) | no effect on viral load. four-fold reduction in the likelihood of immunological failure. Reduced the rate of diarrhea by more than half. No significant difference in mortality | [109] |
| Chelated zinc | 15 mg daily | 12 months | Low: 5 (Z)/7 (P) Normal: 8 (Z)/10 (P) | CD4+ cell count significantly increased | [110] | |
| Zinc sulfate | 20 mg daily | 24 weeks | 26 (Z) 26 (P) | no effect on the increase in CD4%, decrease in viral load, anthropometric indices, and morbidity profile in HIV-infected children started on ART | [111] | |
| Zinc sulfate | 45.5 mg daily | 1 month | 29 (Z) 28 (P) | increase or stabilization in body weight; increase in plasma zinc levels, CD4+ T cells and plasma active zinc-bound thymulin; reduced or delayed frequency of opportunistic infections due to Pneumocystis jiroveci and Candida, not to Cytomegalovirus and Toxoplasma | [112] | |
| Zinc gluconate | 45 mg three time daily | 15 days | 5 (Z) 5 (C) | increased zinc concentrations in red blood, HLA-DR + cells, stimulation of lymphocyte transformation and phagocytosis of opsonized zymosan by neutrophils | [113] | |
| Zinc sulfate | 10 mg daily | 6 months | 44 (Z) 41 (P) | no effect on HIV viral load; decreased morbidity from diarrhea | [114] | |
| Zinc sulfate | 50 mg daily | 1 month | 31 (Z) 34 (P) | no improvements in immune responses to tuberculosis, CD4/CD8 ratio, lymphocyte subsets, and viral load | [115] | |
| Zinc sulfate | 25 mg daily | 6 months | 200 (Z) 200 (P) | when supplemented to pregnant HIV-positive women, no effect on birth outcomes or T-lymphocyte counts, and negative effects on hematological indicators | [116] | |
| Zinc sulfate | 25 mg daily | 6 months | 200 (Z) 200 (P) | increased risk of wasting | [117] | |
| 50 (Z) 50 (P) | no effect on viral load | |||||
| Zinc gluconate | 50 mg daily | 6 days | 44 (Z) 45 (P) | no improvements in antibody responses to a pneumococcal conjugate vaccine | [118] | |
| hepatitis C virus | Not specified | 10 mg | 60 days | 26 (Z + 6400 mg/day Branched-chain amino acids) 27 (P) | BCAA-to-tyrosine ratio (BTR) and zinc levels were significantly increased compared with the placebo group. supplementation reduced the serum α-fetoprotein AFP levels in patients who had elevated serum AFP levels at baseline | [119] |
| Polapre-zinc | 150 mg | 48 weeks | 11 (Z) 12 (C) | serum alanine aminotransferase (ALT) level is lower in zinc group compared to control group. HCV RNA disappeared in all patients in the zinc group and in 80% control patients at 48 week. Polaprezinc supplementation decreased plasma thiobarbituric acid reactive substances and prevented the decrease of polyunsaturated fatty acids of erythrocyte membrane phospholipids | [120] | |
| Polapre-zinc | 17 mg twice a day | 24 weeks | 40 (Z) 35 (C) | zinc supplementation increases serum zinc levels and improves the response to IFN-α therapy | [121] | |
| Zinc gluconate | 50 mg daily | 6 months | 18 (Z) 35 (P) 20 (C) | increased serum zinc levels; decreased incidences of gastrointestinal disturbances, body weight loss, and mild anemia | [122] | |
| Disease | Zinc Species | Zinc Dosage | Period | Participation | Effect of Zinc Supplementation | Reference |
|---|---|---|---|---|---|---|
| Diarrhea | multiple different studies | decreased duration, severity and occurrence of diarrhea | [123] | |||
| Not specified | 20 mg daily | 14 days | 41 (Z) 39 (micronutrient combination * + Vit A) 44 (Z+ Vit A) 43(P) | supplementation with a combination of micronutrients and vitamins was not superior to zinc alone, confirming clinical benefit of zinc in children with diarrhea | [124] | |
| Respiratory tract infections | Zinc sulfate | 20 mg daily | 5 months | 134 (Z) 124 (P) | reduced acute lower respiratory tract infection morbidity | [125] |
| zinc gluconate | 10 mg daily | 60 days | 48 (Z) 48 (P) | reduced episodes of acute lower respiratory infections and severe acute lower respiratory infections. Increased infection free days | [126] | |
| Zinc oxide | 5 mg daily | 12 months | 162 (Z) 167 (C) | decreased incidence of upper respiratory tract infections and diarrhoeal disease episodes | [127] | |
| zinc gluconate | 10 mg daily | 6 months | 298 (Z) 311 (P) | increased plasma zinc levels; decreased episodes of infection | [128] | |
| Zinc acetate | 10 mg twice a day | 5 days | 76 (Z) 74 (P) | increased serum zinc levels and recovery rates from illness and fever in boys | [129] | |
| Zinc sulfate | 15 mg daily | 6 months | 40 (Z) 40 (P) | increased plasma retinol concentrations; earlier sputum conversion and resolution of X-ray lesion area | [130] | |
| Tuberculosis | zinc sulfate | 220 mg daily | 18 months | 8 (Z) | reduced dose of clofazimine; withdrawal of steroids; toleration of dapsone; reduced incidence and severity of erythema nodosum leprosum; gradual decrease in the size of granuloma; gradual increase in the number of lymphocytes | [131] |
| Lepromatous leprosy | zinc sulfate | 220 mg daily | 18 months | 15 (Z) 10 (P) | decreased erythema, edema, and infiltration; regrowth of eyebrows; reduced bacterial index of granuloma; increased serum zinc levels, neovascularization, and endothelial cell proliferation | [132] |
| Zinc acetate | 200 mg twice a day | 13 weeks | 17 (Z) 10 (P) 10 (C) | increased serum zinc levels and delayed hypersensitivity reactions; decreased size of skin nodules; disappearance of erythema; regrowth of eyebrows | [133] | |
| zinc sulfate | 220 mg daily | 4 months | 40 (Z) | improvements on frequency, duration, and severity of erytheme nodosum leprosum reactions; reduction in steroid requirement | [134] | |
| Shigellosis | zinc acetate | 1.3 mg/kg three times a day | 1 month | 16 (Z) 16 (P) | increased intestinal mucosal permeability and better nitrogen absorption; increased serum zinc and alkaline phosphatase activity | [135] |
| zinc acetate | 20 mg daily | 2 weeks | 28 (Z) 28 (P) | increased serum zinc levels, lymphocyte proliferation in response to phytohemagglutinin and plasma invasion plasmid-encoded antigen-specific IgG titers | [136] | |
| zinc acetate | 20 mg daily | 2 weeks | 28 (Z) 28 (P) | increased serum zinc levels, serum shigellacidal antibody titers, CD20+ cells, and CD20+CD38+ cells | [137] | |
| Not specified | 20 mg daily | 2 weeks | 14 (Z) 16 (C) | faster recovery from acute illness. Increased mean body weight. Fewer episodes of diarrhoea | [138] | |
| Helicobacter pylori infection | polapre zinc | 150 mg twice a day | 7 days | 33 (Z) 28 (C) | administration of zinc together with antimicrobial therapy increased cure rate of Helicobacter pylori infection compared with antibiotic treatment alone | [139] |
| Disease | Zinc Species | Zinc Dosage | Period | Participation | Effect of Zinc Supplementation | Reference |
|---|---|---|---|---|---|---|
| Malaria | Not specified | 10 mg 6 times/week | 6 months | 74 (Z + 1 single dose of 200,000 IU Vit A) 74 (P) | significant decrease in the prevalence malaria. Lower malaria episodes. Time to first malaria episode was longer. 22% fewer fever episodes than the placebo group | [140] |
| Zinc gluconate | 10 mg 6 times/week | 46 weeks | 136 (Z) 138 (P) | reduction in Plasmodium falciparum-mediated febrile episodes | [141] | |
| Zinc acetate/zinc gluconate | 70 mg twice a week | 15 months | 55 (Z) 54 (P) | not statistically significant trend towards fewer malaria episodes; no effect on plasma and hair zinc, diarrhea, and respiratory illness | [142] | |
| Zinc sulfate | 12.5 mg 6 times/week | 6 months | 336 (Z) 344 (P) | increased serum zinc levels; reduced prevalence of diarrhea | [143] | |
| Zinc sulfate | 20 or 40 mg daily | 4 days | 473 (Z) 483 (P) | increased plasma zinc, no effect on fever, parasitemia, or hemoglobin concentration | [144] | |
| Zinc sulfate | 20 mg daily | 7 months | 191 (Z) 189 (P) | no significant effect on P. vivax incidence; significantly reduced diarrhea morbidity | [145] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. https://doi.org/10.3390/nu9060624
Gammoh NZ, Rink L. Zinc in Infection and Inflammation. Nutrients. 2017; 9(6):624. https://doi.org/10.3390/nu9060624
Chicago/Turabian StyleGammoh, Nour Zahi, and Lothar Rink. 2017. "Zinc in Infection and Inflammation" Nutrients 9, no. 6: 624. https://doi.org/10.3390/nu9060624
APA StyleGammoh, N. Z., & Rink, L. (2017). Zinc in Infection and Inflammation. Nutrients, 9(6), 624. https://doi.org/10.3390/nu9060624