Microbiota and Probiotics in Health and HIV Infection



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Intestinal Microbiota Functions
2.1. Function and Preservation of the Intestinal Barrier
2.2. Resistance to Pathogenic Colonization
2.3. Development and Stimulation of GALT
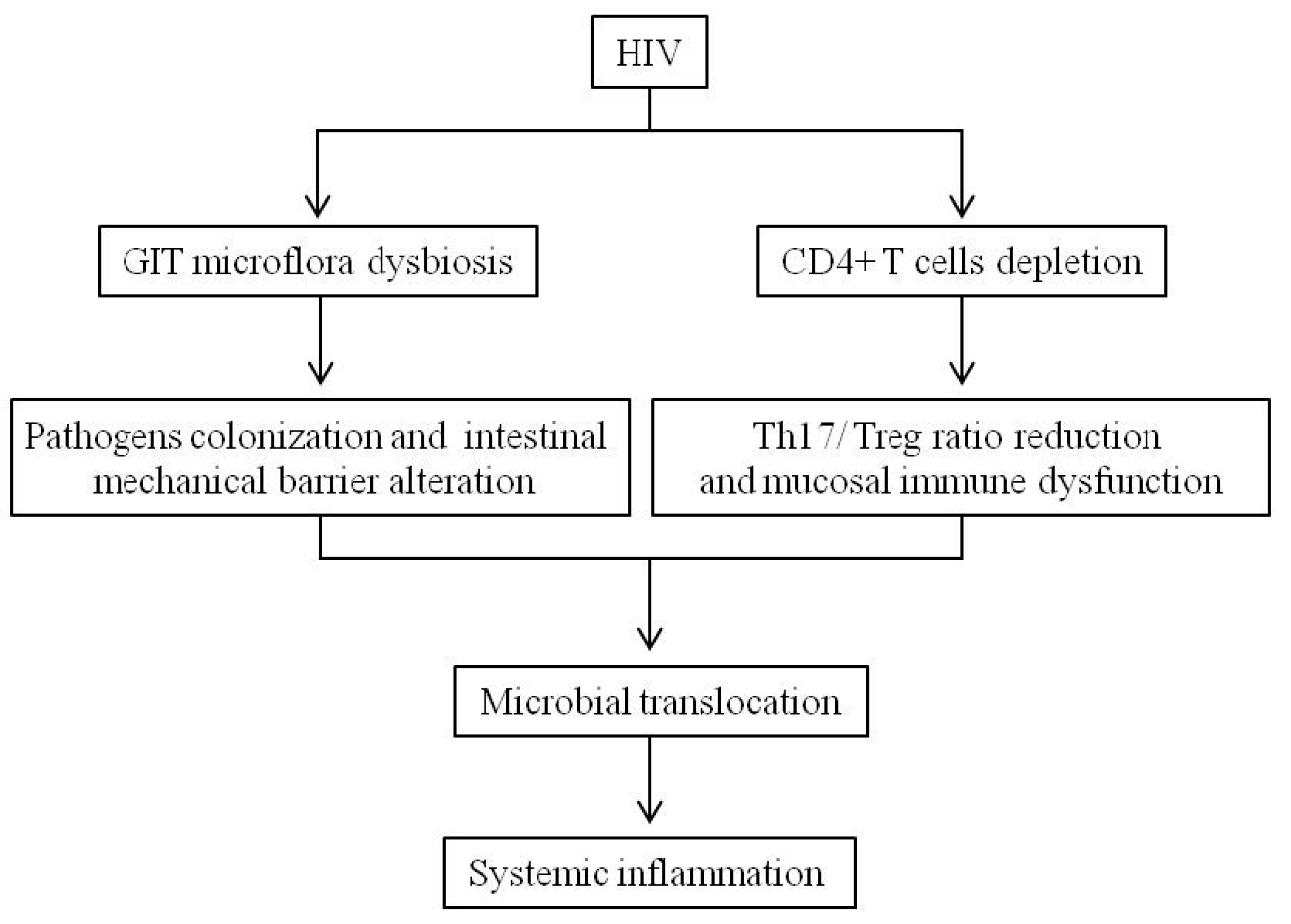
3. Bowel Conditions in People Living with HIV
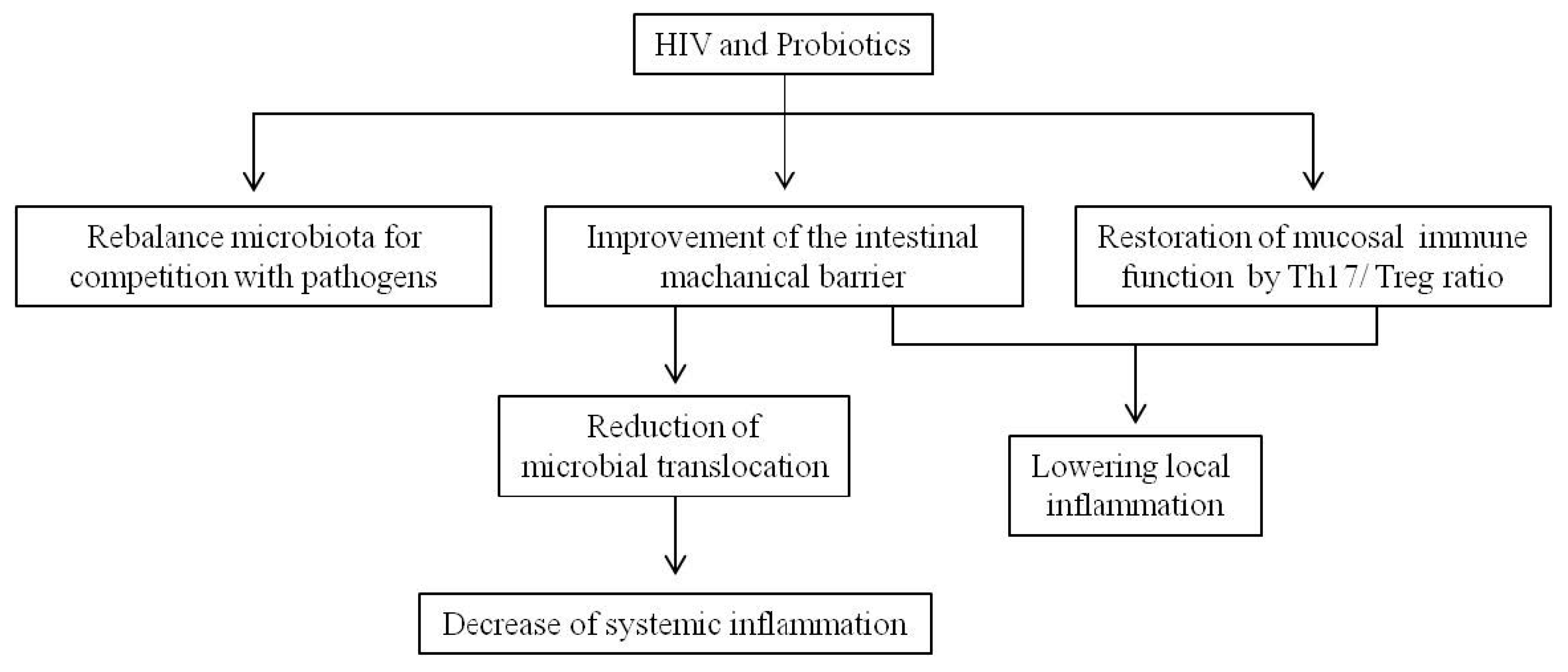
4. Probiotics as a New Therapeutic Approach That Might Improve Life in HIV-Positive Subjects
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parker, R.B. Probiotics, the other half of the antibiotics story. Anim. Nutr. Health 1974, 29, 4–8. [Google Scholar]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [PubMed]
- Salminen, S.; von Wright, A.; Morelli, L.; Marteau, P.; Brassart, D.; de Vos, W.M.; Fonden, R.; Saxelin, M.; Collins, K.; Mogensen, G.; et al. Demonstration of safety of probiotics—A review. Int. J. Food Microbiol. 1998, 44, 93–106. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotics and immune health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Im, S.H. Probiotics as an immune modulator. J. Nutr. Sci. Vitaminol. 2015, 61, 103–105. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, P.W.; Cooney, J.C. Probiotic bacteria influence the composition and function of the intestinal microbiota. Interdiscip. Perspect. Infect. Dis. 2008, 2008, 175285. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E. Impact of probiotics on colonizing microbiota of the gut. J. Clin. Gastroenterol. 2011, 45, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ganji-Arjenaki, M.; Rafieian-Kopaei, M. Probiotics are a good choice in remission of inflammatory bowel diseases: A meta-analysis and systematic review. J. Cell Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Kim, S.S. Probiotics and prebiotics: Present status and future perspectives on metabolic disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Guandalini, S. Probiotics for prevention and treatment of diarrhea. J. Clin. Gastroenterol. 2011, 45, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Sheil, B.; Shanahan, F.; O’Mahony, L. Probiotic effects on inflammatory bowel disease. J. Nutr. 2007, 137, 819–824. [Google Scholar]
- Yang, G.; Liu, Z.; Yang, P.C. Treatment of allergic rhinitis with probiotics: An alternative approach. N. Am. J. Med. Sci. 2013, 5, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Ge, T.; Xiao, Y.; Liao, Y.; Cui, Y.; Zhang, Y.; Ho, W.; Yu, G.; Zhang, T. Probiotics for prevention and treatment of respiratory tract infections in children: A systematic review and meta-analysis of randomized controlled trials. Medicine 2016, 95, 4509. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation? Brain Behav. Immun. 2011, 25, 397. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.P.; Texeira, T.F.; Ferreira, A.B.; Peluzio Mdo, C.; Alfenas Rde, C. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia. Br. J. Nutr. 2012, 108, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Douek, D.C. HIV infection and the gastrointestinal immune system. Mucosal Immunol. 2008, 1, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Kotler, D.P. HIV infection and the gastrointestinal tract. AIDS 2005, 19, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Schacker, T.W.; Ruff, L.E.; Price, D.A.; Taylor, J.H.; Beilman, G.J.; Nguyen, P.L.; Khoruts, A.; Larson, M.; Haase, A.T.; et al. CD4+ T-cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J. Exp. Med. 2004, 200, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.M.; Frank, D.N.; Wilson, C.C. The gut microbiome and HIV-1 pathogenesis: A two-way street. AIDS 2016, 30, 2737–2751. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Williams, B.; Frank, D.; Dillon, S.M.; Wilson, C.C.; Landay, A.L. Inside Out: HIV, the gut microbiome, and the mucosal immune system. J. Immunol. 2017, 198, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Mudd, J.C.; Brenchley, J.M. Gut mucosal barrier dysfunction, microbial dysbiosis, and their role in HIV-1 disease progression. J. Infect. Dis. 2016, 214, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Hamza, O.J.; Matee, M.I.; Simon, E.N.; Kikwilu, E.; Moshi, M.J.; Mugusi, F.; Mikx, F.H.; Verweij, P.E.; van der Ven, A.J. Oral manifestations of HIV infection in children and adults receiving highly active anti-retroviral therapy [HAART] in Dar es Salaam, Tanzania. BMC Oral Health 2006, 6, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci. Transl. Med. 2013, 5, 193ra91. [Google Scholar] [CrossRef] [PubMed]
- Nowak, P.; Troseid, M.; Avershina, E.; Barqasho, B.; Neogi, U.; Holm, K.; Hov, J.R.; Noyan, K.; Vesterbacka, J.; Svärd, J.; et al. Gut microbiota diversity predicts immune status in HIV-1 infection. AIDS 2015, 29, 2409–2418. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.M.; Esmaeili, A.; Shah, H.; Indyk, D.; Johnson, M.; Andreae, M.; Sacks, H.S. Probiotics in Human Immunodeficiency Virus Infection: A systematic review and evidence synthesis of benefits and risks. Open Forum Infect. Dis. 2016, 3, ofw164. [Google Scholar] [CrossRef] [PubMed]
- Klatt, N.R.; Canary, L.A.; Canary, L.A.; Sun, X.; Vinton, C.L.; Funderburg, N.T.; Morcock, D.R.; Quiñones, M.; Deming, C.B.; Perkins, M.; et al. Probiotic/prebiotic supplementation of antiretrovirals improves gastrointestinal immunity in SIV-infected macaques. J. Clin. Investig. 2013, 123, 903–907. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Rizzardini, G.; Van’t Land, B.; Amor, K.B.; van Schaik, J.; Torti, C.; Quirino, T.; Tincati, C.; Bandera, A.; Knol, J.; et al. Specific prebiotics modulate gut microbiota and immune activation in HAART-naïve HIV-infected adults: Results of the “COPA” pilot randomized trial. Nature 2011, 4, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Ohland, C.L.; MacNaughton, W.K. Probiotic bacteria and intestinal epithelial barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef] [PubMed]
- Delcenserie, V.; Martel, D.; Lamoureux, M.; Amiot, J.; Boutin, Y.; Roy, D. Immunomodulatory effects of probiotics in the intestinal tract. Curr. Issues Mol. Biol. 2008, 10, 37–54. [Google Scholar] [PubMed]
- Rijkers, G.T.; Bengmark, S.; Enck, P.; Haller, D.; Herz, U.; Kalliomaki, M.; Kudo, S.; Lenoir-Wijnkoop, I.; Mercenier, A.; Myllyluoma, E.; et al. Guidance for substantiating the evidence for beneficial effects of probiotics: Current status and recommendations for future research. J. Nutr. 2010, 140, 671S–676S. [Google Scholar] [CrossRef] [PubMed]
- Cinque, B.; La Torre, C.; Lombardi, F.; Palumbo, P.; Evtoski, Z.; Santini, S.J.; Falone, S.; Cimini, A.; Amicarelli, F.; Cifone, M.G. Vsl#3 probiotic differently influence iec-6 intestinal epithelial cell status and function. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef]
- Allen, S.J.; Martinez, E.G.; Gregorio, G.V.; Dans, L.F. Probiotics for treating acute infectious diarrhoea. Cochrane Database Syst. Rev. 2010, 11, CD003048. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Buffie, C.G.; Pamer, E.G. Microbiota-mediated colonization resistance against intestinal pathogens. Nat. Rev. Immunol. 2013, 13, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Bäumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Nombela, C. The microbiome as a human organ. Clin. Microbiol. Infect. 2012, 18, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Nishio, J.; Honda, K. Immunoregulation by the gut microbiota. Cell. Mol. Life Sci. 2012, 69, 3635–3650. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S.; Gordon, J.I.; Glimcher, L.H. Homeostasis and inflammation in the intestine. Cell 2010, 140, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Zhou, S.; Lu, Y.; Liu, J.; Jin, X.; Wan, H.; Wang, F. Changes in the expression and distribution of claudins, increased epithelial apoptosis, and a mannan-binding lectin-associated immune response lead to barrier dysfunction in dextran sodium sulfate-induced rat colitis. Gut Liver 2015, 9, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Sakuragi, N.; Takakuwa, A.; Ayabe, T. Paneth cell α-defensins and enteric microbiota in health and disease. Biosci. Microbiota Food Health 2016, 35, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Lugering, N.; Rautenberg, K.; Lugering, A.; Schmidt, M.A.; Stoll, R.; Domschke, W. Role of M-cells in intestinal barrier function. Ann. N. Y. Acad. Sci. 2000, 915, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Zeuthen, L. H.; Fink, L. N.; Frokiaer, H. Epithelial cells prime the immune response to an array of gut- derived commensals towards a tolerogenic phenotype through distinct actions of thymic stromal lymphopoietin and transforming growth factor β. Immunology 2008, 123, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Baba, N.; Samson, S.; Bourdet-Sicard, R.; Rubio, M.; Sarfati, M. Commensal bacteria trigger a full dendritic cell maturation program that promotes the expansion of non-Tr1 suppressor T-cells. J. Leukoc. Biol. 2008, 84, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Rogier, R.; Koenders, M.I.; Abdollahi-Roodsaz, S. Toll-like receptor mediated modulation of T-cell response by commensal intestinal microbiota as a trigger for autoimmune. Arthritis 2015, 2015, 527696. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 11, 215. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenerology 2004, 127, 224–238. [Google Scholar] [CrossRef]
- Melmed, G.; Thomas, L.S.; Lee, N.; Tesfay, S.Y.; Lukasek, K.; Michelsen, K.S.; Hou, Y.; Hu, B.; Arditi, M.; Abreu, M.T. Human intestinal epithelial cells are broadly unresponsive to Toll-like receptor 2-dependent bacterial ligands: Implications for host-microbial interactions in the gut. J. Immunol. 2003, 170, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Ashida, H.; Ogawa, M.; Kim, M.; Mimuro, H.; Sasakawa, C. Bacteria and host interactions in the gut epithelial barrier. Nat. Chem. Biol. 2012, 8, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Caricilli, A.M.; Castoldi, A.; Câmara, N.O. Intestinal barrier: A gentlemen's agreement between microbiota and immunity. World J. Gastrointest. Pathophysiol. 2014, 5, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Ahrne, S.; Hagslatt, M.L. Effect of lactobacilli on paracellular permeability in the gut. Nutrients 2011, 3, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.M.; Vieira, A.T.; Vinolo, M.A.; Oliveira, F.A.; Curi, R.; Martins, F.S. The central role of the gut microbiota in chronic inflammatory diseases. J. Immunol. Res. 2014, 689492. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.K.; Samak, G. Protection and Restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [PubMed]
- Bron, P.A.; Kleerebezem, M.; Brummer, R.J.; Cani, P.D.; Mercenier, A.; MacDonald, T.T.; Garcia-Ródenas, C.L.; Wells, J.M. Can probiotics modulate human disease by impacting intestinal barrier function? Br. J. Nutr. 2017, 117, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Abedi, D.; Feizizadeh, S.; Akbari, S.; Jafarian-Dehkordi, A. In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on Escherichia coli. Res. Pharm. Sci. 2013, 8, 260–268. [Google Scholar] [PubMed]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.L.; Powrie, F. Dendritic cells in intestinal immune regulation. Nat. Rev. Immunol. 2008, 8, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Gresnigt, M.S.; Kullberg, B.J.; van der Meer, J.W.; Joosten, L.A.; Netea, M.G. Th17 responses and host defense against microorganisms: An overview. BMB Rep. 2009, 42, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Donkor, O.N.; Ravikumar, M.; Proudfoot, O.; Day, S.L.; Apostolopoulos, V.; Paukovics, G.; Vasiljevic, T.; Nutt, S.L.; Gill, H. Cytokine profile and induction of T helper type 17 and regulatory T-cells by human peripheral mononuclear cells after microbial exposure. Clin. Exp. Immunol. 2012, 167, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Symons, A.; Budelsky, A.L.; Towne, J.E. Are Th17 cells in the gut pathogenic or protective? Mucosal Immunol. 2012, 5, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Bettelli, E.; Korn, T.; Oukka, M.; Kuchroo, V.K. Induction and effector functions of T(H)17 cells. Nature 2008, 453, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.C.; Tan, X.Y.; Luxenberg, D.P.; Karim, R.; Dunussi-Joannopoulos, K.; Collins, M.; Fouser, L.A. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 2006, 203, 2271–2279. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T-cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Mathias, A.; Pais, B.; Favre, L.; Benyacoub, J.; Corthésy, B. Role of secretory IgA in the mucosal sensing of commensal bacteria. Gut Microbes 2014, 5, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Young, C.; Neu, J. Molecular modulation of intestinal epithelial barrier: Contribution of microbiota. J. Biomed. Biotechnol. 2010, 305879. [Google Scholar] [CrossRef] [PubMed]
- Tincati, C.; Douek, D.C.; Marchetti, G. Gut barrier structure, mucosal immunity and intestinal microbiota in the pathogenesis and treatment of HIV infection. AIDS Res. Ther. 2016, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Neild, P.J.; Nijran, K.S.; Yazaki, E.; Evans, D.F.; Wingate, D.L.; Jewkes, R.; Gazzard, B.G. Delayed gastric emptying in human immunodeficiency virus infection: Correlation with symptoms, autonomic function, and intestinal motility. Dig. Dis. Sci. 2000, 45, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.A.; Keshavarzian, A.; Losurdo, J.; Swanson, G.; Siewe, B.; Forsyth, C.; French, A.; Demarais, P.; Sun, Y.; Koenig, L.; et al. A compositional look at the human gastrointestinal microbiome and immune activation parameters in HIV infected subjects. PLoS Pathog. 2014, 10, e1003829. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Li, M.; Campbell, T.B.; Flores, S.C.; Linderman, D.; Gebert, M.J.; Knight, R.; Fontenot, A.P.; Palmer, B.E. Alterations in the gut microbiota associated with HIV-1 infection. Cell Host Microbe 2013, 14, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.L.; Dong, Z.; Hecht, D.K.; Gianella, S.; Siewe, B.; Smith, D.M.; Landay, A.L.; et al. An altered intestinal mucosal microbiome in HIV-1 infection is associated with mucosal and systemic immune activation and endotoxemia. Mucosal Immunol. 2014, 7, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Perez-Santiago, J.; Gianella, S.; Massanella, M.; Spina, C.A.; Karris, M.Y.; Var, S.R.; Patel, D.; Jordan, P.S.; Young, J.A.; Little, S.J.; et al. Gut Lactobacillales are associated with higher CD4 and less microbial translocation during HIV infection. AIDS 2013, 27, 1921–1931. [Google Scholar] [CrossRef] [PubMed]
- Nwosu, F.C.; Avershina, E.; Wilson, R.; Rudi, K. Gut microbiota in HIV Infection: Implication for disease progression and management. Gastroenterol. Res. Pract. 2014, 2014, 803185. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Assimakopoulos, S.F.; Dimitropoulou, D.; Marangos, M.; Gogos, C.A. Intestinal barrier dysfunction in HIV infection: Pathophysiology, clinical implications and potential therapies. Infection 2014, 42, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Lapenta, C.; Boirivant, M.; Marini, M.; Santini, S.M.; Logozzi, M.; Viora, M.; Belardelli, F.; Fais, S. Human intestinal lamina propria lymphocytes are naturally permissive to HIV-1 infection. Eur. J. Immunol. 1999, 29, 1202–1208. [Google Scholar] [CrossRef]
- Steele, A.K.; Lee, E.J.; Manuzak, J.A.; Dillon, S.M.; Beckham, J.D.; McCarter, M.D.; Santiago, M.L.; Wilson, C.C. Microbial exposure alters HIV-1-induced mucosal CD4+ T-cell death pathways ex vivo. Retrovirology 2014, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Galloway, N.L.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Munoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [PubMed]
- French, M.; Keane, N.; McKinnon, E.; Phung, S.; Price, P. Susceptibility to opportunistic infections in HIV-infected patients with increased CD4 T-cell counts on antiretroviral therapy may be predicted by markers of dysfunctional effector memory CD4 T-cells and B cells. HIV Med. 2007, 8, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Vajpayee, M.; Ali, S.A.; Mojumdar, K.; Chauhan, N.K.; Singh, R. HIV-1 diseases progression associated with loss of Th17 cells in subtype ‘C’ infection. Cytokine 2012, 60, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.Y.; Merchant, A.; Kovacs, C.M.; Loutfy, M.; Persad, D.; Ostrowski, M.A. Virus-specific interleukin-17-producing CD4+ T-cells are detectable in early human immunodeficiency virus type 1 infection. J. Virol. 2008, 82, 6767–6771. [Google Scholar] [CrossRef] [PubMed]
- Ancuta, P.; Monteiro, P.; Sekaly, R.P. Th17 lineage commitment and HIV-1 pathogenesis. Curr. Opin. HIV AIDS 2010, 5, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Lindemans, C.A.; Calafiore, M.; Mertelsmann, A.M.; O’Connor, M.H.; Dudakov, J.A.; Jenq, R.R.; Velardi, E.; Young, L.F.; Smith, O.M.; Lawrence, G.; et al. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature 2015, 528, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, S.; George, M.D.; Baumler, A.J. Th17 cells, HIV and the gut mucosal barrier. Curr. Opin. HIV AIDS 2010, 5, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Chege, D.; Sheth, P.M.; Kain, T.; Kim, C.J.; Kovacs, C.; Loutfy, M.; Halpenny, R.; Kandel, G.; Chun, T.W.; Ostrowski, M.; et al. Sigmoid Th17 populations, the HIV latent reservoir, and microbial translocation in men on long-term antiretroviral therapy. AIDS 2011, 25, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.W. Th17, gut, and HIV: Therapeutic implications. Curr. Opin. HIV AIDS 2010, 5, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Hartigan-O’Connor, D.J.; Hirao, L.A.; McCune, J.M.; Dandekar, S. Th17 cells and regulatory T-cells in elite control over HIV and SIV. Curr. Opin. HIV AIDS 2011, 6, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, B.; Favre, D.; McCune, J.M. Th17 and regulatory T-cells: Implications for AIDS pathogenesis. Curr. Opin. HIV AIDS 2010, 5, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Brandt, L.; Benfield, T.; Mens, H.; Clausen, L.N.; Katzenstein, T.L.; Fomsgaard, A.; Karlsson, I. Low level of regulatory T-cells and maintenance of balance between regulatory T-cells and TH17 cells in HIV-1-infected elite controllers. J. Acquir. Immune Defic. Syndr. 2011, 57, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, M.F.; Weiss, L. The split personality of regulatory T-cells in HIV infection. Blood 2013, 121, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Jacobs, D.R., Jr.; Baker, J.V.; Calmy, A.; Duprez, D.; La Rosa, A.; Kuller, L.H.; Pett, S.L.; Ristola, M.; Ross, M.J.; et al. Markers of inflammation, coagulation, and renal function are elevated in adults with HIV infection. J. Infect. Dis. 2010, 201, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, S.; George, M.D.; Reay, E.; Guadalupe, M.; Flamm, J.; Prindiville, T.; Dandekar, S. Rapid onset of intestinal epithelial barrier dysfunction in primary human immunodeficiency virus infection is driven by an imbalance between immune response and mucosal repair and regeneration. J. Virol. 2008, 82, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.W.; Sinclair, E.; Rodriguez, B.; Shive, C.; Clagett, B.; Funderburg, N.; Robinson, J.; Huang, Y.; Epling, L.; Martin, J.N.; et al. Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J. Infect. Dis. 2014, 210, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Rajasuriar, R.; Wright, E.; Lewin, S.R. Impact of antiretroviral therapy (ART) timing on chronic immune activation/inflammation and end-organ damage. Curr. Opin. HIV AIDS 2015, 10, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Bolsewicz, K.; Debattista, J.; Vallely, A.; Whittaker, A.; Fitzgerald, L. Factors associated with antiretroviral treatment uptake and adherence: A review. Perspectives from Australia, Canada, and the United Kingdom. AIDS Care 2015, 27, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.D.; Romero, L.; Parnell, B.; Anderson, D.A.; Crowe, S.M.; Luchters, S. Feasibility of antiretroviral treatment monitoring in the era of decentralized HIV care: A systematic. AIDS Res. Ther. 2017, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Katlama, C.; Deeks, S.G.; Autran, B.; Martinez-Picado, J.; van Lunzen, J.; Rouzioux, C.; Miller, M.; Vella, S.; Schmitz, J.E.; Ahlers, J.; et al. Barriers to a cure for HIV: New ways to target and eradicate HIV-1 reservoirs. Lancet 2013, 381, 2109–2117. [Google Scholar] [CrossRef]
- Hummelen, R.; Vos, A.P.; van’t Land, B.; van Norren, K.; Reid, G. Altered host-microbe interaction in HIV: A target for intervention with pro- and pre-biotics. Int. Rev. Immunol. 2010, 29, 485–513. [Google Scholar] [CrossRef] [PubMed]
- D’Ettorre, G.; Ceccarelli, G.; Giustini, N.; Serafino, S.; Calantone, N.; De Girolamo, G.; Bianchi, L.; Bellelli, V.; Ascoli-Bartoli, T.; Marcellini, S.; et al. Probiotics reduce inflammation in antiretroviral treated, HIV-infected individuals: Results of the “Probio-HIV” clinical trial. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Surendran Nair, M.; Amalaradjou, M.A.; Venkitanarayanan, K. Antivirulence properties of probiotics in combating microbial pathogenesis. Adv. Appl. Microbiol. 2017, 98, 1–29. [Google Scholar] [PubMed]
- Falasca, K.; Vecchiet, J.; Ucciferri, C.; Di Nicola, M.; D’Angelo, C.; Reale, M. Effect of probiotic supplement on cytokine levels in HIV-infected individuals: A preliminary study. Nutrients 2015, 7, 8335–8347. [Google Scholar] [CrossRef] [PubMed]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, prebiotics and immunomodulation of gut mucosal defences: Homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S. The effect of probiotics and gut microbiota on Th17 cells. Int. Rev. Immunol. 2013, 32, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.K.; Lee, C.G.; So, J.S.; Chae, C.S.; Hwang, J.S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.C.; Im, S.H. Generation of regulatory dendritic cells and CD4+ Foxp3+ T-cells by probiotics administration suppresses immune disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 2159–2164. [Google Scholar] [CrossRef] [PubMed]
- López, P.; González-Rodríguez, I.; Gueimonde, M.; Margolles, A.; Suárez, A. Immune response to Bifidobacterium bifidum strains support Treg/Th17 plasticity. PLoS ONE 2011, 6, e24776. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Walmsley, S.L.; Raboud, J.M.; Kovacs, C.; Coburn, B.; Rousseau, R.; Reinhard, R.; Rosenes, R.; Kaul, R. Can probiotics reduce inflammation and enhance gut immune health in people living with HIV: Study designs for the Probiotic Visbiome for Inflammation and Translocation (PROOV IT) Pilot Trials. HIV Clin. Trials 2016, 17, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Cunningham-Rundles, S.; Ahrne, S.; Johann-Liang, R.; Abuav, R.; Dunn-Navarra, A.M.; Grassey, C.; Bengmark, S.; Cervia, J.S. Effect of probiotic bacteria on microbial host defense, growth, and immune function in human immunodeficiency virus type-1 infection. Nutrients 2011, 3, 1042–1070. [Google Scholar] [CrossRef] [PubMed]
- Yang, O.O.; Kelesidis, T.; Cordova, R.; Khanlou, H. Immunomodulation of antiretroviral drug-suppressed chronic HIV-1 infection in an oral probiotic double-blind placebo-controlled trial. AIDS Res. Hum. Retrovir. 2014, 30, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Irvine, S.L.; Hummelen, R.; Hekmat, S.; Looman, C.W.; Habbema, J.D.; Reid, G. Probiotic yogurt consumption is associated with an increase of CD4 count among people living with HIV/AIDS. J. Clin. Gastroenterol. 2010, 44, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, N.; Takahashi, N.; Xiao, J.Z. Suppressive effects of Bifidobacterium longum on the production of Th2-attracting chemokines induced with T-cell-antigen-presenting cell interactions. FEMS Immunol. Med. Microbiol. 2009, 55, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Mirmonsef, P.; Krass, L.; Landay, A.; Spear, G.T. The role of bacterial vaginosis and trichomonas in HIV transmission across the female genital tract. Curr. HIV Res. 2012, 10, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Hummelen, R.; Changalucha, J.; Butamanya, N.L.; Cook, A.; Habbema, J.D.; Reid, G. Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 to prevent or cure bacterial vaginosis among women with HIV. Int. J. Gynaecol. Obstet. 2010, 111, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Staab, B.; Eick, S.; Knöfler, G.; Jentsch, H. The influence of a probiotic milk drink on the development of gingivitis: A pilot study. J. Clin. Periodontol. 2009, 36, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Ryder, M.I.; Nittayananta, W.; Coogan, M.; Greenspan, D.; Greenspan, J.S. Periodontal disease in HIV/AIDS. Periodontology 2000 2012, 60, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G. Probiotics and periodontal health. J. Med. Life 2011, 4, 387–394. [Google Scholar] [PubMed]
- Noguera-Julian, M.; Guillén, Y.; Peterson, J.; Reznik, D.; Harris, E.V.; Joseph, S.J.; Rivera., J.; Kannanganat, S.; Amara, R.; Nguyen, M.L.; et al. Oral microbiome in HIV-associated periodontitis. Medicine 2017, 96, e5821. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.C.; Bedani, R.; Saad, S.M. Scientific evidence for health effects attributed to the consumption of probiotics and prebiotics: an update for current perspectives and future challenges. Br. J. Nutr. 2015, 114, 1993–2015. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, A.; West, C.E.; Prescott, S.L.; Jenmalm, M.C. Pre- and probiotics for allergy prevention: Time to revisit recommendations? Clin. Exp. Allergy 2016, 46, 1506–1521. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented foods products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Isolauri, E. Probiotics in human disease. Am. J. Clin. Nutr. 2001, 73, 1142S–1146S. [Google Scholar] [PubMed]
- Rajasuriar, R.; Khoury, G.; Kamarulzaman, A.; French, M.A.; Cameron, P.U.; Lewin, S.R. Persistent immune activation in chronic HIV infection: Do any interventions work? AIDS 2013, 27, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, L.A.; Jave-Suarez, L.F.; Fafutis-Morris, M.; Montes-Salcedo, K.E.; Valle-Gutierrez, L.G.; Campos-Loza, A.E.; Enciso-Gómez, L.F.; Andrade-Villanueva, J.F. Synbiotic therapy decreases microbial translocation and inflammation and improves immunological status in HIV-infected patients: A double-blind randomized controlled pilot trial. Nutr. J. 2012, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Amara, A.A.; Shibl, A. Role of Probiotics in health improvement, infection control and disease treatment and management. Saudi Pharm. J. 2015, 23, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Connolly, N.C.; Riddler, S.A.; Rinaldo, C.R. Proinflammatory cytokines in HIV disease—A review and rationale for new therapeutic approaches. AIDS Rev. 2005, 7, 168–180. [Google Scholar] [PubMed]
- Merlini, E.; Bai, F.; Bellistri, G.M.; Tincati, C.; d’Arminio Monforte, A.; Marchetti, G. Evidence for polymicrobic flora translocating in peripheral blood of HIV-infected patients with poor immune response to antiretroviral therapy. PLoS ONE 2011, 6, e18580. [Google Scholar] [CrossRef] [PubMed]
- Gad, M.; Ravn, P.; Søborg, D.A.; Lund-Jensen, K.; Ouwehand, A.C.; Jensen, S.S. Regulation of the IL-10/IL-12 axis in human dendritic cells with probiotic bacteria. FEMS Immunol. Med. Microbiol. 2011, 63, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Hempel, S.; Newberry, S.; Ruelaz, A.; Wang, Z.; Miles, J.; Suttorp, M.J.; Johnsen, B.; Shanman, R.; Slusser, W.; Fu, N.; et al. Safety of probiotics to reduce risk and prevent or treat disease. Evid. Rep. Technol. Assess 2011, 200, 1–645. [Google Scholar]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Angelo, C.; Reale, M.; Costantini, E. Microbiota and Probiotics in Health and HIV Infection. Nutrients 2017, 9, 615. https://doi.org/10.3390/nu9060615
D’Angelo C, Reale M, Costantini E. Microbiota and Probiotics in Health and HIV Infection. Nutrients. 2017; 9(6):615. https://doi.org/10.3390/nu9060615
Chicago/Turabian StyleD’Angelo, Chiara, Marcella Reale, and Erica Costantini. 2017. "Microbiota and Probiotics in Health and HIV Infection" Nutrients 9, no. 6: 615. https://doi.org/10.3390/nu9060615
APA StyleD’Angelo, C., Reale, M., & Costantini, E. (2017). Microbiota and Probiotics in Health and HIV Infection. Nutrients, 9(6), 615. https://doi.org/10.3390/nu9060615